Towards the Biological Control of Devastating Forest Pathogens from the Genus Armillaria

 ,
, 
 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation of Armillaria and Trichoderma
2.2. Identification of Armillaria and Trichoderma Isolates
2.3. Antagonistic Activity Assessment In Vitro by Dual Culture Assay
2.4. Extracellular Enzyme Activity Measurements
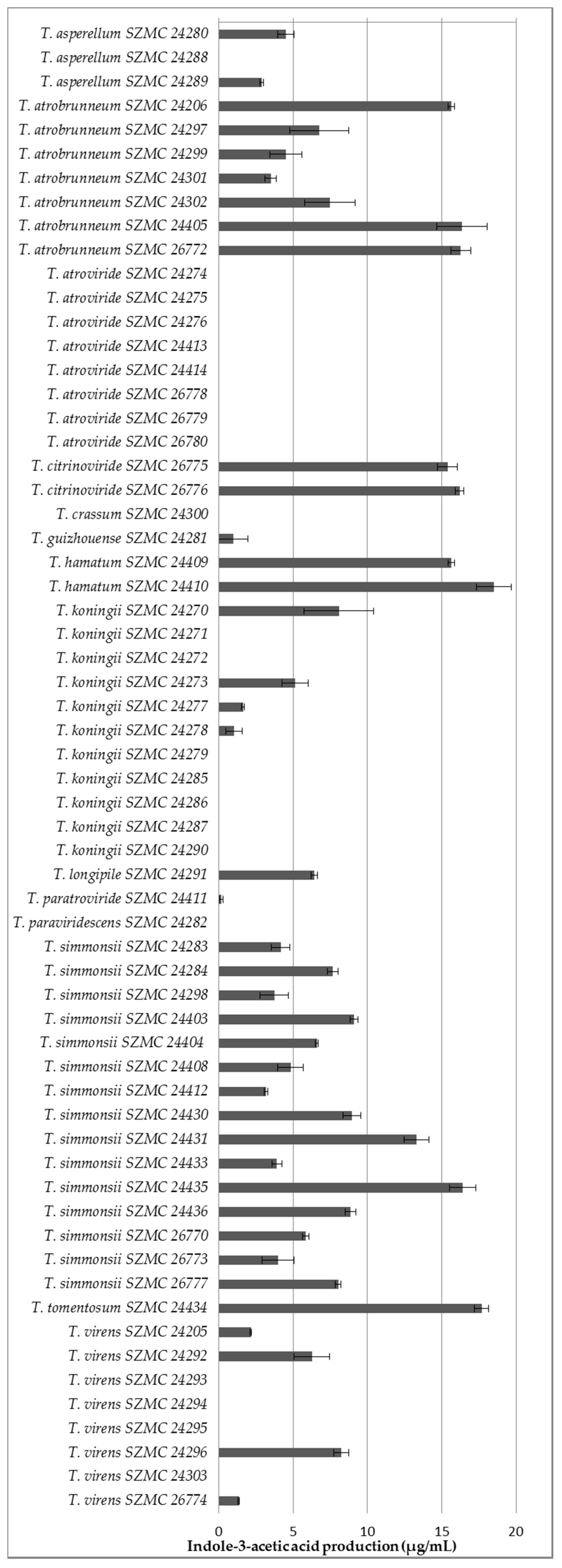
2.5. Quantitative Analysis of Indole-3-Acetic Acid Production
2.6. Siderophore Production
2.7. Field Study in the Keszthely Hills
3. Results
3.1. Diversity of the Genera Armillaria and Trichoderma in Healthy and Armillaria-Damaged Forests
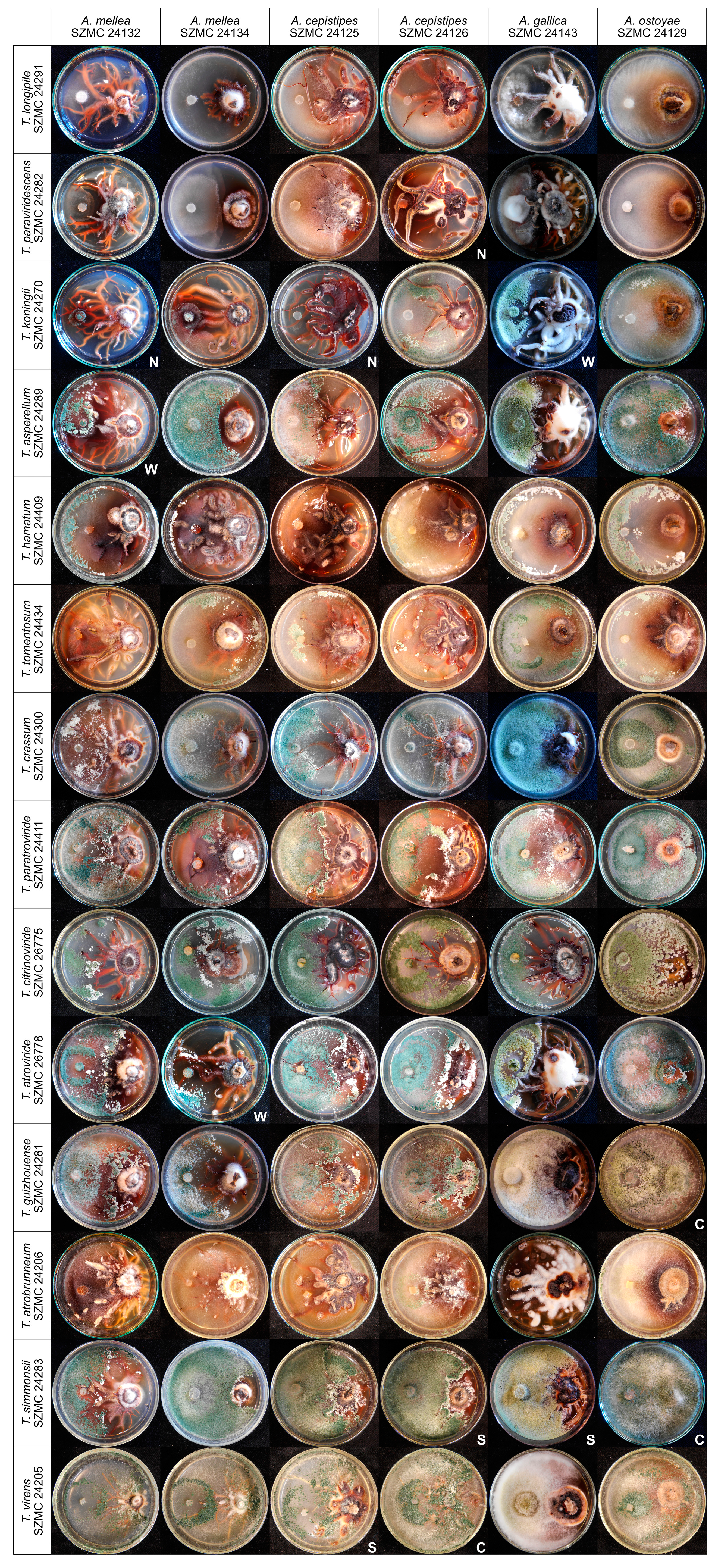
3.2. In Vitro Antagonism of the Isolated Trichoderma Strains Towards Armillaria Species
3.3. Extracellular Enzyme Production of the Trichoderma Isolates
3.4. Potential Plant Growth-Promoting Traits of the Isolated Trichoderma Strains
3.5. Field Experiment in a Heavily Armillaria-Damaged Forest in the Keszthely Hills
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Baumgartner, K.; Coetzee, M.P.; Hoffmeister, D. Secrets of the subterranean pathosystem of Armillaria. Mol. Plant Pathol. 2011, 12, 515–534. [Google Scholar] [CrossRef] [PubMed]
- Heinzelmann, R.; Dutech, C.; Tsykun, T.; Labbé, F.; Soularue, J.-P.; Prospero, S. Latest advances and future perspectives in Armillaria research. Can. J. Plant Pathol. 2019, 41, 1–23. [Google Scholar] [CrossRef]
- Sipos, G.; Anderson, J.B.; Nagy, L.G. Armillaria. Curr. Biol. 2018, 28, R297–R298. [Google Scholar] [CrossRef] [PubMed]
- Sipos, G.; Prasanna, A.N.; Walter, M.C.; O’Connor, E.; Bálint, B.; Krizsán, K.; Kiss, B.; Hess, J.; Varga, T.; Slot, J.; et al. Genome expansion and lineage-specific genetic innovations in the forest pathogenic fungi Armillaria. Nat. Ecol. Evol. 2017, 12, 1931–1941. [Google Scholar] [CrossRef] [PubMed]
- Ross-Davis, A.L.; Stewart, J.E.; Hanna, J.W.; Kim, M.S.; Knaus, B.J.; Cronn, R.; Rai, H.; Richardson, B.A.; Mcdonald, G.I.; Klopfenstein, N.B. Transcriptome of an Armillaria root disease pathogen reveals candidate genes involved in host substrate utilization at the host–pathogen interface. For. Pathol. 2013, 43, 468–477. [Google Scholar] [CrossRef]
- Morrison, D. Epidemiology of Armillaria root disease in Douglas-fir plantations in the cedar-hemlock zone of the southern interior of British Columbia. For. Pathol. 2009, 41, 31–40. [Google Scholar] [CrossRef]
- Kromroy, K.; Blanchette, R.A.; Grigal, D. Armillaria species on small woody plants, small woody debris, and root fragments in red pine stands. Can. J. For. Res. 2005, 35, 1487–1495. [Google Scholar] [CrossRef]
- Lee, C.; Dey, D.; Muzika, R.-M. Oak stump-sprout vigor and Armillaria infection after clearcutting in southeastern Missouri, USA. For. Ecol. Manag. 2016, 374, 211–219. [Google Scholar] [CrossRef]
- Cook, R.J. Making greater use of introduced microorganisms for biological control of plant pathogens. Annu. Rev. Phytopathol. 1993, 31, 53–80. [Google Scholar] [CrossRef]
- Nelson, E.E.; Pearce, M.H.; Malajczuk, N. Effects of Trichoderma spp. and ammonium sulphamate on establishment of Armillaria luteobubalina on stumps of Eucalyptus diversicolor. Mycol. Res. 1995, 99, 957–962. [Google Scholar] [CrossRef]
- Otieno, W.; Jeger, M.; Termorshuizen, A. Effect of infesting soil with Trichoderma harzianum and amendment with coffee pulp on survival of Armillaria. Biol. Control. 2003, 26, 293–301. [Google Scholar] [CrossRef]
- Otieno, W.; Termorshuizen, A.; Jeger, M.; Othieno, C. Efficacy of soil solarization, Trichoderma harzianum, and coffee pulp amendment against Armillaria sp. Crop. Prot. 2003, 22, 325–331. [Google Scholar] [CrossRef]
- Percival, G.; Smiley, E.; Fox, R. Root collar excavation with Trichoderma inoculations as a potential management strategy for honey fungus (Armillaria mellea). Arboric. J. 2011, 33, 267–280. [Google Scholar] [CrossRef]
- Olmedo Monfil, V.; Casas-Flores, S. Molecular mechanisms of biocontrol in Trichoderma spp. and their applications in agriculture. In Biotechnology and Biology of Trichoderma; Gupta, V.K., Schmoll, M., Herrera-Estrella, A., Upadhyay, R.S., Druzhinina, I., Tuohy, M.G., Eds.; Elsevier: Oxford, UK, 2014; pp. 429–453. [Google Scholar] [CrossRef]
- Sharma, V.; Salwan, R.; Sharma, P. Differential response of extracellular proteases of Trichoderma harzianum against fungal phytopathogens. Curr. Microbiol. 2016, 73, 419–425. [Google Scholar] [CrossRef] [PubMed]
- Contreras-Cornejo, H.A.; Macías-Rodríguez, L.; Alfaro-Cuevas, R.; López-Bucio, J. Trichoderma spp. improve growth of Arabidopsis seedlings under salt stress through enhanced root development, osmolite production, and Na+ elimination through root exudates. Mol. Plant Microbe Interact. 2014, 27, 503–514. [Google Scholar] [CrossRef] [PubMed]
- Shaw, C.G.; Roth, L.F. Persistence and distribution of a clone of Armillaria mellea in a ponderosa pine forest. Phytopathology 1976, 66, 1210–1213. [Google Scholar] [CrossRef]
- King, A.D.; Hocking, A.D.; Pitt, J.I. Dichloran-rose bengal medium for enumeration and isolation of molds from foods. Appl. Environ. Microbiol. 1979, 37, 959–964. [Google Scholar]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J.; Innis, M.; Gelfand, D.; Sninsky, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press Inc.: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Oskiera, M.; Szczech, M.; Bartoszewski, G. Molecular identification of Trichoderma strains collected to develop plant growth-promoting and biocontrol agents. J. Hortic. Res. 2015, 23, 75–86. [Google Scholar] [CrossRef]
- Druzhinina, I.; Kopchinskiy, A.; Komoń, M.; Szakacs, G.; Kubicek, C. An oligonucleotide barcode for species identification in Trichoderma and Hypocrea. Fungal Genet. Biol. 2005, 42, 813–828. [Google Scholar] [CrossRef]
- Kopchinskiy, A.; Komoń, M.; Kubicek, C.P.; Druzhinina, I.S. TrichoBLAST: A multilocus database for Trichoderma and Hypocrea identifications. Mycol. Res. 2005, 109, 658–660. [Google Scholar] [CrossRef]
- Szekeres, A.; Leitgeb, B.; Kredics, L.; Manczinger, L.; Vágvölgyi, C. A novel, image analysis-based method for the evaluation of in vitro antagonism. J. Microbiol. Meth. 2006, 65, 619–622. [Google Scholar] [CrossRef] [PubMed]
- Gordon, S.A.; Weber, R.P. Colorimetric estimation of indoleacetic acid. Plant Physiol. 1951, 26, 192–195. [Google Scholar] [CrossRef] [PubMed]
- Milagres, A.M.; Machuca, A.; Napoleao, D. Detection of siderophore production from several fungi and bacteria by a modification of chrome azurol S (CAS) agar plate assay. J. Microbiol. Meth. 1999, 37, 1–6. [Google Scholar] [CrossRef]
- Schwyn, B.; Neilands, J.B. Universal chemical assay for the detection and determination of siderophores. Anal. Biochem. 1987, 160, 47–56. [Google Scholar] [CrossRef]
- Jaklitsch, W. European species of Hypocrea part II: Species with hyaline ascospores. Fungal Divers. 2011, 48, 1–250. [Google Scholar] [CrossRef]
- Jaklitsch, W.M. European species of Hypocrea Part I. The green-spored species. Stud. Mycol. 2009, 63, 1–91. [Google Scholar] [CrossRef]
- Jaklitsch, W.M.; Voglmayr, H. Biodiversity of Trichoderma (Hypocreaceae) in Southern Europe and Macaronesia. Stud. Mycol. 2015, 80, 1–87. [Google Scholar] [CrossRef]
- Błaszczyk, L.; Strakowska, J.; Chełkowski, J.; Gąbka-Buszek, A.; Kaczmarek, J. Trichoderma species occurring on wood with decay symptoms in mountain forests in Central Europe: Genetic and enzymatic characterization. J. Appl. Genet. 2016, 57, 397–407. [Google Scholar] [CrossRef]
- Chaverri, P.; Branco-Rocha, F.; Jaklitsch, W.; Gazis, R.; Degenkolb, T.; Samuels, G.J. Systematics of the Trichoderma harzianum species complex and the re-identification of commercial biocontrol strains. Mycologia 2015, 107, 558–590. [Google Scholar] [CrossRef]
- Newitt, J.T.; Prudence, S.M.M. Biocontrol of cereal crop diseases using Streptomycetes. Pathogens 2019, 8, 78. [Google Scholar] [CrossRef]
- Woo, S.; Ruocco, M.; Vinale, F.; Nigro, M.; Marra, R.; Lombardi, N.; Pascale, A.; Lanzuise, S.; Manganiello, G.; Lorito, M. Trichoderma-based products and their widespread use in agriculture. Open Mycol. J. 2014, 8, 71–126. [Google Scholar] [CrossRef] [Green Version]
- Gajera, H.P.; Hirpara, D.G.; Katakpara, Z.A.; Patel, S.V.; Golakiya, B.A. Molecular evolution and phylogenetic analysis of biocontrol genes acquired from SCoT polymorphism of mycoparasitic Trichoderma koningii inhibiting phytopathogen Rhizoctonia solani Kuhn. Infect. Genet. Evol. 2016, 45, 383–392. [Google Scholar] [CrossRef] [PubMed]
- Chou, H.; Xiao, Y.T.; Tsai, J.N.; Li, T.T.; Wu, H.Y.; Liu, L.D.; Tzeng, D.S.; Chung, C.L. In vitro and in planta evaluation of Trichoderma asperellum TA as a biocontrol agent against Phellinus noxius, the cause of brown root rot disease of trees. Plant Dis. 2019, 103, 2733–2741. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, P.K.; Hurley, J.F.; Taylor, J.T.; Puckhaber, L.; Lehner, S.; Druzhinina, I.; Schumacher, R.; Kenerley, C.M. Ferricrocin, the intracellular siderophore of Trichoderma virens, is involved in growth, conidiation, gliotoxin biosynthesis and induction of systemic resistance in maize. Biochem. Biophys. Res. Commun. 2018, 505, 606–611. [Google Scholar] [CrossRef] [PubMed]
- Lehner, S.M.; Atanasova, L.; Neumann, N.K.; Krska, R.; Lemmens, M.; Druzhinina, I.S.; Schuhmacher, R. Isotope-assisted screening for iron-containing metabolites reveals a high degree of diversity among known and unknown siderophores produced by Trichoderma spp. Appl. Environ. Microbiol. 2013, 79, 18–31. [Google Scholar] [CrossRef] [Green Version]
- Segarra, G.; Casanova, E.; Avilés, M.; Trillas, I. Trichoderma asperellum strain T34 controls Fusarium wilt disease in tomato plants in soilless culture through competition for iron. Microb. Ecol. 2010, 59, 141–149. [Google Scholar] [CrossRef] [PubMed]
- Anke, H.; Kinn, J.; Bergquist, K.-E.; Sterner, O. Production of siderophores by strains of the genus Trichoderma. Biol. Met. 1991, 4, 176–180. [Google Scholar] [CrossRef]
- Wang, C.; Zhuang, W.-Y. Evaluating effective Trichoderma isolates for biocontrol of Rhizoctonia solani causing root rot of Vigna unguiculata. J. Integr. Agric. 2019, 18, 2072–2079. [Google Scholar] [CrossRef]
- Chagas, L.F.B.; De Castro, H.G.; Colonia, B.S.O.; De Carvalho Filho, M.R.; De Oliveira Miller, L.; Chagas, A.F.J. Efficiency of Trichoderma spp. as a growth promoter of cowpea (Vigna unguiculata) and analysis of phosphate solubilization and indole acetic acid synthesis. Braz. J. Bot. 2016, 39, 437–445. [Google Scholar] [CrossRef]
- Schnabel, G.; Rollins, A.P.; Henderson, G.W. Field evaluation of Trichoderma spp. for control of Armillaria root rot of peach. Plant Health Prog. 2011, 10, 3. [Google Scholar] [CrossRef] [Green Version]
- Downer, J.; Faber, B. Non-chemical control of Armillaria mellea infection of Prunus persica. J. Plant Sci. Phytopathol. 2019, 3, 050–055. [Google Scholar] [CrossRef] [Green Version]
- Reza Asef, M.; Goltapeh, E.M.; Danesh, Y.R. Antagonistic effects of Trichoderma species in biocontrol of Armillaria mellea in fruit trees in Iran. J. Plant Prot. Res. 2008, 48, 213–222. [Google Scholar] [CrossRef]
- Szczałba, M.; Kopta, T.; Gąstoł, M.; Sękara, A. Comprehensive insight into arbuscular mycorrhizal fungi, Trichoderma spp. and plant multilevel interactions with emphasis on biostimulation of horticultural crops. J. Appl. Microbiol. 2019, 127, 630–647. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Medina, A.; Roldán, A.; Pascual, J.A. Performance of a Trichoderma harzianum bentonite-vermiculite formulation against Fusarium wilt in seedling nursery melon plants. Hortic. Sci. 2009, 44, 2025–2027. [Google Scholar] [CrossRef] [Green Version]
- De Jaeger, N.; de la Providencia, I.; de Boulois, D.H.; Declerck, S. Trichoderma harzianum might impact phosphorus transport by arbuscular mycorrhizal fungi. FEMS Microbiol. Ecol. 2011, 77, 558–567. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Location | GPS-N | GPS-E | Collection Date | Sample | Isolate Identifier | Diagnosis | ITS (GenBank) | tef1α (GenBank) |
|---|---|---|---|---|---|---|---|---|
| Armillaria Isolates | ||||||||
| Rosalia, Austria | 47° 41.649 | 16° 17.940 | 28.10.2016 | fruiting body | SZMC 24125 | Armillaria cepistipes | - | MN580140 |
| 47° 41.640 | 16° 17.937 | 28.10.2016 | fruiting body | SZMC 24126 | Armillaria cepistipes | - | MN580151 | |
| 47° 41.628 | 16° 17.929 | 28.10.2016 | rhizomorph | SZMC 24128 | Armillaria ostoyae | - | MN580144 | |
| 47° 41.629 | 16° 17.964 | 28.10.2016 | fruiting body | SZMC 24129 | Armillaria ostoyae | - | MN580139 | |
| 47° 41.621 | 16° 17.948 | 28.10.2016 | fruiting body | SZMC 24130 | Armillaria ostoyae | - | MN580142 | |
| Keszthely, Hungary | 46° 48.728 | 17° 16.992 | 20.07.2016 | bulk soil | SZMC 24095 | Armillaria gallica | - | MN580162 |
| 46° 48.712 | 17° 16.994 | 20.07.2016 | bulk soil | SZMC 24098 | Armillaria gallica | - | MN580163 | |
| 46° 48.702 | 17° 16.987 | 29.10.2016 | bulk soil | SZMC 24099 | Armillaria gallica | - | MN580160 | |
| 46° 48.657 | 17° 16.954 | 29.10.2016 | fruiting body | SZMC 24131 | Armillaria mellea | MN585779 | MN580137 | |
| 46° 48.671 | 17° 16.959 | 29.10.2016 | fruiting body | SZMC 24132 | Armillaria mellea | MN585780 | MN580159 | |
| 46° 48.706 | 17° 16.949 | 29.10.2016 | fruiting body | SZMC 24133 | Armillaria mellea | MN585781 | MN580138 | |
| 46° 48.723 | 17° 16.974 | 29.10.2016 | fruiting body | SZMC 24134 | Armillaria mellea | MN585777 | MN580152 | |
| 46° 48.712 | 17° 16.978 | 29.10.2016 | fruiting body | SZMC 24135 | Armillaria mellea | - | MN580145 | |
| 46° 48.736 | 17° 16.992 | 29.10.2016 | fruiting body | SZMC 24651 | Armillaria mellea | - | MN580153 | |
| 46° 48.772 | 17° 16.992 | 29.10.2016 | fruiting body | SZMC 24136 | Armillaria mellea | MN585778 | MN580155 | |
| 46° 48.760 | 17° 16.982 | 29.10.2016 | fruiting body | SZMC 24137 | Armillaria mellea | MN585776 | MN580154 | |
| 46° 48.749 | 17° 16.936 | 29.10.2016 | fruiting body | SZMC 24139 | Armillaria mellea | MN585782 | MN580158 | |
| 46° 48.720 | 17° 17.009 | 29.10.2016 | fruiting body | SZMC 24140 | Armillaria mellea | MN585783 | MN580146 | |
| 46° 48.736 | 17° 16.992 | 03.11.2016 | fruiting body | SZMC 24141 | Armillaria mellea | - | MN580150 | |
| 46° 48.665 | 17° 16.993 | 03.11.2016 | fruiting body | SZMC 24142 | Armillaria mellea | - | MN580143 | |
| 46° 48.738 | 17° 16.956 | 03.11.2016 | fruiting body | SZMC 24143 | Armillaria gallica | - | MN580141 | |
| 46° 48.877 | 17° 17.143 | 03.11.2016 | fruiting body | SZMC 24144 | Armillaria mellea | - | MN580148 | |
| 46° 48.883 | 17° 17.153 | 03.11.2016 | fruiting body | SZMC 24145 | Armillaria mellea | - | MN580149 | |
| 46° 48.917 | 17° 17.062 | 03.11.2016 | fruiting body | SZMC 24146 | Armillaria mellea | - | MN580161 | |
| 46° 48.892 | 17° 16.941 | 03.11.2016 | fruiting body | SZMC 24147 | Armillaria mellea | - | MN580157 | |
| 46° 48.883 | 17° 16.937 | 03.11.2016 | fruiting body | SZMC 24148 | Armillaria mellea | - | MN580156 | |
| 46° 47.935 | 17° 16.958 | 03.11.2016 | fruiting body | SZMC 24149 | Armillaria mellea | - | MN580147 | |
| Trichoderma Isolates | ||||||||
| Rosalia, Austria | 47° 41.649 | 16° 17.940 | 28.10.2016 | bulk soil | SZMC 24270 | Trichoderma koningii | MN516459 | MN520036 |
| SZMC 24271 | Trichoderma koningii | MN516460 | MN520038 | |||||
| SZMC 24272 | Trichoderma koningii | MN516461 | MN520042 | |||||
| SZMC 24273 | Trichoderma koningii | MN516462 | MN520041 | |||||
| 47° 41.640 | 16° 17.937 | 28.10.2016 | bulk soil | SZMC 24274 | Trichoderma atroviride | MN516463 | MN520048 | |
| SZMC 24275 | Trichoderma atroviride | MN516464 | MN520049 | |||||
| SZMC 24276 | Trichoderma atroviride | MN516465 | MN520050 | |||||
| 47° 41.618 | 16° 17.873 | 28.10.2016 | bulk soil | SZMC 24277 | Trichoderma koningii | MN516466 | MN520033 | |
| SZMC 24278 | Trichoderma koningii | MN516467 | MN520040 | |||||
| SZMC 24279 | Trichoderma koningii | MN516468 | MN520039 | |||||
| SZMC 24280 | Trichoderma asperellum | MN516469 | MN520031 | |||||
| 47° 41.629 | 16° 17.964 | 28.10.2016 | bulk soil | SZMC 24285 | Trichoderma koningii | MN516474 | MN520034 | |
| SZMC 24286 | Trichoderma koningii | MN516475 | MN520037 | |||||
| SZMC 24287 | Trichoderma koningii | MN516476 | MN520035 | |||||
| SZMC 24288 | Trichoderma asperellum | MN516477 | MN520032 | |||||
| 47° 41.621 | 16° 17.948 | 28.10.2016 | bulk soil | SZMC 24289 | Trichoderma asperellum | MN516478 | MN520030 | |
| SZMC 24290 | Trichoderma koningii | MN516479 | MN520043 | |||||
| 47° 40.896 | 16° 17.211 | 28.10.2016 | bulk soil | SZMC 24291 | Trichoderma longipile | MN516480 | MN520056 | |
| 47° 41.628 | 16° 17.929 | 28.10.2016 | rhizomorph | SZMC 24281 | Trichoderma guizhouense | MN516470 | MN520084 | |
| SZMC 24282 | Trichoderma paraviridescens | MN516471 | MN520044 | |||||
| SZMC 24283 | Trichoderma simmonsii | MN516472 | MN520079 | |||||
| SZMC 24284 | Trichoderma simmonsii | MN516473 | MN520078 | |||||
| Keszthely, Hungary | 46° 48.657 | 17° 16.954 | 29.10.2016 | bulk soil | SZMC 24429 | Trichoderma simmonsii | - | MN520069 |
| SZMC 24430 | Trichoderma simmonsii | - | MN520071 | |||||
| SZMC 24431 | Trichoderma simmonsii | - | MN520077 | |||||
| SZMC 26770 | Trichoderma simmonsii | - | MN520072 | |||||
| SZMC 24433 | Trichoderma simmonsii | - | MN520081 | |||||
| 46° 48.671 | 17° 16.959 | 29.10.2016 | bulk soil | SZMC 24434 | Trichoderma tomentosum | - | MN520066 | |
| 46° 48.706 | 17° 16.949 | 29.10.2016 | bulk soil | SZMC 24435 | Trichoderma simmonsii | - | MN520075 | |
| SZMC 26771 | Trichoderma simmonsii | - | MN520080 | |||||
| SZMC 24436 | Trichoderma simmonsii | - | MN520068 | |||||
| SZMC 24403 | Trichoderma simmonsii | - | MN520076 | |||||
| SZMC 24404 | Trichoderma simmonsii | - | MN520082 | |||||
| 46° 48.723 | 17° 16.974 | 29.10.2016 | bulk soil | SZMC 26772 | Trichoderma atrobrunneum | - | MN520090 | |
| SZMC 24405 | Trichoderma atrobrunneum | - | MN520091 | |||||
| 46° 48.712 | 17° 16.978 | 29.10.2016 | bulk soil | SZMC 26773 | Trichoderma simmonsii | - | MN520083 | |
| SZMC 26774 | Trichoderma virens | - | MN520058 | |||||
| SZMC 24408 | Trichoderma simmonsii | - | MN520067 | |||||
| 46° 48.736 | 17° 16.992 | 29.10.2016 | bulk soil | SZMC 24409 | Trichoderma hamatum | - | MN520028 | |
| SZMC 24410 | Trichoderma hamatum | - | MN520029 | |||||
| 46° 48.772 | 17° 16.992 | 29.10.2016 | rhizosphere | SZMC 26778 | Trichoderma atroviride | MN516444 | MN520052 | |
| SZMC 26779 | Trichoderma atroviride | MN516445 | MN520051 | |||||
| SZMC 26780 | Trichoderma atroviride | MN516446 | MN520053 | |||||
| 46° 48.769 | 17° 16.961 | 29.10.2016 | bulk soil | SZMC 24411 | Trichoderma paratroviride | - | MN520045 | |
| SZMC 26775 | Trichoderma citrinoviride | - | MN520054 | |||||
| SZMC 26776 | Trichoderma citrinoviride | - | MN520055 | |||||
| SZMC 26777 | Trichoderma simmonsii | - | MN520070 | |||||
| SZMC 24412 | Trichoderma simmonsii | - | MN520074 | |||||
| 46° 48.749 | 17° 16.936 | 29.10.2016 | rhizosphere | SZMC 24413 | Trichoderma atroviride | MN520047 | ||
| SZMC 24414 | Trichoderma atroviride | MN520046 | ||||||
| 46° 48.738 | 17° 16.956 | 20.07.2016 | rhizomorph | SZMC 24292 | Trichoderma virens | MN516447 | MN520059 | |
| SZMC 24293 | Trichoderma virens | MN516448 | MN520061 | |||||
| SZMC 24294 | Trichoderma virens | MN516449 | MN520062 | |||||
| SZMC 24295 | Trichoderma virens | MN516450 | MN520060 | |||||
| SZMC 24296 | Trichoderma virens | MN516451 | MN520064 | |||||
| 46° 48.758 | 17° 16.959 | 20.07.2016 | rhizomorph | SZMC 24297 | Trichoderma atrobrunneum | MN516452 | MN520087 | |
| SZMC 24298 | Trichoderma simmonsii | MN516453 | MN520073 | |||||
| SZMC 24299 | Trichoderma atrobrunneum | MN516454 | MN520086 | |||||
| SZMC 24300 | Trichoderma crassum | MN516455 | MN520057 | |||||
| SZMC 24301 | Trichoderma atrobrunneum | MN516456 | MN520088 | |||||
| SZMC 24302 | Trichoderma atrobrunneum | MN516457 | MN520089 | |||||
| SZMC 24303 | Trichoderma virens | MN516458 | MN520063 | |||||
| 46° 48.722 | 17° 16.993 | 20.07.2016 | rhizomorph | SZMC 24205 | Trichoderma virens | - | MN520065 | |
| SZMC 24206 | Trichoderma atrobrunneum | MN516443 | MN520085 |
| Species | Isolate | Number of Tested Armillaria Isolates | BCI Values ≥ 80 | Percentage of BCI ≥ 80 (%) |
|---|---|---|---|---|
| T. tomentosum | SZMC 24434 | 25 | 15 | 60.00 |
| T. paratroviride | SZMC 24411 | 25 | 19 | 76.00 |
| T. crassum | SZMC 24300 | 25 | 13 | 52.00 |
| T. hamatum | SZMC 24409 | 25 | 9 | 36.00 |
| T. hamatum | SZMC 24410 | 25 | 16 | 64.00 |
| T. citrinoviride | SZMC 26775 | 25 | 12 | 48.00 |
| T. citrinoviride | SZMC 26776 | 25 | 13 | 52.00 |
| T. virens | SZMC 26774 | 25 | 24 | 96.00 |
| T. virens | SZMC 24205 | 25 | 17 | 68.00 |
| T. virens | SZMC 24292 | 25 | 22 | 88.00 |
| T. virens | SZMC 24293 | 25 | 22 | 88.00 |
| T. virens | SZMC 24294 | 25 | 23 | 92.00 |
| T. virens | SZMC 24295 | 25 | 22 | 88.00 |
| T. virens | SZMC 24296 | 25 | 22 | 88.00 |
| T. virens | SZMC 24303 | 25 | 24 | 96.00 |
| T. atrobrunneum | SZMC 26772 | 25 | 19 | 76.00 |
| T. atrobrunneum | SZMC 24405 | 25 | 15 | 60.00 |
| T. atrobrunneum | SZMC 24206 | 25 | 6 | 24.00 |
| T. atrobrunneum | SZMC 24297 | 25 | 10 | 40.00 |
| T. atrobrunneum | SZMC 24299 | 25 | 6 | 24.00 |
| T. atrobrunneum | SZMC 24301 | 25 | 11 | 44.00 |
| T. atrobrunneum | SZMC 24302 | 25 | 10 | 40.00 |
| T. simmonsii | SZMC 24430 | 25 | 15 | 60.00 |
| T. simmonsii | SZMC 24431 | 25 | 17 | 68.00 |
| T. simmonsii | SZMC 26770 | 25 | 21 | 84.00 |
| T. simmonsii | SZMC 24433 | 25 | 16 | 64.00 |
| T. simmonsii | SZMC 24435 | 25 | 19 | 76.00 |
| T. simmonsii | SZMC 24436 | 25 | 18 | 72.00 |
| T. simmonsii | SZMC 24403 | 25 | 17 | 68.00 |
| T. simmonsii | SZMC 24404 | 25 | 18 | 72.00 |
| T. simmonsii | SZMC 26773 | 25 | 19 | 76.00 |
| T. simmonsii | SZMC 24408 | 25 | 10 | 40.00 |
| T. simmonsii | SZMC 26777 | 25 | 17 | 68.00 |
| T. simmonsii | SZMC 24412 | 25 | 16 | 64.00 |
| T. simmonsii | SZMC 24298 | 25 | 13 | 52.00 |
| T. simmonsii | SZMC 24283 | 25 | 18 | 72.00 |
| T. simmonsii | SZMC 24284 | 25 | 16 | 64.00 |
| T. atroviride | SZMC 26778 | 25 | 12 | 48.00 |
| T. atroviride | SZMC 26779 | 25 | 8 | 32.00 |
| T. atroviride | SZMC 26780 | 25 | 10 | 40.00 |
| T. atroviride | SZMC 24413 | 25 | 16 | 64.00 |
| T. atroviride | SZMC 24414 | 25 | 14 | 56.00 |
| T. atroviride | SZMC 24274 | 25 | 14 | 56.00 |
| T. atroviride | SZMC 24275 | 25 | 12 | 48.00 |
| T. atroviride | SZMC 24276 | 25 | 10 | 40.00 |
| T. koningii | SZMC 24270 | 25 | 9 | 36.00 |
| T. koningii | SZMC 24271 | 25 | 9 | 36.00 |
| T. koningii | SZMC 24272 | 25 | 9 | 36.00 |
| T. koningii | SZMC 24273 | 25 | 7 | 28.00 |
| T. koningii | SZMC 24277 | 25 | 9 | 36.00 |
| T. koningii | SZMC 24278 | 25 | 7 | 28.00 |
| T. koningii | SZMC 24279 | 25 | 10 | 40.00 |
| T. koningii | SZMC 24285 | 25 | 7 | 28.00 |
| T. koningii | SZMC 24286 | 25 | 8 | 32.00 |
| T. koningii | SZMC 24287 | 25 | 7 | 28.00 |
| T. koningii | SZMC 24290 | 25 | 10 | 40.00 |
| T. asperellum | SZMC 24280 | 25 | 9 | 36.00 |
| T. asperellum | SZMC 24288 | 25 | 7 | 28.00 |
| T. asperellum | SZMC 24289 | 25 | 8 | 32.00 |
| T. guizhouense | SZMC 24281 | 25 | 16 | 64.00 |
| T. paraviridescens | SZMC 24282 | 25 | 6 | 24.00 |
| T. longipile | SZMC 24291 | 25 | 6 | 24.00 |
| Parcel | Total No. | 1 Dead No. | 2 Growing No. | 3 Survivor No. | 4 FM No. | 5 Corrected Total No. | Dead % | Growing % | Growing + Survivor % |
|---|---|---|---|---|---|---|---|---|---|
| 1 (treated) | 40 | 5 | 24 | 3 | 8 | 32 | 15.6 | 75.0 | 84.4 |
| 2 (treated) | 40 | 5 | 27 | 5 | 3 | 37 | 13.5 | 73.0 | 86.5 |
| 3 (treated) | 40 | 7 | 28 | 4 | 1 | 39 | 17.9 | 71.8 | 82.1 |
| Total | 120 | 17 | 79 | 12 | 12 | 108 | 15.7 | 73.1 | 84.3 |
| 4 (untreated) | 40 | 23 | 5 | 2 | 10 | 30 | 76.7 | 16.7 | 23.3 |
| 5 (untreated) | 39 | 13 | 19 | 2 | 5 | 34 | 38.2 | 55.9 | 61.8 |
| 6 (untreated) | 36 | 7 | 24 | 0 | 5 | 31 | 22.6 | 77.4 | 77.4 |
| Total | 115 | 43 | 48 | 4 | 20 | 95 | 45.3 | 50.5 | 54.7 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, L.; Bóka, B.; Kedves, O.; Nagy, V.D.; Szűcs, A.; Champramary, S.; Roszik, R.; Patocskai, Z.; Münsterkötter, M.; Huynh, T.; et al. Towards the Biological Control of Devastating Forest Pathogens from the Genus Armillaria. Forests 2019, 10, 1013. https://0-doi-org.brum.beds.ac.uk/10.3390/f10111013
Chen L, Bóka B, Kedves O, Nagy VD, Szűcs A, Champramary S, Roszik R, Patocskai Z, Münsterkötter M, Huynh T, et al. Towards the Biological Control of Devastating Forest Pathogens from the Genus Armillaria. Forests. 2019; 10(11):1013. https://0-doi-org.brum.beds.ac.uk/10.3390/f10111013
Chicago/Turabian StyleChen, Liqiong, Bettina Bóka, Orsolya Kedves, Viktor Dávid Nagy, Attila Szűcs, Simang Champramary, Róbert Roszik, Zoltán Patocskai, Martin Münsterkötter, Thu Huynh, and et al. 2019. "Towards the Biological Control of Devastating Forest Pathogens from the Genus Armillaria" Forests 10, no. 11: 1013. https://0-doi-org.brum.beds.ac.uk/10.3390/f10111013