Litter Management as a Key Factor Relieves Soil Respiration Decay in an Urban-Adjacent Camphor Forest under a Short-Term Nitrogen Increment

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Experimental Design
2.3. Resin-Core Preparation and Incubation
2.4. Soil and Resin Bags Analyses
2.5. Soil Respiration Measurement
2.6. Data Analysis
3. Results
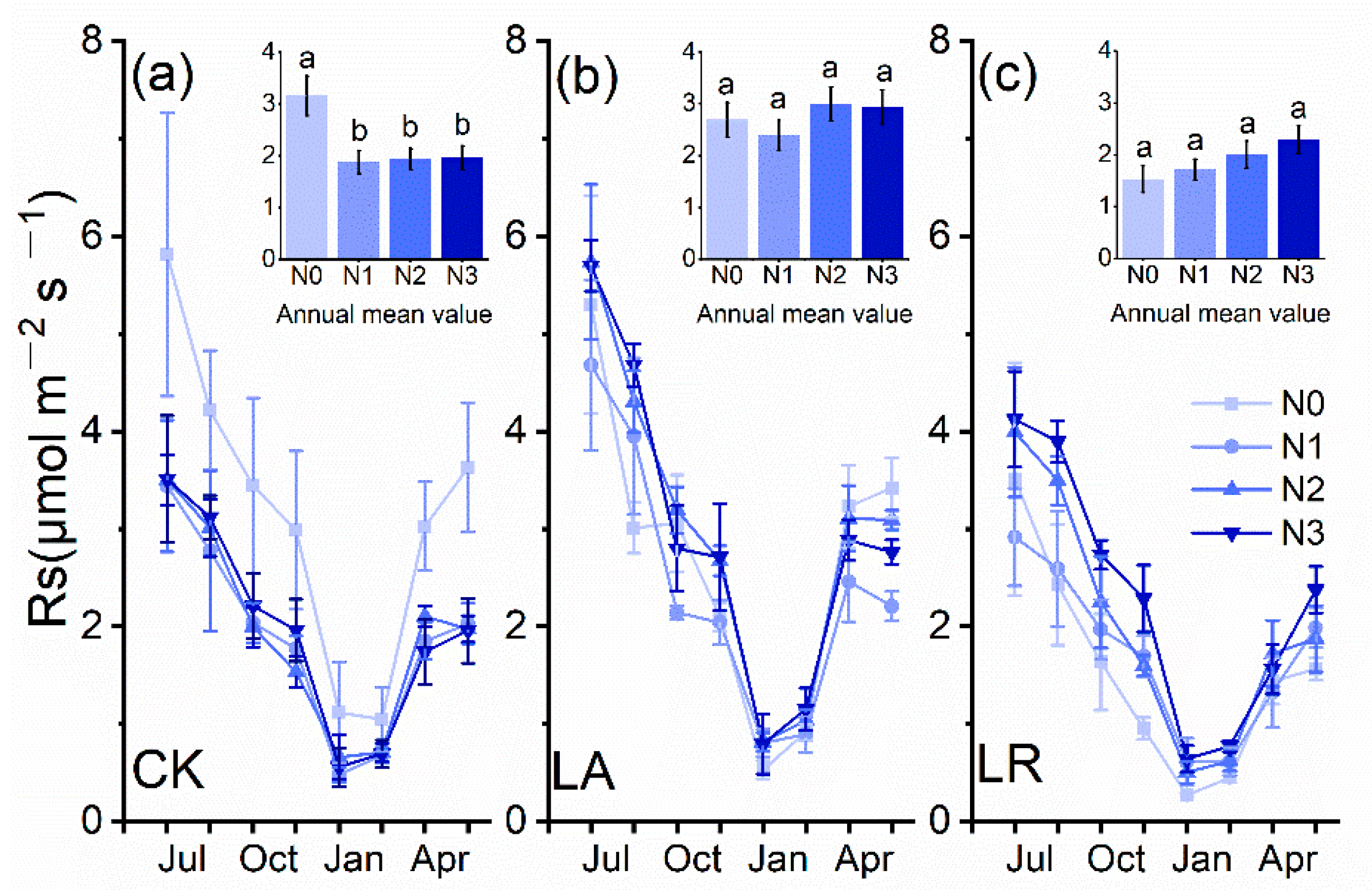
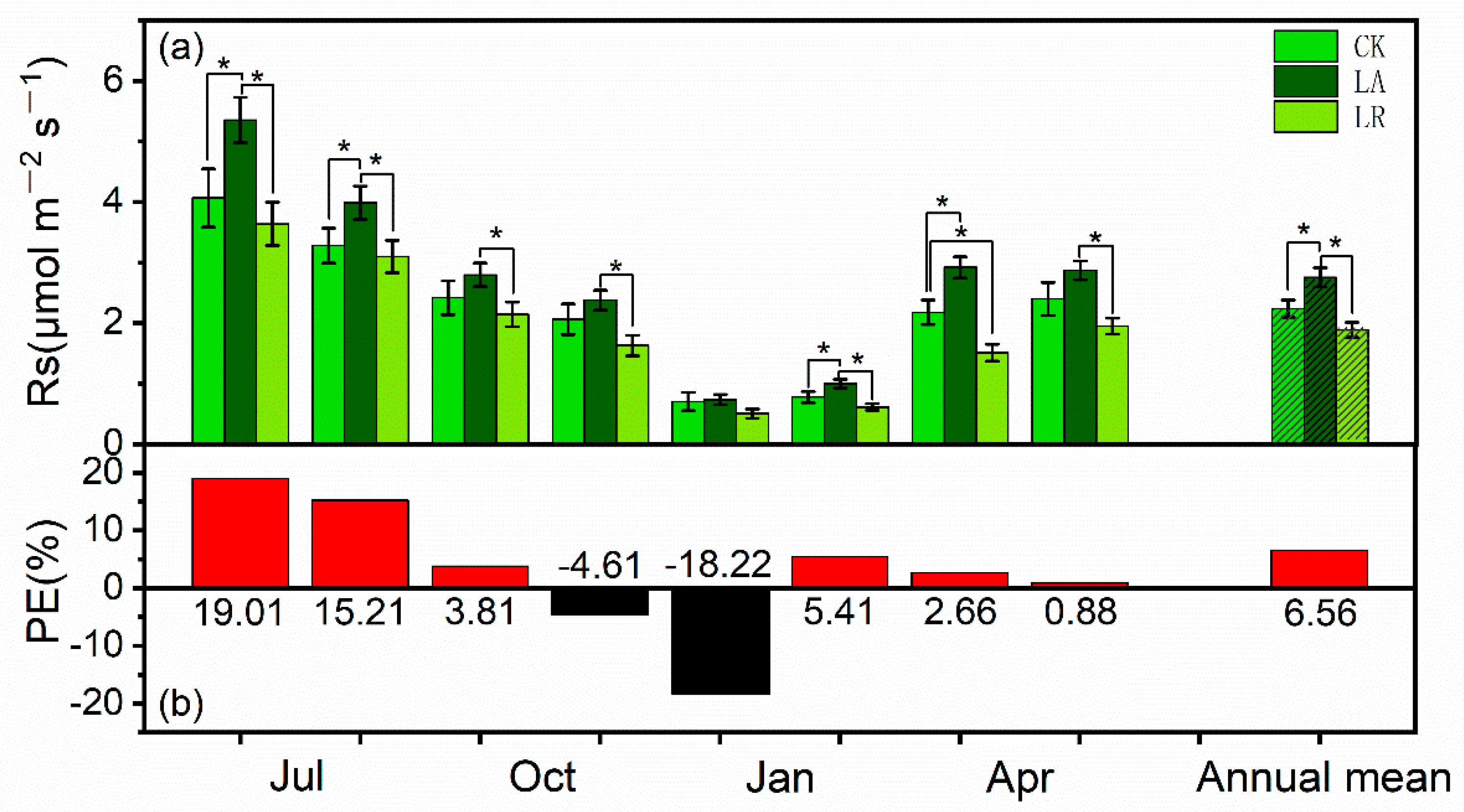
3.1. Soil Respiration (Rs) Rates and Priming Effects (PE)
3.2. Soil Inorganic N and Net N Mineralization
3.3. Effects of N Availability on Soil Respiration
4. Discussion
4.1. Soil Nitrogen Status in Response to N Fertilizers
4.2. Soil Respiration in Response to Incremental N Gradients
4.3. Impacts of Litter Manipulation on N-Saturated Soil
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Solomon, S.; Qin, D.; Manning, M.; Averyt, K.; Marquis, M. Climate Change 2007-the Physical Science Basis: Working Group I Contribution to the Fourth Assessment Report of the IPCC; Cambridge University Press: Cambridge, UK, 2007; Volume 4. [Google Scholar]
- Galloway, J.N.; Townsend, A.R.; Erisman, J.W.; Bekunda, M.; Cai, Z.; Freney, J.R.; Martinelli, L.A.; Seitzinger, S.P.; Sutton, M.A.J.S. Transformation of the nitrogen cycle: Recent trends, questions, and potential solutions. Science 2008, 320, 889–892. [Google Scholar] [CrossRef] [Green Version]
- Gruber, N.; Galloway, J.N. An Earth-system perspective of the global nitrogen cycle. Nature 2008, 451, 293. [Google Scholar] [CrossRef]
- Tian, D.; Niu, S. A global analysis of soil acidification caused by nitrogen addition. Environ. Res. Lett. 2015, 10. [Google Scholar] [CrossRef]
- Zheng, M.; Zhang, W.; Luo, Y.; Li, D.; Wang, S.; Huang, J.; Lu, X.; Mo, J. Stoichiometry controls asymbiotic nitrogen fixation and its response to nitrogen inputs in a nitrogen-saturated forest. Ecology 2018, 99, 2037–2046. [Google Scholar] [CrossRef]
- Phoenix, G.K.; Hicks, W.K.; Cinderby, S.; Kuylenstierna, J.C.I.; Stock, W.D.; Dentener, F.J.; Giller, K.E.; Austin, A.T.; Lefroy, R.D.B.; Gimeno, B.S. Atmospheric nitrogen deposition in world biodiversity hotspots: The need for a greater global perspective in assessing N deposition impacts. Glob. Chang. Biol. 2006, 12, 470–476. [Google Scholar] [CrossRef]
- Cao, J.; Pang, S.; Wang, Q.; Williams, M.A.; Jia, X.; Dun, S.; Yang, J.; Zhang, Y.; Wang, J.; Lü, X.; et al. Plant–bacteria–soil response to frequency of simulated nitrogen deposition has implications for global ecosystem change. Funct. Ecol. 2019. [Google Scholar] [CrossRef]
- Song, L.; Tian, P.; Zhang, J.; Jin, G. Effects of three years of simulated nitrogen deposition on soil nitrogen dynamics and greenhouse gas emissions in a Korean pine plantation of northeast China. Sci. Total Environ. 2017, 609, 1303–1311. [Google Scholar] [CrossRef] [PubMed]
- Reay, D.S.; Dentener, F.; Smith, P.; Grace, J.; Feely, R.A. Global nitrogen deposition and carbon sinks. Nat. Geosci. 2008, 1, 430. [Google Scholar] [CrossRef]
- Yu, Q.; Duan, L.; Yu, L.; Chen, X.; Si, G.; Ke, P.; Ye, Z.; Mulder, J. Threshold and multiple indicators for nitrogen saturation in subtropical forests. Environ. Pollut. 2018, 241, 664–673. [Google Scholar] [CrossRef]
- Bond-Lamberty, B.; Thomson, A. Temperature-associated increases in the global soil respiration record. Nature 2010, 464, 579. [Google Scholar] [CrossRef]
- Janssens, I.A.; Dieleman, W.; Luyssaert, S.; Subke, J.A.; Reichstein, M.; Ceulemans, R.; Ciais, P.; Dolman, A.J.; Grace, J.; Matteucci, G.; et al. Reduction of forest soil respiration in response to nitrogen deposition. Nat. Geosci. 2010, 3, 315–322. [Google Scholar] [CrossRef]
- Chen, Z.; Xu, Y.; He, Y.; Zhou, X.; Fan, J.; Yu, H.; Ding, W. Nitrogen fertilization stimulated soil heterotrophic but not autotrophic respiration in cropland soils: A greater role of organic over inorganic fertilizer. Soil Biol. Biochem. 2018, 116, 253–264. [Google Scholar] [CrossRef]
- Luo, Q.; Gong, J.; Zhai, Z.; Pan, Y.; Liu, M.; Xu, S.; Wang, Y.; Yang, L.; Baoyin, T.-T. The responses of soil respiration to nitrogen addition in a temperate grassland in northern China. Sci. Total Environ. 2016. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Cai, Y.P.; Yang, Z.F.; Yin, X.A.; Tan, Q. Microbial nitrification, denitrification and respiration in the leached cinnamon soil of the upper basin of Miyun Reservoir. Sci. Rep. 2017, 7, 42032. [Google Scholar] [CrossRef] [PubMed]
- Bae, K.; Fahey, T.J.; Yanai, R.D.; Fisk, M. Soil Nitrogen Availability Affects Belowground Carbon Allocation and Soil Respiration in Northern Hardwood Forests of New Hampshire. Ecosystems 2015, 18, 1179–1191. [Google Scholar] [CrossRef]
- Samuelson, L.; Mathew, R.; Stokes, T.; Feng, Y.; Aubrey, D.; Coleman, M. Soil and microbial respiration in a loblolly pine plantation in response to seven years of irrigation and fertilization. For. Ecol. Manag. 2009, 258, 2431–2438. [Google Scholar] [CrossRef]
- Koehler, B.; Corre, M.D.; Veldkamp, E.; Sueta, J. Chronic nitrogen addition causes a reduction in soil carbon dioxide efflux during the high stem-growth period in a tropical montane forest but no response from a tropical lowland forest on a decadal time scale. Biogeosciences 2009, 12, 2973–2983. [Google Scholar] [CrossRef]
- Tu, L.-H.; Hu, T.-X.; Zhang, J.; Li, X.-W.; Hu, H.-L.; Liu, L.; Xiao, Y.-L. Nitrogen addition stimulates different components of soil respiration in a subtropical bamboo ecosystem. Soil Biol. Biochem. 2013, 58, 255–264. [Google Scholar] [CrossRef]
- Xu, Y.; Fan, J.; Ding, W.; Bol, R.; Chen, Z.; Luo, J.; Bolan, N. Stage-specific response of litter decomposition to N and S amendments in a subtropical forest soil. Biol. Fertil. Soils 2016, 52, 711–724. [Google Scholar] [CrossRef]
- Zhang, T.; Chen, H.Y.H.; Ruan, H. Global negative effects of nitrogen deposition on soil microbes. ISME J. 2018, 12, 1817–1825. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Sun, J.; Tian, D.; Wang, J.; Ha, D.; Qu, Y.; Jing, G.; Niu, S. Soil acid cations induced reduction in soil respiration under nitrogen enrichment and soil acidification. Sci. Total Environ. 2017, 615, 1535–1546. [Google Scholar] [CrossRef] [PubMed]
- Fierer, N.; Lauber, C.L.; Ramirez, K.S.; Zaneveld, J.; Bradford, M.A.; Knight, R. Comparative metagenomic, phylogenetic and physiological analyses of soil microbial communities across nitrogen gradients. ISME J. 2012, 6, 1007–1017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maeda, Y.; Tashiro, N.; Enoki, T.; Urakawa, R.; Hishi, T. Effects of species replacement on the relationship between net primary production and soil nitrogen availability along a topographical gradient: Comparison of belowground allocation and nitrogen use efficiency between natural forests and plantations. For. Ecol. Manag. 2018, 422, 214–222. [Google Scholar] [CrossRef]
- Månsson, K.F.; Falkengren-Grerup, U. The effect of nitrogen deposition on nitrification, carbon and nitrogen mineralisation and litter C:N ratios in oak (Quercus robur L.) forests. For. Ecol. Manag. 2003, 179, 455–467. [Google Scholar] [CrossRef]
- Wang, Q.; Wang, S.; He, T.; Liu, L.; Wu, J. Response of organic carbon mineralization and microbial community to leaf litter and nutrient additions in subtropical forest soils. Soil Biol. Biochem. 2014, 71, 13–20. [Google Scholar] [CrossRef]
- Kuzyakov, Y. Priming effects: Interactions between living and dead organic matter. Soil Biol. Biochem. 2010, 42, 1363–1371. [Google Scholar] [CrossRef]
- Prescott, C.E. Litter decomposition: What controls it and how can we alter it to sequester more carbon in forest soils? Biogeochemistry 2010, 101, 133–149. [Google Scholar] [CrossRef]
- Ning, C.; Mueller, G.; Egerton-Warburton, L.; Wilson, A.; Yan, W.; Xiang, W. Diversity and Enzyme Activity of Ectomycorrhizal Fungal Communities Following Nitrogen Fertilization in an Urban-Adjacent Pine Plantation. Forests 2018, 9, 99. [Google Scholar] [CrossRef] [Green Version]
- Gao, Q.; Bai, E.; Wang, J.; Zheng, Z.; Xia, J.; You, W. Effects of litter manipulation on soil respiration under short-term nitrogen addition in a subtropical evergreen forest. For. Ecol. Manag. 2018, 429, 77–83. [Google Scholar] [CrossRef]
- Yan, W.-D.; Chen, X.-Y.; Peng, Y.-Y.; Zhu, F.; Zhen, W.; Zhang, X.-Y. Response of Soil Respiration to Nitrogen Addition in Two Subtropical Forest Types *1. Pedosphere 2017. [Google Scholar] [CrossRef]
- Zou, X.; Valentine, D.W.; Sanford, R.L.; Binkley, D. Resin-core and buried-bag estimates of nitrogen transformations in Costa Rican lowland rainforests. Plant Soil 1992, 139, 275–283. [Google Scholar] [CrossRef]
- Liu, X.-R.; Dong, Y.-S.; Ren, J.-Q.; Li, S.-G. Drivers of soil net nitrogen mineralization in the temperate grasslands in Inner Mongolia, China. Nutr. Cycl. Agroecosyst. 2009, 87, 59–69. [Google Scholar] [CrossRef]
- Deng, H.P.; Wang, G.J.; Geng, G. Response of nitrogen mineralization to litter addition and exclusion in soils of Cinnamomum camphora plantation. J. Beijing For. Univ. 2010, 32, 47–51. [Google Scholar]
- Nelson, D.W.; Sommers, L.E. Total carbon, organic carbon, and organic matter. In Methods of Soil Analysis Part 3. Chemical Methods; Soil Science Society of America Book Series No. 5; Soil Science Society of America, Inc.: Madison, WI, USA, 1996. [Google Scholar]
- Jackson, M.L. Soil Chemical Analysis—Advanced Course; University of Wisconsin: Madison, WI, USA, 1979. [Google Scholar]
- Meyer, N.; Welp, G.; Amelung, W. The Temperature Sensitivity (Q10) of Soil Respiration: Controlling Factors and Spatial Prediction at Regional Scale Based on Environmental Soil Classes. Glob. Biogeochem. Cycles 2018, 32, 306–323. [Google Scholar] [CrossRef]
- Lefcheck, J.S.; Freckleton, R. piecewiseSEM: Piecewise structural equation modelling inr for ecology, evolution, and systematics. Methods Ecol. Evol. 2016, 7, 573–579. [Google Scholar] [CrossRef]
- Isobe, K.; Ikutani, J.; Fang, Y.; Yoh, M.; Mo, J.; Suwa, Y.; Yoshida, M.; Senoo, K.; Otsuka, S.; Koba, K. Highly abundant acidophilic ammonia-oxidizing archaea causes high rates of nitrification and nitrate leaching in nitrogen-saturated forest soils. Soil Biol. Biochem. 2018, 122, 220–227. [Google Scholar] [CrossRef]
- Yu, L.; Kang, R.; Mulder, J.; Zhu, J.; Dörsch, P. Distinct fates of atmogenic NH4+ and NO3− in subtropical, N-saturated forest soils. Biogeochemistry 2017, 133, 279–294. [Google Scholar] [CrossRef]
- Lovett, G.M.; Goodale, C.L. A new conceptual model of nitrogen saturation based on experimental nitrogen addition to an oak forest. Ecosystems 2011, 14, 615–631. [Google Scholar] [CrossRef]
- Niu, S.; Classen, A.T.; Dukes, J.S.; Kardol, P.; Liu, L.; Luo, Y.; Rustad, L.; Sun, J.; Tang, J.; Templer, P.H.; et al. Global patterns and substrate-based mechanisms of the terrestrial nitrogen cycle. Ecol. Lett. 2016, 19, 697–709. [Google Scholar] [CrossRef] [Green Version]
- Mo, J.; Zhang, W.E.I.; Zhu, W.; Gundersen, P.E.R.; Fang, Y.; Li, D.; Wang, H.U.I. Nitrogen addition reduces soil respiration in a mature tropical forest in southern China. Glob. Chang. Biol. 2008, 14, 403–412. [Google Scholar] [CrossRef]
- Sun, Z.; Liu, L.; Ma, Y.; Yin, G.; Zhao, C.; Zhang, Y.; Piao, S. The effect of nitrogen addition on soil respiration from a nitrogen-limited forest soil. Agric. For. Meteorol. 2014, 197, 103–110. [Google Scholar] [CrossRef]
- Sun, Q.; Wang, R.; Wang, Y.; Du, L.; Zhao, M.; Gao, X.; Hu, Y.; Guo, S. Temperature sensitivity of soil respiration to nitrogen and phosphorous fertilization: Does soil initial fertility matter? Geoderma 2018, 325, 172–182. [Google Scholar] [CrossRef]
- Peng, Y.; Thomas, S.C. Influence of Non-nitrogenous Soil Amendments on Soil CO2 Efflux and Fine Root Production in an N-Saturated Northern Hardwood Forest. Ecosystems 2010, 13, 1145–1156. [Google Scholar] [CrossRef]
- Fan, H.; Wu, J.; Liu, W.; Yuan, Y.; Huang, R.; Liao, Y.; Li, Y. Nitrogen deposition promotes ecosystem carbon accumulation by reducing soil carbon emission in a subtropical forest. Plant Soil 2014, 379, 361–371. [Google Scholar] [CrossRef]
- Chen, H.; Li, D.; Gurmesa, G.A.; Yu, G.; Li, L.; Zhang, W.; Fang, H.; Mo, J. Effects of nitrogen deposition on carbon cycle in terrestrial ecosystems of China: A meta-analysis. Environ. Pollut. 2015, 206, 352–360. [Google Scholar] [CrossRef]
- Lohse, K.A.; Matson, P. Consequences of nitrogen additions for soil processes and solution losses from wet tropical forests. Ecol. Appl. 2005, 15, 1629–1648. [Google Scholar] [CrossRef]
- Bowden, R.D.; Davidson, E.; Savage, K.; Arabia, C.; Steudler, P. Chronic nitrogen additions reduce total soil respiration and microbial respiration in temperate forest soils at the Harvard Forest. For. Ecol. Manag. 2004, 196, 43–56. [Google Scholar] [CrossRef]
- Treseder, K.K. Nitrogen additions and microbial biomass: A meta-analysis of ecosystem studies. Ecol. Lett. 2008, 11, 1111–1120. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Zhang, X.-Y.; Zou, H.-T.; Kou, L.; Yang, Y.; Wen, X.-F.; Li, S.-G.; Wang, H.-M.; Sun, X.-M. Contrasting effects of ammonium and nitrate additions on the biomass of soil microbial communities and enzyme activities in subtropical China. Biogeosciences 2017, 14, 4815–4827. [Google Scholar] [CrossRef] [Green Version]
- Cleveland, C.C.; Houlton, B.Z.; Smith, W.K.; Marklein, A.R.; Reed, S.C.; Parton, W.; Del Grosso, S.J.; Running, S.W. Patterns of new versus recycled primary production in the terrestrial biosphere. Proc. Natl. Acad. Sci. USA 2013, 110, 12733–12737. [Google Scholar] [CrossRef] [Green Version]
- Rennenberg, H.; Dannenmann, M. Nitrogen Nutrition of Trees in Temperate Forests—The Significance of Nitrogen Availability in the Pedosphere and Atmosphere. Forests 2015, 6, 2820–2835. [Google Scholar] [CrossRef]
- Miao, R.; Ma, J.; Liu, Y.; Liu, Y.; Yang, Z.; Guo, M. Variability of Aboveground Litter Inputs Alters Soil Carbon and Nitrogen in a Coniferous–Broadleaf Mixed Forest of Central China. Forests 2019, 10, 188. [Google Scholar] [CrossRef] [Green Version]
- Riaz, M.; Mian, I.A.; Cresser, M.S. Litter effects on ammonium dynamics in an acid soil under grassland. Geoderma 2010, 159, 198–208. [Google Scholar] [CrossRef]
- Nair, R.K.; Perks, M.P.; Mencuccini, M. Decomposition nitrogen is better retained than simulated deposition from mineral amendments in a temperate forest. Glob. Chang. Biol. 2017, 23, 1711–1724. [Google Scholar] [CrossRef] [Green Version]
- Joly, F.-X.; Fromin, N.; Kiikkilä, O.; Hättenschwiler, S. Diversity of leaf litter leachates from temperate forest trees and its consequences for soil microbial activity. Biogeochemistry 2016, 129, 373–388. [Google Scholar] [CrossRef]
- Chen, X.; Chen, H.Y.H. Global effects of plant litter alterations on soil CO2 to the atmosphere. Glob. Chang. Biol. 2018, 24, 3462–3471. [Google Scholar] [CrossRef]
- Eberwein, J.R.; Oikawa, P.Y.; Allsman, L.A.; Jenerette, G.D. Carbon availability regulates soil respiration response to nitrogen and temperature. Soil Biol. Biochem. 2015, 88, 158–164. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Basic Soil Characteristics (n = 36) | |
|---|---|
| Soil bulk density (g cm−3) | 1.50 ± 0.11 |
| Soil pH | 3.98 ± 0.22 |
| Total organic carbon (mg g−1) | 13.97 ± 1.70 |
| Total nitrogen (mg g−1) | 1.34 ± 0.14 |
| C/N | 10.43 |
| Litter layer thickness | 1–2 cm |
| Annual litterfall production (kg m−2 year−1) | 0.45 ± 0.03 |
| Rs | (NH4-N) | (NO3-N) | RA | RN | RM | |
|---|---|---|---|---|---|---|
| N fertilizers (N) | 0.535 | 0.490 | <0.001 a | 0.561 | <0.001 a | 0.161 |
| Litter inputs (L) | <0.001 a | 0.001 a | 0.028 a | 0.841 | 0.061 | 0.287 |
| Seasonal (S) | <0.001 a | <0.001 a | 0.012 a | 0.015 a | 0.009 a | <0.001 a |
| N × L | 0.449 | 0.699 | 0.003 a | 0.983 | 0.256 | 0.581 |
| N × S | 0.284 | 0.059 | <0.001 a | 0.112 | <0.001 a | 0.584 |
| L × S | 0.505 | 0.095 | <0.001 a | 0.551 | 0.672 | 0.451 |
| N × L × S | 0.892 | 0.102 | <0.001 a | 0.509 | 0.924 | 0.509 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, X.; Li, Y.; Ning, C.; Zheng, W.; Zhao, D.; Li, Z.; Yan, W. Litter Management as a Key Factor Relieves Soil Respiration Decay in an Urban-Adjacent Camphor Forest under a Short-Term Nitrogen Increment. Forests 2020, 11, 216. https://0-doi-org.brum.beds.ac.uk/10.3390/f11020216
Zhang X, Li Y, Ning C, Zheng W, Zhao D, Li Z, Yan W. Litter Management as a Key Factor Relieves Soil Respiration Decay in an Urban-Adjacent Camphor Forest under a Short-Term Nitrogen Increment. Forests. 2020; 11(2):216. https://0-doi-org.brum.beds.ac.uk/10.3390/f11020216
Chicago/Turabian StyleZhang, Xuyuan, Yong Li, Chen Ning, Wei Zheng, Dayong Zhao, Ziqian Li, and Wende Yan. 2020. "Litter Management as a Key Factor Relieves Soil Respiration Decay in an Urban-Adjacent Camphor Forest under a Short-Term Nitrogen Increment" Forests 11, no. 2: 216. https://0-doi-org.brum.beds.ac.uk/10.3390/f11020216