Three Invasive Tree Species Change Soil Chemistry in Guam Forests


College of Natural and Applied Sciences, University of Guam, Mangilao, GU 96923, USA
Forests 2020, 11(3), 279; https://0-doi-org.brum.beds.ac.uk/10.3390/f11030279
Submission received: 28 January 2020
/
Revised: 23 February 2020
/
Accepted: 26 February 2020
/
Published: 28 February 2020
(This article belongs to the Section Forest Ecology and Management)
Abstract
:Research Highlights: Established stands of Leucaena leucocephala (Lam.) de Wit, Spathodea campanulata P. Beauv., and Vitex parviflora Juss. modified soils in Guam’s limestone forests, reducing storage pools of carbon, nitrogen, and phosphorus. Background and Objectives: Invasive plants may engineer negative changes in ecosystem properties. This study was conducted to determine changes in soil chemistry following infestations of three problematic tree species on Guam. Materials and Methods: Minerals, metals, and mineralization dynamics were measured in invaded sites and paired sites with biodiverse native tree cover. Results: Most soil properties were significantly changed by long-term infestations of the invasive tree species. The soils within invaded sites exhibited total carbon, total nitrogen, and available phosphorus that were less than native sites. In contrast, the carbon/nitrogen ratio increased for every species-site combination. The other chemical properties were idiosyncratic among the sites and species. Conclusions: Mitigation and restoration activities that include the removal of these trees from project sites may require many years for the below-ground ecosystems to return to their native state. These three invasive trees decrease the ability of Guam soils to sequester recalcitrant forms of carbon, nitrogen, and phosphorus.
1. Introduction
Non-native plant species may cause profound changes to invaded forests. Species that are able to displace resident native plants may create unique habitat process rates and controls which affect ecosystem nutrient and carbon cycles [1]. Some invasive plants may alter habitat functioning to the extent that ecosystem services are threatened [2,3,4,5]. Additionally, the changes that are engineered by some invasive plant species may adjust soil properties in a manner that benefits their own continued competitive growth [6,7,8,9]. The invasive plant species that possess the ability to engineer such consequential changes to forested habitats are sometimes called transformer species [10]. Recent assessments of plant invasion research illuminate the need to distinguish non-native species that carry minimal risk from invasive species that are high risk and link the likelihood of naturalization to these consequential impacts on the invaded habitats [11,12].
No international consensus exists concerning how to measure the impacts of plant invasions, and case study conclusions may be idiosyncratic due to inconsistent methods [13,14]. Often the impacts of non-native plant species are circumstantial and contingent on traits of the invaded habitat [15]. These context-dependent characteristics of plant invasions indicate that research outcomes from distant localities or invasive species that are not locally pertinent may have little relevance for understanding local ecosystem responses to plant invasions [16]. This means that an understanding of the impacts of plant invasions on tropical island forests need to be studied under local conditions, as the vast majority of published research on the subject of plant invasions is biased toward temperate habitats of continental North America and Europe [15].
The Mariana Island of Guam has been showcased as an insular example of ecosystem damage due to invasive species, and the effectiveness of responsive programs have begun to be studied [17]. While most of the funding and research focus has been on animal invasions, three non-native tree species have established a foothold on the calcareous soils of the island. Leucaena leucocephala is from Central America and is among the most serious environmental weeds worldwide [18]. Copious seed production, persistent seed bank, and ability to capitalize on disturbed sites are among the traits that facilitate its invasive potential [19]. Allelopathic methods are employed by the tree to exclude competitive plant growth [20]. The invasion of this aggressive legume has been shown to influence successional pathways and community assembly [21]. The properties of soils beneath L. leucocephala stands have been reported from the Mariana Island of Tinian [22]. The results indicated increased mineralization generated losses from the forest ecosystem such that recalcitrant carbon and nitrogen pools decreased. No comparable study has been reported from Guam. Spathodea campanulata is from tropical Africa and has been introduced throughout the tropical regions of the world for use as street trees [23]. The tree naturalizes in many habitat types to displace native trees, especially in fertile soils. Vitex parviflora is from tropical Asia and is used in urban landscapes outside of its native range. This tree is the only large tree species that exhibited a substantial increase in the Guam population between a 2002 forest inventory and a 2013 forest inventory [24].
These three tree species are among the invasive plants that impose overt threats to the health of Pacific island ecosystems [25]. They are particularly skilled at displacing existing native tree species in Guam forests to occupy large areas with mono-specific closed canopy. They occupy vast areas of Guam lands that are under the custody of the United States government. A massive military buildup is currently being imposed on Guam [26], and restoration efforts of forests are a component of the accompanying terrestrial resource management plans. Therefore, the role of these three invasive tree species in ecosystem degradation needs to be studied to ensure adequate knowledge for guiding funded conservation and restoration strategies. However, to my knowledge no studies have been funded or carried out toward that goal. My first objective was to repeat the methods that were used in Tinian [22] to determine if the changes in two Guam soils caused by L. leucocephala were consistent with the results from Tinian. My second objective was to use the same methods to determine the changes that occurred in Guam following invasive establishment of V. parviflora and S. campanulata.
2. Materials and Methods
Two soil series were included for this study. One series was a sandy soil which was formed in water-deposited coral sand derived from coral reef formations (Carbonatic, isohyperthermic Typic Ustipsamments) [27]. Leucaena leucocephala was the only tree species studied on these soils. The other locations were situated on the uplifted plateau where the dominant soil is Guam cobbly clay. These soils were formed from sediment overlying porous limestone on uplifted plateaus (Clayey, gibbsitic, nonacid, isohyperthermic Lithic Ustorthents) [27]. Most of the forest locations that are currently used for mitigation and conservation activities associated with the ongoing military buildup [26] are located on this clay soil series. All three tree species were studied on this soil.
2.1. Field Sampling
Within each habitat, large stands of the non-native trees were selected as “invaded” sampling locations. The size of the mono-specific stands varied, and was 9–15 ha for V. parviflora, 1.2–7 ha for L leucocephala, and 0.6–0.7 ha for S. campanulata. The study sites were ecotones where the distinct edge of the invaded locations was adjacent to secondary native forest, and paired locations within these multi-species forests were used as “native” sampling locations. The field work was conducted on 15 September 2014 for V. parviflora, on 6 November 2014 for L. leucocephala, and on 7 November 2014 for S. campanulata.
Paired sampling locations were positioned close to the ecotone edge, whereby the invaded site and paired native site were each located 10 m from the edge. Therefore, each pair of samples included soil sampling sites that were 20 m apart. Soil was harvested from the 0–15 cm stratum for each sample. At each sampling site, a linear transect 9 m in length was positioned perpendicular to the ecotone edge. Ten subsamples were collected at 1 m intervals along the transect, and combined into one homogenized sample to form one replication. Six locations were selected for each tree species, yielding a total of six replications. This approach required the collection of 120 soil cores for each species-soil combination, and 480 soil cores for the entire study.
The dominant native trees in the L. leucocephala study site on the sand soils were Artocarpus mariannensis Trécul, Morinda citrifolia L., and Pandanus tectorius Parkinson ex Du Roi. The dominant native trees in the L. leucocephala study site on clay soils were A. mariannensis, Hibiscus tiliaceus L., and M. citrifolia. The dominant native trees in the V. parviflora site were Elaeocarpus joga Merr., M. citrifolia, and P. tectorius. The dominant native trees in the S. campanulata study site were A. mariannensis, Eugenia L. sp., and H. tiliaceus.
2.2. Analyses and Incubations
Each fresh soil replication was separated into three subsamples. One subsample was dried at 105 °C for standard chemical assays. Total carbon and nitrogen contents were determined by dry combustion (FLASH EA1112 CHN analyzer; Thermo Fisher, Waltham, MA., USA). Available P was determined by the Olsen method [28]. Extractable essential minerals were quantified following digestion with diethylenetriaminepentaacetic acid [29]. Total metal content was quantified following digestion with nitric acid [30]. Analysis was by inductively coupled plasma optical emission spectrometry (Spectro Genesis; SPECTRO Analytical Instruments, Kleve, Germany). The second fresh subsample was used to determine nitrate and ammonium colorimetrically following 2M potassium chloride extraction [31]. The third fresh subsample was incubated in a mesocosm constructed with the same field soil for about one month, using the buried bag method [32]. Vitex parviflora soils were harvested from the mesocosm on 17 October 2014 for 32 days (temperature 24–33 °C), L. leucocephala soils were harvested on 8 December 2014 for 32 days (temperature 25–34 °C), and S. campanulata soils were harvested on 8 December 2014 for 31 days (temperature 24–34 °C). Nitrate and ammonium were quantified at the end of the incubation period. Net nitrification was calculated by subtracting initial from final nitrate concentration and dividing by the duration of the incubation period. Net ammonification was calculated by subtracting initial from final ammonium concentration and dividing by the duration of the incubation period. Net mineralization was calculated as the sum of nitrification and ammonification. The choice to use a mesocosm for the incubations was to provide relative values that were comparable for the three species in this study. Moreover, the feral pig (Sus scrofa L.) population on Guam is beyond control, and inadvertent vandalism of experimental sites like freshly dug incubation sites is common. The use of the mesocosm mitigated these threats to experimental methods that accompany the use of field incubations.
2.3. Statistics
Variables that met parametric requirements were subjected to paired t test to determine differences between invaded and native soils. Some of the rare metals were undetected or present in minute quantities in some of the replications, and in these cases no transformations were able to satisfy parametric requirements. For these metals, the Mann-Whitney U test [33] was used, as this test does not require any assumption about distribution of the data.
3. Results
3.1. Vitex parviflora
The pH, calcium, and zinc soil concentrations were not influenced by V. parviflora, but the remainder of properties differed between the V. parviflora stands and the native forest (Table 1, Figure 1). The concentrations of every mineral and metal within the V. parviflora sites were less than the paired native sites, with the exception of selenium. The concentration of this rare metal in the invaded sites was 50% greater than in the native tree sites.
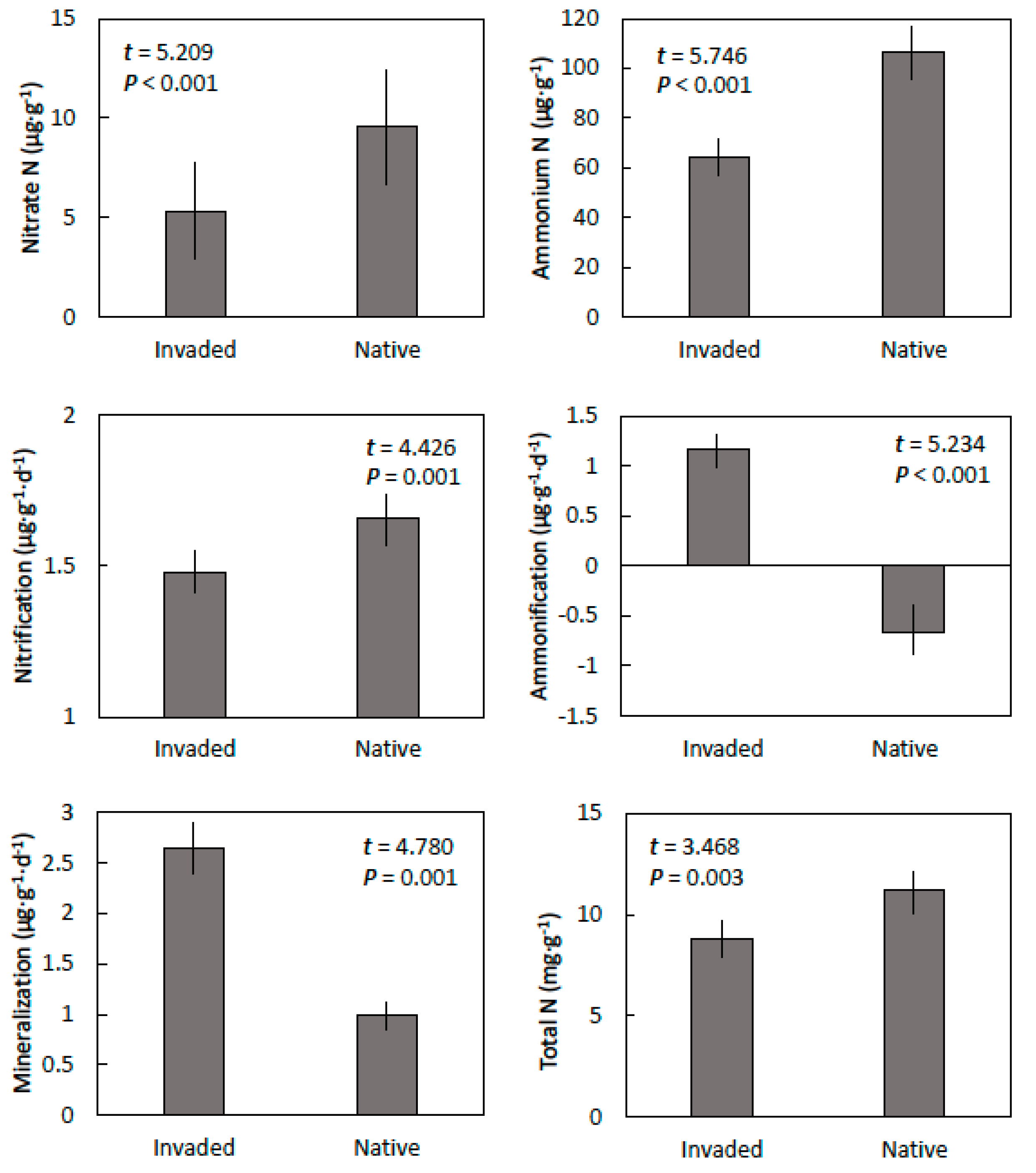
Nitrate and ammonium concentrations were greater in the soils from the native sites than from the invaded sites occupied by V. parviflora (Figure 1). Therefore, bioavailable nitrogen was much greater in the soils from the native sites than in the invaded sites. Net nitrification was less but net ammonification was greater in the soils from the invaded site when compared with the native sites. Therefore, net mineralization of organic nitrogen to available nitrogen was more than 2.5-fold greater within the soils from the invaded sites than the native sites. In contrast, total nitrogen was greater in the soil from the native sites than in the invaded sites with V. parviflora canopy.
3.2. Leucaena leucocephala
In the Guam cobbly clay locations, every measured soil property was influenced by L. leucocephala with the exception of the concentrations of cobalt, copper, and zinc (Table 2). Soil pH, carbon, and phosphorus concentrations were greater in the native sites than in the invaded sites. The remainder of the significantly influenced mineral and metal concentrations were greater in the invaded sites than in the native sites.
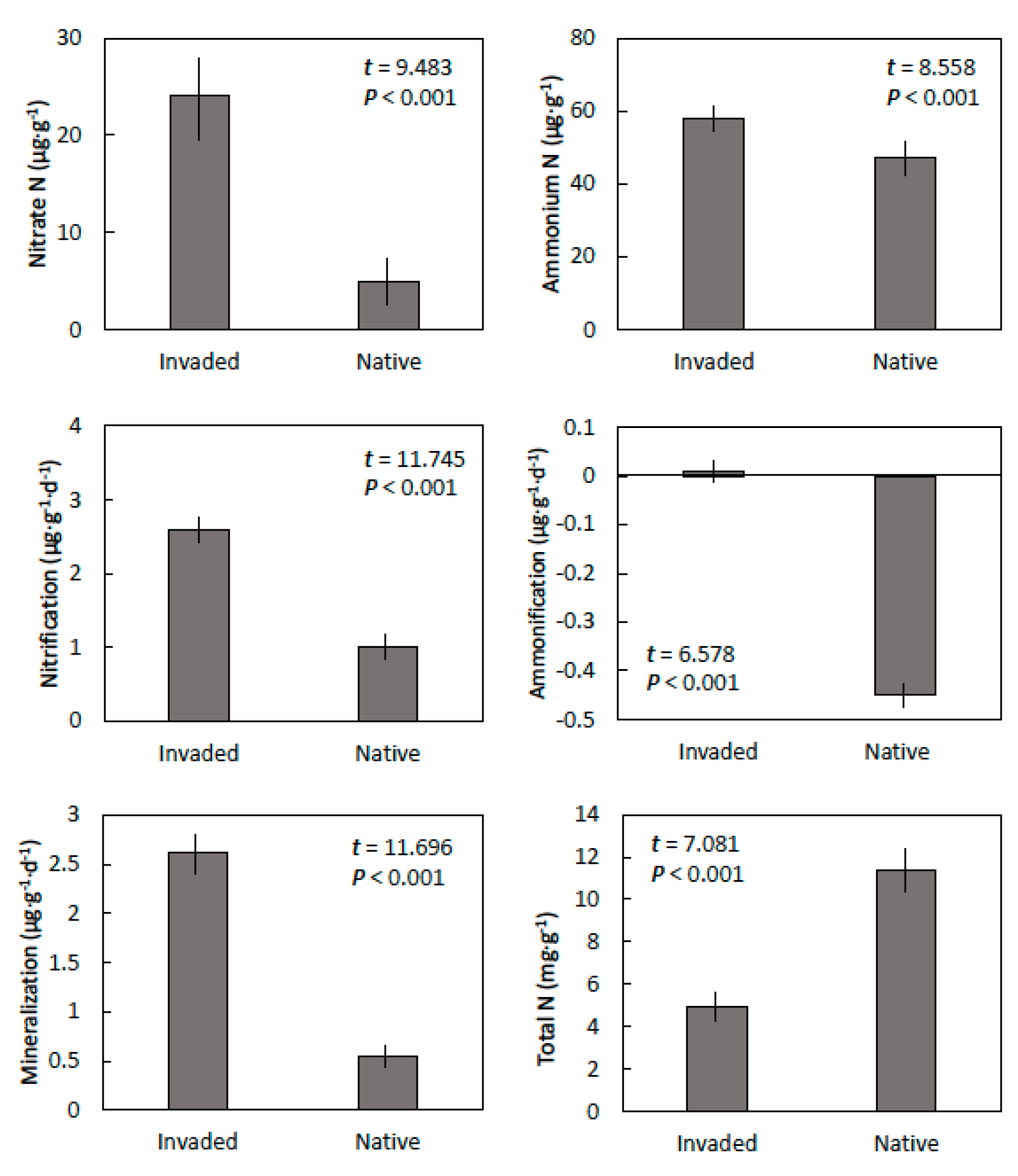
Nitrate and ammonium concentrations in this clay soil were greater in invaded sites than in the native sites (Figure 2). Net nitrification and net ammonification were also greater in the soils from the invaded sites than from the native sites. Therefore, net mineralization of organic nitrogen to available nitrogen was more than 5-fold greater within the soil from the invaded sites than the native sites. In contrast, total nitrogen was greater in the soil from the native sites than in the sites that were occupied by L. leucocephala.
In the coastal sand location, every measured soil property was influenced by L. leucocephala with the exception of soil pH, calcium, and chromium (Table 3). Potassium and selenium concentrations were greater in the invaded sites than in the native sites. The remainder of the significantly influenced mineral and metal concentrations were greater in the native sites than in the invaded sites.
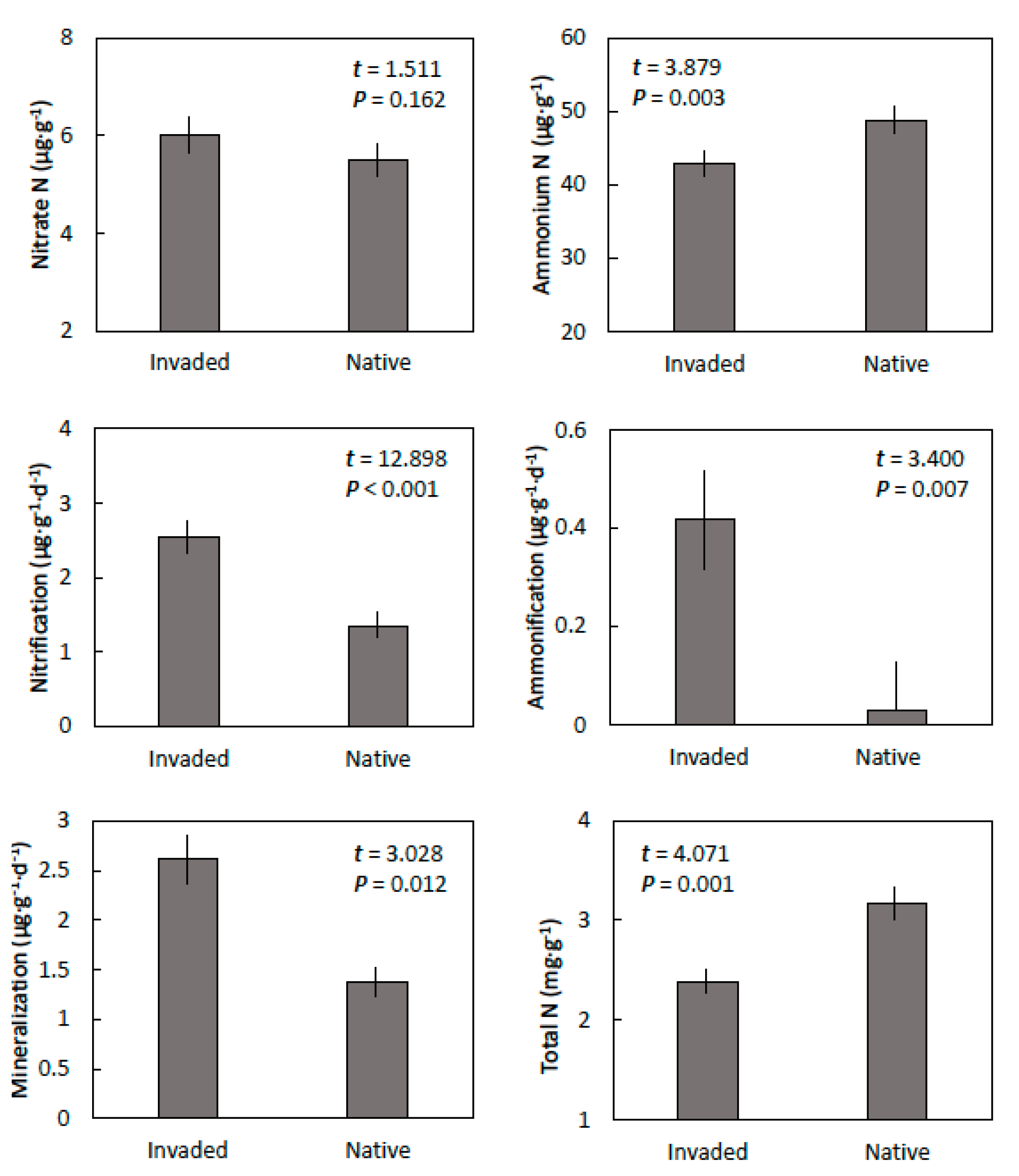
Nitrate concentration did not differ between the soils from invaded and native sites, but ammonium concentrations were greater in soils from the native sites than from the invaded sites (Figure 3). Net nitrification and net ammonification were also greater in the soils from the invaded sites than from the native sites. Therefore, net mineralization of organic nitrogen to available nitrogen of the soils from the invaded sites was about double that of the soils from the native sites. In contrast, total nitrogen was greater in the soils from the native sites than in the sites that were occupied by L. leucocephala.
3.3. Spathodea campanulata
Soil potassium, magnesium, cobalt, and copper concentrations were not influenced by S. campanulata, but the remainder of measured properties differed between the invaded stands and the native forest (Table 4). Soil pH, carbon, phosphorus, calcium, and selenium concentrations were greater in the native sites than in the invaded sites. In contrast, manganese, iron, chromium, nickel, and zinc concentrations were greater in the invaded sites than in the native sites.
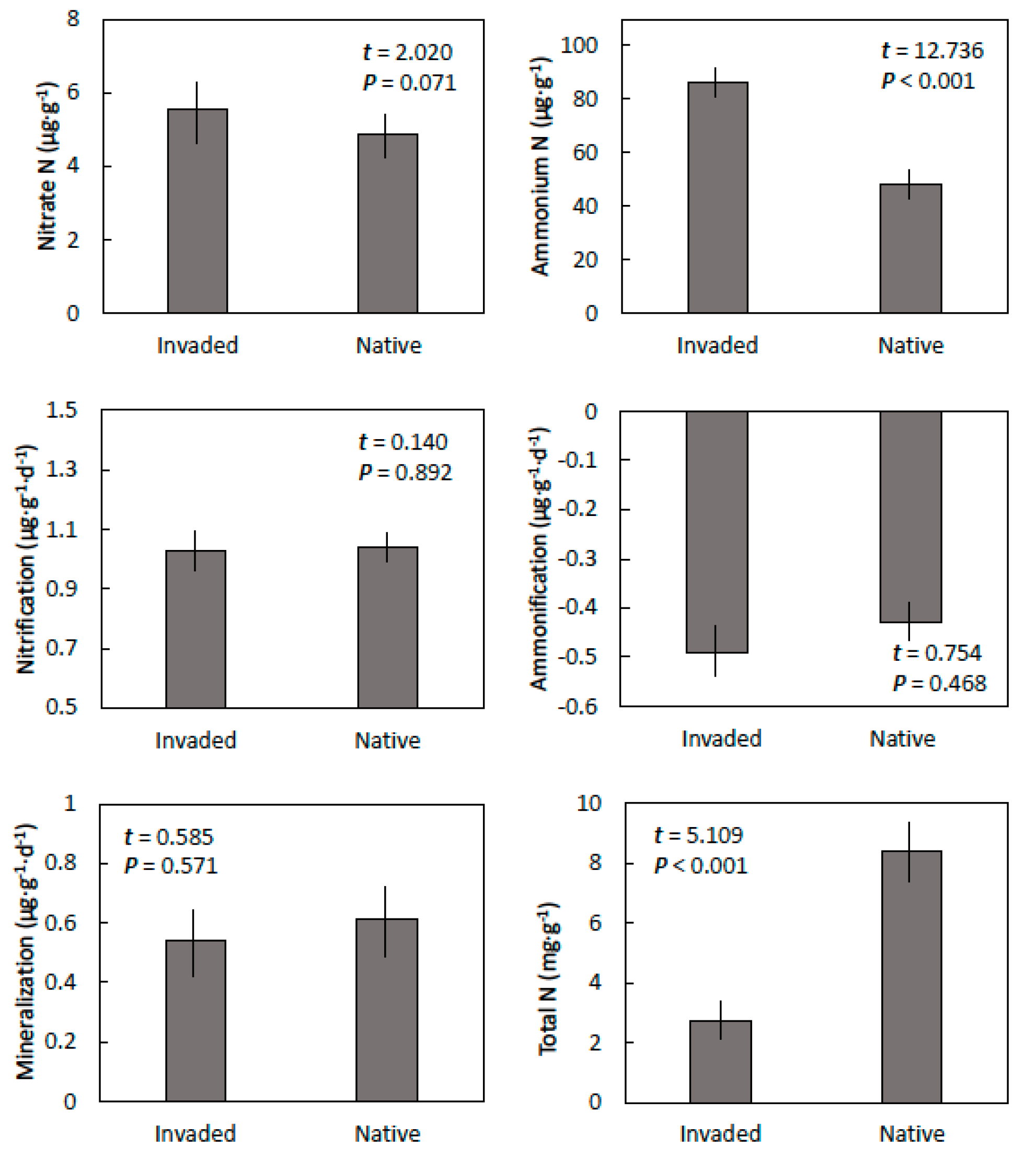
Nitrate concentration did not differ between the soils from the invaded and native sites, but ammonium concentrations were greater in soils from invaded sites than soils from the native sites (Figure 4). Net nitrification, net ammonification, and net mineralization also did not differ between the two sites. In contrast, total nitrogen was greater in the soils from the native sites than from the sites that were occupied by S. campanulata.
4. Discussion
Non-native plant species may alter soil properties in their invaded habitat, but the magnitude and direction of these changes are sometimes unique to each case study [15,34]. Attempts to define canonical drivers of how invasive plants degrade soil quality are hindered by the fact that most of the published research on the subject is biased toward temperate habitats of North America and Europe [15]. My addition of three prominent invasive trees to this research agenda addresses this bias by adding a tropical island ecosystem to the relevant literature. For the island of Guam, perhaps the most important plant trait predicting large-scale negative impacts on soil is the ability to commandeer the emergent forest canopy to displace other resident plants, then expand in occupancy to encompass large areas with mono-specific closed canopy. The trees L. leucocephala and V. parviflora are exceptionally adept at this in Guam’s forests. This first Guam case study on biogeochemistry of invaded sites revealed that L. leucocephala, S. campanulata, and V. parviflora act as transformer species by engineering substantial changes to soil chemistry. Carbon, nitrogen, and phosphorus are highly influential of many ecosystem processes, and the direction of change for these three elements was universal among the species and sites in this study. The biodiverse native soils always exhibited greater carbon, nitrogen, and phosphorus than the invaded soils. These changes following the invasions clearly decreased the recalcitrant forms of these elements, indicating further research is needed to fully understand the negative consequences of these invasive tree species to in situ carbon and nitrogen cycles.
Ecological stoichiometry based on carbon:nitrogen:phosphorus relations is often invoked to understand ecosystem function and cycles [35]. Carbon/nitrogen always increased in the invaded Guam sites when compared with the paired native sites. In contrast, carbon/phosphorus and nitrogen/phosphorus increased for some of the trees but decreased for others. These outcomes indicate the relative decreases in soil carbon and nitrogen within the invaded soils were similar among the species, but the relative decreases in phosphorus were dissimilar to the other elements among the species.
The invaded range of L. leucocephala is substantial, and the only soils in the Mariana Islands that are not invaded by this legume tree are the acid soils. The two soils in this study and the soil from the Tinian study [22] exhibited several similarities in the changes imposed by L. leucocephala invasion. This invasive tree reduced total carbon, total nitrogen, and net ammonification in all of these soils when compared with the soils under adjacent native biodiverse forest cover. In contrast, net nitrification and net mineralization of the soils beneath L. leucocephala stands were always greater than the soils beneath native tree cover. Several of the measured soil properties exhibited dissimilar responses to the presence of chronic L. leucocephala cover, indicating idiosyncratic responses that may be mediated by soil properties or the traits of the native species at each locality.
The differences in the minerals and metals that were measured in the soils beneath the invasive tree stands versus the adjacent native biodiverse forest cover could have been due to background spatial heterogeneity. My methods of selecting distinct edges to the ecotones defined by invaded versus native cover allowed the paired sampling sites to be only 20 m apart. Therefore, disparity in the sequestration of the minerals and metals within the sizeable plant organs was the most likely explanation for the differences in soil mineral and metal concentrations.
The Integrated Natural Resources Management Plan for the Mariana Islands [36] defined the control of invasive species as a critical endeavor. The Environmental Impact Statement for the Guam and Tinian military relocation [37] specified control of these and other non-native trees as necessary to avert continued forest degradation. Numerous resource assessments and proposed management plans that have been conducted for military operations in the Mariana Islands call for the removal of these and other invasive species [38,39,40,41,42]. Despite this institutionalized acknowledgement of landscape dysfunction due to these invasive tree species, to date there have been no empirical studies funded and conducted to identify the means by which these trees damage Guam’s forests. The lack of funded research on this topic is surprising, given the responsibilities of the United States government to manage large expanses of lands under federal control.
Forest enhancement projects are underway on Guam as a component of mitigation efforts associated with the military buildup. My results illuminate three relevant issues for consideration. First, the substantial changes to the soils caused by invasive plants may persist long after removal of the non-native species from a project site, and these invasion legacies may create persistent negative soil conditions that reduce objectives of reestablishing native vegetation [43,44,45,46,47,48,49,50]. Therefore, the culling of invasive trees from project sites should be viewed as a first step in a long process of restoring the soils to their native state. Second, planting native trees to enhance degraded forest locations requires consideration of which native species to select for this purpose. Threatened tree species that are listed on the United States Endangered Species Act [51] are the subject of salvage projects on Guam, and mature trees are being rescued from construction sites and transplanted to recipient forest enhancement areas [52]. Until further research is conducted, the use of these threatened trees for restoring degraded forests with native vegetation is not advisable if the recipient sites have historical infestations of L. leucocephala, V. parviflora, or S. campanulata. Indeed, many failures in reintroductions of threatened species have been due to selection of poor habitat conditions and an inability of conservationists to provide the conditions required to achieve self-sustaining reintroduced populations [53]. Healthy Guam recipient sites with native vegetation would better provide the transplanted threatened trees with abiotic and biotic traits that these trees co-evolved with for their successful establishment. Third, restoration projects vary in the form of interventions from passive approaches to full-scale reclamation approaches [54], and the integration of habitat restoration and species-targeted conservation may take on many forms [55]. One annoying issue with conservation and restoration programs is how to select the goals and indicators that meaningfully prove success [56]. When projects are funded with public funds, taxpayers need to know that the best available science is employed to define success. The large-scale threatened tree transplant projects on Guam carry a requirement of 50% survival after one year, and this metric of project success has little relevance to long-term restoration success [52]. The inclusion of proof that native edaphic conditions return within the forest enhancement sites may be one of the more relevant metrics to include in restoration and conservation projects that use public funds.
The documented increase in V. parviflora population on Guam in recent years [24] confirms the need to expand the control efforts of this aggressive invader. Early detection of invasive species and development of a rapid response are current thrusts in the fight against invasive species [57]. These early detection mandates indicate Guam ecologists need to watch the nascent evidence that Cedrela odorata L. can displace L. leucocephala (personal observations). This large tree is capable of germinating and establishing beneath mature L. leucocephala stands. A single juvenile stem rapidly penetrates the emergent canopy of L. leucocephala, after which a branched architecture develops above the mature L. leucocephala. The large resulting C. odorata canopy excludes incoming light to kill the subtending L. leucocephala trees.
5. Conclusions
This Guam study adds a tropical island case study to the literature covering the influence of invasive plants on biogeochemistry, and provides an example of how tree invasions of islands can damage forest ecological processes by displacing native plant species then engineering unique changes to the soil ecosystems. The results indicate that L. leucocephala, V. parviflora, and S. campanulata are transformer species, and their below-ground ecosystems become less proficient at storing carbon, nitrogen, and phosphorus. Mitigation activities that include the removal of these trees from restoration sites may require many years for the below-ground ecosystems to return to their native state. Future research should focus on strategies that restore these degraded soils to their native status to better support native vegetation restoration.
Funding
This research was funded in part by United States Department of Agriculture Natural Resources Conservation Service Grant Agreement number 69-3A75-10-148.
Acknowledgments
Support and companionship was provided by Nirmala Dongol for all field work. Management of the mesocosm provided by Nirmala Dongol.
Conflicts of Interest
The author declares no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Liao, C.; Peng, R.; Luo, Y.; Zhou, X.; Wu, X.; Fang, C.; Chen, J.; Li, B. Altered ecosystem carbon and nitrogen cycles by plant invasion: a meta-analysis. New Phytol. 2008, 177, 706–714. [Google Scholar] [CrossRef] [PubMed]
- Pejchar, L.; Mooney, H.A. Invasive species, ecosystem services and human well-being. Trends Ecol. Evol. 2009, 24, 497–504. [Google Scholar] [CrossRef]
- Ehrenfeld, J.G. Ecosystem consequences of biological invasions. Annu. Rev. Ecol. Evol. Syst. 2010, 41, 59–80. [Google Scholar] [CrossRef] [Green Version]
- Pyšek, P.; Richardson, D.M. Invasive species, environmental change and management, and health. Annu. Rev. Environ. Resources 2010, 35, 25–55. [Google Scholar] [CrossRef] [Green Version]
- Elgersma, K.J.; Ehrenfeld, J.G.; Yu, S.; Vor, T. Legacy effects overwhelm the short-term effects of exotic plant invasion and restoration on soil microbial community structure, enzyme activities, and nitrogen cycling. Oecologia 2011, 167, 733–745. [Google Scholar] [CrossRef] [PubMed]
- Kulmatiski, A.; Beard, K.H.; Stevens, J.R.; Cobbold, S.M. Plant-soil feedbacks: a meta-analytical review. Ecol. Lett. 2008, 11, 980–992. [Google Scholar] [CrossRef]
- Sanon, A.; Duponnois, R.; Andrianjaka, Z.N.; Prin, Y.; Bally, R.; Thioulouse, J.; Comte, G. Rhizosphere microbiota interferes with plant–plant interactions. Plant Soil 2009, 321, 259–278. [Google Scholar] [CrossRef]
- Coykendall, K.E.; Houseman, G.R. Lespedeza cuneata invasion alters soils facilitating its own growth. Biol. Invasions 2014, 16, 1735–1742. [Google Scholar] [CrossRef]
- Anning, A.K.; Gyamfi, B.; Effah, A.T. Broussonetia papyrifera controls nutrient return to soil to facilitate its own invasion in a tropical forest of Ghana. J. Plant Ecol. 2018, 11, 909–918. [Google Scholar] [CrossRef]
- Richardson, D.M.; Pyšek, P.; Rejmanek, M.; Barbour, M.G.; Panetta, F.D.; West, C.J. Naturalization and invasion of alien plants: Concepts and definitions. Divers. Distrib. 2000, 6, 93–107. [Google Scholar] [CrossRef]
- Davis, M.; Chew, M.K.; Hobbs, R.J.; Lugo, A.E.; Ewel, J.J.; Vermeij, G.J.; Brown, J.H.; Rosenzweig, M.L.; Gardener, M.R.; Carroll, S.P.; et al. Don’t judge species on their origins. Nature 2011, 474, 153–154. [Google Scholar] [CrossRef]
- Reaser, J.K.; Simpson, A.; Guala, G.F.; Morisette, J.T.; Fuller, P. Envisioning a national invasive species information framework. Biol. Invasions 2020, 22, 21–36. [Google Scholar] [CrossRef] [Green Version]
- Hulme, P.E. Weed risk assessment: A way forward or a waste of time? J. Appl. Ecol. 2012, 49, 10–19. [Google Scholar] [CrossRef]
- Hulme, P.E.; Pyšek, P.; Jarosik, V.; Pergl, J.; Schaffner, U.; Vilá, M. Bias and error in understanding plant invasion impacts. Trends Ecol. Evol. 2013, 28, 212–218. [Google Scholar] [CrossRef]
- Pyšek, P.; Jarošík, V.; Hulme, P.E.; Pergl, J.; Hejda, M.; Schaffner, U.; Vilà, M. A global assessment of invasive plant impacts on resident species, communities and ecosystems: The interaction of impact measures, invading species’ traits and environment. Glob. Change Biol. 2012, 18, 1725–1737. [Google Scholar] [CrossRef]
- Prescott, C.E.; Zukswert, J.M. Invasive plant species and litter decomposition: time to challenge assumptions. New Phytol. 2016, 209, 5–7. [Google Scholar] [CrossRef] [Green Version]
- Ward, D.M.; Nelson, K.A.; Gawel, A.M.; Rogers, H.S. The role of trust in public attitudes toward invasive species management on Guam: A case study. J. Environ. Manag. 2019, 229, 133–144. [Google Scholar]
- Weber, E. Invasive Plant Species of the World: A Reference Guide to Environmental Weeds; CABI: Boston, MA, USA, 2003. [Google Scholar]
- Global Invasive Species Database. Leucaena leucocephala. Invasive Species Specialist Group. Available online: www.iucngisd.org (accessed on 28 February 2020).
- Chou, H.-C.; Kuo, Y.-L. Allelopathic research of subtropical vegetation in Taiwan. III. Allelopathic exclusion of understory by Leucaena leucocephala (Lam.) de Wit. J. Chem. Ecol. 1986, 12, 1431–1448. [Google Scholar] [CrossRef]
- Yoshida, K.; Oka, S. Impact of biological invasion of Leucaena leucocephala on successional pathway and species diversity of secondary forest on Hahajima Island, Ogasawara (Bonin) Islands, northwestern Pacific. Japanese J. Ecol. 2000, 50, 111–119. [Google Scholar]
- Marler, T.E.; Dongol, N.; Cruz, G.N. Leucaena leucocephala and adjacent native limestone forest habitats contrast in soil properties on Tinian Island. Commun. Integr. Biol. 2016, 9, e12127–e12192. [Google Scholar] [CrossRef] [Green Version]
- Global Invasive Species Database. Spathodea campanulata. Invasive Species Specialist Group. Available online: www.iucngisd.org (accessed on 28 February 2020).
- United States Department of Agriculture Forest Service. The Pacific Northwest Forest Inventory and Analysis. Available online: https://www.fs.fed.us/pnw/rma/fia-topics/inventory-data/index.php (accessed on 27 January 2020).
- United States Forest Service. Pacific Island Ecosystems at Risk. Available online: http://www.hear.org/pier/ (accessed on 23 February 2020).
- Marler, T.E.; Moore, A. Military threats to terrestrial resources not restricted to wartime: A case study from Guam. J. Environ. Sci. Eng. 2011, 5, 1198–1214. [Google Scholar]
- Young, F.J. Soil Survey of Territory of Guam; U. S. Dept. of Agric. Soil Conservation Service: Washington, DC, USA, 1988. [Google Scholar]
- Olsen, S.R.; Cole, C.V.; Watanabe, F.S.; Dean, L.A. Estimation of Available Phosphorus in Soils by Extraction with Sodium Bicarbonate; U.S. Dept. of Agric. Circ. No. 939: Washington, DC, USA, 1954. [Google Scholar]
- Berghage, R.D.; Krauskopf, D.M.; Warncke, D.D.; Widders, I. Micronutrient Testing of Plant Growth Media Extractant, Identification and Evaluation. Commun. Soil Sci. Plant Anal. 1987, 18, 1089–1109. [Google Scholar] [CrossRef]
- Zheljazkov, V.D.; Warman, P.R. Comparison of three digestion methods for the recovery of 17 plant essential nutrients and trace elements from six composts. Compost Sci. Utiliz. 2002, 10, 197–203. [Google Scholar] [CrossRef]
- Cataldo, D.A.; Haroon, M.; Schrader, L.E.; Youngs, V.L. Rapid colorimetric determination of nitrate in plant tissue by nitration of salicylic acid. Commun. Soil Sci. Plant Anal. 1975, 6, 71–80. [Google Scholar] [CrossRef]
- Eno, C.F. Nitrate production in the field by incubating the soil in polyethylene bags. Proc. Soil Sci. Soc. Amer. 1960, 24, 277–279. [Google Scholar] [CrossRef]
- Mann, H.B.; Whitney, D.R. On a test of whether one of two random variables is stochastically larger than the other. Ann. Math. Stat. 1947, 18, 50–60. [Google Scholar] [CrossRef]
- Castro-Díez, P.; Godoy, O.; Alonso, A.; Gallardo, A.; Saldaña, A. What explains variation in the impacts of exotic plant invasions on the nitrogen cycle? A meta-analysis. Ecol. Lett. 2014, 17, 1–12. [Google Scholar]
- Sardans, J.; Rivas-Ubach, A.; Peñuelas, J. The C:N:P stoichiometry of organisms and ecosystems in a changing world: A review and perspectives. Perspect. Plant Ecol. Evol. Syst. 2012, 14, 33–47. [Google Scholar] [CrossRef]
- Joint Region Marianas. Final Integrated Natural Resources Management Plan-Joint Region Marianas. Guam, Tinian, and Farallon de Medinilla; Prepared under Contract #SF1449-N40192-10-R-9915; 2012. Available online: sablan.house.gov (accessed on 28 February 2020).
- Joint Guam Program Office. Final Environmental Impact Statement. Guam and CNMI Military Relocation. 2010. Available online: guambuildupeis.us (accessed on 28 February 2020).
- Hickam Air Force Base, Hawaii. Biological Assessment for Beddown of Training and Support Initiatives at Northwest Field, Andersen Air Force Base, Guam; Hickam Air Force Base: Hawaii, HI, USA, 2006. [Google Scholar]
- Andersen Air Force Base. Cycas micronesica Management Plan for Andersen Air Force Base, Guam; Prepared under Contract #N40192-12-P-5008; Andersen Air Force Base: Yigo, GU, USA, 2014. [Google Scholar]
- Naval Facilities Engineering Command Marianas. Haputo Ecological Reserve Area General Management Plan, Naval Base Guam; Naval Facilities Engineering Command Marianas: Piti, GU, USA, 2010. [Google Scholar]
- Naval Facilities Engineering Command Marianas. Restoration Plan for the Habitat Management Unit, Naval Support Activity Andersen, Guam; Prepared under Cooperative Agreement N40192-14-2-8003; 2017. Available online: sablan.jouse.gov (accessed on 28 February 2020).
- WESMURPH Consulting. Off-Site Conservation of Threatened Cycas micronesica, Navy Leased Lands, Tinian; Prepared under Contract #N40192-16-M-5006; WESMURPH Consulting: Santa Rita, GU, USA, 2017. [Google Scholar]
- Simberloff, D.; Von Holle, B. Positive interactions of nonindigenous species: Invasional meltdown? Biol. Invasions 1999, 1, 21–32. [Google Scholar] [CrossRef]
- Ehrenfeld, J.G.; Kourtev, P.; Huang, W. Changes in soil functions following invasions of exotic understory plants in deciduous forests. Ecol. Appl. 2001, 11, 1287–1300. [Google Scholar] [CrossRef]
- Corbin, J.D.; D’Antonio, C.M. Effects of exotic species on soil nitrogen cycling: Implications for restoration. Weed Technol. 2004, 18, 1464–1467. [Google Scholar] [CrossRef]
- Kulmatiski, A.; Beard, K.H. Long-term plant growth legacies overwhelm short-term plant growth effects on soil microbial community structure. Soil Biol. Biochem. 2011, 43, 823–830. [Google Scholar] [CrossRef]
- Suding, K.N.; Harpole, W.S.; Fukami, T.; Kulmatiski, A.; MacDougall, A.S.; Stein, C.; van der Putten, W.H. Consequences of plant–soil feedbacks in invasion. J. Ecol. 2013, 101, 298–308. [Google Scholar] [CrossRef]
- van der Putten, W.H.; Bardgett, R.D.; Bever, J.D.; Bezemer, T.M.; Casper, B.B.; Fukami, T.; Kardol, P.; Klironomos, J.N.; Kulmatiski, A.; Schweitzer, J.A.; et al. Plant-soil feedbacks: The past, the present and future challenges. J. Ecol. 2013, 101, 265–276. [Google Scholar] [CrossRef]
- Gibbons, S.M.; Lekberg, Y.; Mummey, D.L.; Sangwan, N.; Ramsey, P.W.; Gilbert, J.A. Invasive plants rapidly reshape soil properties in a grassland ecosystem. mSystems 2017, 2, e00178–e00216. [Google Scholar] [CrossRef] [Green Version]
- Nsikani, M.M.; van Wilgen, B.W.; Gaertner, M. Barriers to ecosystem restoration presented by soil legacy effects of invasive alien N2-fixing woody species: implications for ecological restoration. Restor. Ecol. 2018, 26, 235–244. [Google Scholar] [CrossRef]
- United States Fish & Wildlife Service. Endangered and threatened wildlife and plants; endangered status for 16 species and threatened status for 7 species in Micronesia. Fed. Regist. 2015, 80, 59424–59497. [Google Scholar]
- Marler, T.E.; Lindström, A.J. First, do no harm. Communic. Integr. Biol. 2017, 10, e1393593. [Google Scholar] [CrossRef]
- Osborne, P.E.; Seddon, P.J. Selecting suitable habitats for reintroductions: variation, change and the role of species distribution modelling. In Reintroduction Biology: Integrating Science and Management; Ewen, J.G., Armstrong, D.P., Parker, K.A., Seddon, P.J., Eds.; Wiley-Blackwell: Oxford, UK, 2012; pp. 73–105. [Google Scholar]
- Prach, K.; Šebelíková, L.; Řehounková, K.; del Moral, R. Possibilities and limitations of passive restoration of heavily disturbed sites. Landsc. Res. 2020, 45, 247–253. [Google Scholar] [CrossRef]
- Volis, S. Conservation-oriented restoration—A two for one method to restore both threatened species and their habitats. Plant Divers. 2019, 41, 50–58. [Google Scholar] [CrossRef]
- Prach, K.; Durigan, G.; Fennessy, S.; Overbeck, G.E.; Torezan, J.M.; Murphy, S.D. A primer on choosing goals and indicators to evaluate ecological restoration success. Restor. Ecol. 2019, 27, 917–923. [Google Scholar] [CrossRef]
- Reaser, J.K.; Burgiel, S.W.; Kirkey, J.; Brantley, K.A.; Veatch, S.D.; Burgos-Rodríguez, J. The early detection of and rapid response (EDRR) to invasive species: A conceptual framework and federal capacities assessment. Biol. Invasions 2020, 22, 1–19. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
The nitrogen properties of Guam cobbly clay soils within invaded mature stands of. Vitex parviflora and in adjacent native forest in central Guam. Means ± SE, n = 6.
Figure 1.
The nitrogen properties of Guam cobbly clay soils within invaded mature stands of. Vitex parviflora and in adjacent native forest in central Guam. Means ± SE, n = 6.

Figure 2.
The nitrogen properties of Guam cobbly clay soils within invaded mature stands of Leucaena leucocephala and in adjacent native forest in central Guam. Means ± SE, n = 6.
Figure 2.
The nitrogen properties of Guam cobbly clay soils within invaded mature stands of Leucaena leucocephala and in adjacent native forest in central Guam. Means ± SE, n = 6.

Figure 3.
The nitrogen properties of coastal sand soils within invaded mature stands of. Leucaena leucocephala and in adjacent native forest in central Guam. Means ± SE, n = 6.
Figure 3.
The nitrogen properties of coastal sand soils within invaded mature stands of. Leucaena leucocephala and in adjacent native forest in central Guam. Means ± SE, n = 6.

Figure 4.
The nitrogen properties of Guam cobbly clay soils within invaded mature stands of. Spathodea campanulata and in adjacent native forest in central Guam. Means ± SE, n = 6.
Figure 4.
The nitrogen properties of Guam cobbly clay soils within invaded mature stands of. Spathodea campanulata and in adjacent native forest in central Guam. Means ± SE, n = 6.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
The chemical properties of Guam cobbly clay soils within invaded mature stands of. Vitex parviflora and in adjacent native forest in central Guam. Means ± SE, n = 6.
Table 1.
The chemical properties of Guam cobbly clay soils within invaded mature stands of. Vitex parviflora and in adjacent native forest in central Guam. Means ± SE, n = 6.
| Soil Property | Invaded | Native | t-test Statistic | Significance |
|---|---|---|---|---|
| pH | 7.72 ± 0.07 | 7.65 ± 0.06 | t = 0.74 | 0.238 |
| Carbon 1 | 120.28 ± 3.44 | 141.68 ± 4.60 | t = 3.36 | 0.004 |
| Phosphorus 2 | 55.45 ± 2.36 | 87.17 ± 5.22 | t = 10.70 | <0.001 |
| Potassium 3 | 32.53 ± 1.39 | 64.56 ± 4.13 | t = 7.35 | <0.001 |
| Calcium 3 | 10.63 ± 0.45 | 11.69 ± 0.50 | t = 1.72 | 0.058 |
| Magnesium 3 | 335.83 ± 14.33 | 381.87 ± 19.85 | t = 1.88 | 0.045 |
| Manganese 3 | 96.84 ± 4.13 | 123.17 ± 5.30 | t = 3.92 | 0.001 |
| Iron 3 | 29.90 ± 1.28 | 52.67 ± 3.19 | t = 6.65 | <0.001 |
| Cobalt 4 | 3.24 ± 0.14 | 9.56 ± 0.41 | t = 14.69 | <0.001 |
| Chromium 4 | 156.49 ± 7.36 | 494.97 ± 21.12 | t = 15.13 | <0.001 |
| Copper 4 | 9.38 ± 0.42 | 25.98 ± 1.11 | t = 14.01 | <0.001 |
| Nickel 4 | 21.03 ± 0.93 | 69.04 ± 2.95 | t = 15.53 | <0.001 |
| Selenium 4 | 0.96 ± 0.05 | 0.63 ± 0.03 | t = 5.70 | <0.001 |
| Zinc 4 | 23.40 ± 0.98 | 23.12 ± 0.99 | t = 0.20 | 0.422 |
1 Dry combustion, mg·g−1. 2 Olsen’s method, µg·g−1. 3 diethylenetriaminepentaacetic acid extractable, mg·g−1 for calcium, µg·g−1 for remainder. 4 nitric acid digestible, µg·g−1.
Table 2.
The chemical properties of Guam cobbly clay soils within invaded mature stands of. Leucaena leucocephala and in adjacent native forest in central Guam. Means ± SE, n = 6.
Table 2.
The chemical properties of Guam cobbly clay soils within invaded mature stands of. Leucaena leucocephala and in adjacent native forest in central Guam. Means ± SE, n = 6.
| Soil Property | Invaded | Native | t-test Statistic | Significance |
|---|---|---|---|---|
| pH | 7.55 ± 0.04 | 7.72 ± 0.05 | t = 2.41 | 0.018 |
| Carbon 1 | 80.83 ± 2.44 | 105.35 ± 4.49 | t = 4.60 | 0.001 |
| Phosphorus 2 | 23.63 ± 1.55 | 39.06 ± 1.67 | t = 6.78 | <0.001 |
| Potassium 3 | 61.74 ± 4.34 | 33.21± 1.42 | t = 6.24 | <0.001 |
| Calcium 3 | 10.09 ± 0.39 | 8.66 ± 0.37 | t = 2.66 | 0.024 |
| Magnesium 3 | 627.17 ± 30.39 | 483.91 ± 15.73 | t = 4.19 | 0.002 |
| Manganese 3 | 26.73 ± 1.97 | 13.86 ± 0.63 | t = 6.23 | <0.001 |
| Iron 3 | 27.67 ± 0.62 | 14.33 ± 0.69 | t = 14.43 | <0.001 |
| Cobalt 4 | 15.91 ± 1.38 | 21.61 ± 3.49 | t = 1.52 | 0.080 |
| Chromium 4 | 178.30 ± 11.52 | 148.32 ± 10.44 | t = 1.93 | 0.043 |
| Copper 4 | 31.21 ± 4.08 | 49.81 ± 11.15 | t = 1.57 | 0.074 |
| Nickel 4 | 69.73 ± 10.71 | 29.54 ± 6.32 | t = 3.23 | 0.005 |
| Selenium 4 | 1.76 ± 0.30 | 0.38 ± 0.10 | t = 4.36 | 0.001 |
| Zinc 4 | 89.18 ± 6.49 | 77.29 ± 5.50 | t = 1.40 | 0.096 |
1 Dry combustion, mg·g−1. 2 Olsen’s method, µg·g−1. 3 diethylenetriaminepentaacetic acid extractable, mg·g−1 for calcium, µg·g−1 for remainder. 4 nitric acid digestible, µg·g−1.
Table 3.
The chemical properties of coastal sand soils within invaded mature stands of. Leucaena leucocephala and in adjacent native forest in central Guam. Means ± SE, n = 6.
Table 3.
The chemical properties of coastal sand soils within invaded mature stands of. Leucaena leucocephala and in adjacent native forest in central Guam. Means ± SE, n = 6.
| Soil Property | Invaded | Native | t-test Statistic | Significance |
|---|---|---|---|---|
| pH | 7.67 ± 0.04 | 7.73 ± 0.03 | t = 1.69 | 0.061 |
| Carbon 1 | 113.9 ± 2.55 | 137.20 ± 2.64 | t = 4.36 | 0.001 |
| Phosphorus 2 | 46.50 ± 1.18 | 60.33 ± 1.52 | t = 7.20 | <0.001 |
| Potassium 3 | 53.83 ± 1.62 | 41.33 ± 1.05 | t = 6.46 | <0.001 |
| Calcium 3 | 6.51 ± 0.71 | 6.90 ± 0.62 | t = 0.42 | 0.343 |
| Magnesium 3 | 400.33 ± 6.85 | 508.67 ± 7.25 | t = 10.86 | <0.001 |
| Manganese 3 | 19.50 ± 0.76 | 25.11 ± 1.13 | t = 3.61 | 0.002 |
| Iron 3 | 9.33 ± 0.42 | 14.83 ± 1.08 | t = 4.75 | 0.004 |
| Cobalt 4 | 0.19 ± 0.01 | 0.25 ± 0.01 | t = 4.12 | 0.001 |
| Chromium 4 | 4.89 ± 0.25 | 5.22 ± 0.22 | t = 0.99 | 0.174 |
| Copper 4 | 0.02 ± 0.01 | 1.96 ± 0.08 | t = 23.25 | <0.001 |
| Nickel 4 | 0.41 ± 0.04 | 1.23 ± 0.07 | t = 10.91 | <0.001 |
| Selenium 4 | 0.51 ± 0.03 | 0.34 ± 0.02 | t = 4.86 | 0.003 |
| Zinc 4 | 10.47 ± 0.53 | 12.03 ± 0.51 | t = 2.12 | 0.030 |
1 Dry combustion, mg·g−1. 2 Olsen’s method, µg·g−1. 3 diethylenetriaminepentaacetic acid extractable, mg·g−1 for calcium, µg·g−1 for remainder. 4 nitric acid digestible, µg·g−1.
Table 4.
The chemical properties of Guam cobbly clay soils within invaded mature stands of. Spathodea campanulata and in adjacent native forest in central Guam. Means ± SE, n = 6.
Table 4.
The chemical properties of Guam cobbly clay soils within invaded mature stands of. Spathodea campanulata and in adjacent native forest in central Guam. Means ± SE, n = 6.
| Soil Property | Invaded | Native | Statistic 1 | Significance |
|---|---|---|---|---|
| pH | 7.52 ± 0.06 | 7.72 ± 0.05 | t = 2.35 | 0.040 |
| Carbon 2 | 64.64 ± 3.27 | 107.17 ± 5.01 | t = 7.11 | <0.001 |
| Phosphorus 3 | 30.86 ± 1.33 | 40.95 ± 1.56 | t = 4.92 | <0.001 |
| Potassium 4 | 34.71 ± 1.80 | 32.71 ± 1.31 | t = 0.90 | 0.391 |
| Calcium 4 | 7.17 ± 0.32 | 8.76 ± 0.34 | t = 3.42 | 0.007 |
| Magnesium 4 | 513.83 ± 21.86 | 488.88 ± 14.23 | t = 0.96 | 0.361 |
| Manganese 4 | 32.88 ± 1.42 | 15.39 ± 0.99 | t = 10.06 | <0.001 |
| Iron 4 | 20.43 ± 0.91 | 15.90 ± 0.86 | t = 3.61 | 0.005 |
| Cobalt 5 | 26.66 ± 2.52 | 21.61 ± 3.49 | t = 1.17 | 0.134 |
| Chromium 5 | 210.73 ± 9.87 | 148.32 ± 10.44 | t = 4.34 | 0.001 |
| Copper 5 | 62.24 ± 4.52 | 49.81 ± 11.15 | t = 1.03 | 0.163 |
| Nickel 5 | 54.78 ± 4.96 | 29.54 ± 6.32 | t = 3.14 | 0.005 |
| Selenium 5 | 0.05 ± 0.301 | 0.38 ± 0.10 | U = 0.5 | 0.007 |
| Zinc 5 | 94.86 ± 3.51 | 77.29 ± 5.50 | t = 2.69 | 0.011 |
1 Mann-Whitney U test, or two-tailed t test. 2 Dry combustion, mg·g−1. 3 Olsen’s method, µg·g−1. 4 diethylenetriaminepentaacetic acid extractable, mg·g−1 for calcium, µg·g−1 for remainder. 5 nitric acid digestible, µg·g−1.
© 2020 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Marler, T.E. Three Invasive Tree Species Change Soil Chemistry in Guam Forests. Forests 2020, 11, 279. https://0-doi-org.brum.beds.ac.uk/10.3390/f11030279
AMA Style
Marler TE. Three Invasive Tree Species Change Soil Chemistry in Guam Forests. Forests. 2020; 11(3):279. https://0-doi-org.brum.beds.ac.uk/10.3390/f11030279
Chicago/Turabian StyleMarler, Thomas E. 2020. "Three Invasive Tree Species Change Soil Chemistry in Guam Forests" Forests 11, no. 3: 279. https://0-doi-org.brum.beds.ac.uk/10.3390/f11030279
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.