Effect of Bark-Stripping on Mechanical Stability of Norway Spruce
by
 , , and
, , and
Oskars Krisans
1, 
Renate Saleniece
1,
Steffen Rust
2,
Didzis Elferts
1 ,
,
Rolands Kapostins
1,
Aris Jansons
1,* and
Roberts Matisons
1 1
Latvian State Forest Research Institute ‘Silava’, Forest Tree Breeding and Adaptation, 111 Rigas street, LV-2169 Salaspils, Latvia
2
University of Applied Sciences and Arts, Faculty of Resource Mananagement, Büsgenweg 1a, 37077 Göttingen, Germany
*
Author to whom correspondence should be addressed.
Forests 2020, 11(3), 357; https://0-doi-org.brum.beds.ac.uk/10.3390/f11030357
Submission received: 28 February 2020
/
Revised: 17 March 2020
/
Accepted: 20 March 2020
/
Published: 21 March 2020
(This article belongs to the Special Issue Natural Disturbance Dynamics Analysis for Forest Ecosystem Management)
Abstract
:The increasing effects of storms are considered the main abiotic disturbance affecting forest ecosystems. Bark-stripping damage from the growing ungulate populations, in turn, are among the main biotic risks, which might burden the stability of trees and stands. Therefore, the aim of our study is to estimate the effect of cervid bark-stripping on the mechanical stability of Norway spruce using a static tree-pulling test. For the test, eight damaged and 11 undamaged canopy trees were selected from a 40-year old stand (plantation with 1 × 3 m spacing) growing on mineral mesotrophic soil. The selected trees were bark-stripped 7–9 years prior to the experiment. Uprooting was the most frequent type of failure; only two trees broke at the stem. For the damaged trees, the resistance to pulling was significantly reduced (p-value < 0.001). Stem volume and presence of bark-stripping were the best linear predictors of the basal bending moment at the primary failure (irreversible deformation of wood structure) and secondary failure (collapse of the tree). A significant (p-value < 0.001) interaction between stem–wood volume and presence of bark-stripping was observed for primary failure, indicating a size-dependent reduction of stability of the damaged trees. Such interaction lacked significance (p-value = 0.43) for the secondary failure (mostly uprooting), indicating a decrease in stability irrespectively of tree size. Somewhat surprisingly, the decrease in the overall mechanical stability of the bark-stripped trees appeared not to be related to a direct reduction of the strength of the stems, but rather to physiological effects such as altered allocation of carbon, increased drought stress because of interfered hydraulic conductance of wood, or secondary infestation. The reduced stability also suggests that bark-stripped trees can act as the weak spots decreasing the collective stability of stands in the long term, thus increasing the susceptibility to storms.
1. Introduction
In European forests, storms are the main abiotic disturbance [1], which have caused more than half of the losses in standing stock during the last 50 years [1,2]. The impact of storms is expected to further increase due to climatic change [3]. In Northern Europe, increasing damage of storms is related to unfrozen soil conditions under warming winters, hence the mechanical stability of individual trees is essential for the sustainability of stands [4]. Furthermore, the impact of storms is intensified by the accompanying legacy effects, such as pest outbreaks and predisposal to pathogens as the trees are weakened [1,2]. Accordingly, storms are projected to further intensify the negative effects of climate changes on Norway spruce (Picea abies L. Karst), which is already associated with lower mechanical stability against wind, particularly in pure stands [5,6,7].
European forests suffer increasing damage caused by the growing cervid populations [8]. Middle-aged (stem exclusion phase; cf. Oliver and Larson [9]) Norway spruce stands are frequently subjected to bark-stripping [10,11], as bark can make more than 10% out of the red deer diet [12]. Bark-stripping damage intensifies with an increasing proportion of spruce in stands and with increasing stand density [13], which subjects pure stands to increased risk [6]. Stem breakage of Norway spruces damaged by moose at the wound height has been observed by Randveer and Heikkilä [14], linking the wildlife damage with the stand stability and susceptibility to wind damage [4]. The interactions between the biotic and abiotic agents can also amplify their individual influence [1]. For example, bark-stripping results in the loss of timber value because of the removal of phloem, while the uncovering of xylem interferes with the physiological processes, thus reducing increment and weakening trees [15], as well as providing a gateway for infections [16].
The effects of storms on stands are shaped by collective and individual stability of trees, which depend on stand density, openings in the canopy, which are the starting points for wind damage, and mechanical resistance of the individuals [2,17,18]. Injuries to the stem can cause a loss of mechanical strength of the tree, resulting in weakened parts in the canopy of a stand, thus decreasing the collective stability [2,18]. Additionally, stripped bark acts as a gateway for pathogens [16,19]; hence, stem rot has been found on 60%–100% of damaged trees [15,16,18]. Rot-infected trees are more prone to stem breakage due to increased susceptibility to wind damage [6], causing negative effects on the stability and growth of a stand. However, the empirical data on the effect of bark-stripping on the wind stability of individual trees, which are needed for a deeper understanding and more precise modelling of the expected effects of wind in the future, are still lacking [1,18,20].
The aim of the study is to evaluate the effect of bark-stripping on the mechanical stability of middle-aged Norway spruce growing in a pure stand. We hypothesise that bark-stripping reduces the mechanical stability of trees, causing stem breakage (snapping) at the wound.
2. Materials and Methods
2.1. Study Site and Sample Trees
A 40-year-old monoculture (planting density 3 × 1 m) of Norway spruce established in lowland conditions (113 m above sea level) on oligotrophic well-drained soil in the central part of Latvia (56°42′ N; 25°53′ E) was studied. Such selection was made because dense Norway spruce stands are common in the eastern Baltics. The studied stand is located in a national forest research station, hence information on bark-stripping and sanitary conditions were available.
The area of the stand was 0.89 ha, and it was exposed to a clear-cut on the south and southwest sides, and to mature Norway spruce stands on the other sides. No management has been done except weed control after the establishment. The climate at the site location is temperate. The 30-year mean annual precipitation is 600 mm and the mean monthly temperature ranges from −7 to 16.9 °C in January and July, respectively. Westerlies are the dominant winds with a mean annual velocity of 3 ms−1. The strongest winds mostly occur during winter. Five storms stronger than 22 ms−1 have occurred over the last 18 years [21].
In the studied stand, eight canopy trees with stripped bark on the stems and 11 undamaged trees, representing the diameter distribution of the plantation, were selected for the destructive static pulling tests. The selected trees appeared visually healthy (apart from bark-stripping wounds). Additional criteria for the selection of the damaged trees were the exposure of the wounds to the north and northeast direction. Trees on the edges of the stand were avoided. The undamaged trees had slightly larger stem diameter at breast height (DBH), height and stem volume (Table 1). More details on each tree are shown in the supplementary material (Table S1). Bark-stripping wounds on the damaged tree stems were at a height of 80–150 cm above the ground. The age of the wounds, as determined from stem discs collected after the pulling test, was 7–9 years. The shapes of the bark-stripping wounds were transferred to transparent film, and, later in the laboratory, measured using a planimeter (Planix 10S; Tamaya, Japan). The area of the damage ranged from 603 to 2375 cm2. Bark-stripped trees did not have any other visual damage. Presence of wood rot was evaluated on stem discs.
2.2. Static Pulling Tests
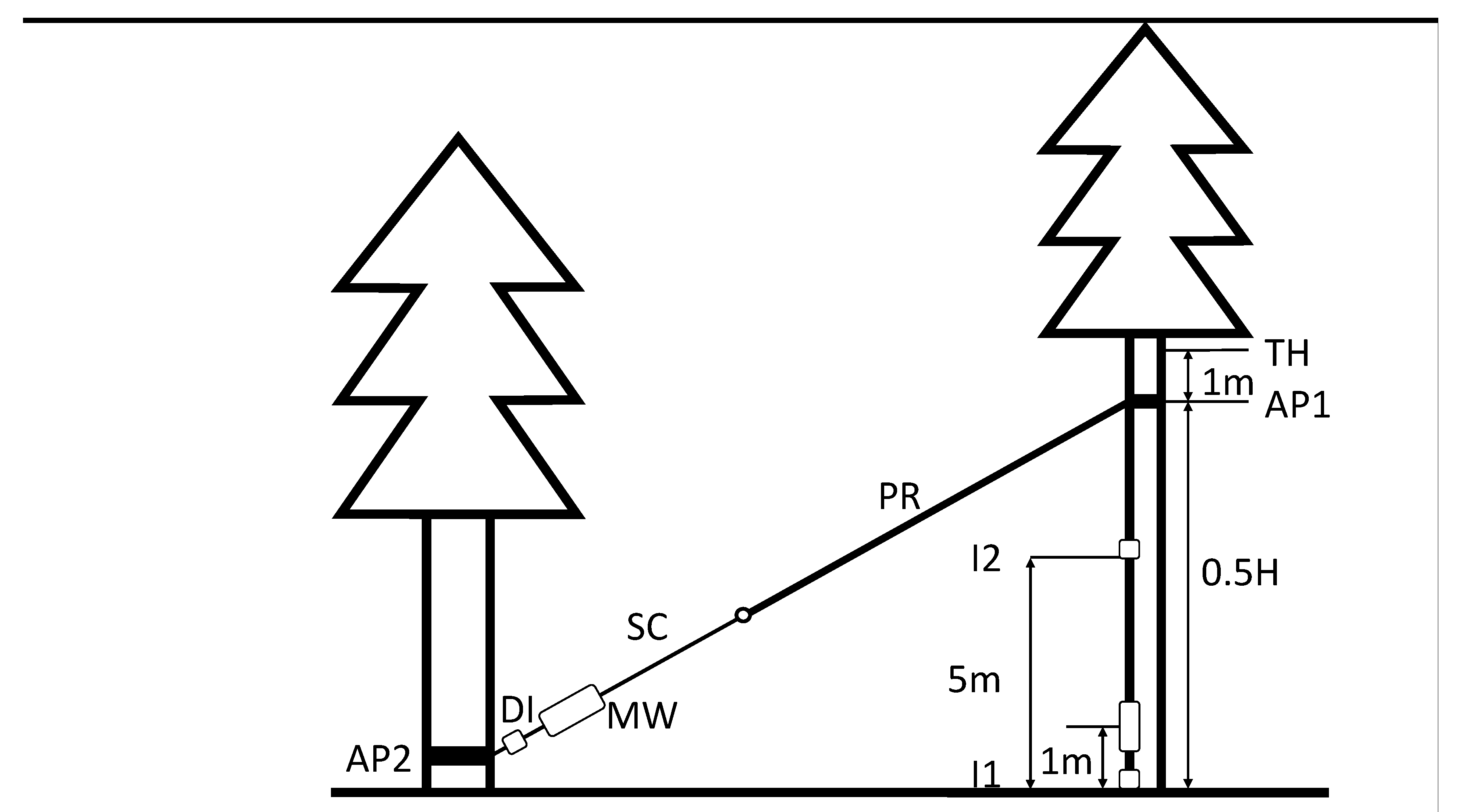
Destructive static pulling tests [22] were used to assess the effect of bark-stripping on the stability of Norway spruce. The pulling line was anchored at 50% of the height of each sample tree (Figure 1). Before the pulling, trees were topped 1 m above the anchorage point to prevent the influence of wind and canopy weight on the measurements.
The trees were pulled using a manual winch (working load limit 32 kN) and steel cable (diameter 16 mm) anchored at the ground level. The pulling cable was extended with a static polyester rope (Tenex Tec 16; Samson Rope Technologies Inc., Ferndale, WA, USA). Damaged trees were pulled in the direction of the wound. Measurements were done with the TreeQinetic System (Argus electronic GmbH, Rostock, Germany). A dynamometer, recording pulling force and rope angle was placed between the winch and the rope (Figure 1). The inclination of the root plate and the curvature of the stem were measured with two inclinometers at the base and at the height of 5 m.
2.3. Data Processing and Analysis
For each tree, the bending moment at the base of the stem (BBM, in kNm), was calculated as follows:
where F is the pulling force, hanchor is the height of the anchor point of the sample tree, and medianα rope is the median of the rope angle. The difference between two simultaneous inclinometer measurements (N5m at the height of 5 m; Nbase at the base, 0 m) was used to characterise the curvature of the stem NΔ:
BBM = F × hanchor × cos (medianα rope),
NΔ = N5m − Nbase
The primary failure of the stem occurs when the structure of wood starts to change due to stress caused by the external force. At that point, the wood fibers start to buckle, while the damage might not be visually detectable. The limit of proportionality between NΔ and BBM was considered as the point when the primary failure occurs [23]. This point was determined by graphical inspection. The secondary failure was considered to occur when the maximum BBM was reached, after which tree collapse (either uprooted or its stem broke) followed.
Linear models were used to assess the main tree-level variables affecting primary and secondary failure. The full model was fit to describe both individual and interacting effects of the studied factors on the BBM at primary and secondary failure (Table 2). As the study had a limited scope, a restricted number of factors were analyzed together according to the arbitrary selection principle. The tested predictors were tree dimensions (DBH, height, and stem volume), presence and size of bark-stripping, and presence of wood rot. Pulling direction (cosine) was tested as a proxy for the effects of dominant winds. Model performance was assessed using diagnostic plots. If models had similar performance, they were compared using ANOVA; if no statistical difference between the models was estimated, the simplest was used. Stem–wood volume was calculated using local functions by Liepa [24]. The data were processed in R Software (version 3.5.3) using packages: “readr”, “tidyverse”, “DBI”, “zoo”, “RSQLite”, “ggplot2” [25].
3. Results
Uprooting was the most frequent type of failure, as only two of the 19 trees studied had stem breakage (one bark-stripped and one undamaged tree). However, the limited sample size of the fractured trees prevented statistical analysis of factors influencing failure type. Both broken trees had similar DBH and height. The breakage on the bark-stripped tree occurred a few meters above the bark-stripping wound. Wood decay was not observed on the stem disc at the wound. The undamaged broken tree showed notably higher BBM at the primary and secondary failure compared to the uprooted ones; therefore, it was excluded from further analysis as an outlier.
The best performing models describing primary and secondary failure contained the same two predictors: stem–wood volume and presence of a bark-stripping wound, which were strictly significant (p-value < 0.001; Table 2). The interaction between the predictors, however, was significant (p-value < 0.001) only in case of primary failure. Nevertheless, both models showed a good fit (R2 ≥ 0.95), and their residuals were independent and followed the normal distribution. The BBM at primary and secondary failure showed a clear linear dependency on the stem–wood volume (Figure 2). The presence of bark-stripping wounds on stems significantly (Table 2) reduced the BBM necessary for tree failure to occur (Figure 2). The effect of bark-stripping was particularly pronounced for the occurrence of primary failure. With the increasing size of the trees, the effect of bark-stripping became larger. On average, the damaged trees required 61% less load to fail (Figure 2A). The presence of bark-stripping wounds had a tree-size-independent effect on the resistance of trees to secondary failure, reducing the BBM by ca. 16 kNm per m3, indicating higher susceptibility of the smaller trees.
4. Discussion
Beyond the general rule that bark-stripping enables fungal infection [18] and wood decay reduces the mechanical strength of trees [26], little is known about the quantitative effects of ungulate bark-stripping on the stability of individual trees [27,28]. Because this type of damage is widespread in commercial forests, models for storm risk assessment might benefit from more comprehensive information.
To quantify the effect of bark-stripping on tree stability, damaged and undamaged Norway spruce trees were pulled to failure. Contrary to expectations, most trees with wounds did not snap but were uprooted, which is the most frequent wind damage for shallow-rooted species such as the Norway spruce [2]. However, the presence of bark-stripping reduced bending moments at primary and secondary failures significantly (Figure 2; Table 2), confirming our initial hypothesis at least in part. The critical bending moments were comparable to the results found for Norway spruce in other parts of Europe [4,29,30].
Bark-stripping wounds reduced not only stem strength but also anchorage. Bark-stripping disrupts conductive tissues of trees, causing physiological stress [18], and triggers the allocation of resources to healing [15]. Both effects reduce growth in the long term. Under such conditions, we speculate that trees invest less carbon into the root system, thus reducing anchorage and facilitating uprooting [28,31]. Bark-stripping also acts as a gateway for pathogens [16,18,32], which can reduce the growth and vigour of trees, reducing the mechanical stability of trees [28]. One of the most common Norway spruce pathogens, Heterobasidion annosum, has been related to increased uprooting and stem breakage [28,33]. Although wood rot was not detected on the stem discs at the wound, the spread of pathogens in the stem can differ by species [16,34]; hence, fungal infection still might have had affected roots of the bark-stripped trees, although not visible higher in the stem.
The marked increase in susceptibility of the bark-stripped trees to primary failure (Figure 2A), which disrupts conductivity of wood facilitating physiological drought and weakening of trees [23], suggested that the studied damage by wildlife might enhance the negative legacy effects of storms, such as increased susceptibility to pests [1,2]. The legacy effects of bark-stripping are decreased mechanical stability of trees and increased susceptibility of stands to wind damage [18]. Weaker resistance of the bark-stripped trees (Figure 2) implies that they reach primary failure at considerably slower wind speeds, facilitating storm legacy effects, which in turn are increasing susceptibility to wind damage, thus resulting in a negative feedback loop and drawing negative perspectives to Norway spruce stands in the eastern Baltic region. In addition, the primary failure creates irreversible damage to the wood fibers (even if not visually detectable) [23], reducing the quality and value of timber [28,35]. Furthermore, the negative effect of bark-stripping on tree stability (Figure 2; Table 2) suggests that damaged trees, as the triggering points, might reduce the collective stability of stands to wind loads [2,17,18]. The growing frequency of stand-replacing storms [2,3,36] in combination with intensifying cervid damage [10,20] are expected to cause increasing calamities to Norway spruce in Northern Europe.
5. Conclusions
Under the growing influence of storms, bark-stripping damages caused by growing cervid populations will intensify the direct and legacy effects of storms on Norway spruce. Considering the long-term and short-term effects and risks allocated to bark-stripping on trees and stand, the damaged trees are to be harvested soon after the damage, while the legacy effects have not caused more extensive damage, although the harvesting of the damaged trees should be carried out when the risk of storms is the lowest, allowing stands to restore collective stability. In this sense, prevention of wildlife damage could notably improve the sustainability of Norway spruce stands against wind damage.
Supplementary Materials
The following are available online at https://0-www-mdpi-com.brum.beds.ac.uk/1999-4907/11/3/357/s1.
Author Contributions
Conceptualization, O.K. and A.J.; methodology, O.K., R.M., S.R.; formal analysis, D.E., R.M., data curation, R.K., R.M.; writing—original draft preparation, R.S., O.K.; writing—review and editing, R.M., S.R.; project administration, A.J. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by ERDF grant number 1.1.1.1/16/A/260.
Acknowledgments
This study was funded by the European Regional Development Fund project development of decision support tool for prognosis of storm damages in forest stands on peat soils (No. 1.1.1.1/16/A/260).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Seidl, R.; Rammer, W. Climate change amplifies the interactions between wind and bark beetle disturbances in forest landscapes. Landsc. Ecol. 2017, 32, 1485–1498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gardiner, B.; Schuck, A.R.T.; Schelhaas, M.J.; Orazio, C.; Blennow, K.; Nicoll, B. Living with Storm Damage to Forests; European Forest Institute: Joensuu, Finland, 2013; pp. 1–132. [Google Scholar]
- Seidl, R.; Schelhaas, M.J.; Rammer, W.; Verkerk, P.J. Increasing forest disturbances in Europe and their impact on carbon storage. Nat. Clim. Chang. 2014, 4, 806–810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peltola, H.; Ikonen, V.P.; Gregow, H.; Strandman, H.; Kilpeläinen, A.; Venäläinen, A.; Kellomäki, S. Impacts of climate change on timber production and regional risks of wind-induced damage to forests in Finland. For. Ecol. Manag. 2010, 260, 833–845. [Google Scholar] [CrossRef]
- Albrecht, A.; Hanewinkel, M.; Bauhus, J.; Kohnle, U. How does silviculture affect storm damage in forests of south-western Germany? Results from empirical modeling based on long-term observations. Eur. J. For. Res. 2012, 131, 229–247. [Google Scholar] [CrossRef]
- Richter, C. Wood Characteristics: Description, Causes, Prevention, Impact on Use and Technological Adaptation; Springer Internationale Publishing: Basel, Switzerland, 2015; p. 222. [Google Scholar]
- Lodin, I. Choice of Tree Species in the Aftermath of Two Major Storms—A Qualitative Study of Private Forest Owners in Southern Sweden. Master’s Thesis, Swedish University of Agricultural Sciences, Uppsala, Sweden, 2016. [Google Scholar]
- Kurek, T.; Jacek, T.; Pazdrowski, W.; Szymański, M.; Łukowski, A. Intensity of stripping and sugar content in the bark and the bast of European beech (Fagus sylvatica). Open Life Sci. 2019, 14, 19–28. [Google Scholar] [CrossRef]
- Oliver, C.D.; Larson, B.C. Forest Stand Dynamics; John Wiley & Sons: New York, NY, USA, 1996; p. 520. [Google Scholar]
- Apollonio, M.; Andersen, R.; Putman, R. European Ungulates and Their Management in the 21st Century, 1st ed.; Cambridge University Press: Cambridge, UK, 2010; p. 618. [Google Scholar]
- Månsson, J.; Jarnemo, A. Bark-stripping on Norway spruce by red deer in Sweden: Level of damage and relation to tree characteristics. Scan. J. For. Res. 2013, 28, 117–125. [Google Scholar] [CrossRef]
- Verheyden, H.; Ballon, P.; Bernard, V.; Saint-Andrieux, C. Variations in bark-stripping by red deer Cervus elaphus across Europe. Mamm. Rev. 2006, 36, 217–234. [Google Scholar] [CrossRef]
- Vospernik, S. Probability of bark stripping damage by red deer (Cervus elaphus) in Austria. Silva Fenn. 2006, 40, 589–601. [Google Scholar] [CrossRef] [Green Version]
- Randveer, T.; Heikkilä, R. Damage cause by moose (Alces alces L.) by bark stripping of Picea abies. Scan. J. For. Res. 1996, 11, 153–158. [Google Scholar] [CrossRef]
- Vasiliauskas, R. Damage to trees due to forestry operations and its pathological significance in temperate forests: A literature review. Forestry 2001, 74, 319–336. [Google Scholar] [CrossRef] [Green Version]
- Burneviča, N.; Jansons, Ā.; Zaļuma, A.; Kļaviņa, D.; Jansons, J.; Gaitnieks, T. Fungi Inhabiting Bark Stripping Wounds Made by Large Game on Stems of Picea abies (L.) Karst. in Latvia. Balt. For. 2016, 22, 2–7. [Google Scholar]
- Díaz-Yáñez, O.; Mola-Yudego, B.; González-Olabarria, J.R.; Pukkala, T. How does forest composition and structure affect the stability against wind and snow? For. Ecol. Manag. 2017, 401, 215–222. [Google Scholar] [CrossRef]
- Cukor, J.; Vacek, Z.; Linda, R.; Vacek, S.; Marada, P.; Šimůnek, V.; Havránek, F. Effects of bark stripping on timber production and structure of Norway spruce forests in relation to climatic factors. Forests 2019, 10, 320. [Google Scholar] [CrossRef] [Green Version]
- McLaughlin, J.; Šica, L. Aļņu radītie mizas plēsumi parastai eglei un ar tiem saistītās trupes izraisītāju infekcijas. Mežzinātne 1996, 39, 26–37. [Google Scholar]
- Vlad, R.; Sidor, C. Assessment of the wood volume with stem decay in Norway spruce stands damaged by red deer. In Proceedings of the Biennial International Symposium Forest and Sustainable Development, Brasov, Romania, 24–25 October 2014; Transylvania University Press: Brasov, Romania, 2015; pp. 381–386. [Google Scholar]
- Harris, I.; Jones, P.; Osborn, T.; Lister, D. Updated high-resolution grids of monthly climatic observations—The CRU TS3.10 Dataset. Int. J. Climatol. 2014, 34, 623–642. [Google Scholar] [CrossRef] [Green Version]
- Peltola, H.M. Mechanical stability of trees under static loads. Am. J. Bot. 2006, 93, 1501–1511. [Google Scholar] [CrossRef]
- Detter, A.; Richter, K.; Rust, C.; Rust, S. Aktuelle Untersuchungen zum Primärversagen von grünem Holz—Current studies on primary failure in green wood. In Proceedings of the Conference Deutsche Baumpflegetage, Augsburg, Germany, 5–7 May 2015; pp. 156–167. [Google Scholar]
- Liepa, I. Pieauguma Mācība; [Increment Theory]; Latvia University of Agriculture: Jelgava, Latvia, 1996; p. 123. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018; Available online: https://www.R-project.org/ (accessed on 24 November 2019).
- Wagener, W.W. Judging Hazard from Native Trees in California Recreational Areas: A Guide for Professional Foresters; USFS Research Paper PSW-P1; CFS: Berkeley, CA, USA, 1963; p. 29. [Google Scholar]
- Shibata, E.; Torazawa, Y. Effects of bark stripping by sika deer, Cervus nippon, on wind damage to coniferous trees in subalpine forest of central Japan. J. For. Res. 2008, 13, 296–301. [Google Scholar] [CrossRef]
- Honkaniemi, J.; Lehtonen, M.; Väisänen, H.; Peltola, H. Effects of wood decay by Heterobasidion annosum on the vulnerability of Norway spruce stands to wind damage: A mechanistic modelling approach. Can. J. For. Res. 2017, 47, 777–787. [Google Scholar] [CrossRef]
- Lundström, T.; Jonas, T.; Stöckli, S.; Ammann, W. Anchorage of mature conifers: Resistive turning moment, root–soil plate geometry and root growth orientation. Tree Physiol. 2007, 27, 1217–1227. [Google Scholar] [CrossRef] [Green Version]
- Jillich, S.; Köhler, J.; Rust, C.; Rust, S.; Detter, A. Brechen schlanke Fichten leichter? In Jahrbuch der Baumpflege; Dujesiefken, D., Ed.; Haymarket Media: Braunschweig, Germany, 2013; pp. 267–273. [Google Scholar]
- Szoradova, A.; Praus, L.; Kolarik, J. Evaluation of the root system resistance against failure of urban trees using principal component analysis. Biosyst. Eng. 2013, 115, 244–249. [Google Scholar] [CrossRef]
- Vasiliauskas, R. Five basidiomycetes in living stems of Picea abies, associated with 7–25 year-old wounds. Balt. For. 1998, 1, 29–35. [Google Scholar]
- Deflorio, G.; Johnson, C.R.; Fink, S.; Schwarze, F.W.M.R. Decay development in living sapwood of coniferous and deciduous trees inoculated with six wood decay fungi. For. Ecol. Manag. 2008, 255, 2373–2383. [Google Scholar] [CrossRef]
- Schmidt, O. Wood and Tree Fungi; Springer: Berlin/Heidelberg, Germany, 2006; p. 334. [Google Scholar]
- Herrmann, S.; Huber, M.O.; Bont, Z.; Rigling, A.; Wunder, J. Decay detection in Norway spruce (Picea abies) with the Rotfinder instrument. For. Ecol. Manag. 2019, 448, 549–558. [Google Scholar] [CrossRef]
- Schelhaas, M.J. Impacts of natural disturbances on the development of European forest resources: Application of model approaches from tree and stand levels to large-scale scenarios. Alterra Sci. Contrib. 2008, 23, 168. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Schematic depiction of the static destructive pulling test setup. AP: anchorage point, DI: dynamometer (includes inclinometer for rope angle), I: inclinometer, MW: manual winch, TH: topping height; SC: steel cable; PR: polyester rope.
Figure 1.
Schematic depiction of the static destructive pulling test setup. AP: anchorage point, DI: dynamometer (includes inclinometer for rope angle), I: inclinometer, MW: manual winch, TH: topping height; SC: steel cable; PR: polyester rope.

Figure 2.
Bending moment of the Norway spruce stem at the primary failure (A) and secondary failure (B) according to stem–wood volume and presence of bark-stripping wound.
Figure 2.
Bending moment of the Norway spruce stem at the primary failure (A) and secondary failure (B) according to stem–wood volume and presence of bark-stripping wound.


{kind=link}
{kind=link}
Table 1.
Statistics of the studied dataset. Mean values ± 95% confidence intervals are shown.
| Undamaged | Bark-Stripped | |
|---|---|---|
| Number of trees | 11 | 8 |
| Stem circumference (cm) | 86.0 ± 13.7 | 82.2 ± 17.3 |
| Damage circumference (cm) | - | 22.5 ± 6.2 |
| Damage proportion (%) | - | 28.1 ± 7.3 |
| DBH (cm) | 27.3 ± 4.3 | 25.8 ± 5.1 |
| Tree height (m) | 24.3 ± 2.6 | 23.1 ± 3.3 |
| Stem volume (m3) | 0.76 ± 0.24 | 0.64 ± 0.27 |
| Pulling direction (cosine) | 0.33 ± 0.46 | 0.83 ± 0.23 |
Table 2.
Regression between tree stability parameters and stem volume and cervid damage for the damaged trees.
Table 2.
Regression between tree stability parameters and stem volume and cervid damage for the damaged trees.
| Parameter | F-Value | p-Value |
|---|---|---|
| Primary failure | ||
| Stem volume | 268.9 | <0.001 |
| Presence of bark-stripping wound | 159.3 | <0.001 |
| Stem volume by presence of bark-stripping interaction | 28.9 | <0.001 |
| R2 | 0.96 | |
| Model overall significance, p-value | <0.001 | |
| Secondary failure | ||
| Stem volume | 294.5 | <0.001 |
| Presence of bark-stripping wound | 38.0 | <0.001 |
| Stem volume by presence of bark-stripping interaction | 0.6441 | 0.43 |
| R2 | 0.95 | |
| Model overall significance, p-value | <0.001 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Krisans, O.; Saleniece, R.; Rust, S.; Elferts, D.; Kapostins, R.; Jansons, A.; Matisons, R. Effect of Bark-Stripping on Mechanical Stability of Norway Spruce. Forests 2020, 11, 357. https://0-doi-org.brum.beds.ac.uk/10.3390/f11030357
AMA Style
Krisans O, Saleniece R, Rust S, Elferts D, Kapostins R, Jansons A, Matisons R. Effect of Bark-Stripping on Mechanical Stability of Norway Spruce. Forests. 2020; 11(3):357. https://0-doi-org.brum.beds.ac.uk/10.3390/f11030357
Chicago/Turabian StyleKrisans, Oskars, Renate Saleniece, Steffen Rust, Didzis Elferts, Rolands Kapostins, Aris Jansons, and Roberts Matisons. 2020. "Effect of Bark-Stripping on Mechanical Stability of Norway Spruce" Forests 11, no. 3: 357. https://0-doi-org.brum.beds.ac.uk/10.3390/f11030357
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.