Climatic, Edaphic and Biotic Controls over Soil δ13C and δ15N in Temperate Grasslands

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Sites
2.2. Analysis of Soil δ13C and δ15N
2.3. Analysis of Soil Properties and Microbial Biomass
2.4. Calculations and Statistics
3. Results
3.1. Climatic, Edaphic and Biotic Factors along the Transect
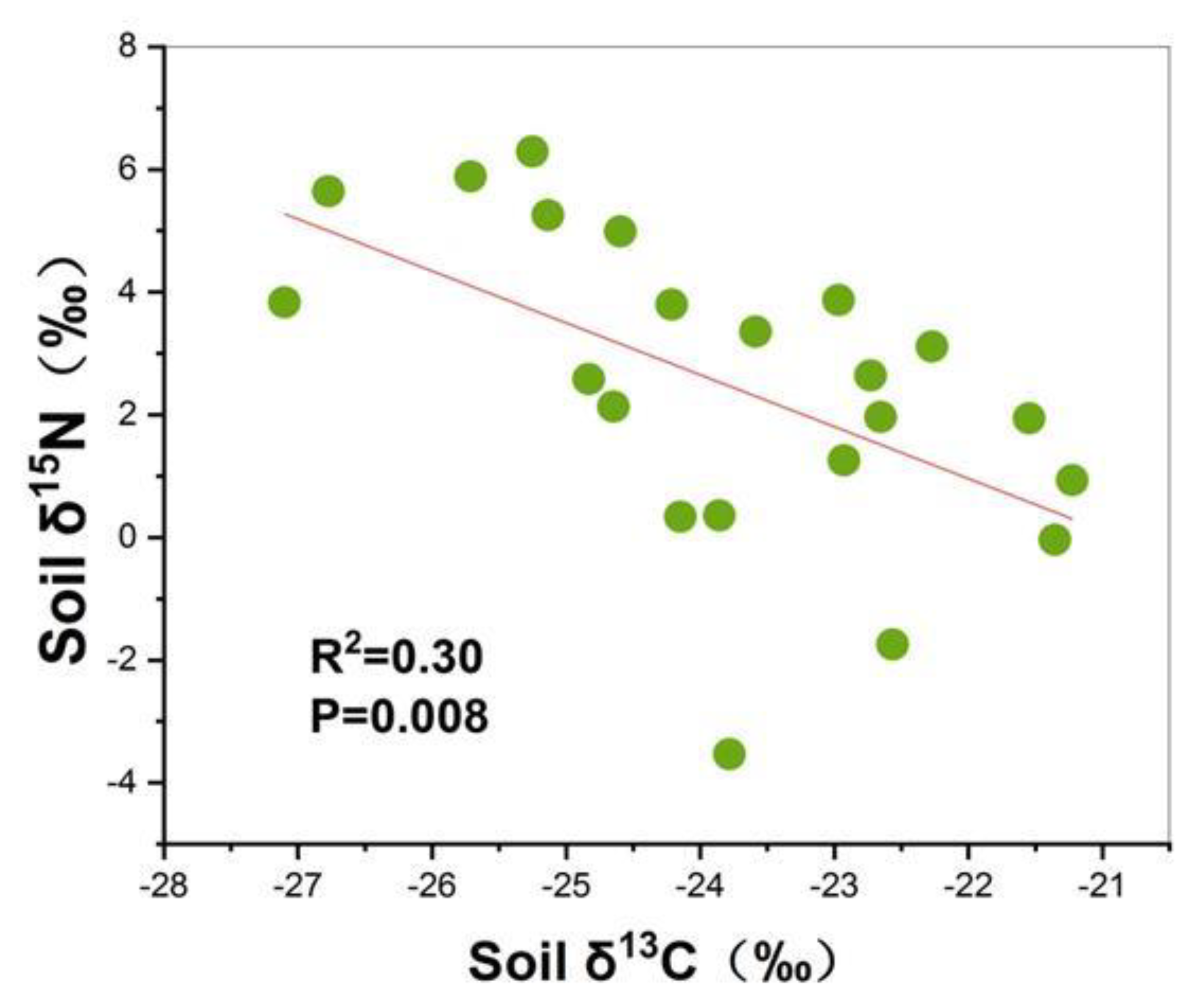
3.2. Soil δ15N and δ13C
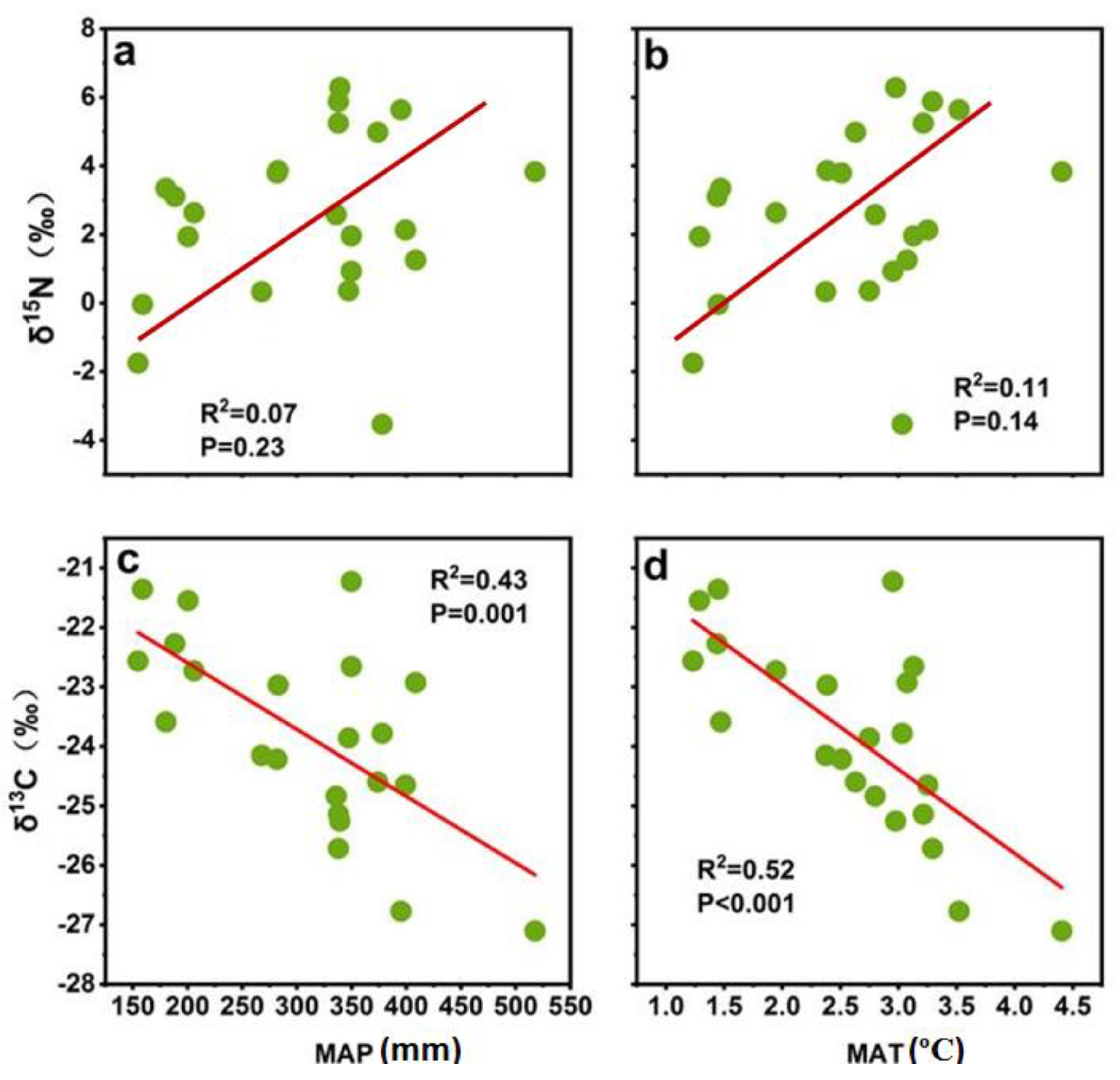
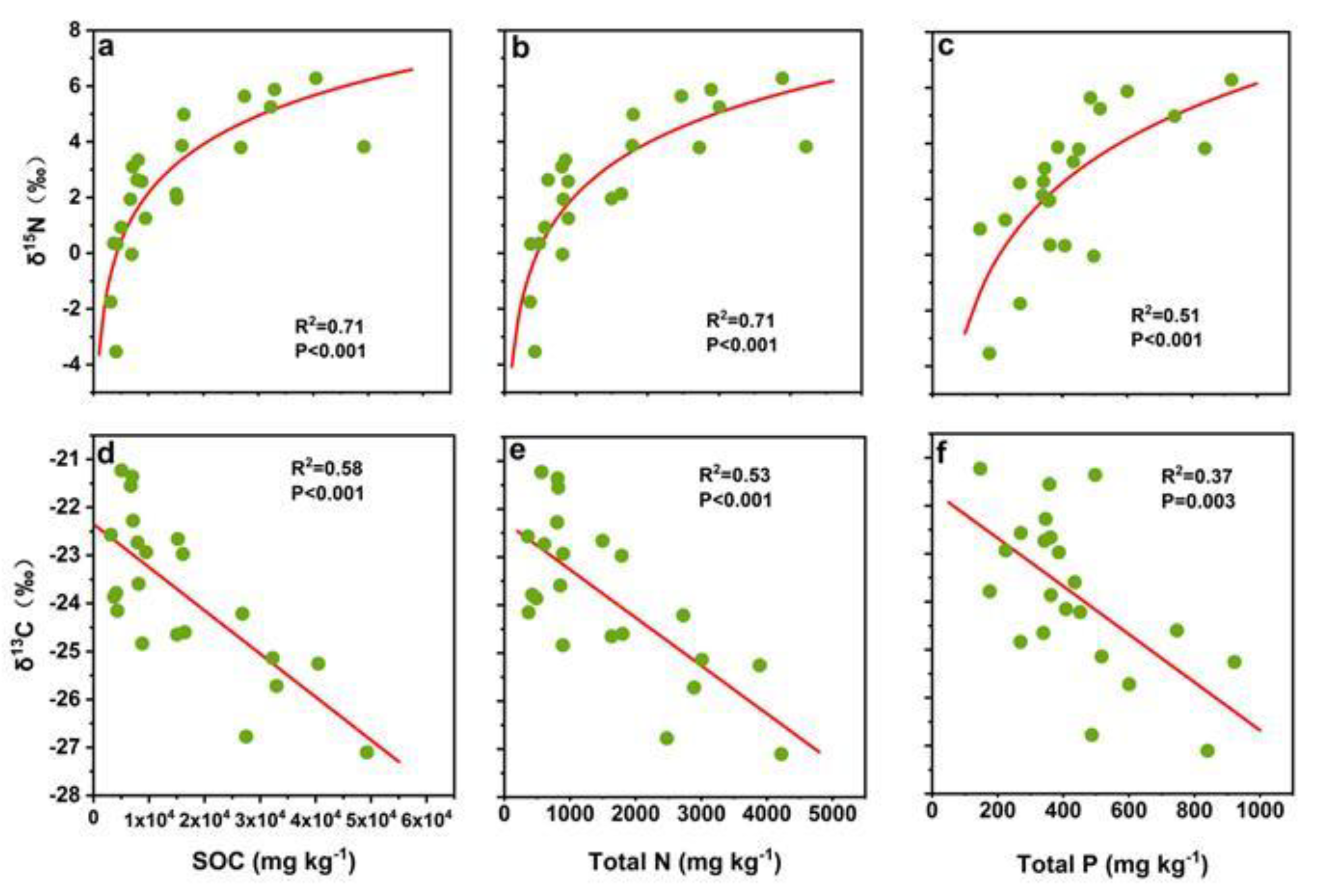
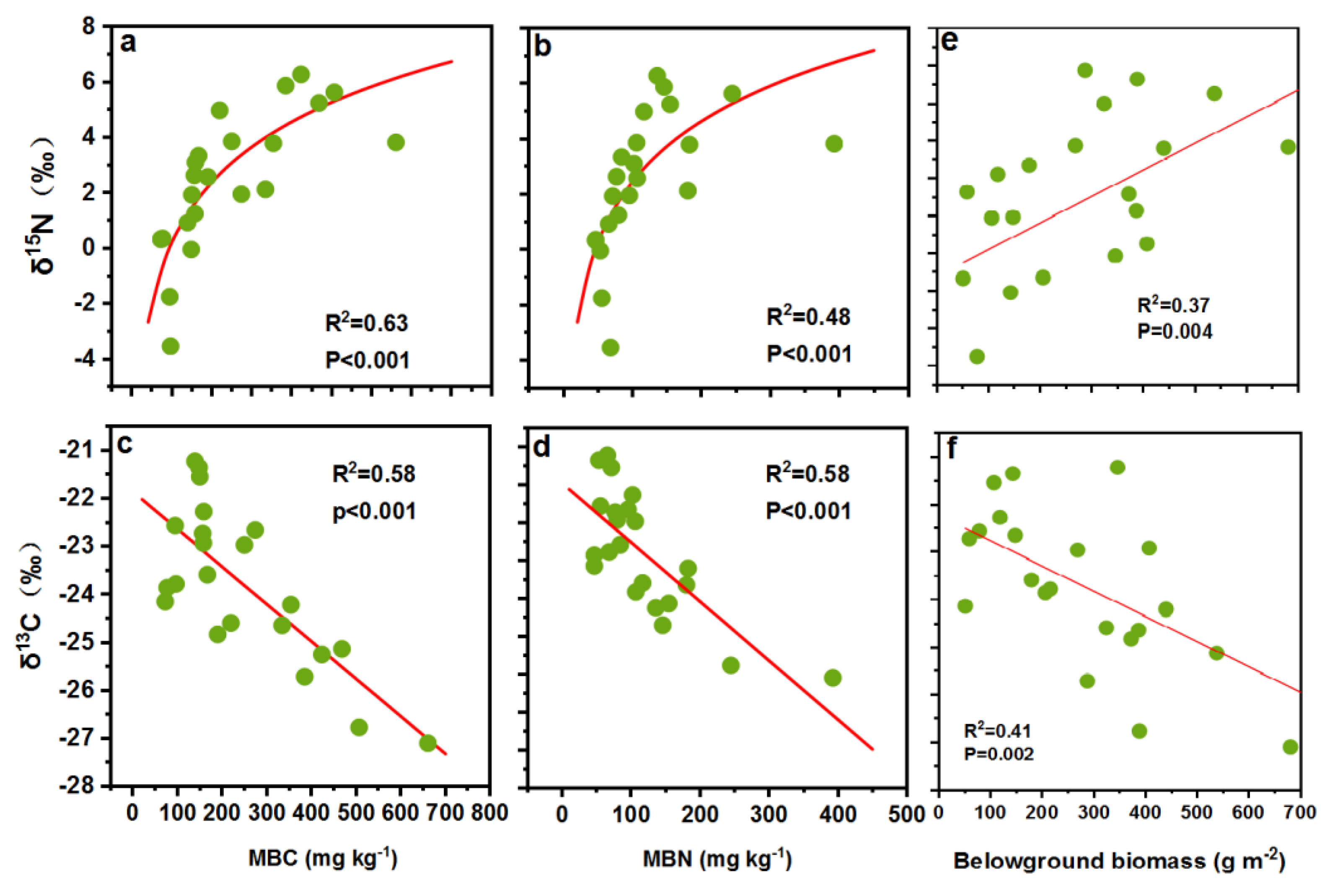
3.3. Correlation Climatic, Edaphic and Biotic Factors with Soil δ15N and δ13C
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Miltner, A.; Bombach, P.; Schmidt-Bruecken, B.; Kaestner, M. SOM genesis: Microbial biomass as a significant source. Biogeochemistry 2012, 111, 41–55. [Google Scholar] [CrossRef]
- Schnitzer, M. A lifetime perspective on the chemistry of soil organic matter. Adv. Agron. 1999, 68, 1–58. [Google Scholar] [CrossRef]
- Batjes, N.H. Total carbon and nitrogen in the soils of the world. Eur. J. Soil Sci. 1996, 47, 151–163. [Google Scholar] [CrossRef]
- Post, W.M.; Pastor, J.; Zinke, P.J.; Stangenberger, A.G. Global patterns of soil nitrogen storage. Nature 1985, 317, 613–616. [Google Scholar] [CrossRef]
- Janzen, H.H.; Campbell, C.A.; Ellert, B.H.; Bremer, E. Soil organic matter dynamics and their relationship to soil quality. Dev. Soil Sci. 1997, 25, 277–291. [Google Scholar] [CrossRef]
- Schmidt, M.W.I.; Torn, M.S.; Abiven, S.; Dittmar, T.; Guggenberger, G.; Janssens, I.A.; Kleber, M.; Kögel-Knabner, I.; Lehmann, J.; Manning, D.A.C.; et al. Persistence of soil organic matter as an ecosystem property. Nature 2011, 478, 49–56. [Google Scholar] [CrossRef] [Green Version]
- Goh, K.M. Carbon sequestration and stabilization in soils: Implications for soil productivity and climate change. Soil Sci. Plant Nutr. 2004, 50, 467–476. [Google Scholar] [CrossRef] [Green Version]
- Lal, R. Managing soils and ecosystems for mitigating anthropogenic carbon emissions and advancing global food Security. Bioscience 2010, 60, 708–721. [Google Scholar] [CrossRef]
- Amundson, R.; Austin, A.T.; Schuur, E.A.G.; Yoo, K.; Matzek, V.; Kendall, C.; Uebersax, A.; Brenner, D.L.; Baisden, W.T. Global patterns of the isotopic composition of soil and plant nitrogen. Glob. Biogeochem. Cycles 2003, 17, 1031. [Google Scholar] [CrossRef]
- Cheng, W.X.; Chen, Q.S.; Xu, Y.Q.; Han, X.G.; Li, L.H. Climate and ecosystem 15N natural abundance along a transect of Inner Mongolian grasslands: Contrasting regional patterns and global patterns. Glob. Biogeochem. Cycles 2009, 23, GB2005. [Google Scholar] [CrossRef]
- Craine, J.M.; Elmore, A.J.; Wang, L.; Augusto, L.; Baisden, W.T.; Brookshire, E.N.J.; Cramer, M.D.; Hasselquist, N.J.; Hobbie, E.A.; Kahmen, A.; et al. Convergence of soil nitrogen isotopes across global climate gradients. Sci. Rep. 2015, 5, 8280. [Google Scholar] [CrossRef] [Green Version]
- Ehleringer, J.R.; Buchmann, N.; Flanagan, L.B. Carbon isotope ratios in belowground carbon cycle processes. Ecol. Appl. 2000, 10, 412–422. [Google Scholar] [CrossRef]
- Garten, C.T.; Cooper, L.W.; Post, W.M.; Hanson, P.J. Climate controls on forest soil C isotope ratios in the southern appalachian mountains. Ecology 2000, 81, 1108–1119. [Google Scholar] [CrossRef]
- Wang, C.; Houlton, B.Z.; Liu, D.; Hou, J.; Cheng, W.; Bai, E. Stable isotopic constraints on global soil organic carbon turnover. Biogeosciences 2018, 15, 987–995. [Google Scholar] [CrossRef] [Green Version]
- Stable Isotopes in Ecology and Environmental Science, 2nd ed.; Michener, R.H.; Lajtha, K. (Eds.) Blackwell Publishing Ltd.: Oxford, UK, 2007; ISBN 978-1-4051-2680-9. [Google Scholar]
- Bernoux, M.; Cerri, C.C.; Neill, C.; de Moraes, J.F.L. The use of stable carbon isotopes for estimating soil organic matter turnover rates. Geoderma 1998, 82, 43–58. [Google Scholar] [CrossRef]
- Conen, F.; Zimmermann, M.; Leifeld, J.; Seth, B.; Alewell, C. Relative stability of soil carbon revealed by shifts in δ15N and C:N ratio. Biogeosciences 2008, 5, 123–128. [Google Scholar] [CrossRef] [Green Version]
- Natelhoffer, K.J.; Fry, B. Controls on natural 15N and 13C abundances in forest soil organic matter. Soil Sci. Soc. Am. J. 1988, 52, 1633–1640. [Google Scholar] [CrossRef]
- Bai, E.; Boutton, T.W.; Liu, F.; Wu, X.B.; Archer, S.R. Spatial patterns of soil δ13C reveal grassland-to-woodland successional processes. Org. Geochem. 2012, 42, 1512–1518. [Google Scholar] [CrossRef]
- Balesdent, J.; Mariotti, A. Measurement of soil organic matter turnover using 13C natural abundance. In Mass Spectrometry of Soil; Boutton, T.W., Yamasaki, S.I., Eds.; Marcel Dekker, Inc.: New York, NY, USA, 1996; pp. 83–111. ISBN 0-8247-9699-3. [Google Scholar]
- Kramer, M.G.; Sollins, P.; Sletten, R.S.; Swart, P.K. N isotope fractionation and measures of organic matter alteration during decomposition. Ecology 2003, 84, 2021–2025. [Google Scholar] [CrossRef] [Green Version]
- Biedenbender, S.H.; McClaran, M.P.; Quade, J.; Weltz, M.A. Landscape patterns of vegetation change indicated by soil carbon isotope composition. Geoderma 2004, 119, 69–83. [Google Scholar] [CrossRef]
- Mayor, J.R.; Wright, S.J.; Schuur, E.A.; Brooks, M.E.; Turner, B.L. Stable nitrogen isotope patterns of trees and soils altered by long-term nitrogen and phosphorus addition to a lowland tropical rainforest. Biogeochemistry 2014, 119, 293–306. [Google Scholar] [CrossRef] [Green Version]
- Stevenson, B.A.; Parfitt, R.L.; Schipper, L.A.; Baisden, W.T.; Mudge, P. Relationship between soil δ15N, C/N and N losses across land uses in New Zealand. Agric. Ecosyst. Environ. 2010, 139, 736–741. [Google Scholar] [CrossRef]
- Bai, E.; Boutton, T.W.; Liu, F.; Ben Wu, X.; Hallmark, C.T.; Archer, S.R. Spatial variation of soil δ13C and its relation to carbon input and soil texture in a subtropical lowland woodland. Soil Biol. Biochem. 2012, 44, 102–112. [Google Scholar] [CrossRef]
- Wynn, J.G.; Bird, M.I. Environmental controls on the stable carbon isotopic composition of soil organic carbon: Implications for modelling the distribution of C3 and C4 plants, Australia. Tellus 2008, 60, 604–621. [Google Scholar] [CrossRef] [Green Version]
- Craine, J.M.; Brookshire, E.N.J.; Cramer, M.D.; Hasselquist, N.J.; Koba, K.; Marin-Spiotta, E.; Wang, L. Ecological interpretations of nitrogen isotope ratios of terrestrial plants and soils. Plant Soil 2015, 396, 1–26. [Google Scholar] [CrossRef] [Green Version]
- Dawson, T.E.; Mambelli, S.; Plamboeck, A.H.; Templer, P.H.; Tu, K.P. Stable isotopes in plant ecology. Annu. Rev. Ecol. Syst. 2002, 33, 507–559. [Google Scholar] [CrossRef]
- Högberg, P. 15N natural abundance in soil-plant systems. New Phytol. 1997, 137, 179–203. [Google Scholar] [CrossRef]
- Makarov, M.I. The nitrogen isotopic composition in soils and plants: Its use in environmental studies (a review). Eurasian Soil Sci. 2009, 42, 1335–1347. [Google Scholar] [CrossRef]
- Pardo, L.H.; Nadelhoffer, K.J. Using nitrogen isotope ratios to assess terrestrial ecosystems at regional and global scales. In Isoscapes: Understanding Movement, Pattern, and Process on Earth through Isotope Mapping; West, J.B., Bowen, G.J., Dawson, T.E., Tu, K.P., Eds.; Springer: New York, NY, USA, 2010; pp. 221–249. ISBN 978-90-481-3353-6. [Google Scholar]
- Robinson, D. δ15N as an integrator of the nitrogen cycle. Trends Ecol. Evol. 2001, 16, 153–162. [Google Scholar] [CrossRef]
- Powers, J.S.; Veldkamp, E. Regional variation in soil carbon and δ13C in forests and pastures of northeastern Costa Rica. Biogeochemistry 2005, 72, 315–336. [Google Scholar] [CrossRef]
- Wang, C.; Wang, X.; Liu, D.; Wu, H.; Lu, X.; Fang, Y.; Cheng, W.; Luo, W.; Jiang, P.; Shi, J.; et al. Aridity threshold in controlling ecosystem nitrogen cycling in arid and semi-arid grasslands. Nat. Commun. 2014, 5, 4799. [Google Scholar] [CrossRef] [Green Version]
- Handley, L.L.; Raven, J.A. The use of natural abundance of nitrogen isotopes in plant physiology and ecology. Plant Cell Environ. 1992, 15, 965–985. [Google Scholar] [CrossRef]
- Aranibar, J.N.; Otter, L.; Macko, S.A.; Feral, C.J.W.; Epstein, H.E.; Dowty, P.R.; Eckardt, F.; Shugart, H.H.; Swap, R.J. Nitrogen cycling in the soil-plant system along a precipitation gradient in the Kalahari sands. Glob. Chang. Biol. 2004, 10, 359–373. [Google Scholar] [CrossRef]
- Austin, A.T.; Vitousek, P.M. Nutrient dynamics on a precipitation gradient in Hawai’i. Oecologia 1998, 113, 519–529. [Google Scholar] [CrossRef]
- Handley, L.L.; Austin, A.T.; Robinson, D.; Scrimgeour, C.M.; Raven, J.A.; Heaton, T.H.E.; Schmidt, S.; Stewart, G.R. The 15N natural abundance (δ15N) of ecosystem samples reflects measures of water availability. Aust. J. Plant Physiol. 1999, 26, 185–199. [Google Scholar] [CrossRef]
- Qiao, N.; Xu, X.; Cao, G.; Ouyang, H.; Kuzyakov, Y. Land use change decreases soil carbon stocks in Tibetan grasslands. Plant Soil 2015, 395, 231–241. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; D’Odorico, P.; Ries, L.; Macko, S.A. Patterns and implications of plant-soil δ13C and δ15N values in African savanna ecosystems. Quat. Res. 2010, 73, 77–83. [Google Scholar] [CrossRef]
- Conant, R.T.; Cerri, C.E.P.; Osborne, B.B.; Paustian, K. Grassland management impacts on soil carbon stocks: A new synthesis. Ecol. Appl. 2017, 27, 662–668. [Google Scholar] [CrossRef] [Green Version]
- Ramankutty, N.; Evan, A.T.; Monfreda, C.; Foley, J.A. Farming the planet: 1. Geographic distribution of global agricultural lands in the year 2000. Glob. Biogeochem. Cycles 2008, 22, GB1003. [Google Scholar] [CrossRef]
- Foley, J.A.; Ramankutty, N.; Brauman, K.A.; Cassidy, E.S.; Gerber, J.S.; Johnston, M.; Mueller, N.D.; O’Connell, C.; Ray, D.K.; West, P.C.; et al. Solutions for a cultivated planet. Nature 2011, 478, 337–342. [Google Scholar] [CrossRef] [Green Version]
- Climate Change 2014: Synthesis Report. Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Core Writing Team, Pachauri, R.K., Meyer, L.A., Eds.; IPCC: Geneva, Switzerland, 2014; ISBN 978-92-9169-143-2. [Google Scholar]
- Bredenkamp, G.J.; Spada, F.; Kazmierczak, E. On the origin of northern and southern hemisphere grasslands. Plant Ecol. 2002, 163, 209–229. [Google Scholar] [CrossRef]
- Farquhar, G.D.; Richards, R.A. Isotopic composition of plant carbon correlates with water use efficiency of wheat genotypes. Aust. J. Plant Physiol. 1984, 11, 539–552. [Google Scholar] [CrossRef]
- Soil Survey Staff. Keys to Soil Taxonomy, 9th ed.; USDA-Natural Resources Conservation Service: Washington, DC, USA, 2003.
- Grimshaw, H.M. The determination of total phosphorus in soils by acid digestion. In Chemical Analysis in Environmental Research; Rowland, A.P., Ed.; NERC/ITE: Abbots Ripton, UK, 1987; pp. 92–95. ISBN 0-904282-98-8. [Google Scholar]
- Parkinson, J.A.; Allen, S.E. A wet oxidation procedure suitable for the determination of nitrogen and mineral nutrients in biological material. Commun. Soil Sci. Plant Anal. 1975, 6, 1–11. [Google Scholar] [CrossRef]
- Brookes, P.C.; Landman, A.; Pruden, G.; Jenkinson, D.S. Chloroform fumigation and the release of soil nitrogen: A rapid direct extraction method to measure microbial biomass nitrogen in soil. Soil Biol. Biochem. 1985, 17, 837–842. [Google Scholar] [CrossRef]
- Vance, E.D.; Brookes, P.C.; Jenkinson, D.S. An extraction method for measuring soil microbial biomass C. Soil Biol. Biochem. 1987, 19, 703–707. [Google Scholar] [CrossRef]
- Wu, J.S.; Joergensen, R.G.; Pommerening, B.; Chaussod, R.; Brookes, P.C. Measurement of soil microbial biomass C by fumigation extraction—An automated procedure. Soil Biol. Biochem. 1990, 22, 1167–1169. [Google Scholar] [CrossRef]
- Burnham, K.P.; Anderson, D.R.; Huyvaert, K.P. AIC model selection and multimodel inference in behavioral ecology: Some background, observations, and comparisons. Behav. Ecol. Sociobiol. 2011, 65, 23–35. [Google Scholar] [CrossRef]
- Cheng, S.L.; Fang, H.J.; Yu, G.R.; Zhu, T.H.; Zheng, J.J. Foliar and soil 15N natural abundances provide field evidence on nitrogen dynamics in temperate and boreal forest ecosystems. Plant Soil 2010, 337, 285–297. [Google Scholar] [CrossRef]
- Xu, Y.; He, J.; Cheng, W.; Xing, X.; Li, L. Natural 15N abundance in soils and plants in relation to N cycling in a rangeland in Inner Mongolia. J. Plant Ecol. 2010, 3, 201–207. [Google Scholar] [CrossRef]
- Kahmen, A.; Wanek, W.; Buchmann, N. Foliar δ15N values characterize soil N cycling and reflect nitrate or ammonium preference of plants along a temperate grassland gradient. Oecologia 2008, 156, 861–870. [Google Scholar] [CrossRef] [Green Version]
- Weintraub, S.R.; Cole, R.J.; Schmitt, C.G.; All, J.D. Climatic controls on the isotopic composition and availability of soil nitrogen across mountainous tropical forest. Ecosphere 2016, 7, e01412. [Google Scholar] [CrossRef]
- Shan, Y.; Huang, M.; Suo, L.; Zhao, X.; Wu, L. Composition and variation of soil δ15N stable isotope in natural ecosystems. Catena 2019, 183, 104236. [Google Scholar] [CrossRef]
- Nissenbaum, A.; Schallinger, K.M. The distribution of stable carbon isotope (13C/12C) in fractions of soil organic matter. Geoderma 1974, 11, 137–145. [Google Scholar] [CrossRef]
- Vankessel, C.; Farrell, R.E.; Pennock, D.J. 13C and 15N natural abundance in crop residues and soil organic matter. Soil Sci. Soc. Am. J. 1994, 58, 382–389. [Google Scholar] [CrossRef]
- Santruckova, H.; Bird, M.I.; Lloyd, J. Microbial processes and carbon-isotope fractionation in tropical and temperate grassland soils. Funct. Ecol. 2000, 14, 108–114. [Google Scholar] [CrossRef]
- Du, B.; Liu, C.; Kang, H.; Zhu, P.; Yin, S.; Shen, G.; Hou, J.; Ilvesniemi, H. Climatic control on plant and soil delta C-13 along an altitudinal transect of Lushan mountain in subtropical China: Characteristics and interpretation of soil carbon dynamics. PLoS ONE 2014, 9, e86440. [Google Scholar] [CrossRef] [Green Version]
- Guo, Q.; Hu, Z.; Li, S.; Li, X.; Sun, X.; Yu, G. Spatial variations in aboveground net primary productivity along a climate gradient in Eurasian temperate grassland: Effects of mean annual precipitation and its seasonal distribution. Glob. Chang. Biol. 2012, 18, 3624–3631. [Google Scholar] [CrossRef]
- Jiao, C.; Yu, G.; He, N.; Ma, A.; Ge, J.; Hu, Z. Spatial pattern of grassland aboveground biomass and its environmental controls in the Eurasian steppe. J. Geogr. Sci. 2017, 27, 3–22. [Google Scholar] [CrossRef]
- Bai, X.; Cheng, J.; Zheng, S.; Zhan, S.; Bai, Y. Ecophysiological responses of Leymus chinensis to nitrogen and phosphorus additions in a typical steppe. Chin. J. Plant Ecol. 2014, 38, 103–115. [Google Scholar] [CrossRef]
- Dong, C.; Wang, W.; Liu, H.; Xu, X.; Zeng, H. Temperate grassland shifted from nitrogen to phosphorus limitation induced by degradation and nitrogen deposition: Evidence from soil extracellular enzyme stoichiometry. Ecol. Indic. 2019, 101, 453–464. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | type | latitude | longitude | Altitude (m) | MAP (mm) | MAT (°C) | g kg−1 | mg kg−1 | Biomass (g m−2) | ‰ | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| SOC | Total N | Total P | MBC | MBN | soil pH | Aboveground | Belowground | δ13C | δ15N | |||||||
| 1 | Temperate meadow steppe | 50°12′ | 119°43′ | 550 | 339.1 | 3.0 | 36.10 ± 0.79 | 3.36 ± 0.08 | 0.9 | 422.9 ± 89.0 | 135.7 ± 29.7 | 6.9 | 26.9 ± 3.4 | 785.6 ± 64.8 | -25.3 ± 0.1 | 6.3 ± 0.6 |
| 2 | Temperate meadow steppe | 49°12′ | 120°22′ | 590 | 394.9 | 3.5 | 31.30 ± 1.16 | 2.82 ± 0.13 | 0.5 | 506.2 ± 56.8 | 244.5 ± 115.9 | 7.3 | 55.7 ± 15.5 | 388.2 ± 20.2 | -26.8 ± 0.1 | 5.6 ± 0.2 |
| 3 | Temperate meadow steppe | 49°17′ | 119°56′ | 590 | 337.7 | 3.2 | 29.77 ± 0.12 | 2.76 ± 0.11 | 0.5 | 467.8 ± 41.7 | 154.4 ± 21.1 | — | 37.4 ± 22.9 | 537.7 ± 92.9 | -25.1 ± 0.2 | 5.3 ± 0.4 |
| 4 | Temperate meadow steppe | 49°9′ | 119°50′ | 500 | 337.7 | 3.3 | 34.64 ± 0.89 | 2.96 ± 0.09 | 0.6 | 384.3 ± 52.9 | 145.7 ± 20.5 | 7.1 | 42.6 ± 52.0 | 287.3 ± 97.1 | -25.7 ± 0.0 | 5.9 ± 0.2 |
| 5 | Temperate meadow steppe | 49°33′″ | 117°19′ | 683 | 281.7 | 2.5 | 17.84 ± 1.55 | 1.88 ± 0.15 | 0.5 | 353.4 ± 47.7 | 182.5 ± 51.5 | 8.0 | 146.6 ± 42.0 | 439.6 ± 117.8 | -24.2 ± 0.3 | 3.8 ± 0.3 |
| 6 | Temperate meadow steppe | 48°22′ | 122°17′ | 450 | 517.5 | 4.4 | 53.36 ± 0.66 | 4.69 ± 0.10 | 0.8 | 660.7 ± 50.2 | 392.4 ± 63.7 | 7.2 | 66.7 ± 14.0 | 680.7 ± 170.6 | -27.1 ± 0.1 | 3.8 ± 0.6 |
| 7 | Temperate steppe | 43°96′ | 115°86′ | 1000 | 282.6 | 2.4 | 12.60 ± 0.32 | 1.37 ± 0.03 | 0.4 | 249.4 ± 57.3 | 105.8 ± 14.3 | 7.5 | 125.9 ± 63.7 | 268.4 ± 183.3 | -23.0 ± 0.4 | 3.9 ± 0.6 |
| 8 | Temperate steppe | 43°52′ | 119°22′ | 644 | 349.5 | 3.1 | 19.57 ± 0.99 | 1.90 ± 0.11 | 0.4 | 273.4 ± 19.6 | 95.4 ± 14.0 | 7.3 | 20.0 ± 0.9 | 147.6 ± 72.0 | -22.7 ± 0.4 | 2.0 ± 0.5 |
| 9 | Temperate steppe | 43°15′ | 118°09′ | 749 | 377.8 | 3.0 | 2.48 ± 0.45 | 0.27 ± 0.04 | 0.2 | 95.9 ± 63.0 | 68.1 ± 50.4 | 7.3 | 70.0 ± 16.0 | 215.3 ± 168.3 | -23.8 ± 0.6 | -3.5 ± 3.7 |
| 10 | Temperate steppe | 43°15′ | 117°11′ | 1296 | 399.1 | 3.3 | 18.06 ± 1.30 | 1.85 ± 0.11 | 0.3 | 333.7 ± 55.2 | 180.1 ± 21.7 | 7.2 | 62.2 ± 12.5 | 386.5 ± 71.6 | -24.6 ± 0.6 | 2.1 ± 0.7 |
| 11 | Temperate steppe | 43°43′ | 112°50′ | 993 | 158.8 | 1.5 | 3.21 ± 0.19 | 0.36 ± 0.03 | 0.5 | 147.7 ± 10.5 | 65.4 ± 16.4 | 8.1 | 25.4 ± 4.0 | 143.0 ± 132.0 | -21.4 ± 0.7 | 0.0 ± 2.1 |
| 12 | Temperate steppe | 43°12′ | 116°09′ | 1298 | 335.7 | 2.8 | 7.64 ± 0.54 | 0.72 ± 0.03 | 0.3 | 189.4 ± 13.6 | 79.5 ± 10.6 | 7.8 | 31.8 ± 5.1 | 372.0 ± 62.2 | -24.8 ± 0.6 | 2.6 ± 1.3 |
| 13 | Temperate steppe | 43°19′ | 119°35′ | 453 | 349.5 | 3.0 | 6.12 ± 0.09 | 0.73 ± 0.03 | 0.1 | 138.5 ± 35.0 | 116.5 ± 24.8 | 8.1 | 11.0 ± 2.3 | 345.8 ± 196.8 | -21.2 ± 0.3 | 0.9 ± 0.9 |
| 14 | Temperate steppe | 42°32′ | 118°53′ | 794 | 408.3 | 3.1 | 9.23 ± 0.91 | 1.02 ± 0.09 | 0.2 | 157.1 ± 40.1 | 53.2 ± 12.0 | 7.9 | 77.2 ± 46.3 | 407.1 ± 206.4 | -22.9 ± 0.6 | 1.3 ± 0.7 |
| 15 | Temperate steppe | 41°20′ | 112°51′ | 1760 | 373.7 | 2.6 | 10.21 ± 0.50 | 1.07 ± 0.04 | 0.7 | 218.9 ± 54.7 | 106.7 ± 20.1 | 8.1 | 35.5 ± 9.1 | 324.1 ± 38.5 | -24.6 ± 0.3 | 5.0 ± 1.4 |
| 16 | Temperate desert | 43°21′ | 111°52′ | 960 | 154.5 | 1.2 | 1.78 ± 0.12 | 0.20 ± 0.02 | 0.3 | 94.0 ± 26.3 | 55.4 ± 26.7 | 8.1 | 7.8 ± 3.5 | 78.0 ± 37.9 | -22.6 ± 0.7 | -1.7 ± 2.4 |
| 17 | Temperate desert | 42°56′ | 110°50′ | 1071 | 200.3 | 1.3 | 4.03 ± 0.40 | 0.55 ± 0.05 | 0.4 | 149.4 ± 22.5 | 71.2 ± 13.5 | 8.1 | 6.5 ± 5.7 | 106.4 ± 95.5 | -21.5 ± 0.8 | 1.9 ± 1.8 |
| 18 | Temperate desert | 42°25′ | 109°49′ | 1158 | 188.2 | 1.4 | 3.95 ± 0.58 | 0.56 ± 0.07 | 0.3 | 158.1 ± 10.8 | 102.0 ± 52.3 | 8.3 | 19.6 ± 8.8 | 118.1 ± 64.8 | -22.3 ± 0.4 | 3.1 ± 1.1 |
| 19 | Temperate desert | 41°54′ | 108°42′ | 1533 | 180.1 | 1.5 | 5.65 ± 0.24 | 0.67 ± 0.04 | 0.4 | 166.2 ± 15.4 | 84.1 ± 13.4 | 8.0 | 11.6 ± 3.2 | 179.0 ± 187.0 | -23.6 ± 0.3 | 3.4 ± 1.5 |
| 20 | Temperate desert | 40°01′ | 110°03′ | 1339 | 347.1 | 2.8 | 2.88 ± 0.10 | 0.26 ± 0.02 | 0.4 | 76.2 ± 13.8 | 46.5 ± 16.0 | 8.3 | 31.5 ± 9.2 | 205.8 ± 92.0 | -23.9 ± 0.6 | 0.4 ± 1.7 |
| 21 | Temperate desert | 39°14′ | 107°16′ | 1281 | 205.8 | 2.0 | 5.62 ± 0.55 | 0.51 ± 0.04 | 0.3 | 155.7 ± 14.3 | 76.8 ± 18.2 | 8.1 | 22.6 ± 5.4 | 58.5 ± 27.3 | -22.7 ± 0.2 | 2.6 ± 0.9 |
| 22 | Temperate desert | 38°44′ | 107°45′ | 1345 | 267.6 | 2.4 | 1.97 ± 0.16 | 0.17 ± 0.02 | 0.4 | 71.8 ± 22.6 | 46.5 ± 24.8 | 8.7 | 87.4 ± 85.1 | 50.9 ± 44.9 | -24.1 ± 0.6 | 0.3 ± 3.4 |
| MAP | MAT | SOC | Total N | Total P | pH | MBC | MBN | BB | |
|---|---|---|---|---|---|---|---|---|---|
| MAP | 1.000 | ||||||||
| MAT | 0.959 * | 1.000 | |||||||
| SOC | 0.629 * | 0.742 * | 1.000 | ||||||
| Total N | 0.627 * | 0.732 * | 0.996 * | 1.000 | |||||
| Total P | 0.273 | 0.320 | 0.721 * | 0.716 * | 1.000 | ||||
| pH | −0.584 * | −0.676 * | −0.780 * | −0.788 | −0.402 | 1.000 | |||
| MBC | 0.580 * | 0.688 * | 0.966 * | 0.973 * | 0.677 * | −0.742 * | 1.000 | ||
| MBN | 0.626 * | 0.675 * | 0.867 * | 0.870 * | 0.575 * | −0.579 * | 0.919 * | 1.000 | |
| BB | 0.677 * | 0.697 * | 0.781 * | 0.802 * | 0.602 * | −0.670 * | 0.771 * | 0.677 * | 1.000 |
| MBC | Total N | MAP | Total P | SOC | MAT | |
|---|---|---|---|---|---|---|
| Soil 15N | 0.52 (0.012) | 0.21 (1.398) | 0.20 (−0.007) | 0.16 (8.559) | 0.12 (0.119) | |
| Soil 13C | 0.29 (−0.006) | 0.29 (−0.005) | 0.71 (−3.420) | 0.71 (−1.152) | ||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, X.; Xu, X.; Wang, F.; Greenberg, I.; Liu, M.; Che, R.; Zhang, L.; Cui, X. Climatic, Edaphic and Biotic Controls over Soil δ13C and δ15N in Temperate Grasslands. Forests 2020, 11, 433. https://0-doi-org.brum.beds.ac.uk/10.3390/f11040433
Zhao X, Xu X, Wang F, Greenberg I, Liu M, Che R, Zhang L, Cui X. Climatic, Edaphic and Biotic Controls over Soil δ13C and δ15N in Temperate Grasslands. Forests. 2020; 11(4):433. https://0-doi-org.brum.beds.ac.uk/10.3390/f11040433
Chicago/Turabian StyleZhao, Xing, Xingliang Xu, Fang Wang, Isabel Greenberg, Min Liu, Rongxiao Che, Li Zhang, and Xiaoyong Cui. 2020. "Climatic, Edaphic and Biotic Controls over Soil δ13C and δ15N in Temperate Grasslands" Forests 11, no. 4: 433. https://0-doi-org.brum.beds.ac.uk/10.3390/f11040433