The Pattern of Genetic Variation, Survival and Growth in the Abies alba Mill. Population within the Introgression Zone of Two Refugial Lineages in the Carpathians


Abstract
:1. Introduction
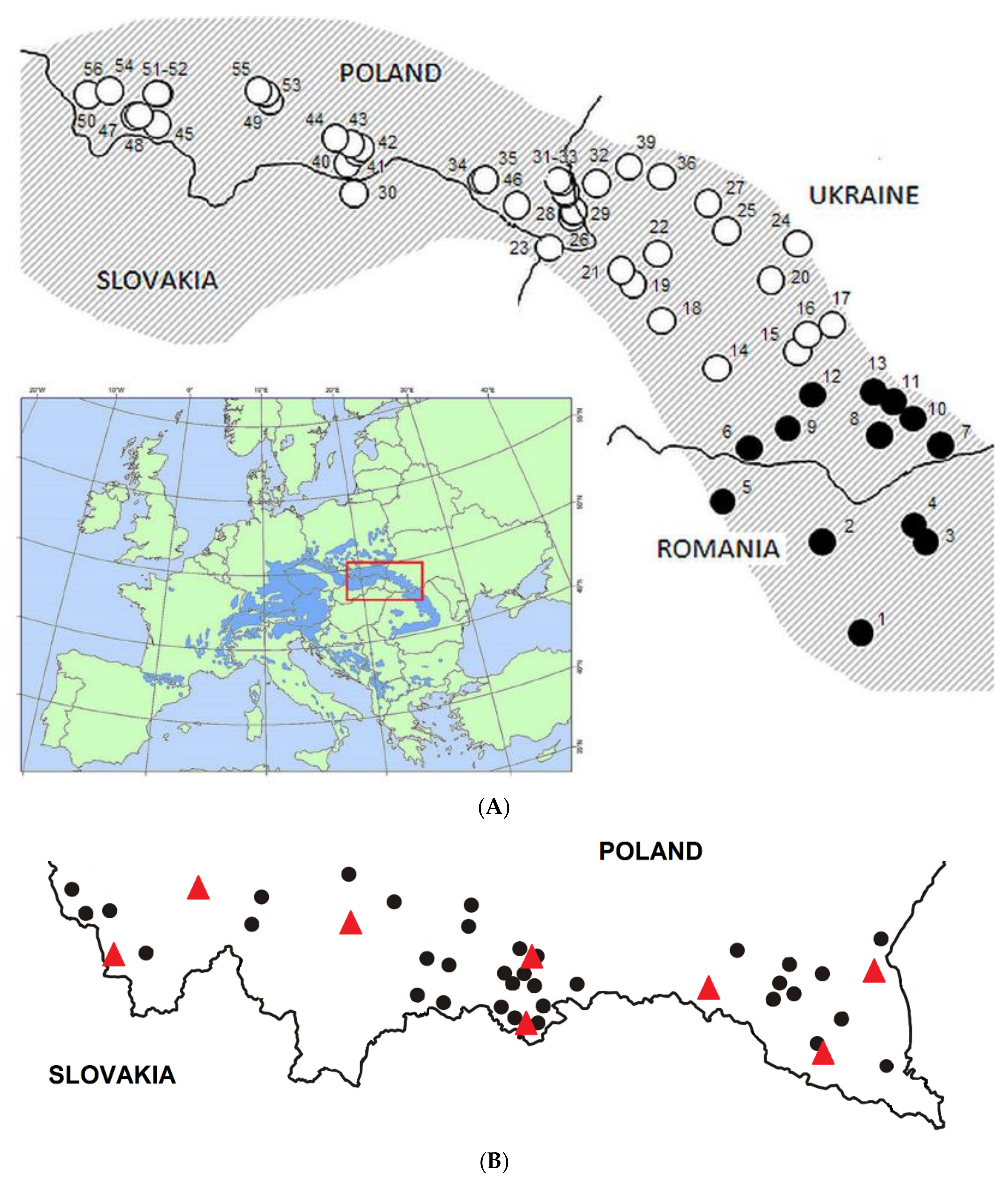
2. Materials and Methods
2.1. Molecular Analyses
2.2. Provenance Trial
2.3. Data Analysis
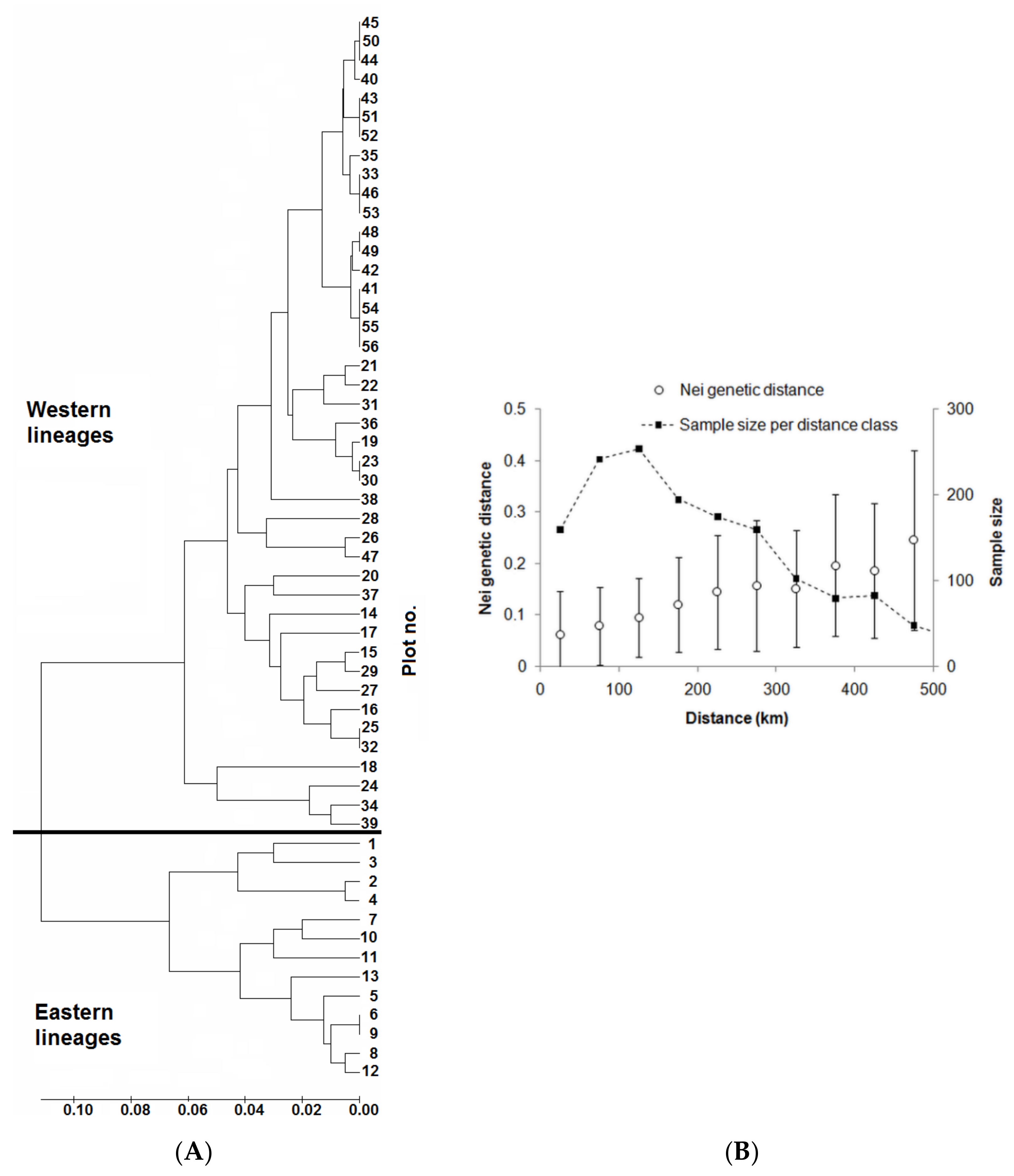
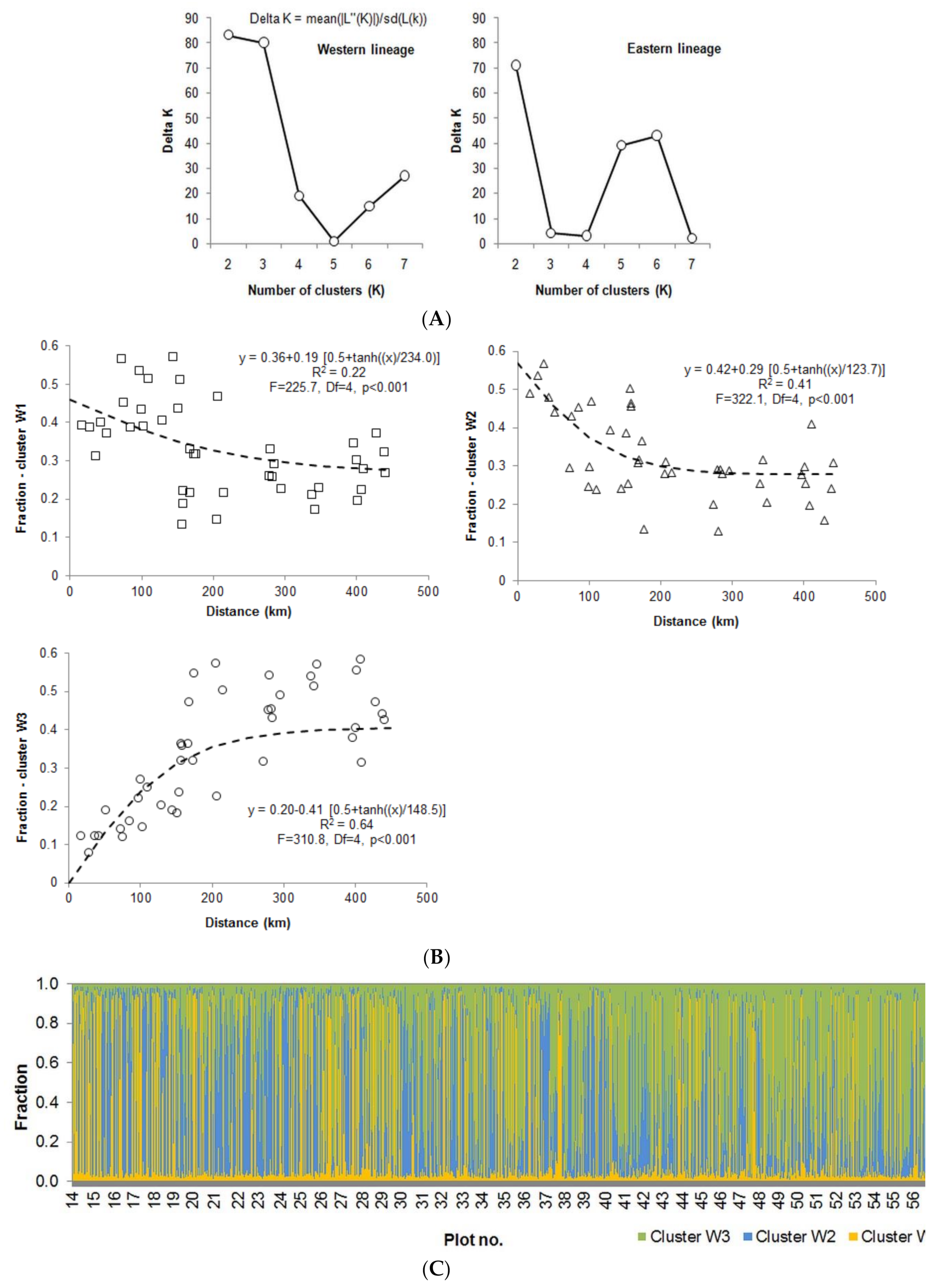
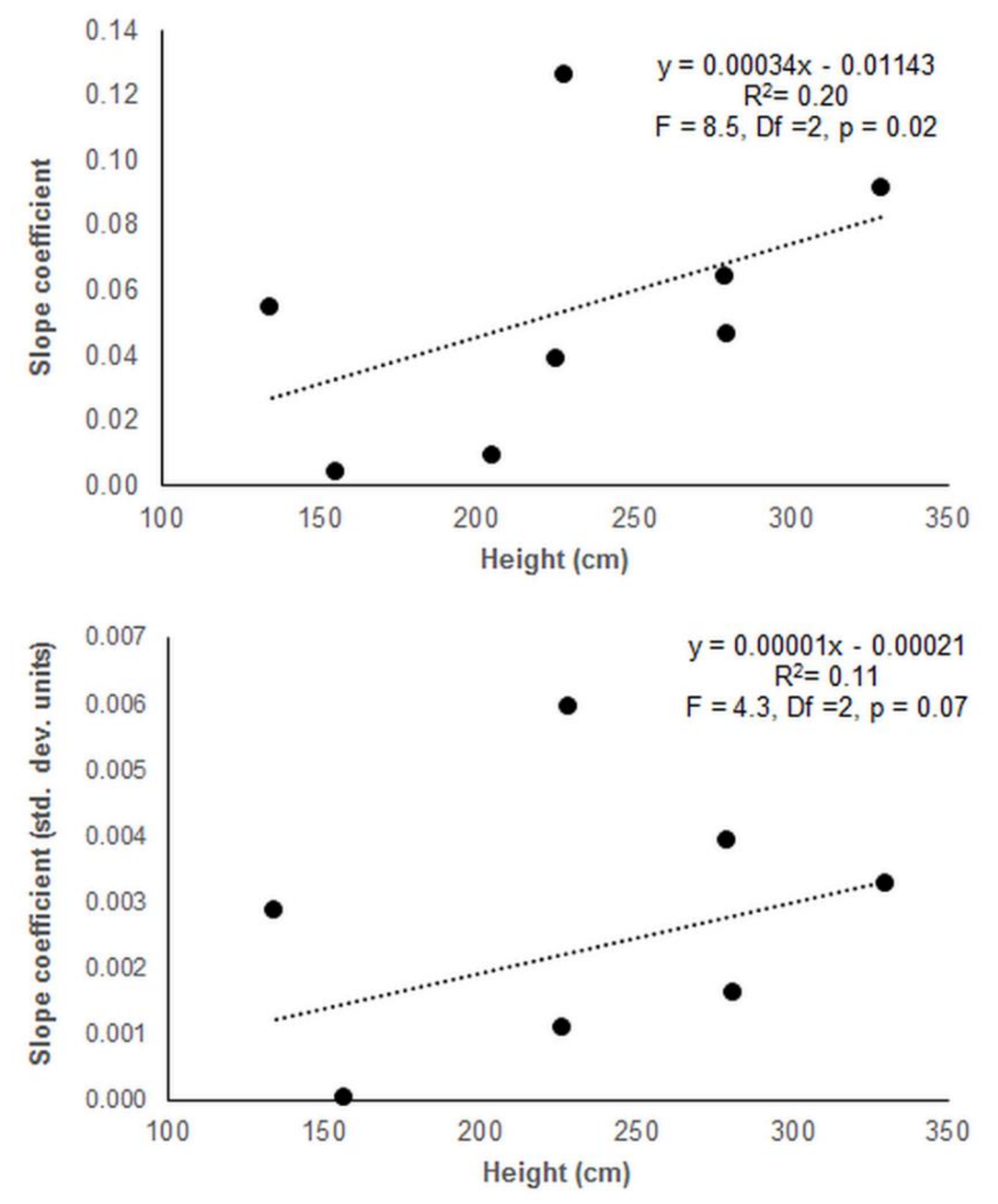
3. Results
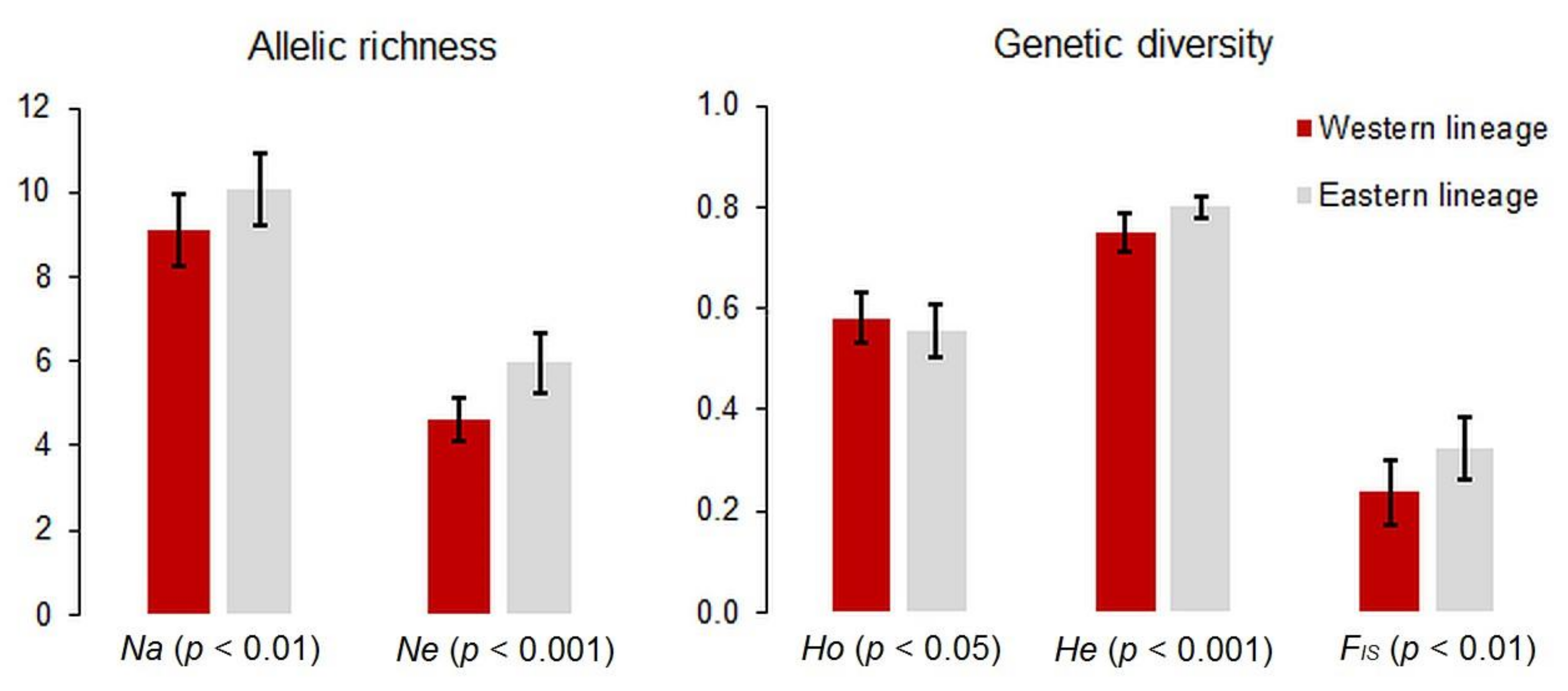
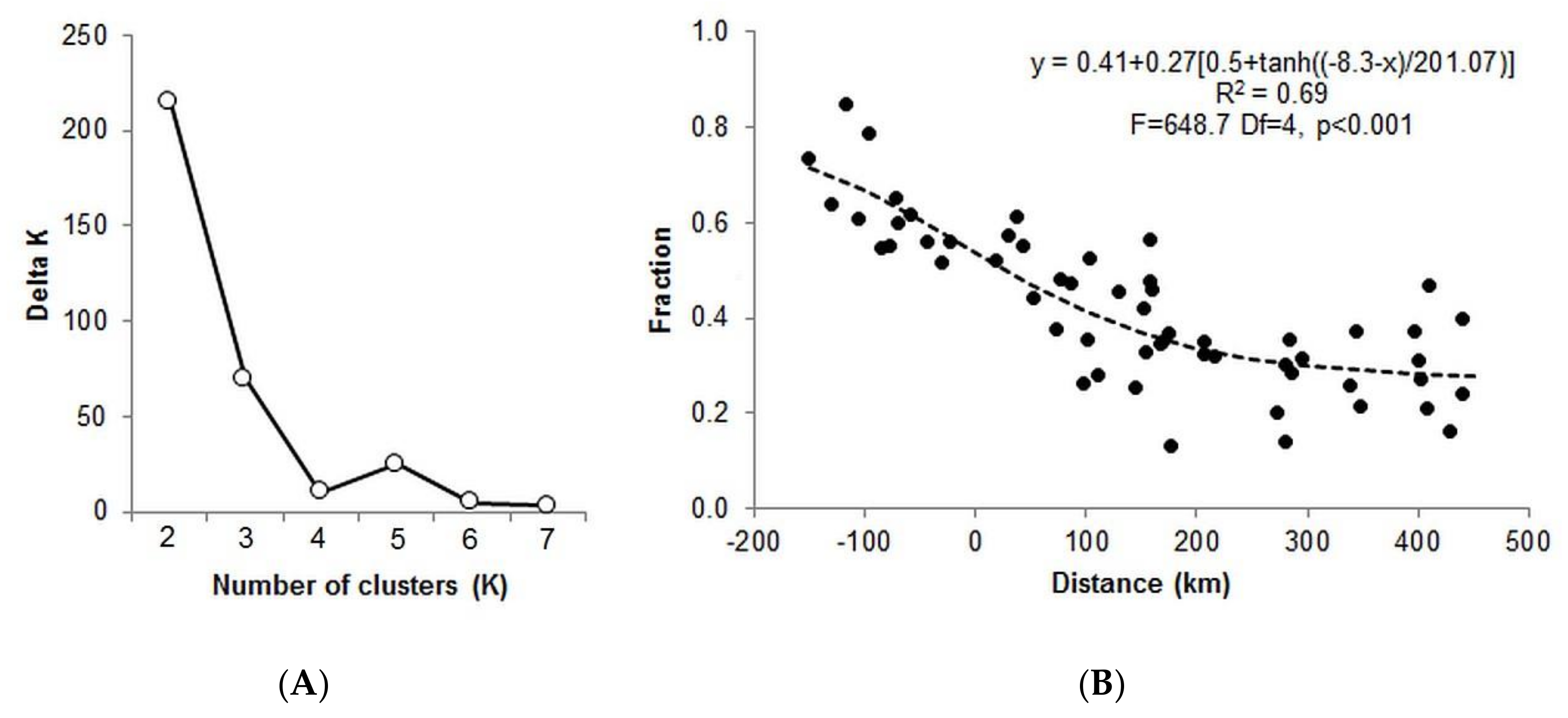
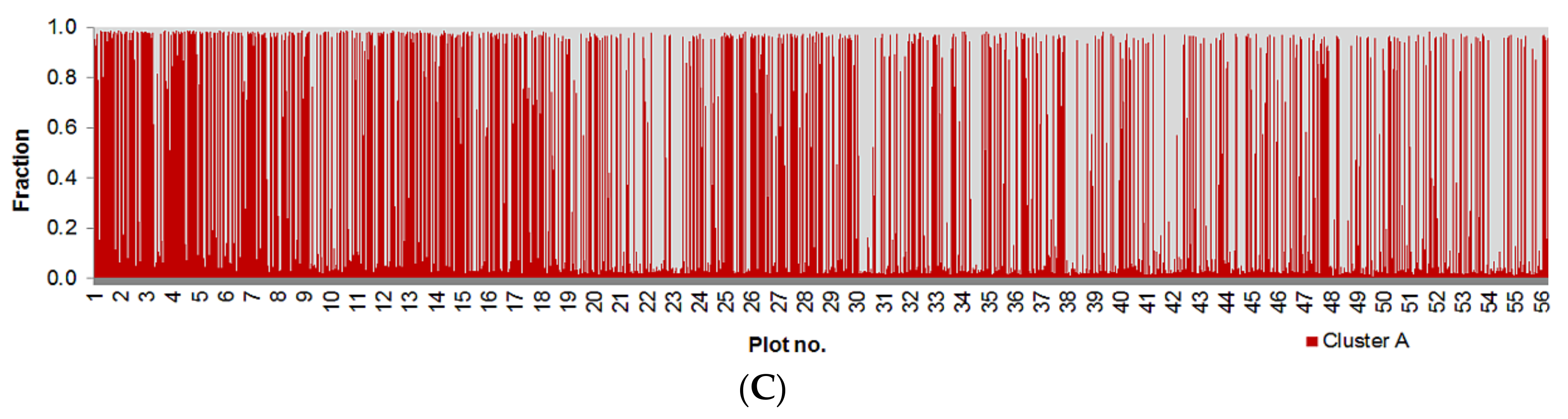
3.1. Genetic Analyses
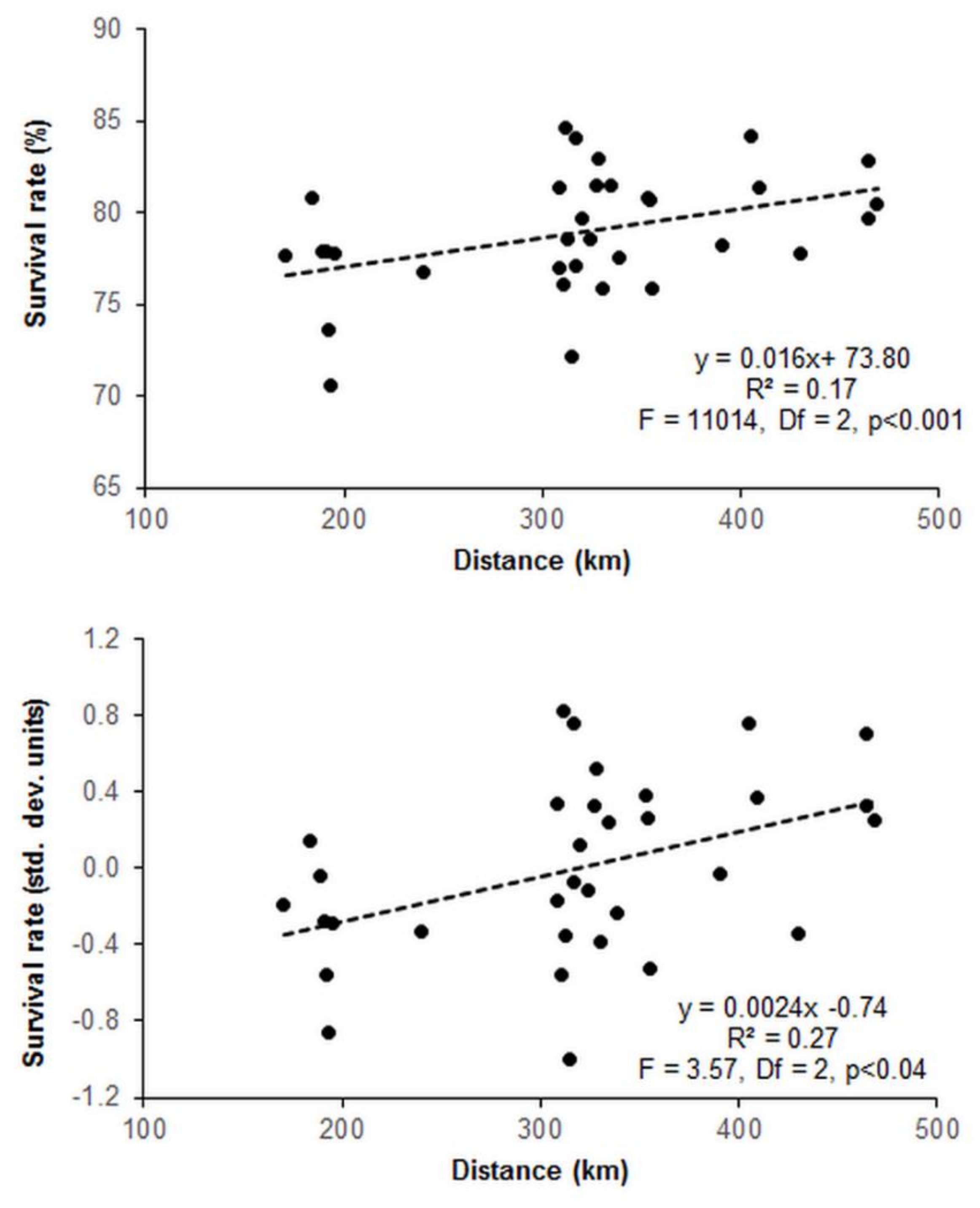
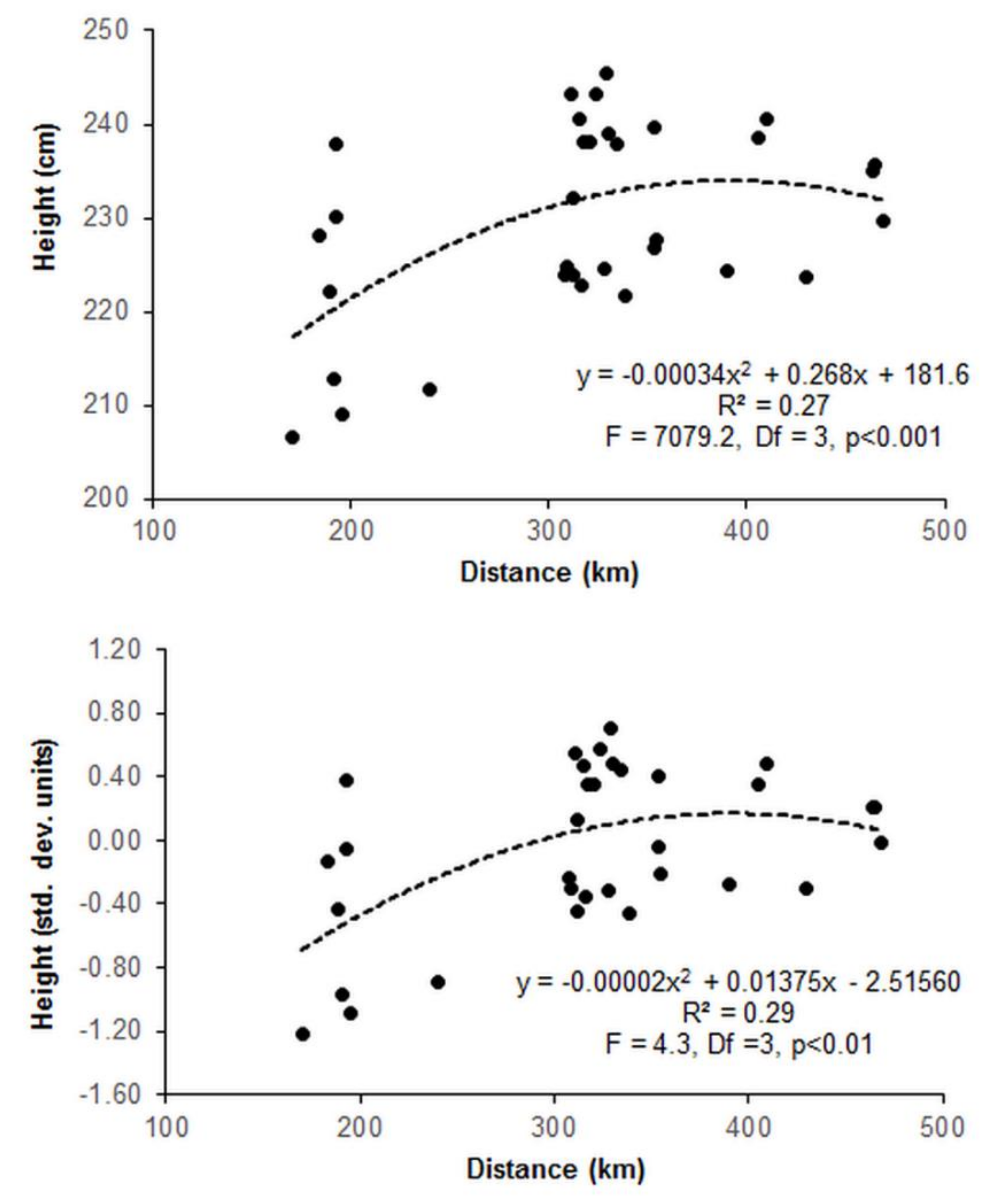
3.2. Provenance Trial
4. Discussion
4.1. Pattern of Genetic Variation
4.2. Survival and Growth Pattern in the Provenance Trial
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hewitt, G. The genetic legacy of the quaternary ice ages. Nature 2000, 405, 907–913. [Google Scholar] [CrossRef] [PubMed]
- Hamrick, J.L. Response of forest trees to global environmental changes. Ecol. Manag. 2004, 197, 323–335. [Google Scholar] [CrossRef]
- Kremer, A.; Ronce, O.; Robledo-Arnuncio, J.J.; Guillaume, F.; Bohrer, G.; Nathan, R.; Bridle, J.R.; Gomulkiewicz, R.; Klein, E.K.; Ritland, K.; et al. Long-distance gene flow and adaptation of forest trees to rapid climate change. Ecol. Lett. 2012, 15, 378–392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alfaro, R.I.; Fady, B.; Vendramin, G.G.; Dawson, I.K.; Fleming, R.A.; Sáenz-Romero, C.; Lindig-Cisneros, R.A.; Murdock, T.; Vinceti, B.; Navarro, C.M.; et al. The role of forest genetic resources in responding to biotic and abiotic factors in the context of anthropogenic climate change. Ecol. Manag. 2014, 333, 76–87. [Google Scholar] [CrossRef]
- Lefèvre, F.; Boivin, T.; Bontemps, A.; Courbet, F.; Davi, H.; Durand-Gillmann, M.; Fady, B.; Gauzere, J.; Gidoin, C.; Karam, M.J.; et al. Considering evolutionary processes in adaptive forestry. Ann. Sci. 2014, 71, 723–739. [Google Scholar] [CrossRef] [Green Version]
- Widmer, A.; Lexer, C. Glacial refugia: Sanctuaries for allelic richness, but not for gene diversity. Trends Ecol. Evol. 2001, 16, 267–269. [Google Scholar] [CrossRef]
- Petit, R.J.; Aguinagalde, I.; De Beaulieu, J.L.; Bittkau, C.; Brewer, S.; Cheddadi, R.; Ennos, R.; Fineschi, S.; Grivet, D.; Lascoux, M.; et al. Glacial refugia: Hotspots but not melting pots of genetic diversity. Science 2003, 300, 1563–1565. [Google Scholar] [CrossRef] [Green Version]
- Rull, V. On microrefugia and cryptic refugia. J. Biogeogr. 2010, 37, 1623–1625. [Google Scholar] [CrossRef]
- Stewart, J.R.; Lister, A.M. Cryptic northern refugia and the origins of the modern biota. Trends Ecol. Evol. 2001, 16, 608–613. [Google Scholar] [CrossRef]
- Alleaume-Benharira, M.; Pen, I.R.; Ronce, O. Geographical patterns of adaptation within a species’ range: Interactions between drift and gene flow. J. Evol. Biol. 2006, 19, 203–215. [Google Scholar] [CrossRef]
- Bell, M.A.; Travis, M.P. Hybridization, transgressive segregation, genetic covariation, and adaptive radiation. Trends Ecol. Evol. 2005, 20, 358–361. [Google Scholar] [CrossRef] [PubMed]
- Wachowiak, W.; Żukowska, W.B.; Wójkiewicz, B.; Cavers, S.; Litkowiec, M. Hybridization in contact zone between temperate European pine species. Tree Genet. Genom. 2016, 12, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Mitsopulos, D.; Panetsos, C. Origin of variation in fir forests of Greece. Silvae Genet. 1987, 36, 1–15. [Google Scholar]
- Krajmerová, D.; Paule, L.; Zhelev, P.; Voleková, M.; Evtimov, I.; Gagov, V.; Gömöry, D. Natural hybridization in eastern-Mediterranean firs: The case of Abies borisii-regis. Plant Biosyst. 2016, 150, 1189–1199. [Google Scholar] [CrossRef]
- Dobrowolska, D.; Bončina, A.; Klumpp, R. Ecology and silviculture of silver fir (Abies alba Mill.): A review. J. For. Res. 2017, 22, 326–335. [Google Scholar] [CrossRef]
- Svenning, J.C.; Skov, F. Limited filling of the potential range in European tree species. Ecol. Lett. 2004, 7, 565–573. [Google Scholar] [CrossRef]
- Tinner, W.; Colombaroli, D.; Heiri, O.; Henne, P.D.; Steinacher, M.; Untenecker, J.; Vescovi, E.; Allen, J.R.M.; Carraro, G.; Conedera, M.; et al. The past ecology of Abies alba provides new perspectives on future responses of silver fir forests to global warming. Ecol. Monogr. 2013, 83, 419–439. [Google Scholar] [CrossRef] [Green Version]
- Büntgen, U.; Tegel, W.; Kaplan, J.O.; Schaub, M.; Hagedorn, F.; Bürgi, M.; Brázdil, R.; Helle, G.; Carrer, M.; Heussner, K.-U.; et al. Placing unprecedented recent fir growth in a European-wide and Holocene-long context. Front. Ecol. Env. 2014, 12, 100–106. [Google Scholar] [CrossRef]
- Ruosch, M.; Spahni, R.; Joos, F.; Henne, P.D.; van der Knaap, W.O.; Tinner, W. Past and future evolution of Abies alba forests in Europe-comparison of a dynamic vegetation model with palaeo data and observations. Glob. Chang. Biol. 2016, 22, 727–740. [Google Scholar] [CrossRef]
- Hansen, J.K.; Larsen, J.B. European silver fir (Abies alba Mill.) provenances from Calabria, southern Italy: 15-year results from Danish provenance field trials. Eur. J. Res. 2004, 123, 127–138. [Google Scholar] [CrossRef]
- Carrer, M.; Nola, P.; Motta, R.; Urbinati, C. Contrasting tree-ring growth to climate responses of Abies alba toward the southern limit of its distribution area. Oikos 2010, 119, 1515–1525. [Google Scholar] [CrossRef]
- Koprowski, M. Reaction of silver fir (Abies alba) growing outside its natural range to extreme weather events and a long-term increase in march temperature. Tree Ring Res. 2013, 69, 49–61. [Google Scholar] [CrossRef] [Green Version]
- Gazol, A.; Camarero, J.J.; Gutiérrez, E.; Popa, I.; Andreu-Hayles, L.; Motta, R.; Nola, P.; Ribas, M.; Sangüesa-Barreda, G.; Urbinati, C.; et al. Distinct effects of climate warming on populations of silver fir (Abies alba) across Europe. J. Biogeogr. 2015, 42, 1150–1162. [Google Scholar] [CrossRef] [Green Version]
- George, J.P.; Schueler, S.; Karanitsch-Ackerl, S.; Mayer, K.; Klumpp, R.T.; Grabner, M. Inter-and intra-specific variation in drought sensitivity in Abies spec. and its relation to wood density and growth traits. Agric. Meteorol. 2015, 214–215, 430–443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kerr, G.; Stokes, V.; Peace, A.; Jinks, R. Effects of provenance on the survival, growth and stem form of European silver fir (Abies alba Mill.) in Britain. Eur. J. Res. 2015, 134, 349–363. [Google Scholar] [CrossRef] [Green Version]
- Liepelt, S.; Cheddadi, R.; de Beaulieu, J.-L.L.; Fady, B.; Gömöry, D.; Hussendörfer, E.; Konnert, M.; Litt, T.; Longauer, R.; Terhürne-Berson, R.; et al. Postglacial range expansion and its genetic imprints in Abies alba (Mill.)—A synthesis from palaeobotanic and genetic data. Rev. Palaeobot. Palynol. 2009, 153, 139–149. [Google Scholar] [CrossRef]
- Heer, K.; Behringer, D.; Piermattei, A.; Bässler, C.; Brandl, R.; Fady, B.; Jehl, H.; Liepelt, S.; Lorch, S.; Piotti, A.; et al. Linking dendroecology and association genetics in natural populations: Stress responses archived in tree rings associate with SNP genotypes in silver fir (Abies alba Mill.). Mol. Ecol. 2018, 27, 1428–1438. [Google Scholar] [CrossRef]
- Liepelt, S.; Bialozyt, R.; Ziegenhagen, B. Wind-dispersed pollen mediates postglacial gene flow among refugia. Proc. Natl. Acad. Sci. USA 2002, 99, 14590–14594. [Google Scholar] [CrossRef] [Green Version]
- Gömöry, D.; Longauer, R.; Liepelt, S.; Ballian, D.; Brus, R.; Kraigher, H.; Parpan, V.I.; Parpan, T.V.; Paule, L.; Stupar, V.I.; et al. Variation patterns of mitochondrial DNA of Abies alba Mill. in suture zones of postglacial migration in Europe. Acta Soc. Bot. Pol. 2004, 73, 203–206. [Google Scholar] [CrossRef] [Green Version]
- Gömöry, D.; Paule, L.; Krajmerová, D.; Romšáková, I.; Longauer, R. Admixture of genetic lineages of different glacial origin: A case study of Abies alba Mill. in the Carpathians. Plant Syst. Evol. 2012, 298, 703–712. [Google Scholar] [CrossRef]
- Kaltenrieder, P.; Belis, C.A.; Hofstetter, S.; Ammann, B.; Ravazzi, C.; Tinner, W. Environmental and climatic conditions at a potential glacial refugial site of tree species near the Southern Alpine glaciers. New insights from multiproxy sedimentary studies at Lago della Costa (Euganean Hills, Northeastern Italy). Quat. Sci. Rev. 2009, 28, 2647–2662. [Google Scholar] [CrossRef]
- Magri, D.; Agrillo, E.; Di Rita, F.; Furlanetto, G.; Pini, R.; Ravazzi, C.; Spada, F. Holocene dynamics of tree taxa populations in Italy. Rev. Palaeobot. Palynol. 2015, 218, 267–284. [Google Scholar] [CrossRef]
- Cheddadi, R.; Birks, H.J.B.; Tarroso, P.; Liepelt, S.; Gömöry, D.; Dullinger, S.; Meier, E.S.; Hülber, K.; Maiorano, L.; Laborde, H. Revisiting tree-migration rates: Abies alba (Mill.), a case study. Veg. Hist. Archaeobot. 2014, 23, 113–122. [Google Scholar] [CrossRef]
- Bosela, M.; Popa, I.; Gömöry, D.; Longauer, R.; Tobin, B.; Kyncl, J.; Kyncl, T.; Nechita, C.; Petráš, R.; Sidor, C.G.; et al. Effects of post-glacial phylogeny and genetic diversity on the growth variability and climate sensitivity of European silver fir. J. Ecol. 2016, 104, 716–724. [Google Scholar] [CrossRef] [Green Version]
- Teodosiu, M.; Mihai, G.; Fussi, B.; Ciocîrlan, E. Genetic diversity and structure of silver fir (Abies alba Mill.) at the south-eastern limit of its distribution range. Ann. Res. 2019, 62, 139–156. [Google Scholar] [CrossRef]
- WISL. Wielkoobszarowa Inwentaryzacja Stanu Lasu (Okr. 2014–2018); Biuro Urządzania Lasu i Geodezji Leśnej: Warszawa, Poland, 2018. [Google Scholar]
- Kramer, W. Die Weißtanne (Abies alba Mill.) in Ost-und Südosteuropa; Gustav Fischer Verlag: Stuttgart, Germany, 1992. [Google Scholar]
- Matuszkiewicz, J.M. Zespoły leśne Polski; Wydawnictwo Naukowe PWN: Warszawa, Poland, 2001. [Google Scholar]
- Fabijanowski, J.; Jaworski, A. Gospodarstwo leśne (Silviculture). In Karpaty Polskie, Człowiek i Jego Działalność; Warszyńska, J., Ed.; Wydawnictwo UJ: Kraków, Poland, 1995; pp. 253–263. [Google Scholar]
- Khanuja, S.; Shasany, A.; Darokar, M.P.; Kumar, S. Rapid Isolation of DNA from Dry and Fresh Samples of Plants Producing Large Amounts of Secondary Metabolites and Essential Oils. Plant Mol. Biol. Rep. 1999, 17, 74. [Google Scholar] [CrossRef]
- Cremer, E.; Liepelt, S.; Sebastiani, F.; Buonamici, A.; Michalczyk, I.M.; Ziegenhagen, B.; Vendramin, G.G. Identification and characterization of nuclear microsatellite loci in Abies alba Mill. Mol. Ecol. Notes 2006, 6, 374–376. [Google Scholar] [CrossRef]
- Kempf, M.; Sabor, J.; Stanuch, H. Ocena cech adaptacyjnych i morfologicznych potomstwa drzewostanow jodlowych objetych ochrona w Karpackim Banku Genow. Sylwan 2003, 147, 3–15. [Google Scholar]
- Kempf, M.; Sabor, J. Evaluation of the variability of adaptive traits in 5−year−old silver fir progenies from provenances protected on conservation plots in the Carpathian Gene Bank. Sylwan 2009, 153, 651–661. [Google Scholar]
- Wypych, A.; Ustrnul, Z.; Schmatz, D.R. Long-term variability of air temperature and precipitation conditions in the Polish Carpathians. J. Mt. Sci. 2018, 15, 237–253. [Google Scholar] [CrossRef]
- Paszyński, J.; Niedźwiedź, T. Klimat. In Geografia Polski. Środowisko Przyrodnicze.; Starkel, L., Ed.; Wydawnictwo Naukowe PWN: Warszawa, Poland, 1999; pp. 288–343. [Google Scholar]
- Van Oosterhout, C.; Hutchinson, W.F.; Wills, D.P.M.; Shipley, P. MICRO-CHECKER: Software for identifying and correcting genotyping errors in microsatellite data. Mol. Ecol. Notes 2004, 4, 535–538. [Google Scholar] [CrossRef]
- Van Oosterhout, C.; Weetman, D.; Hutchinson, W.F. Estimation and adjustment of microsatellite null alleles in nonequilibrium populations. Mol. Ecol. Notes 2006, 6, 255–256. [Google Scholar] [CrossRef]
- Brookfield, J.F.Y. A simple new method for estimating null allele frequency from heterozygote deficiency. Mol. Ecol. 1996, 5, 453–455. [Google Scholar] [CrossRef] [PubMed]
- Raymond, M.; Rousset, F. GENEPOP (version 1.2): Population genetics software for exact tests and ecumenicism. J. Hered. 1995, 86, 248–249. [Google Scholar] [CrossRef]
- Nei, M. Estimation of average heterozygosity and genetic distance from a small number of individuals. Genetics 1978, 89, 583–590. [Google Scholar]
- Peakall, R.; Smouse, P.E. GENALEX 6: Genetic analysis in Excel. Population genetic software for teaching and research. Mol. Ecol. Notes 2006, 6, 288–295. [Google Scholar] [CrossRef]
- Peakall, R.; Smouse, P.E. GenALEx 6.5: Genetic analysis in Excel. Population genetic software for teaching and research-an updat. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef] [Green Version]
- Efron, B.; Tibshirani, R.J. An Introduction to the Bootstrap; Chapmann&Hall/CRC: New York, NY, USA, 1993; ISBN 0412042312. [Google Scholar]
- Slatkin, M. A measure of population subdivision based on microsatellite allele frequencies. Genetics 1995, 139, 457–462. [Google Scholar]
- Nei, M. Genetic Distance between Populations. Am. Nat. 1972, 106, 283–292. [Google Scholar] [CrossRef]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software STRUCTURE: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Earl, D.A.; VonHoldt, B.M. STRUCTURE HARVESTER: A website and program for visualizing STRUCTURE output and implementing the Evanno method. Conserv. Genet. Resour. 2012, 4, 359–361. [Google Scholar] [CrossRef]
- Kopelman, N.M.; Mayzel, J.; Jakobsson, M.; Rosenberg, N.A.; Mayrose, I. Clumpak: A program for identifying clustering modes and packaging population structure inferences across K. Mol. Ecol. Resour. 2015, 1179–1191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pawlaczyk, E.; Kroplewska, I.; Bobowicz, M. Postglacial migration of silver fir (Abies alba Mill.) to Poland-Analysis on the basis of mitochondrial DNA polymorphism. Sylwan 2013, 157, 458–463. [Google Scholar]
- Litkowiec, M.; Lewandowski, A.; Raczka, G. Spatial pattern of the mitochondrial and chloroplast genetic variation in poland as a result of the migration of Abies alba Mill. from different glacial refugia. Forests 2016, 7, 1–13. [Google Scholar] [CrossRef]
- Lewandowski, A.; Filipiak, M.; Burczyk, J. Genetic variation of Abies alba Mill. in Polish part of Sudety Mts. Acta Soc. Bot. Pol. 2001, 70, 215–219. [Google Scholar] [CrossRef]
- Longauer, R.; Paule, L.; Andonoski, A. Genetic diversity of southern populations of Abies alba Mill. Int. J. For. Genet. 2003, 10, 1–10. [Google Scholar]
- Parducci, L.; Szmidt, A.E.; Villani, F.; Wang, X.R.; Cherubini, M. Genetic variation of Abies alba in Italy. Hereditas 1996, 125, 11–18. [Google Scholar] [CrossRef]
- Mejnartowicz, L. Genetic analysis of silver-fir populations in the north Carpathian and Sudeten mountains. Acta Soc. Bot. Pol. 2004, 73, 285–292. [Google Scholar] [CrossRef] [Green Version]
- Ballian, D.; Kajba, D. Estimation of the isoenzyme genetic variability of the silver fir (Abies alba Mill.) the area of Gorski Kotar (Croatia). Period. Biol. 2005, 107, 67–72. [Google Scholar]
- Kormuťák, A.; Kádasi-Horáková, M.; Vooková, B.; Gömöry, D. Genetic structure of silver fir primeval forests in Slovakia. Lesn. Časopis For. J. 2008, 54, 37–42. [Google Scholar]
- Cremer, E.; Ziegenhagen, B.; Schulerowitz, K.; Mengel, C.; Donges, K.; Bialozyt, R.; Hussendörfer, E.; Liepelt, S. Local seed dispersal in European silver fir (Abies alba Mill.): Lessons learned from a seed trap experiment. Trees Struct. Funct. 2012, 26, 987–996. [Google Scholar] [CrossRef]
- Dering, M.; Sekiewicz, K.; Boratyńska, K.; Litkowiec, M.; Iszkuło, G.; Romo, A.; Boratyński, A. Genetic diversity and inter-specific relations of western Mediterranean relic Abies taxa as compared to the Iberian, Abies alba. Flora Morphol. Distrib. Funct. Ecol. Plants 2014, 209, 367–374. [Google Scholar] [CrossRef] [Green Version]
- Cvrčková, H.; Máchová, P.; Malá, J. Use of nuclear microsatellite loci for evaluating genetic diversity among selected populations of Abies alba Mill. in the Czech Republic. J. Sci. 2015, 61, 345–351. [Google Scholar] [CrossRef] [Green Version]
- Belletti, P.; Ferrazzini, D.; Ducci, F.; De Rogatis, A.; Mucciarelli, M. Genetic diversity of italian populations of Abies alba. Dendrobiology 2017, 77, 147–159. [Google Scholar] [CrossRef]
- White, G.M.; Boshier, D.H.; Powell, W. Increased pollen flow counteracts fragmentation in a tropical dry forest: An example from Swietenia humilis Zuccarini. Proc. Natl. Acad. Sci. USA 2002, 99, 2038–2042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Craft, K.J.; Ashley, M.V. Pollen-mediated gene flow in isolated and continuous stands of bur oak, Quercus macrocarpa (Fagaceae). Am. J. Bot. 2010, 97, 1999–2006. [Google Scholar] [CrossRef] [Green Version]
- Ballian, D.; Bogunić, F.; Bajrić, M.; Kajba, D.; Kraigher, H.; Konnert, M. The genetic population study of Balkan silver fir (Abies alba Mill.). Period. Biol. 2012, 114, 55–65. [Google Scholar]
- Willis, K.J.; Van Andel, T.H. Trees or no trees? The environments of central and eastern Europe during the Last Glaciation. Quat. Sci. Rev. 2004, 23, 2369–2387. [Google Scholar] [CrossRef]
- Willis, K.J.; Rudner, E.; Sümegi, P. The full-glacial forests of central and southeastern Europe. Quat. Res. 2000, 53, 203–213. [Google Scholar] [CrossRef]
- Terhürne-Berson, R.; Litt, T.; Cheddadi, R. The spread of Abies throughout Europe since the last glacial period: Combined macrofossil and pollen data. Veg. Hist. Archaeobot. 2004, 13, 257–268. [Google Scholar] [CrossRef]
- Culiberg, M. Late Glacial Vegetation in Slovenia; SAZU: Ljubljana, Sovenia, 1991; ISBN 9788671310512. [Google Scholar]
- Obidowicz, A.; Szczepanek, K.; Madeyska, E.; Nalepka, D. Abies alba Mill.—Fir. In Late Glacial and Holocene History of Vegetation in Poland Based on Isopollen Maps; Ralska-Jasiewiczowa, M., Latałowa, M., Wasylikowa, K., Tobolski, K., Madeyska, E., Wright, H.E.J., Turner, C., Eds.; W. Szafer Institute of Botany, Polish Academy of Sciences: Kraków, Poland, 2004; pp. 31–38. [Google Scholar]
- Tinner, W.; Lotter, A.F. Central European vegetation response to abrupt climate change at 8.2 ka. Geology 2001, 29, 551–554. [Google Scholar] [CrossRef] [Green Version]
- Roschanski, A.M.; Csilléry, K.; Liepelt, S.; Oddou-Muratorio, S.; Ziegenhagen, B.; Huard, F.; Ullrich, K.K.; Postolache, D.; Vendramin, G.G.; Fady, B. Evidence of divergent selection for drought and cold tolerance at landscape and local scales in Abies alba Mill. in the French Mediterranean Alps. Mol. Ecol. 2016, 25, 776–794. [Google Scholar] [CrossRef] [PubMed]
- Csilléry, K.; Ovaskainen, O.; Sperisen, C.; Buchmann, N.; Widmer, A.; Gugerli, F. Adaptation to local climate in multi-trait space: Evidence from silver fir (Abies alba Mill.) populations across a heterogeneous environment. Heredity 2020, 124, 77–92. [Google Scholar] [CrossRef] [Green Version]
- Matías, L.; Gonzalez-Díaz, P.; Quero, J.L.; Camarero, J.J.; Lloret, F.; Jump, A.S. Role of geographical provenance in the response of silver fir seedlings to experimental warming and drought. Tree Physiol. 2016, 36, 1236–1246. [Google Scholar] [CrossRef] [Green Version]
- Konôpková, A.; Kurjak, D.; Kmeť, J.; Klumpp, R.; Longauer, R.; Ditmarová, Ľ; Gömöry, D. Differences in photochemistry and response to heat stress between silver fir (Abies alba Mill.) provenances. Trees Struct. Funct. 2018, 32, 73–86. [Google Scholar] [CrossRef]
- Skrzyszewska, K. Wartość selekcyjna jodły pospolitej (Abies alba Mill.) polskich pochodzeń w okresie juwenilnego wzrostu w zróżnicowanych warunkach siedliskowych. Zesz. Nauk. AR Krak. 2013, 377, 7–31. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source of Variation | Degrees of Freedom (Df) | Variance Component (%) | R Statistic | Variance Component (%) | F Statistic | ||
|---|---|---|---|---|---|---|---|
| Between lineages | 1 | 1.7 | *** | 0.017 | 0.0 | 0.000 | |
| Between subpopulations | 54 | 8.2 | *** | 0.084 | 2.8 | *** | 0.028 |
| Within subpopulations | 3466 | 90.1 | *** | 0.099 | 97.2 | *** | 0.016 |
| Total | 3521 | 100 | - | 100 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kempf, M.; Zarek, M.; Paluch, J. The Pattern of Genetic Variation, Survival and Growth in the Abies alba Mill. Population within the Introgression Zone of Two Refugial Lineages in the Carpathians. Forests 2020, 11, 849. https://0-doi-org.brum.beds.ac.uk/10.3390/f11080849
Kempf M, Zarek M, Paluch J. The Pattern of Genetic Variation, Survival and Growth in the Abies alba Mill. Population within the Introgression Zone of Two Refugial Lineages in the Carpathians. Forests. 2020; 11(8):849. https://0-doi-org.brum.beds.ac.uk/10.3390/f11080849
Chicago/Turabian StyleKempf, Marta, Marcin Zarek, and Jarosław Paluch. 2020. "The Pattern of Genetic Variation, Survival and Growth in the Abies alba Mill. Population within the Introgression Zone of Two Refugial Lineages in the Carpathians" Forests 11, no. 8: 849. https://0-doi-org.brum.beds.ac.uk/10.3390/f11080849