Does Shrub Encroachment Indicate Ecosystem Degradation? A Perspective Based on the Spatial Patterns of Woody Plants in a Temperate Savanna-Like Ecosystem of Inner Mongolia, China

Abstract
:1. Introduction
2. Materials and Methods
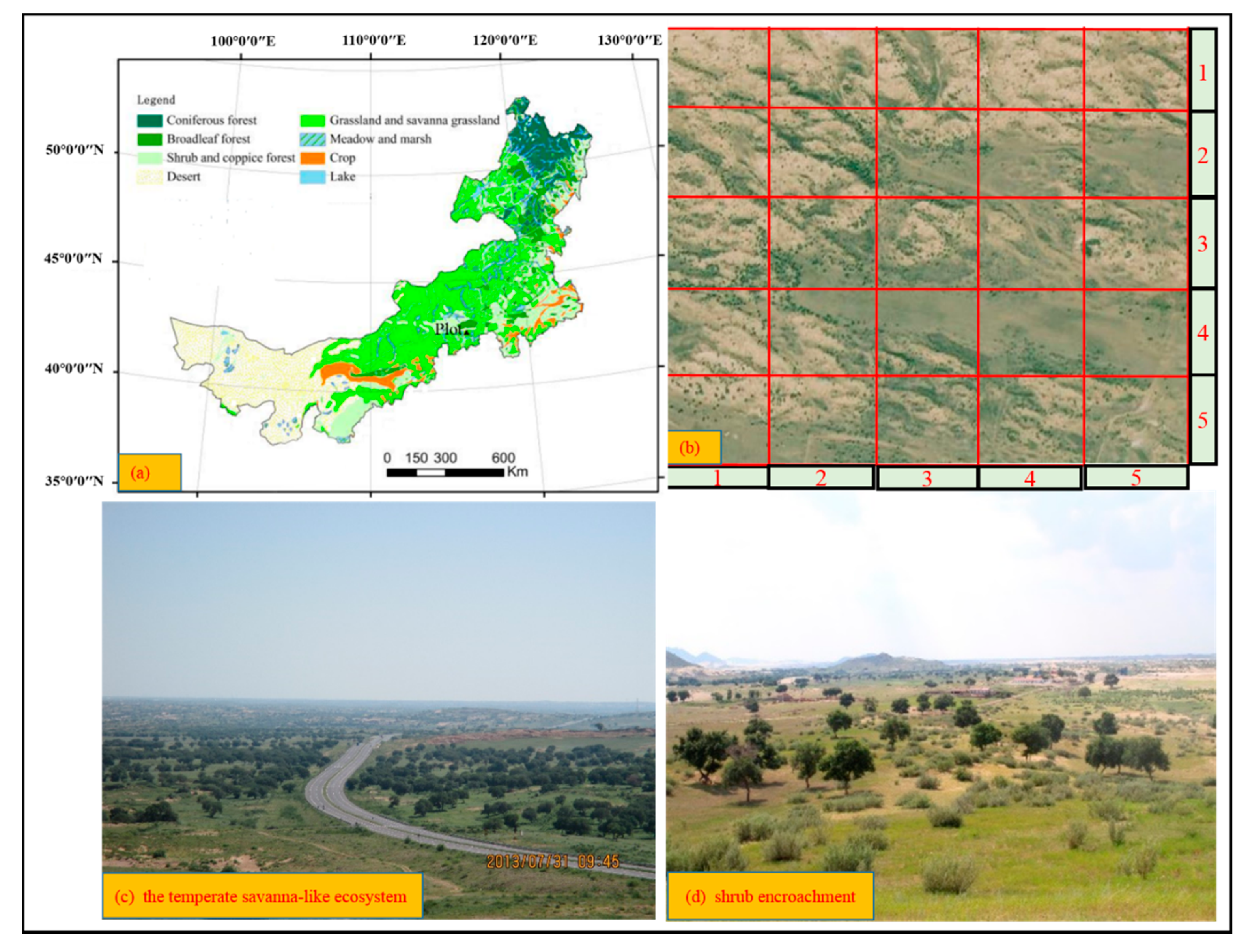
2.1. Study Area
2.2. Research Site and Data Collection
2.3. Data Analysis
3. Results
3.1. Characteristics of the Temperate Savanna Ecosystem
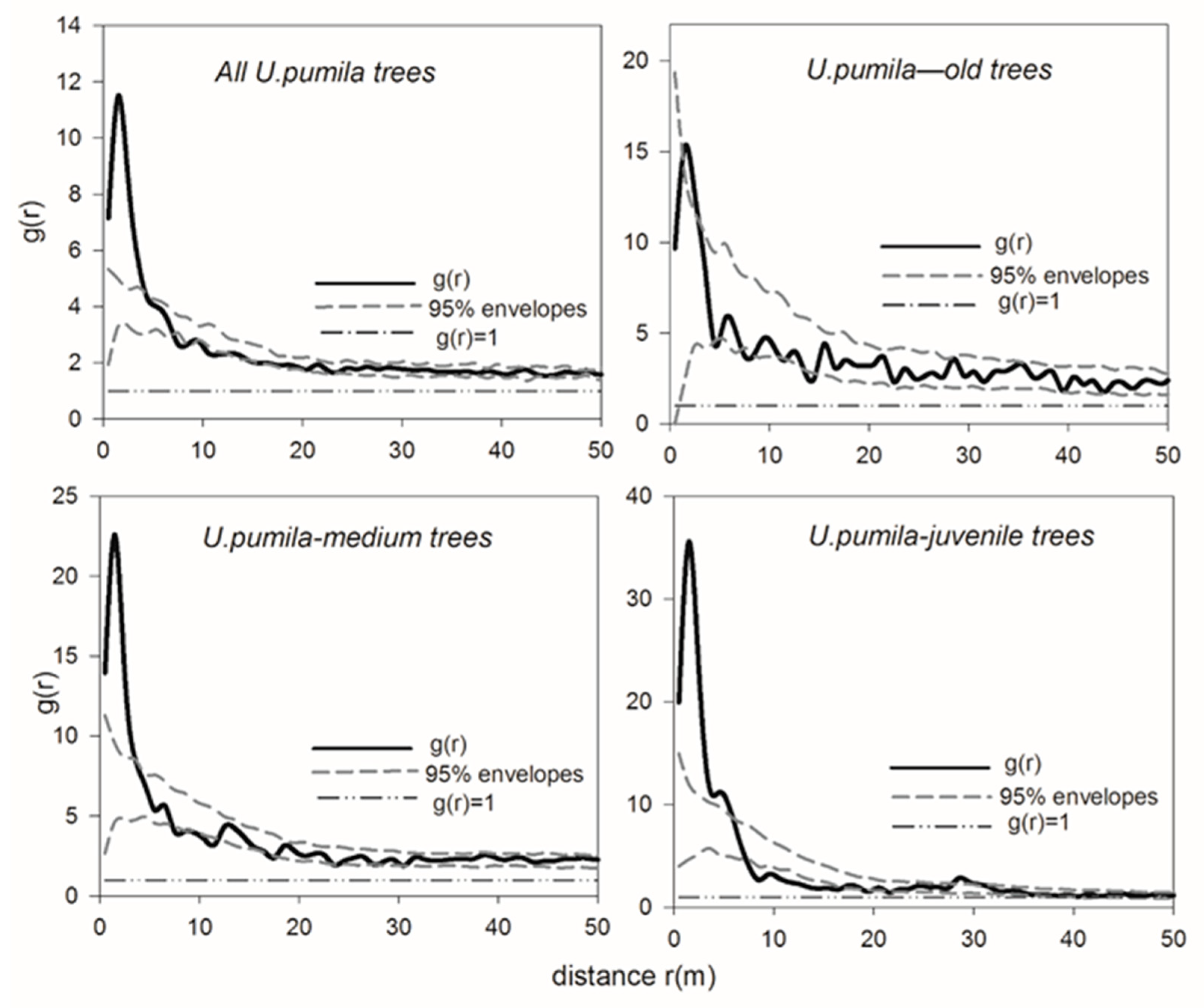
3.2. Spatial Distribution Patterns of U. pumila Trees—A Random Distribution of Old Trees and an Aggregated Distribution of Juvenile Trees
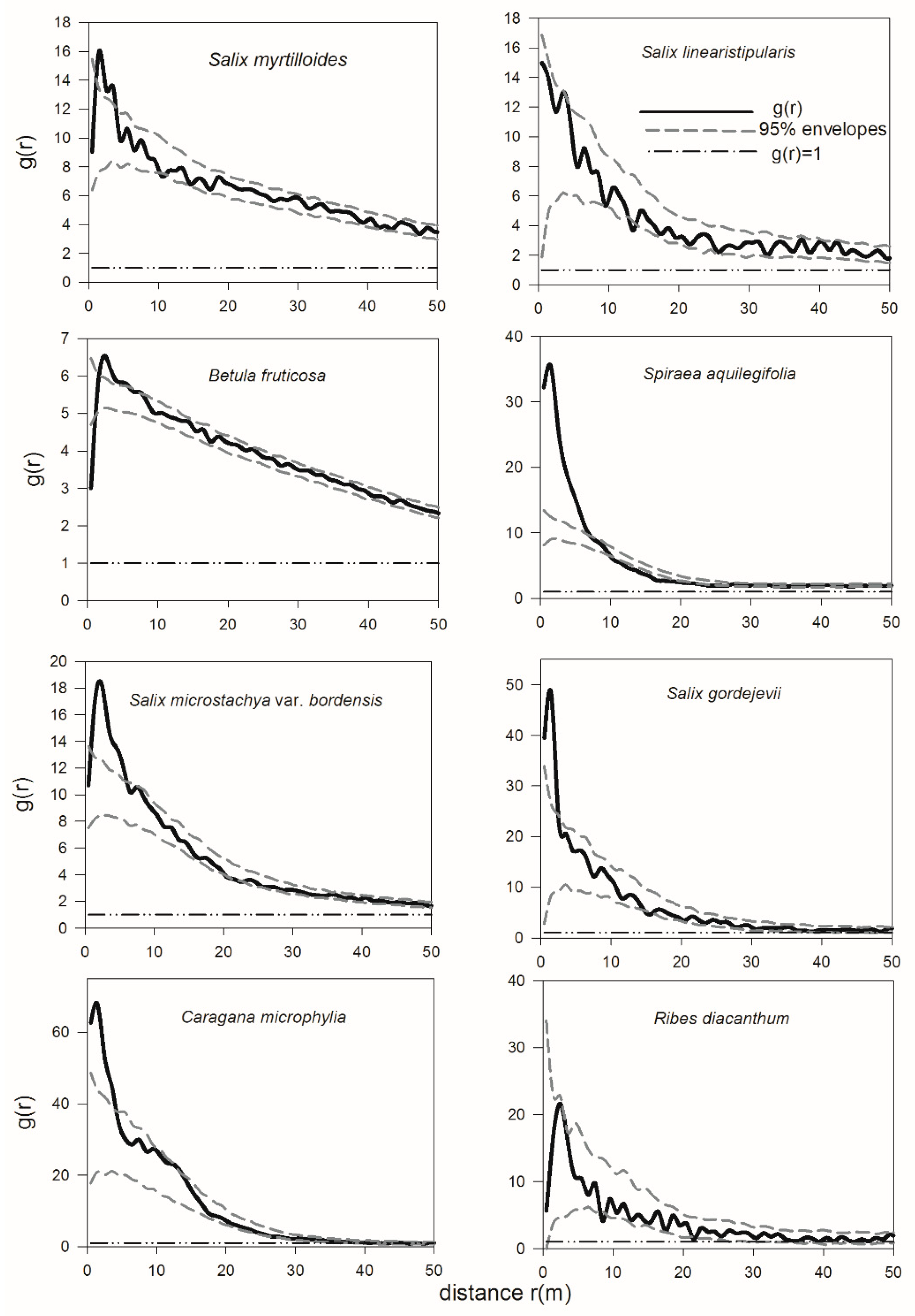
3.3. Aggregated Distribution of Shrubs at Specific Scales
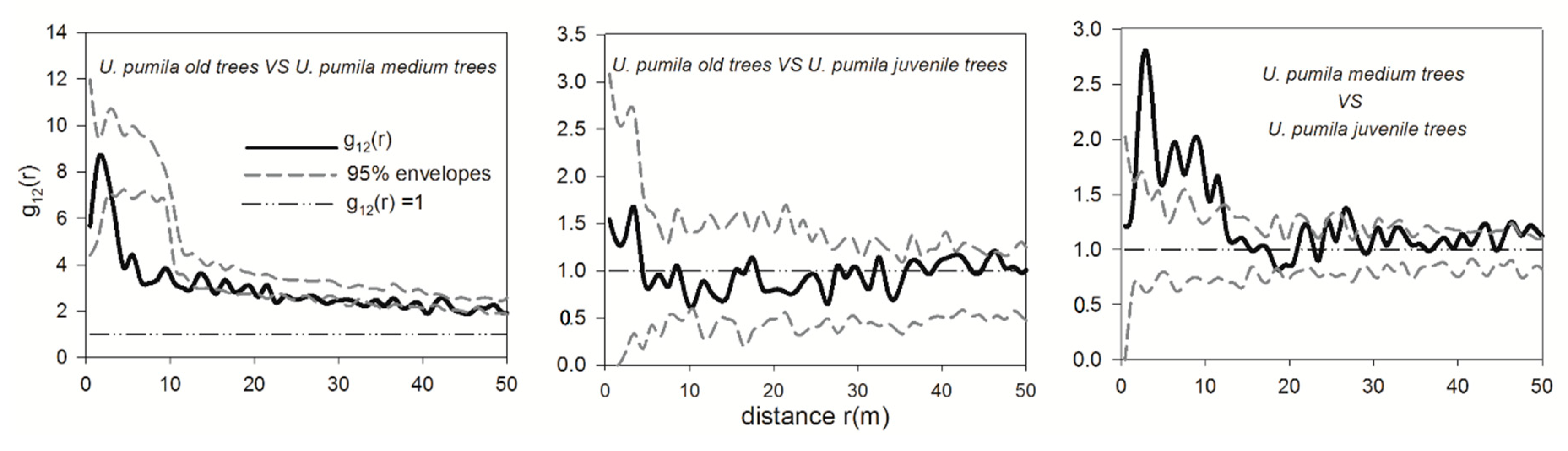
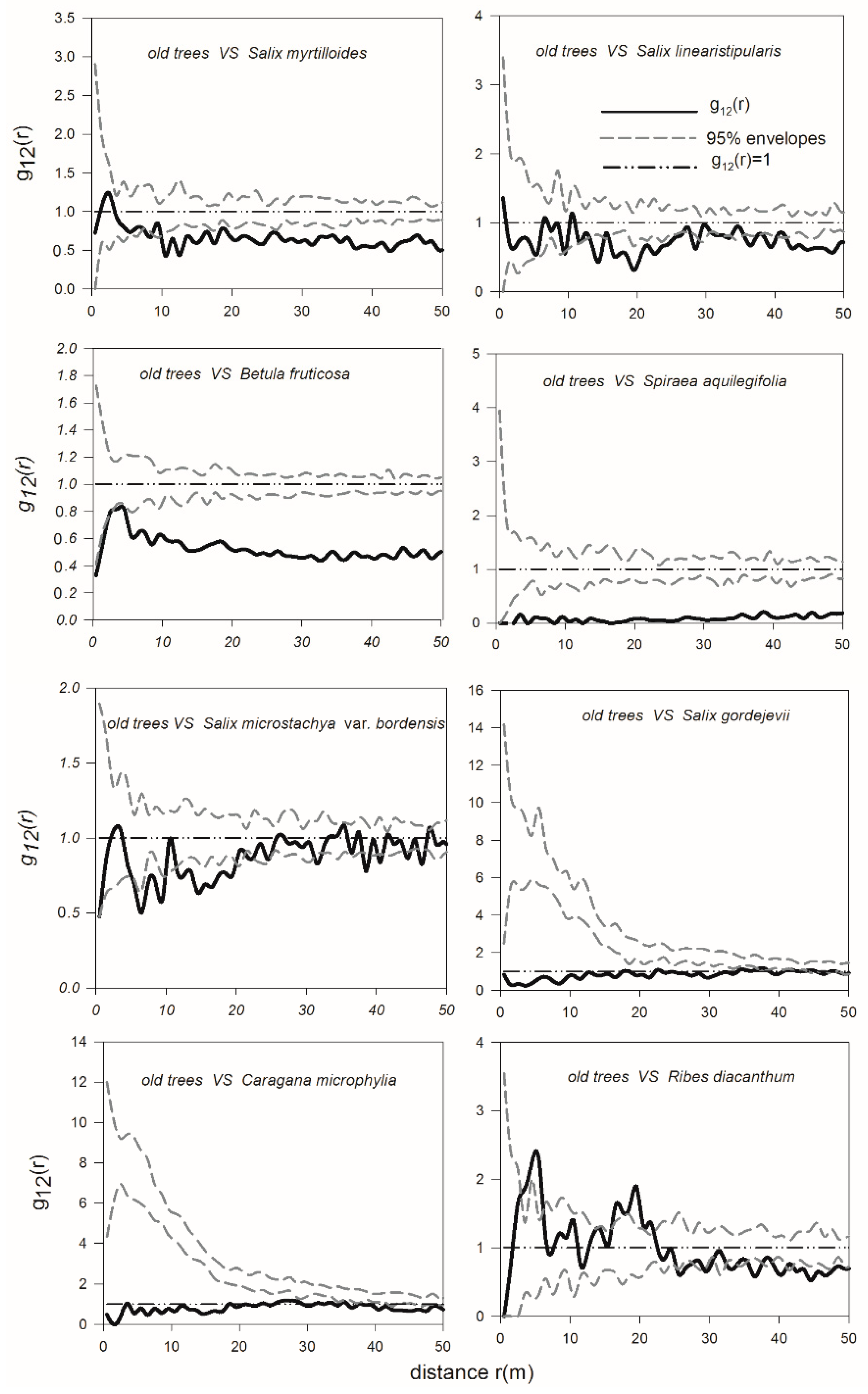
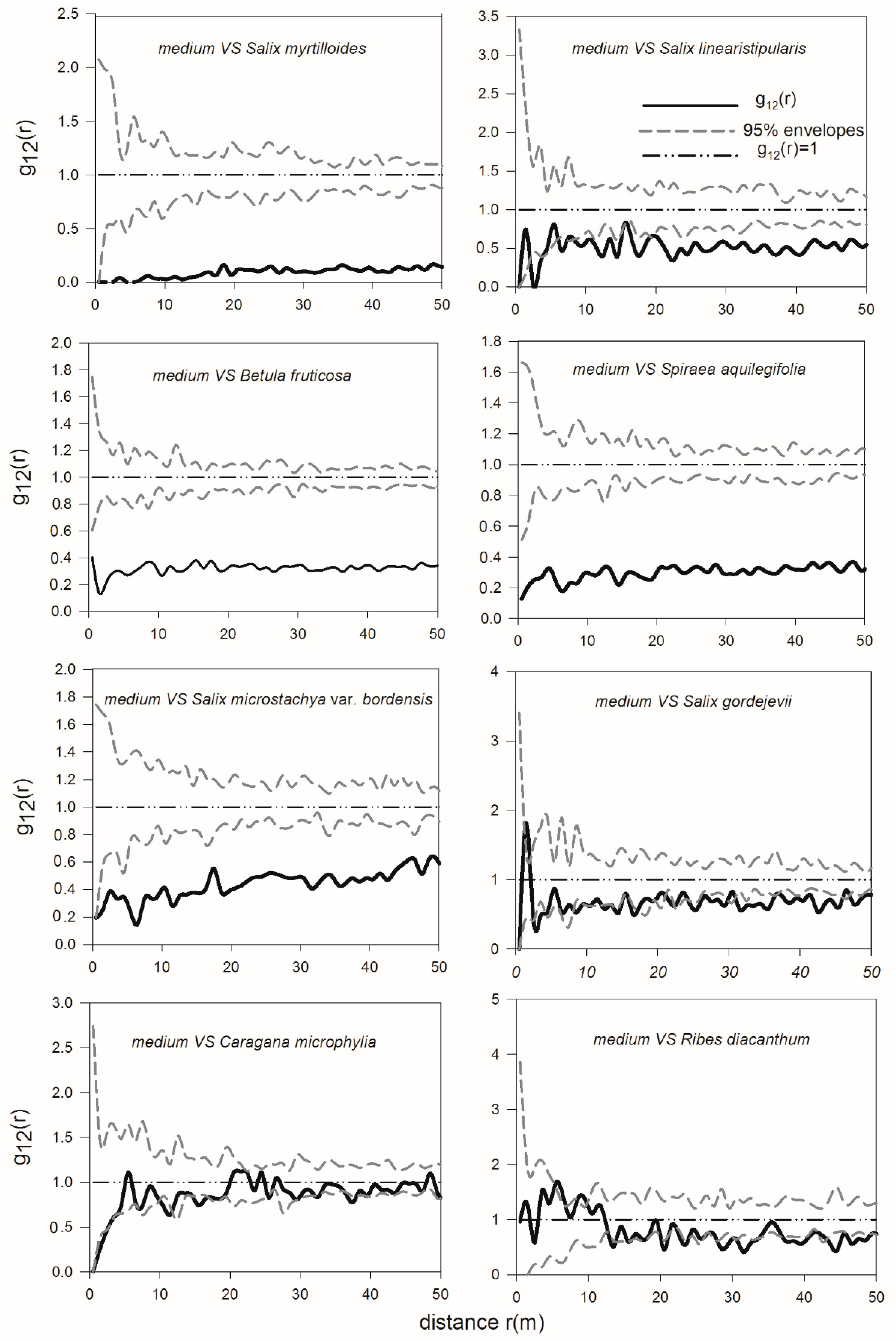
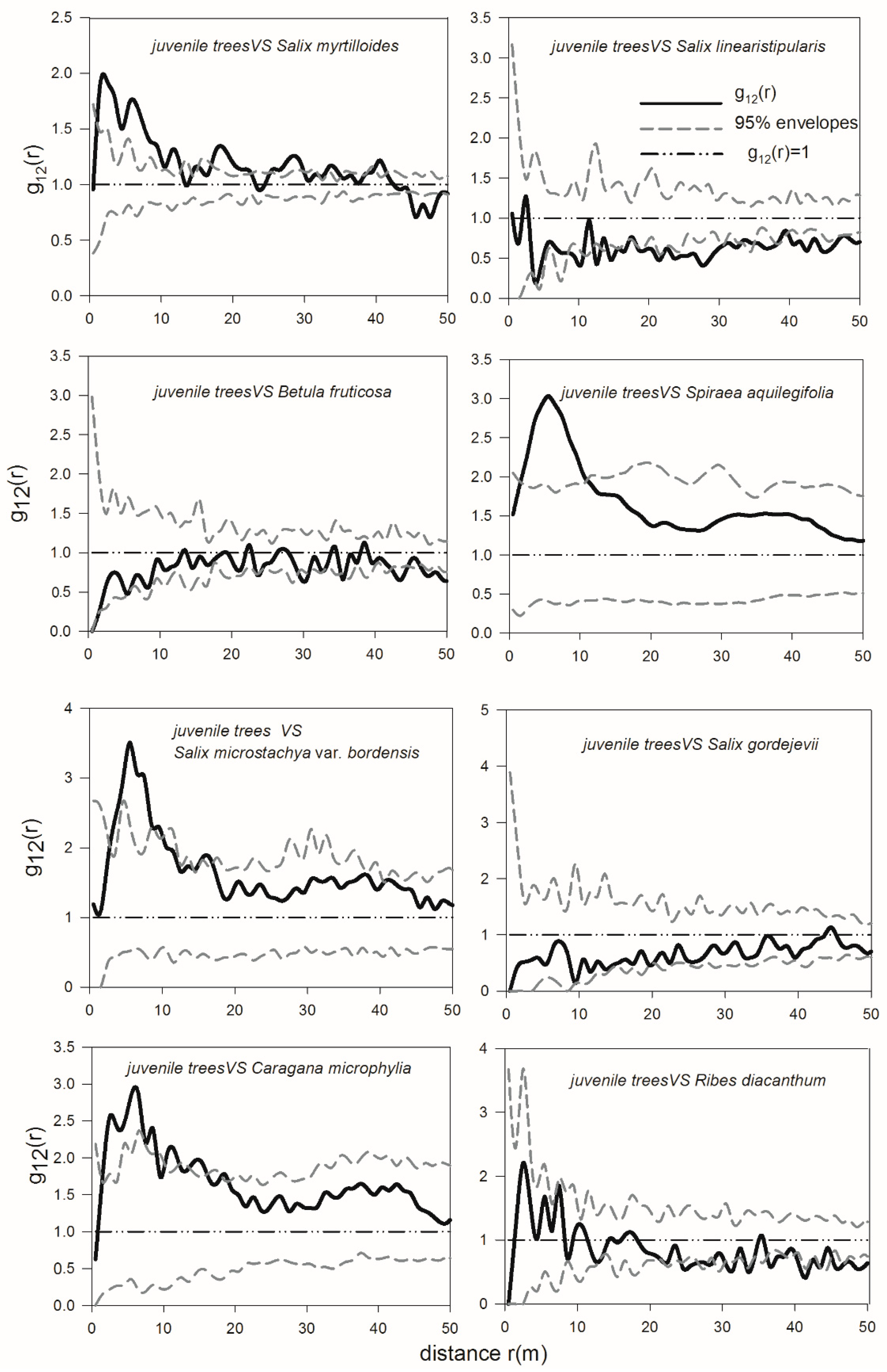
3.4. Spatial Interactions of Woody Plants—Competition between Shrubs and Adult Trees and Mutualism between Dominant Shrub Species and Juvenile Trees
4. Discussion
4.1. Spatial Patterns of U. pumila Trees and Their Mechanism of Formation in the Temperate Savanna-Like Ecosystem
4.2. Spatial Interactions among Trees and Shrubs—Old U. pumila Trees Inhibit the Survival of Juvenile Trees and Shrubs
4.3. The Development of the Original Tree-Dominated Temperate Savanna-Like Ecosystem after Shrub Encroachment
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Archer, S.R.; Andersen, E.M.; Predick, K.I.; Schwinning, S.; Woods, S.R. Woody Plant Encroachment: Causes and Consequences; Rangeland Systems; Springer: Cham, Switzerland, 2017; ISBN 978-3-319-46707-8. [Google Scholar]
- Stevens, N.; Lehmann, C.E.R.; Murphy, B.P.; Durigan, G. Savanna woody encroachment is widespread across three continents. Glob. Chang. Biol. 2017, 23, 235–244. [Google Scholar] [CrossRef] [Green Version]
- Naito, A.T.; Cairns, D.M. Patterns and processes of global shrub expansion. Prog. Phys. Geogr. 2011, 35, 423–442. [Google Scholar] [CrossRef]
- Ravi, S.; D’Odorico, P.; Wang, L.; White, C.S.; Okin, G.S.; Macko, S.A.; Collins, S.L. Post-fire resource redistribution in desert grasslands: A possible negative feedback on land degradation. Ecosystems 2009, 12, 434–444. [Google Scholar] [CrossRef]
- Ratajczak, Z.; Nippert, J.B.; Collins, S.L. Woody encroachment decreases diversity across North American grasslands and savannas. Ecology 2012, 93, 697–703. [Google Scholar] [CrossRef] [PubMed]
- Grover, H.D.; Musick, H.B. Shrubland encroachment in southern new Mexico, USA.: An analysis of desertification processes in the american southwest. Clim. Chang. 1990, 17, 305–330. [Google Scholar] [CrossRef]
- D’Odorico, P.; Okin, G.S.; Bestelmeyer, B.T. A synthetic review of feedbacks and drivers of shrub encroachment in arid grasslands. Ecohydrology 2012, 5, 520–530. [Google Scholar] [CrossRef]
- Van Auken, O.W.; Bush, J. Invasion of Woody Legumes; Springer: Berlin/Heidelberg, Germany, 2013; p. 67. [Google Scholar]
- Tölgyesi, C.; Bátori, Z.; Gallé, R.; Urák, I.; Hartel, T. Shrub encroachment under the trees diversifies the herb layer in a Romanian silvopastoral system. Rangel. Ecol. Manag. 2017, 71, 571–577. [Google Scholar] [CrossRef]
- Tabares, X.; Zimmermann, H.; Dietze, E.; Ratzmann, G.; Herzschuh, U. Vegetation state changes in the course of shrub encroachment in an african savanna since about 1850ce and their potential drivers. Ecol. Evol. 2020, 10, 962–979. [Google Scholar] [CrossRef]
- Li, Y.; Chen, J.; Cui, J.; Zhao, X.; Zhang, T. Nutrient resorption in caragana microphylla along a chronosequence of plantations: Implications for desertified land restoration in north china. Ecol. Eng. 2013, 53, 299–305. [Google Scholar] [CrossRef]
- Devine, A.P.; McDonald, R.A.; Quaife, T.; Maclean, I.M.D. Determinants of woody encroachment and cover in African savannas. Oecologia 2017, 183, 939–951. [Google Scholar] [CrossRef] [Green Version]
- Eldridge, D.J.; Bowker, M.A.; Maestre, F.T.; Roger, E.; Reynolds, J.F.; Whitford, W.G. Impacts of shrub encroachment on ecosystem structure and functioning: Towards a global synthesis. Ecol. Lett. 2011, 14, 709–722. [Google Scholar] [CrossRef] [PubMed]
- Erdős, L.; Cserhalmi, D.; Bátori, Z.; Kiss, T.; Morschhauser, T.; Benyhe, B.; Dénes, A. Shrub encroachment in a wooded-steppe mosaic: Combining GIS methods with landscape historical analysis. Appl. Ecol. Environ. Res. 2013, 11, 371–384. [Google Scholar] [CrossRef]
- Hilbig, W. The Vegetation of Mongolia; SPB Academic Publishing: Amsterdam, The Netherlands, 1995; 246p. [Google Scholar]
- Fang, J.; Wang, Z.; Tang, Z. Atlas of Woody Plants in China: Distribution and Climate; Springer: Berlin/Heidelberg, Germany, 2011; 2020p. [Google Scholar]
- Fu, L.; Xin, Y. Elms of China. In The Elms: Breeding, Conservation, and Disease Management; Dunn, C.P., Ed.; Kluwer Academic Publishers: New York, NY, USA, 2000; 384p. [Google Scholar]
- Zhang, X.S. Vegetation of China and Its Geographic Patterns; Geological publishing House: Beijing, China, 2007; 1175p. [Google Scholar]
- Zhao, H.-L.; Zhou, R.-L.; Su, Y.-Z.; Zhang, H.; Zhao, L.-Y.; Drake, S. Shrub facilitation of desert land restoration in the Horqin Sand Land of Inner Mongolia. Ecol. Eng. 2007, 31, 1–8. [Google Scholar] [CrossRef]
- Wigley, B.J.; Bond, W.J.; Hoffman, M.T. Thicket expansion in a south african savanna under divergent land use: Local vs. global drivers? Glob. Chang. Biol. 2010, 16, 964–976. [Google Scholar] [CrossRef]
- Venter, Z.S.; Cramer, M.D.; Hawkins, H.-J. Drivers of woody plant encroachment over africa. Nat. Commun. 2018, 9, 1–7. [Google Scholar] [CrossRef] [Green Version]
- O’Connor, R.C.; Taylor, J.H.; Nippert, J.B. Browsing and fire decreases dominance of a resprouting shrub in woody encroached grassland. Ecology 2020, 101, e02935. [Google Scholar] [CrossRef] [Green Version]
- Liu, F.; Wu, X.; Bai, E.; Boutton, T.W.; Archer, S.R. Spatial scaling of ecosystem c and n in a subtropical savanna landscape. Glob. Chang. Biol. 2010, 16, 2213–2223. [Google Scholar] [CrossRef]
- Nie, W.; Yuan, Y.; Kepner, W.; Erickson, C.; Jackson, M. Hydrological impacts of mesquite encroachment in the upper san pedro watershed. J. Arid Environ. 2012, 82, 147–155. [Google Scholar] [CrossRef]
- Li, L.; He, W.; Changcun, L.; Deli, W. Vegetation and community changes of elm (ulmus pumila) woodlands in northeastern china in 1983–2011. Chin. Geogr. Sci. 2013, 23, 321–330. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Wang, X.-J.; Liu, G.-H.; Niu, H.-L.; Zhen, P. Dynamics, patterns and structure of major population in ulmus pumila var. sabulosa sparse forest in hunshandake sandland. J. Desert Res. 2009, 29, 508–513. (In Chinese) [Google Scholar]
- Chen, L.; Li, H.; Zhang, P.; Zhao, X.; Zhou, L.; Liu, T.; Hu, H.; Bai, Y.; Shen, H.; Fang, J. Shrub-encroached grassland: A new vegetation type. Chin. J. Nat. 2014, 36, 391–396. (In Chinese) [Google Scholar]
- Su, H.; Li, Y.; Liu, W.; Xu, H.; Sun, O.J. Changes in water use with growth in Ulmus pumila in semiarid sandy land of northern China. Trees 2014, 28, 41–52. [Google Scholar] [CrossRef]
- Dale, M.R.T. Spatial Pattern Analysis in Plant. Ecology; Cambridge University Press: Cambridge, UK, 1999; 340p. [Google Scholar]
- Nathan, R. Long distance dispersal of plants. Science 2006, 313, 786–788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Druckenbrod, D.L.; Shugart, H.H.; Davies, I. Spatial pattern and process in forest stands with in the Virginia piedmont. J. Veg. Sci. 2005, 16, 37–48. [Google Scholar] [CrossRef]
- Wiegand, T.; Moloney, K.A. Handbook of Spatial Point-Pattern Analysis in Ecology; CRC Press: Boca Raton, FL, USA, 2014; 481p. [Google Scholar]
- He, F.; Legendre, P.; La Frankie, J.V. Distribution patterns of tree species in a Malaysian tropical rain forest. J. Veg. Sci. 1997, 8, 105–114. [Google Scholar]
- Condit, R.; Ashton, P.S.; Baker, P.; Bunyavejchewin, S.; Gunatilleke, S.; Gunatilleke, N.; Hubbell, S.P.; Foster, R.B.; Itoh, A.; LaFrankie, J.V.; et al. Spatial patterns in the distribution of tropical tree species. Science 2000, 288, 1414–1418. [Google Scholar] [CrossRef]
- Wang, Y.; Yang, X.; Shi, Z. The formation of the patterns of desert shrub communities on the western Ordos Plateau, China: The roles of seed dispersal and sand burial. PLoS ONE 2013, 8, e69970. [Google Scholar] [CrossRef]
- Wu, B.; Yang, H. Spatial Patterns and Natural Recruitment of Native Shrubs in a Semi-arid Sandy Land. PLoS ONE 2013, 8, e58331. [Google Scholar] [CrossRef]
- Chun, X.; Yong, M.; Liu, J.; Liang, W. Monitoring land cover change and its dynamic mechanism on the qehan lake basin, inner mongolia, north china, during 1977–2013. Environ. Monit. Asses. 2018, 190, 205. [Google Scholar] [CrossRef]
- Bai, Y.F.; Wu, J.G.; Xing, Q.; Pan, Q.M.; Huang, J.H.; Yang, D.L.; Han, X.G. Primary production and rain use efficiency across a precipitation gradient on the Mongolia plateau. Ecology 2008, 89, 2140–2153. [Google Scholar] [CrossRef]
- Editorial Committee of Flora of China. Flora of China, Revised ed.; Scientific Press: Beijing, China, 2013. [Google Scholar]
- Li, S.-L.; Yu, F.-H.; Werger, M.J.A.; Dong, M.; Ramula, S.; Zuidema, P.A. Understanding the effects of a new grazing policy: The impact of seasonal grazing on shrub demography in the inner mongolian steppe. J. Appl. Ecol. 2013, 50, 1377–1386. [Google Scholar] [CrossRef]
- Condit, R. Research in large, long-term tropical forest plots. Trends Ecol. Evol. 1995, 10, 18–22. [Google Scholar] [CrossRef]
- Yu, P. Restoring Degraded Ecosystem in Hunshandak Sand Land through Nature Reserve. Ph.D. Thesis, Chinese Academy of Science, Beijing, China, 2005; 114p. [Google Scholar]
- Stoyan, D.; Stoyan, H. Improving ratio estimators of second order point process characteristics. Scand. J. Stat. 2000, 27, 641–656. [Google Scholar] [CrossRef]
- Wiegand, T.; Moloney, K.A. Rings, circles and null-models for point pattern analysis in ecology. Oikos 2004, 104, 209–229. [Google Scholar] [CrossRef]
- Illian, J.; Penttinen, A.; Stoyan, H.; Stoyan, D. (Eds.) Statistical Analysis and Modelling of Spatial Point Patterns; John Wiley & Sons: Chichester, UK, 2008; 560p. [Google Scholar]
- Kim, D.; Campbell, J.J.N. Effects of tree size, shade tolerance, and spatial pattern on the mortality of woody plants in a seminatural forest in central kentucky. Prof. Geogr. 2016, 68, 436–450. [Google Scholar] [CrossRef]
- Skarpe, C. Spatial patterns and dynamics of woody vegetation in an arid savanna. J. Veg. Sci. 2010, 2, 565–572. [Google Scholar] [CrossRef]
- Landry, S.; St-Laurent, M.H.; Nelson, P.R.; Pelletier, G.; Villard, M.A. Canopy cover estimation from landsat images: Understory impact on top-of-canopy reflectance in a northern hardwood forest. Can. J. Remote Sens. 2018, 44, 435–446. [Google Scholar] [CrossRef]
- Sitch, S.; Smith, B.; Prentice, I.C.; Arneth, A.; Bondeau, A.; Cramer, W.; Kaplan, J.O.; Levis, S.; Lucht, W.; Sykes, M.T.; et al. Evaluation of ecosystem dynamics, plant geography and terrestrial carbon cycling in the lpj dynamic global vegetation model. Glob. Chang. Biol. 2003, 9, 161–185. [Google Scholar] [CrossRef]
- Hartmann, H. Will a 385 million year-struggle for light become a struggle for water and for carbon?–How trees may cope with more frequent climate change-type drought events. Glob. Chang. Biol. 2011, 17, 642–655. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhang, B.; Zhang, X.; Yang, X.; Shi, Z.; Liu, Y. Grazing altered the pattern of woody plants and shrub encroachment in a temperate savanna ecosystem. Int. J. Environ. Res. Public Health 2019, 16, 330. [Google Scholar] [CrossRef] [Green Version]
- Guo, K.; Liu, H. A comparative researches on the development of elm seedlings in four habitats in the Otindag Sandland, Inner Mongolia, China. Acta Ecol. Sin. 2004, 24, 2024–2028. (In Chinese) [Google Scholar]
- Wang, X.; Hu, C.; Li, G.; Zuo, H. Analysis of the factors affecting seed disperal and seedling survival rate of Ulmus pumila in the Otindag sandy land. Arid Zone Res. 2011, 28, 542–547. (In Chinese) [Google Scholar]
- Dai, J.; Liu, H.; Wang, Y.; Guo, Q.; Hu, T.; Quine, T.; Green, S.; Hartmann, H.; Xu, C.; Liu, X.; et al. Drought-modulated allometric patterns of trees in semi-arid forests. Commun. Biol. 2020, 405, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhang, B.; Zhang, K.; Zhou, J.; Ahmad, B. The spatial pattern and interactions of woody plants on the temperate savanna of inner mongolia, china: The effects of alternating seasonal grazing-mowing regimes. PLoS ONE 2015, 10, e0133277. [Google Scholar] [CrossRef] [Green Version]
- Qi, J.; Chen, J.; Wan, S.; Ai, L. Understanding the coupled natural and human systems in dryland east asia. Environ. Res. Lett. 2012, 7, 15202–15207. [Google Scholar] [CrossRef] [Green Version]
- Priyadarshini, K.V.R.; Prins, H.H.T.; De Bie, S.; Heitkönig, I.M.; Woodborne, S.; Gort, G.; Kirkman, K.P.; Ludwig, F.; Dawson, T.E.; De Kroon, H. Seasonality of hydraulic redistribution by trees to grasses and changes in their water-source use that change tree-grass interactions. Ecohydrology 2015, 9, 217–228. [Google Scholar] [CrossRef]
- Stumpp, M.; Wesche, K.; Retzer, V.; Miehe, G. Impact of grazing livestock and distance from water source on soil fertility in southern mongolia. Mt. Res. Dev. 2005, 25, 244–251. [Google Scholar] [CrossRef] [Green Version]
- Anthony, E.; Bernard, B.; Henry, M.M.; Paul, N. Piosphere syndrome and rangeland degradation in Karamoja sub-region, Uganda. Resour. Environ. 2015, 5, 73–89. [Google Scholar] [CrossRef]
- Zhang, Z. Spatial Distribution of Vegetation and Soil in Temperate Savanna Ecosystem, Inner Mongolia. Ph.D. Thesis, Chinese Academy Forestry, Beijing, China, 2017; 137p. [Google Scholar]
- Dohn, J.; Augustine, D.J.; Hanan, N.P.; Ratnam, J.; Sankaran, M. Spatial vegetation patterns and neighborhood competition among woody plants in an east african savanna. Ecology 2016, 98, 478. [Google Scholar] [CrossRef] [Green Version]
- Mckinley, D.C.; Blair, J.M. Woody plant encroachment by juniperus virginiana in a mesic native grassland promotes rapid carbon and nitrogen accrual. Ecosystems 2008, 11, 454–468. [Google Scholar] [CrossRef]
- Schlesinger, W.H.; Raikes, J.A.; Hartley, A.E.; Cross, A.F. On the spatial pattern of soil nutrients in desert ecosystems. Ecology 1996, 77, 364–374. [Google Scholar] [CrossRef] [Green Version]
- Dulamsuren, C.; Hauck, M.; Nyambayar, S. Establishment of Ulmus pumila seedlings on steppe slopes of the northern Mongolian mountain taiga. Acta Oecol. 2009, 35, 563–572. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Category | Number | Height (m) | Maximum Crown Radius (m) | Minimum Crown Radius (m) |
|---|---|---|---|---|---|
| U. pumila trees | Old | 750 | 8.4 ± 0.94 | 6.2 ± 0.83 | 5.0 ± 1.53 |
| Medium | 250 | 5.3 ± 0.93 | 4.1 ± 0.25 | 3.2 ± 1.27 | |
| Juvenile | 400 | 1.7 ± 0.30 | 1.5 ± 0.30 | 0.4 ± 0.22 | |
| Shrubs | Salix linearistipularis | 292 | 1.0 ± 0.13 | 1.1 ± 0.28 | 0.4 ± 0.10 |
| S. microstachya var. bordensis | 547 | 0.50 ± 0.17 | 0.4 ± 0.15 | 0.20 ± 0.35 | |
| S. gordejevii | 238 | 1.30 ± 0.22 | 0.6 ± 0.11 | 0.40 ± 0.27 | |
| S. myrtilloides | 837 | 1.20 ± 0.31 | 0.7 ± 0.13 | 0.4 ± 0.34 | |
| Betula fruticosa | 2417 | 1.0 ± 0.14 | 0.5 ± 0.18 | 0.3 ± 0.10 | |
| Ribes diacanthum | 168 | 0.8 ± 0.16 | 0.6 ± 0.23 | 0.1 ± 0.11 | |
| Caragana microphylla | 414 | 0.3 ± 0.12 | 0.4 ± 0.23 | 0.35 ± 0.14 | |
| Spiraea aquilegifolia | 1038 | 0.6 ± 0.15 | 0.8 ± 0.14 | 0.6 ± 0.12 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, X.; Jiang, L.; Yang, X.; Shi, Z.; Yu, P. Does Shrub Encroachment Indicate Ecosystem Degradation? A Perspective Based on the Spatial Patterns of Woody Plants in a Temperate Savanna-Like Ecosystem of Inner Mongolia, China. Forests 2020, 11, 1248. https://0-doi-org.brum.beds.ac.uk/10.3390/f11121248
Wang X, Jiang L, Yang X, Shi Z, Yu P. Does Shrub Encroachment Indicate Ecosystem Degradation? A Perspective Based on the Spatial Patterns of Woody Plants in a Temperate Savanna-Like Ecosystem of Inner Mongolia, China. Forests. 2020; 11(12):1248. https://0-doi-org.brum.beds.ac.uk/10.3390/f11121248
Chicago/Turabian StyleWang, Xiao, Lina Jiang, Xiaohui Yang, Zhongjie Shi, and Pengtao Yu. 2020. "Does Shrub Encroachment Indicate Ecosystem Degradation? A Perspective Based on the Spatial Patterns of Woody Plants in a Temperate Savanna-Like Ecosystem of Inner Mongolia, China" Forests 11, no. 12: 1248. https://0-doi-org.brum.beds.ac.uk/10.3390/f11121248