
Enhanced Summer Planting Survival of Japanese Larch Container-Grown Seedlings

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Irrigation and Potassium Fertilizer Treatments
2.3. Root Membrane Integrity and Leaf Water Relations
2.4. Application of Antitranspirant and Topping at Planting
2.5. Summer Planting Experiment
2.6. Leaf Gas Exchange
2.7. Statistical Analysis
3. Results
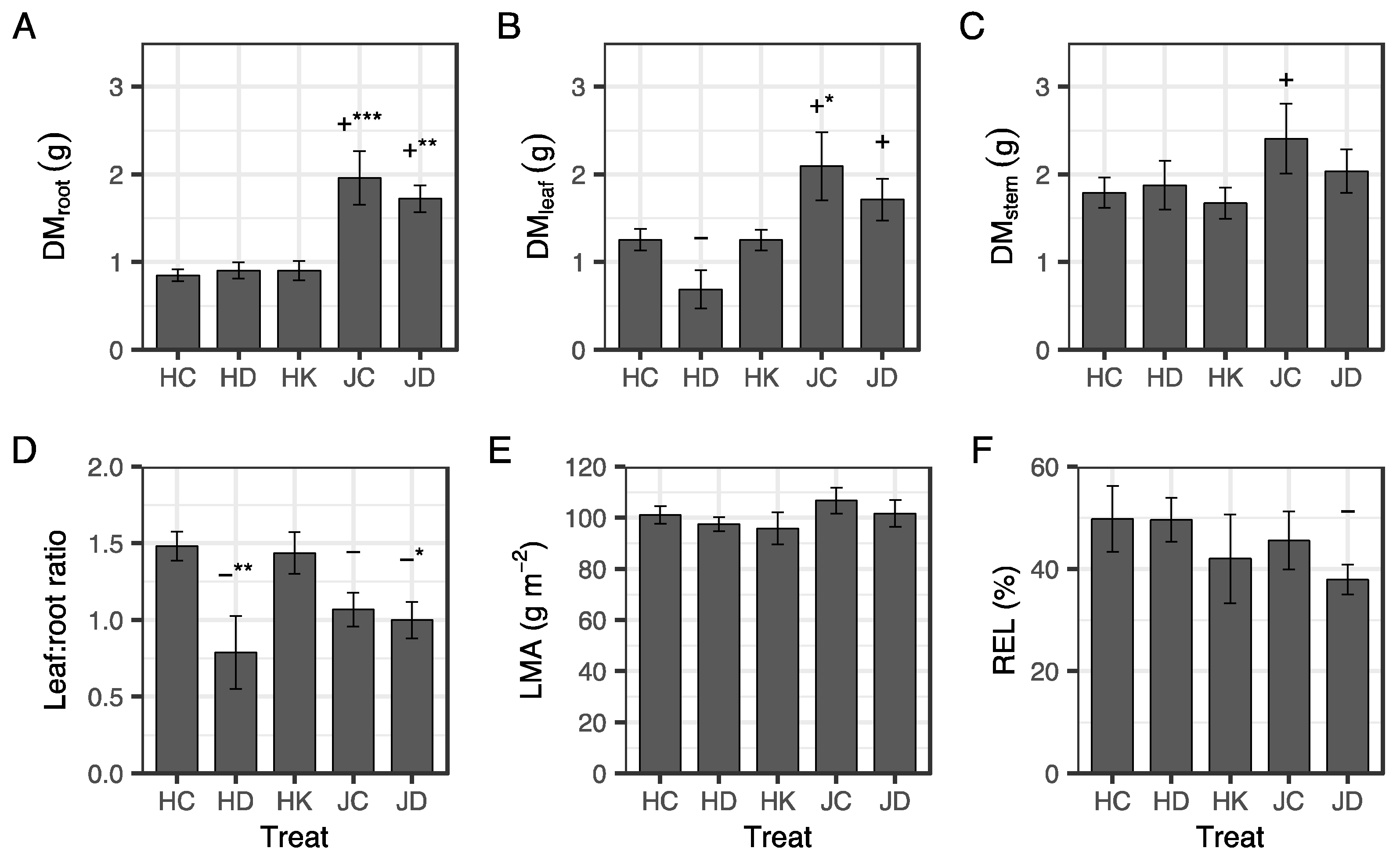
3.1. Dry Mass and Relative Electrolyte Leakage from Fine Roots after the Two Month Nursery Treatment
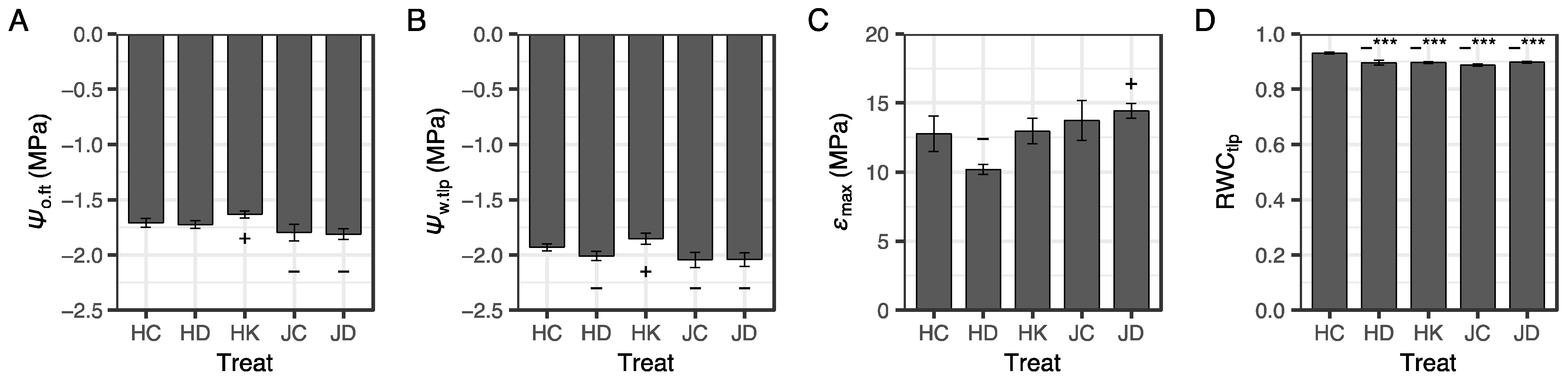
3.2. Leaf Water Relations after Two Months of Nursery Treatment
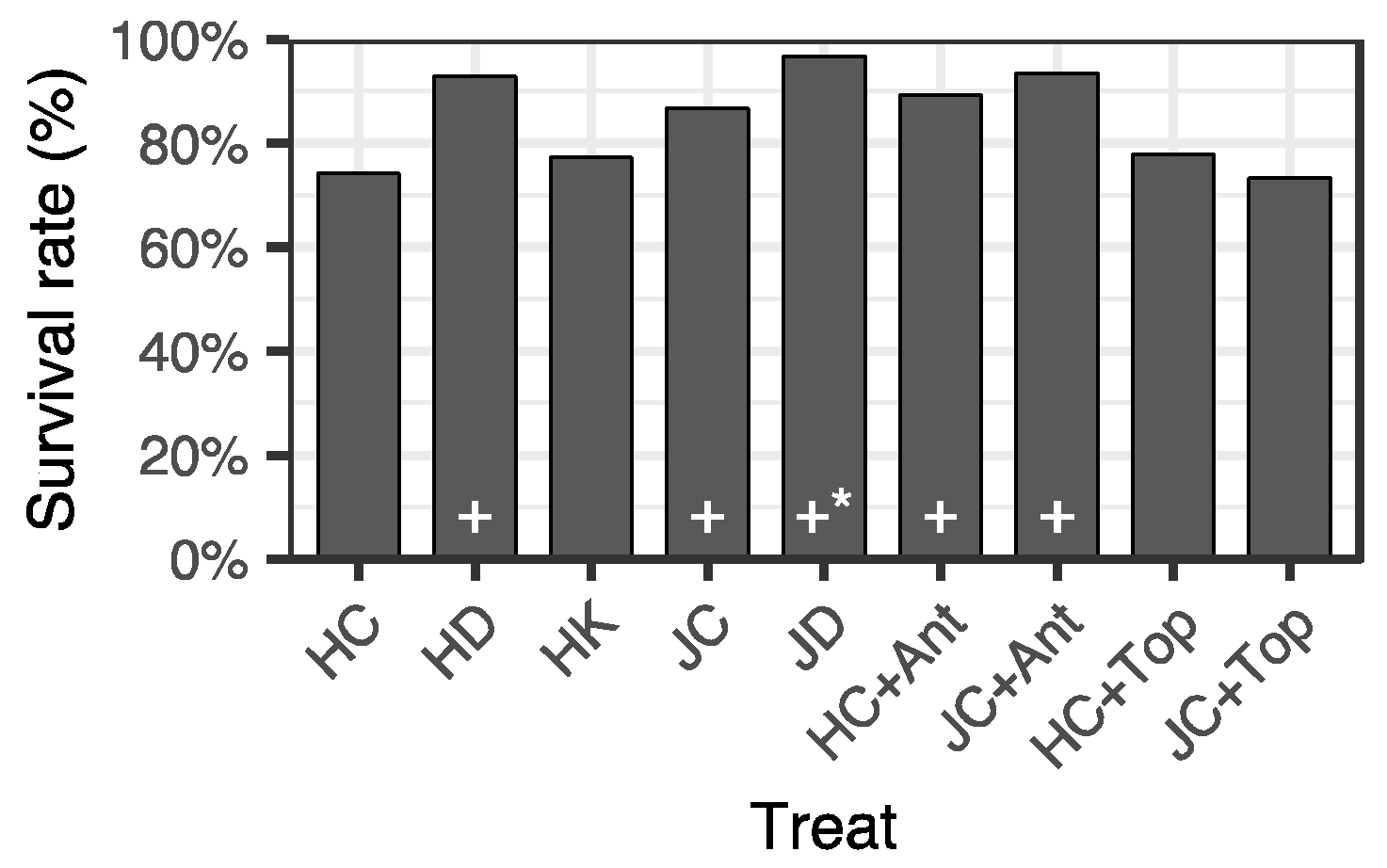
3.3. Survival after Summer Planting
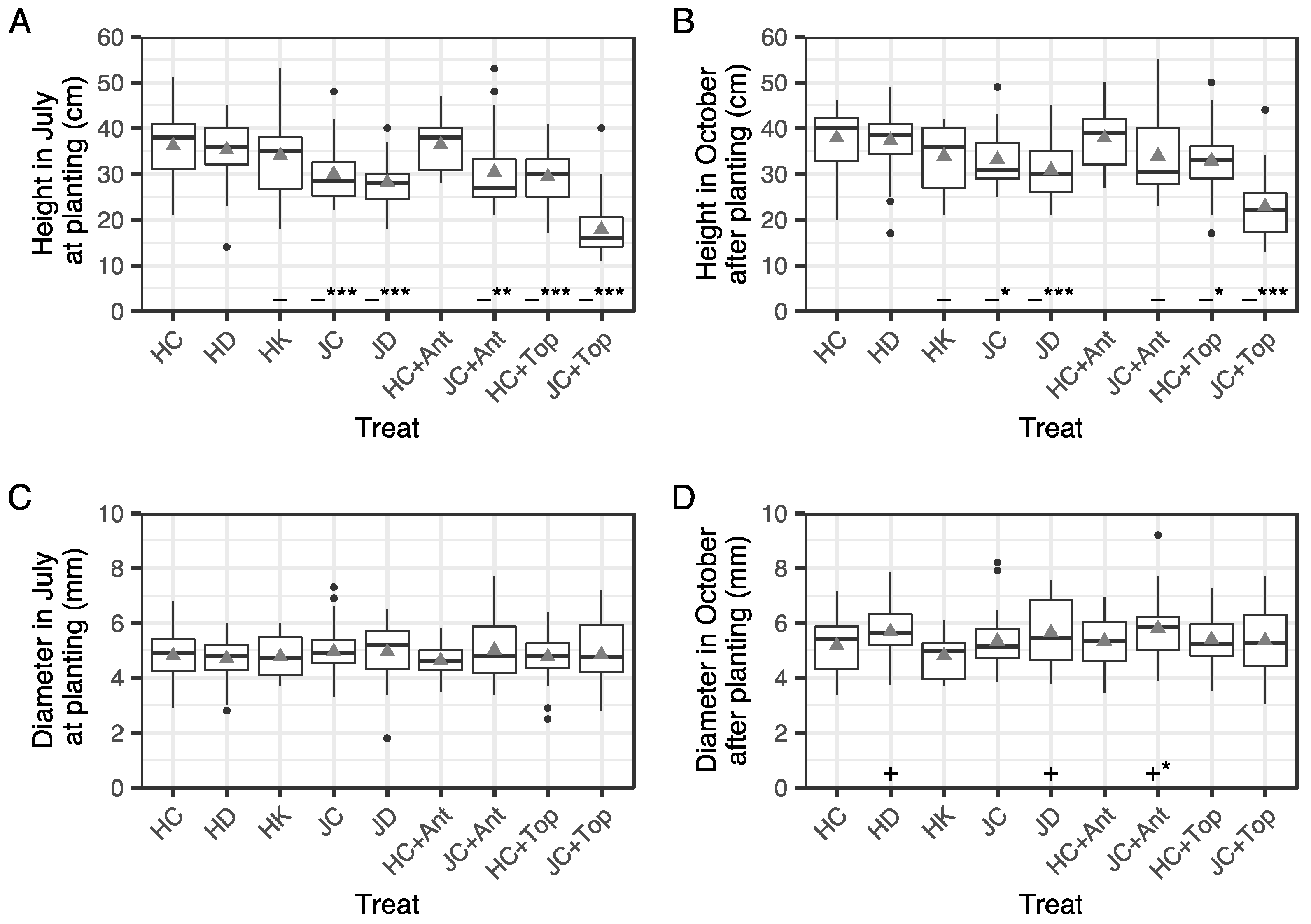
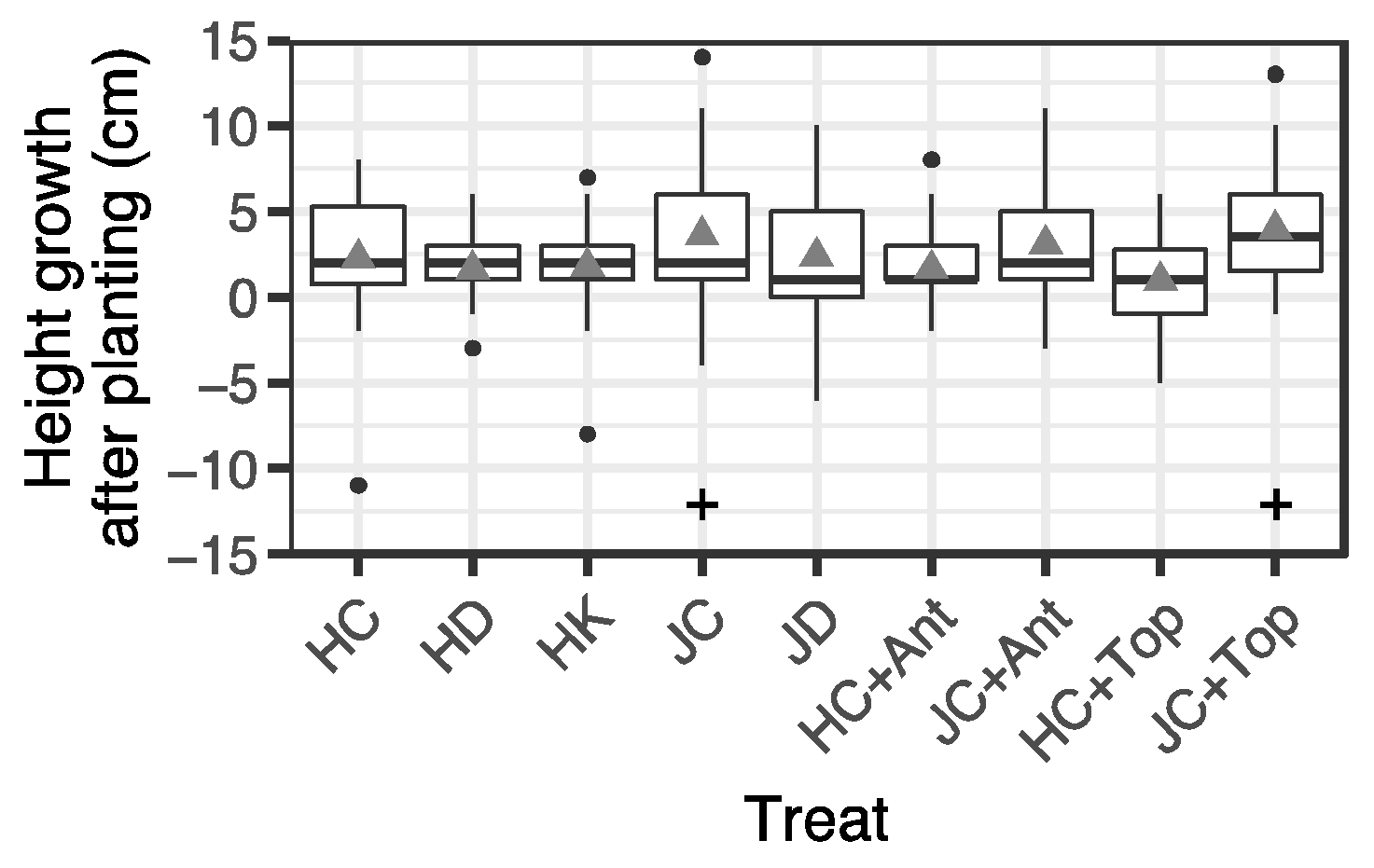
3.4. Height and Root Collar Diameter of Seedlings before and after Summer Planting
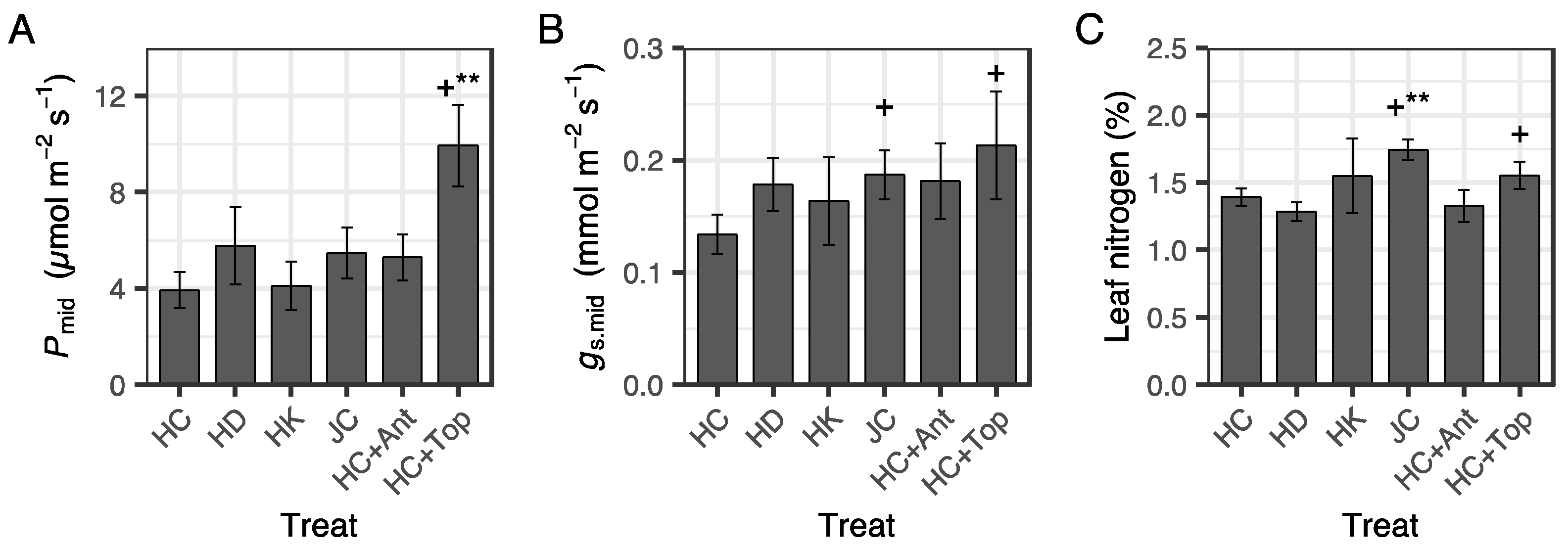
3.5. Gas Exchange after Summer Planting
4. Discussion
4.1. Effects of Irrigation and K Fertilization for Two Months after Winter Storage
4.2. Effects of Antitranspirant and Topping at Summer Planting
4.3. Effect of Container Type
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mattsson, A. Predicting field performance using seedling quality assessment. New For. 1997, 13, 227–252. [Google Scholar] [CrossRef]
- Grossnickle, S.C. Why seedlings survive: Influence of plant attributes. New For. 2012, 43, 711–738. [Google Scholar] [CrossRef]
- Grossnickle, S.C.; MacDonald, J.E. Why seedlings grow: Influence of plant attributes. New For. 2018, 49, 1–34. [Google Scholar] [CrossRef]
- Radoglou, K.; Raftoyannis, Y. The impact of storage, desiccation and planting date on seedling quality and survival of woody plant species. Forestry 2002, 75, 179–190. [Google Scholar] [CrossRef] [Green Version]
- Thiffault, N.; Jobidon, R.; Munson, A.D. Comparing large containerized and bareroot conifer stock on sites of contrasting vegetation composition in a non-herbicide scenario. New For. 2014, 45, 875–891. [Google Scholar] [CrossRef]
- Mason, W.L.; Biggin, P. Comparative performance of containerised and bare-root sitka spruce and lodgepole pine seedlings in upland Britain. Forestry 1988, 61, 149–163. [Google Scholar] [CrossRef]
- Grossnickle, S.C.; El-Kassaby, Y.A. Bareroot versus container stocktypes: A performance comparison. New For. 2016, 47, 1–51. [Google Scholar] [CrossRef]
- Paul Jackson, D.; Kasten Dumroese, R.; Barnett, J.P. Nursery response of container Pinus palustris seedlings to nitrogen supply and subsequent effects on outplanting performance. For. Ecol. Manag. 2012, 265, 1–12. [Google Scholar] [CrossRef]
- Duryea, M.L. Nursery cultural practices: Impacts on seedling quality. In Forestry Nursery Manual: Production of Bareroot Seedlings; Duryea, M.L., Landis, T.D., Perry, C.R., Eds.; Springer: Dordrecht, The Netherlands, 1984; Volume 11, pp. 143–164. [Google Scholar]
- Leugner, J.; Jurásek, A.; Martincová, J. Comparison of morphological and physiological parameters of the planting material of Norway spruce (Picea abies [L.] Karst.) from intensive nursery technologies with current bareroot plants. J. For. Sci. 2009, 55, 511–517. [Google Scholar] [CrossRef] [Green Version]
- Shi, W.; Grossnickle, S.C.; Li, G.; Su, S.; Liu, Y. Fertilization and irrigation regimes influence on seedling attributes and field performance of Pinus tabuliformis Carr. Forestry 2019, 92, 97–107. [Google Scholar] [CrossRef]
- Pinto, J.R.; Marshall, J.D.; Dumroese, R.K.; Davis, A.S.; Cobos, D.R. Establishment and growth of container seedlings for reforestation: A function of stocktype and edaphic conditions. For. Ecol. Manag. 2011, 261, 1876–1884. [Google Scholar] [CrossRef]
- Allen, C.D.; Breshears, D.D.; McDowell, N.G. On underestimation of global vulnerability to tree mortality and forest die-off from hotter drought in the Anthropocene. Ecosphere 2015, 6, 1–55. [Google Scholar] [CrossRef]
- IPCC. Climate Change 2014 Impacts, Adaptation, and Vulnerability Part A: Global and Sectoral Aspects; Field, C.B., Barros, V.R., Eds.; Cambridge University Press: New York, NY, USA, 2014.
- Japan Forestry Agency. Annual Report on Forest and Forestry in Japan—Fiscal Year 2017. Available online: http://www.rinya.maff.go.jp/j/kikaku/hakusyo/29hakusyo/index.html (accessed on 5 July 2021).
- Masaki, T.; Oguro, M.; Yamashita, N.; Otani, T.; Utsugi, H. Reforestation following harvesting of conifer plantations in Japan: Current issues from silvicultural and ecological perspectives. Reforesta 2017, 3, 125–141. [Google Scholar] [CrossRef] [Green Version]
- Agathokleous, E.; Kitao, M.; Komatsu, M.; Tamai, Y.; Saito, H.; Harayama, H.; Uemura, A.; Tobita, H.; Koike, T. Effects of soil nutrient availability and ozone on container-grown Japanese larch seedlings and role of soil microbes. J. For. Res. 2020, 31, 2295–2311. [Google Scholar] [CrossRef] [Green Version]
- Japan Meteorological Agency. Available online: https://www.data.jma.go.jp/obd/stats/etrn/view/nml_sfc_ym.php?prec_no=14&block_no=47412 (accessed on 5 July 2021).
- Kurinobu, S. Forest tree breeding for Japanese larch. Eurasian J. For. Res. 2005, 8, 127–134. [Google Scholar]
- Da Ronch, F.; Caudullo, G.; Tinner, W.; de Rigo, D. Larix decidua and other larches in Europe: Distribution, habitat, usage and threats. In European Atlas of Forest Tree Species; San-Miguel-Ayanz, J., de Rigo, D., Caudullo, G., Houston Durrant, T., Mauri, A., Eds.; Publication Office of the European Union: Luxembourg, 2016. [Google Scholar]
- Pâques, L.E. A critical review of larch hybridization and its incidence on breeding strategies. Ann. For. Sci. 1989, 46, 141–153. [Google Scholar] [CrossRef] [Green Version]
- Perron, M. A strategy for the second breeding cycle of Larix x marschlinsii in Québec, Canada including experiments to guide interspecific tree breeding programme. Silvae Genet. 2008, 57, 282–291. [Google Scholar] [CrossRef]
- Greenwood, M.S.; Roth, B.E.; Maass, D.D.; Irland, L.C. Near rotation-length performance of selected hybrid larch in Central Maine, USA. Silvae Genet. 2015, 64, 73–80. [Google Scholar] [CrossRef] [Green Version]
- Vanoni, M.; Bugmann, H.; Nötzli, M.; Bigler, C. Drought and frost contribute to abrupt growth decreases before tree mortality in nine temperate tree species. For. Ecol. Manag. 2016, 382, 51–63. [Google Scholar] [CrossRef] [Green Version]
- Levesque, M.; Saurer, M.; Siegwolf, R.; Eilmann, B.; Brang, P.; Bugmann, H.; Rigling, A. Drought response of five conifer species under contrasting water availability suggests high vulnerability of Norway spruce and European larch. Glob. Chang. Biol. 2013, 19, 3184–3199. [Google Scholar] [CrossRef]
- Peng, K.; Peng, J.; Huo, J.; Yang, L. Assessing the adaptability of alien (Larix kaempferi) and native (Pinus armandii) tree species at the Baiyunshan Mountain, central China. Ecol. Indic. 2018, 95, 108–116. [Google Scholar] [CrossRef]
- Harayama, H.; Kita, K.; Kon, H.; Ishizuka, W.; Tobita, H.; Utsugi, H. Effect of planting season on survival rate, growth and ecophysiological properties of container seedlings of Japanese larch (Larix kaempferi). J. Jpn. For. Soc. 2016, 98, 158–166. [Google Scholar] [CrossRef] [Green Version]
- Saiki, S.-T.; Ando, Y.; Yazaki, K.; Tobita, H. Drought hardening contributes to the maintenance of proportions of non-embolized xylem and cambium status during consecutive dry treatment in container-grown seedling of Japanese cedar (Cryptomeria japonica). Forests 2020, 11, 441. [Google Scholar] [CrossRef]
- Sigala, J.A.; Uscola, M.; Oliet, J.A.; Jacobs, D.F. Drought tolerance and acclimation in Pinus ponderosa seedlings: The influence of nitrogen form. Tree Physiol. 2020, 40, 1165–1177. [Google Scholar] [CrossRef] [PubMed]
- Royo, A.; Gil, L.; Pardos, J.A. Effect of water stress conditioning on morphology, physiology and field performance of Pinus halepensis Mill. seedlings. New For. 2001, 21, 127–140. [Google Scholar] [CrossRef]
- Cuesta, B.; Villar-Salvador, P.; Puértolas, J.; Jacobs, D.F.; Rey Benayas, J.M. Why do large, nitrogen rich seedlings better resist stressful transplanting conditions? A physiological analysis in two functionally contrasting Mediterranean forest species. For. Ecol. Manag. 2010, 260, 71–78. [Google Scholar] [CrossRef]
- Wang, L.; Katzensteiner, K.; Schume, H.; Van Loo, M.; Godbold, D.L. Potassium fertilization affects the distribution of fine roots but does not change ectomycorrhizal community structure. Ann. For. Sci. 2016, 73, 691–702. [Google Scholar] [CrossRef] [Green Version]
- Fernández, M.; Royo, A.; Gil, L.; Pardos, J.A. Effects of temperature on growth and stress hardening development of phytotron-grown seedlings of Aleppo pine (Pinus halepensis Mill.). Ann. For. Sci. 2003, 60, 277–284. [Google Scholar] [CrossRef] [Green Version]
- Grzebisz, W.; Gransee, A.; Szczepaniak, W.; Diatta, J. The effects of potassium fertilization on water-use efficiency in crop plants. J. Plant Nutr. Soil Sci. 2013, 176, 355–374. [Google Scholar] [CrossRef]
- Römheld, V.; Kirkby, E.A. Research on potassium in agriculture: Needs and prospects. Plant Soil 2010, 335, 155–180. [Google Scholar] [CrossRef]
- Gu, S.L.; Fuchigami, L.H.; Guak, S.H.; Shin, C. Effects of short-term water stress, hydrophilic polymer amendment, and antitranspirant on stomatal status, transpiration, water loss, and growth in ‘better boy’ tomato plants. J. Am. Soc. Hortic. Sci. 1996, 121, 831–837. [Google Scholar] [CrossRef] [Green Version]
- Colombo, S.J.; Odlum, K.D. Efficacy of six antitranspirants on black spruce container seedlings. New For. 1987, 1, 239–244. [Google Scholar] [CrossRef]
- Del Amor, F.M.; Cuadra-Crespo, P.; Walker, D.J.; Camara, J.M.; Madrid, R. Effect of foliar application of antitranspirant on photosynthesis and water relations of pepper plants under different levels of CO2 and water stress. J. Plant Physiol. 2010, 167, 1232–1238. [Google Scholar] [CrossRef]
- Yamashita, N.; Okuda, S.; Suwa, R.; Lei, T.T.; Tobita, H.; Utsugi, H.; Kajimoto, T. Impact of leaf removal on initial survival and growth of container-grown and bare-root seedlings of Hinoki cypress (Chamaecyparis obtusa). For. Ecol. Manag. 2016, 370, 76–82. [Google Scholar] [CrossRef]
- Zaczek, J.J.; Steiner, K.C.; Bowersox, T.W. Northern red oak planting stock: 6-year results. New For. 1996, 13, 177–191. [Google Scholar] [CrossRef]
- Landis, T.D.; Nisley, R.G. The Container Tree Nursery Manual; US Department of Agriculture, Forest Service: Washington, DC, USA, 1990.
- McKay, H.M. Electrolyte leakage from fine roots of conifer seedlings: A rapid index of plant vitality following cold storage. Can. J. For. Res. 1992, 22, 1371–1377. [Google Scholar] [CrossRef]
- McKay, H. Root electrolyte leakage and root growth potential as indicators of spruce and larch establishment. Silva Fenn. 1998, 32, 241–252. [Google Scholar] [CrossRef] [Green Version]
- Tyree, M.T.; Hammel, H.T. The measurement of the turgor pressure and the water relations of plants by the pressure-bomb technique. J. Exp. Bot. 1972, 23, 267–282. [Google Scholar] [CrossRef]
- Bartlett, M.K.; Klein, T.; Jansen, S.; Choat, B.; Sack, L. The correlations and sequence of plant stomatal, hydraulic, and wilting responses to drought. Proc. Natl. Acad. Sci. USA 2016, 113, 13098–13103. [Google Scholar] [CrossRef] [Green Version]
- Harayama, H.; Ikeda, T.; Ishida, A.; Yamamoto, S.-I. Seasonal variations in water relations in current-year leaves of evergreen trees with delayed greening. Tree Physiol. 2006, 26, 1025–1033. [Google Scholar] [CrossRef] [Green Version]
- Schulte, P.J.; Hinckley, T.M. A comparison of pressure-volume curve data analysis techniques. J. Exp. Bot. 1985, 36, 1590–1602. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; 4.0.3; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Villar-Salvador, P.; Planelles, R.; Oliet, J.; Penuelas-Rubira, J.L.; Jacobs, D.F.; Gonzalez, M. Drought tolerance and transplanting performance of holm oak (Quercus ilex) seedlings after drought hardening in the nursery. Tree Physiol. 2004, 24, 1147–1155. [Google Scholar] [CrossRef] [Green Version]
- Kozlowski, T.T.; Pallardy, S.G. Acclimation and adaptive responses of woody plants to environmental stresses. Bot. Rev. 2002, 68, 270–334. [Google Scholar] [CrossRef]
- Doi, K.; Morikawa, Y.; Hinckley, T.M. Seasonal trends of several water relation parameters in Cryptomeria japonica seedlings. Can. J. For. Res. 1986, 16, 74–77. [Google Scholar] [CrossRef]
- Rhizopoulou, S.; Mitrakos, K. Water relations of evergreen sclerophylls. I. Seasonal changes in the water relations of eleven species from the same environment. Ann. Bot. 1990, 65, 171–178. [Google Scholar] [CrossRef]
- Colombo, S.J.; Teng, Y. Seasonal variation in the tissue water relations of Picea glauca. Oecologia 1992, 92, 410–415. [Google Scholar] [CrossRef] [PubMed]
- Zolfaghari, R.; Rezaei, K.; Fayyaz, P.; Naghiha, R.; Namvar, Z. The effect of indigenous phosphate-solubilizing bacteria on Quercus brantii seedlings under water stress. J. Sustain. For. 2021, 40, 733–747. [Google Scholar] [CrossRef]
- Huang, X.; Lakso, A.N.; Eissenstat, D.M. Interactive effects of soil temperature and moisture on Concord grape root respiration. J. Exp. Bot. 2005, 56, 2651–2660. [Google Scholar] [CrossRef] [Green Version]
- Zorb, C.; Senbayram, M.; Peiter, E. Potassium in agriculture–status and perspectives. J. Plant Physiol. 2014, 171, 656–669. [Google Scholar] [CrossRef]
- Jacobs, D.F.; Salifu, K.F.; Seifert, J.R. Growth and nutritional response of hardwood seedlings to controlled-release fertilization at outplanting. For. Ecol. Manag. 2005, 214, 28–39. [Google Scholar] [CrossRef]
- Hummel, R.L. Water relations of container-grown woody and Herbaceous plants following antitranspirant sprays. HortScience 1990, 25, 772–775. [Google Scholar] [CrossRef] [Green Version]
- Emerson, J.L.; Hildreth, A.C. Preliminary report on reducing transpiration of transplanted evergreens. Science 1933, 77, 433–434. [Google Scholar] [CrossRef]
- Shirley, H.L.; Meuli, L.J. Influence of foliage sprays on drought resistance of conifers. Plant Physiol. 1938, 13, 399–406. [Google Scholar] [CrossRef] [Green Version]
- Mphande, W.; Kettlewell, P.S.; Grove, I.G.; Farrell, A.D. The potential of antitranspirants in drought management of arable crops: A review. Agric. Water Manag. 2020, 236, 106143. [Google Scholar] [CrossRef]
- Helliwell, D.R.; Harrison, A.F. Effects of light and weed competition on the growth of seedlings of four tree species on a range of soil. Q. J. For. 1979, 73, 160–167. [Google Scholar]
- Oester, P.T. Ten-year response of western larch and Douglas-fir seedlings to mulch mats, sulfometuron, and shade in northeast Oregon. Tree Plant. Notes 2009, 53, 29–36. [Google Scholar]
- Harayama, H.; Tsuyama, I.; Kuramoto, S.; Uemura, A.; Kitao, M.; Han, Q.; Yamada, T.; Sasaki, S. Effects of weed competition on the survival and initial growth of planted seedlings of Japanese larch (Larix kaempferi). J. Jpn. For. Soc. 2018, 100, 158–164. [Google Scholar] [CrossRef]
- Smith, P.F. Inhibition of growth in guayule as affected by topping and defoliation. Am. J. Bot. 1944, 31, 328–336. [Google Scholar] [CrossRef]
- Aghai, M.M.; Pinto, J.R.; Davis, A.S. Container volume and growing density influence western larch (Larix occidentalis Nutt.) seedling development during nursery culture and establishment. New For. 2014, 45, 199–213. [Google Scholar] [CrossRef]
- Ritchie, G.A.; Landis, T.D. Assessing plant quality. In The Container Tree Nursery Manual; Landis, T.D., Nisley, R.G., Eds.; RNGR: Houston, TX, USA, 2010; Volume 7, pp. 17–80. [Google Scholar]
- Dominguez-Lerena, S.; Herrero Sierra, N.; Carrasco Manzano, I.; Ocaña Bueno, L.; Peñuelas Rubira, J.L.; Mexal, J.G. Container characteristics influence Pinus pinea seedling development in the nursery and field. For. Ecol. Manag. 2006, 221, 63–71. [Google Scholar] [CrossRef]
- Aphalo, P.; Rikala, R. Field performance of silver-birch planting-stock grown at different spacing and in containers of different volume. New For. 2003, 25, 93–108. [Google Scholar] [CrossRef]
- Rune, G. Slits in container wall improve root structure and stem straightness of outplanted Scots pine seedlings. Silva Fenn. 2003, 37, 333–342. [Google Scholar] [CrossRef] [Green Version]
- Thomas, B.R.; Schreiber, S.G.; Kamelchuk, D.P. Impact of planting container type on growth and survival of three hybrid poplar clones in central Alberta, Canada. New For. 2016, 47, 815–827. [Google Scholar] [CrossRef]
- Matthes-Sears, U.; Larson, D.W. Limitations to seedling growth and survival by the quantity and quality of rooting space: Implications for the establishment of Thuja occidentalis on cliff faces. Int. J. Plant Sci. 1999, 160, 122–128. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Container Type | Watering | Fertilization | Antitranspirant | Topping | Abbreviation | No. of Seedlings |
|---|---|---|---|---|---|---|
| Hiko-V-120 | Control | Control | No | No | HC | 27 |
| Dry | Control | No | No | HD | 28 | |
| Control | High-K | No | No | HK | 23 | |
| Control | Control | Yes | No | HC + Ant | 28 | |
| Control | Control | No | Yes | HC + Top | 27 | |
| JFA-150 | Control | Control | No | No | JC | 30 |
| Dry | Control | No | No | JD | 30 | |
| Control | Control | Yes | No | JC + Ant | 30 | |
| Control | Control | No | Yes | JC + Top | 30 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Harayama, H.; Tobita, H.; Kitao, M.; Kon, H.; Ishizuka, W.; Kuromaru, M.; Kita, K. Enhanced Summer Planting Survival of Japanese Larch Container-Grown Seedlings. Forests 2021, 12, 1115. https://0-doi-org.brum.beds.ac.uk/10.3390/f12081115
Harayama H, Tobita H, Kitao M, Kon H, Ishizuka W, Kuromaru M, Kita K. Enhanced Summer Planting Survival of Japanese Larch Container-Grown Seedlings. Forests. 2021; 12(8):1115. https://0-doi-org.brum.beds.ac.uk/10.3390/f12081115
Chicago/Turabian StyleHarayama, Hisanori, Hiroyuki Tobita, Mitsutoshi Kitao, Hirokazu Kon, Wataru Ishizuka, Makoto Kuromaru, and Kazuhito Kita. 2021. "Enhanced Summer Planting Survival of Japanese Larch Container-Grown Seedlings" Forests 12, no. 8: 1115. https://0-doi-org.brum.beds.ac.uk/10.3390/f12081115