
Climate-Growth Relations of Abies georgei along an Altitudinal Gradient in Haba Snow Mountain, Southwestern China


Abstract
:1. Introduction
2. Materials and Methods
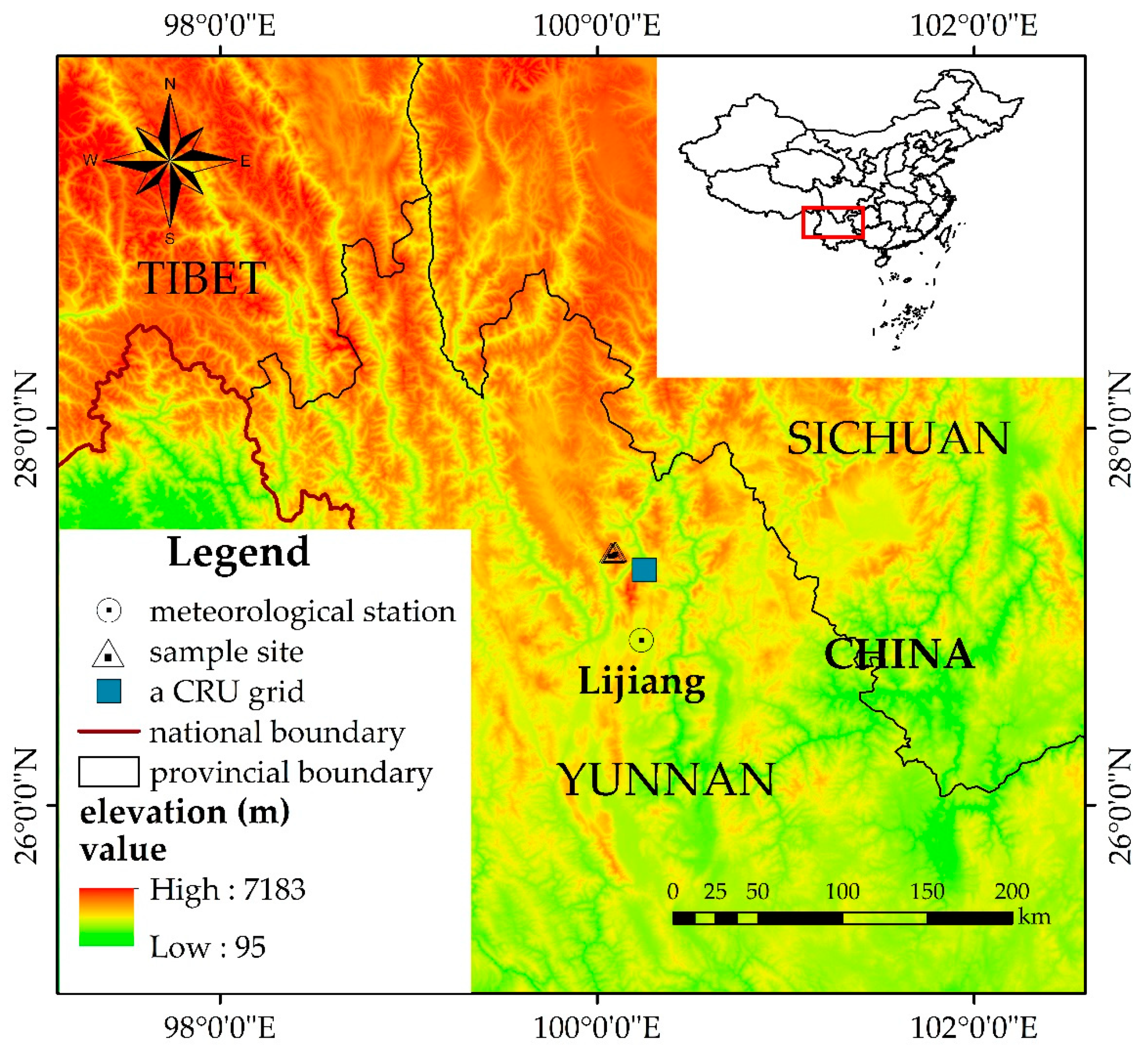
2.1. Study Area
2.2. Tree-Ring Sampling and Chronnology Development
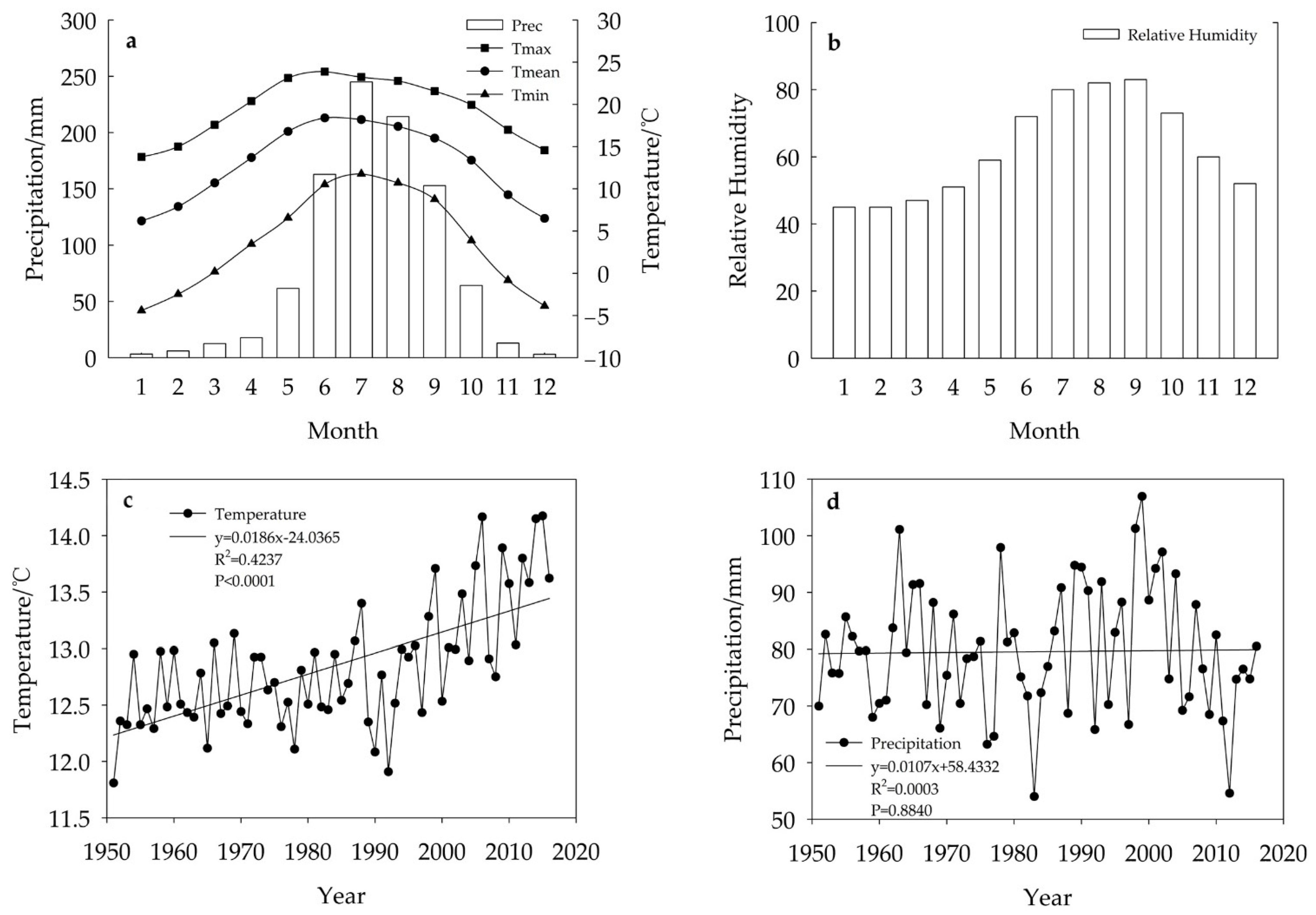
2.3. Climate Data
2.4. Climate-Growth Relationships
3. Results
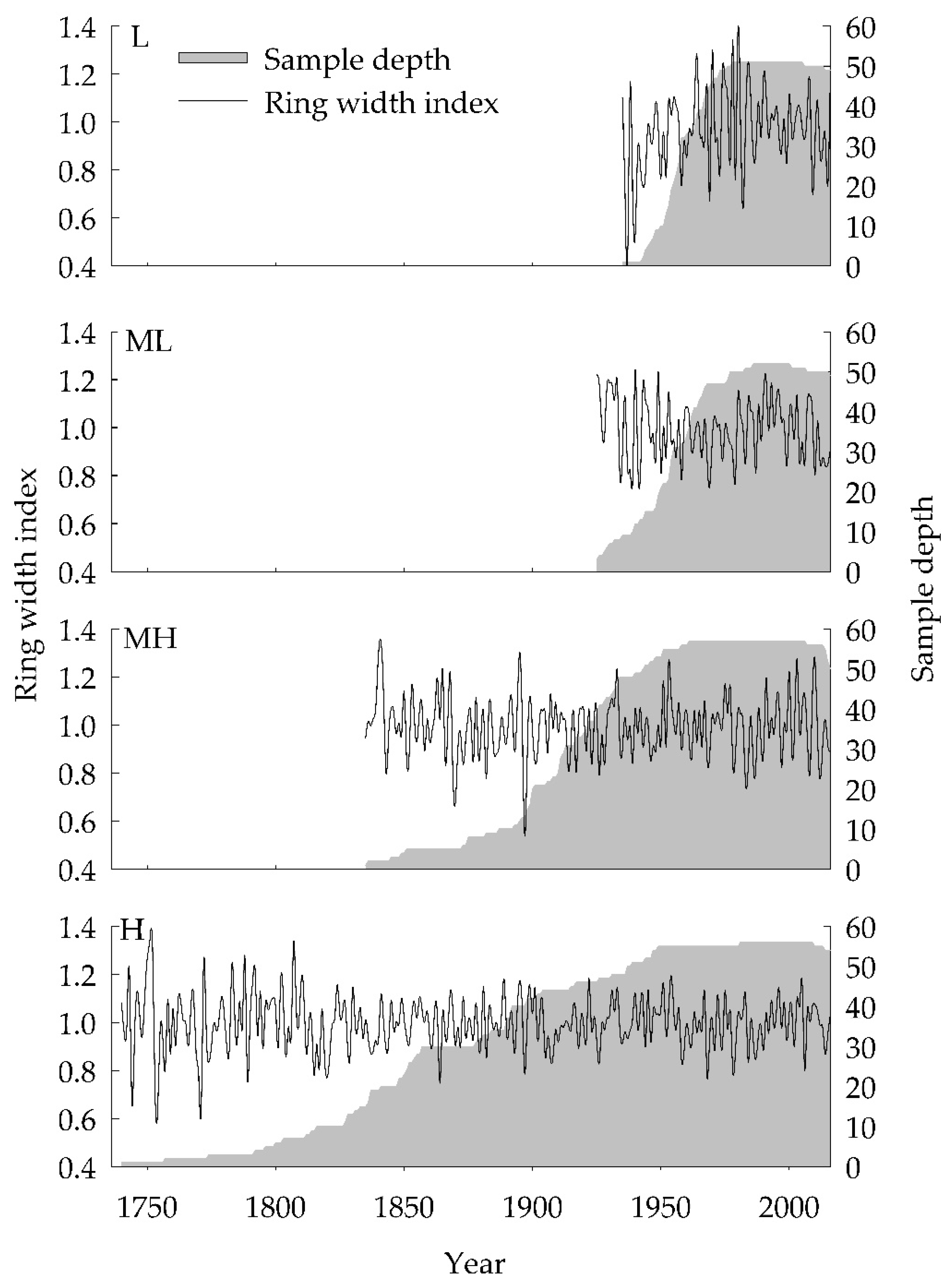
3.1. Tree-Ring Chronologies
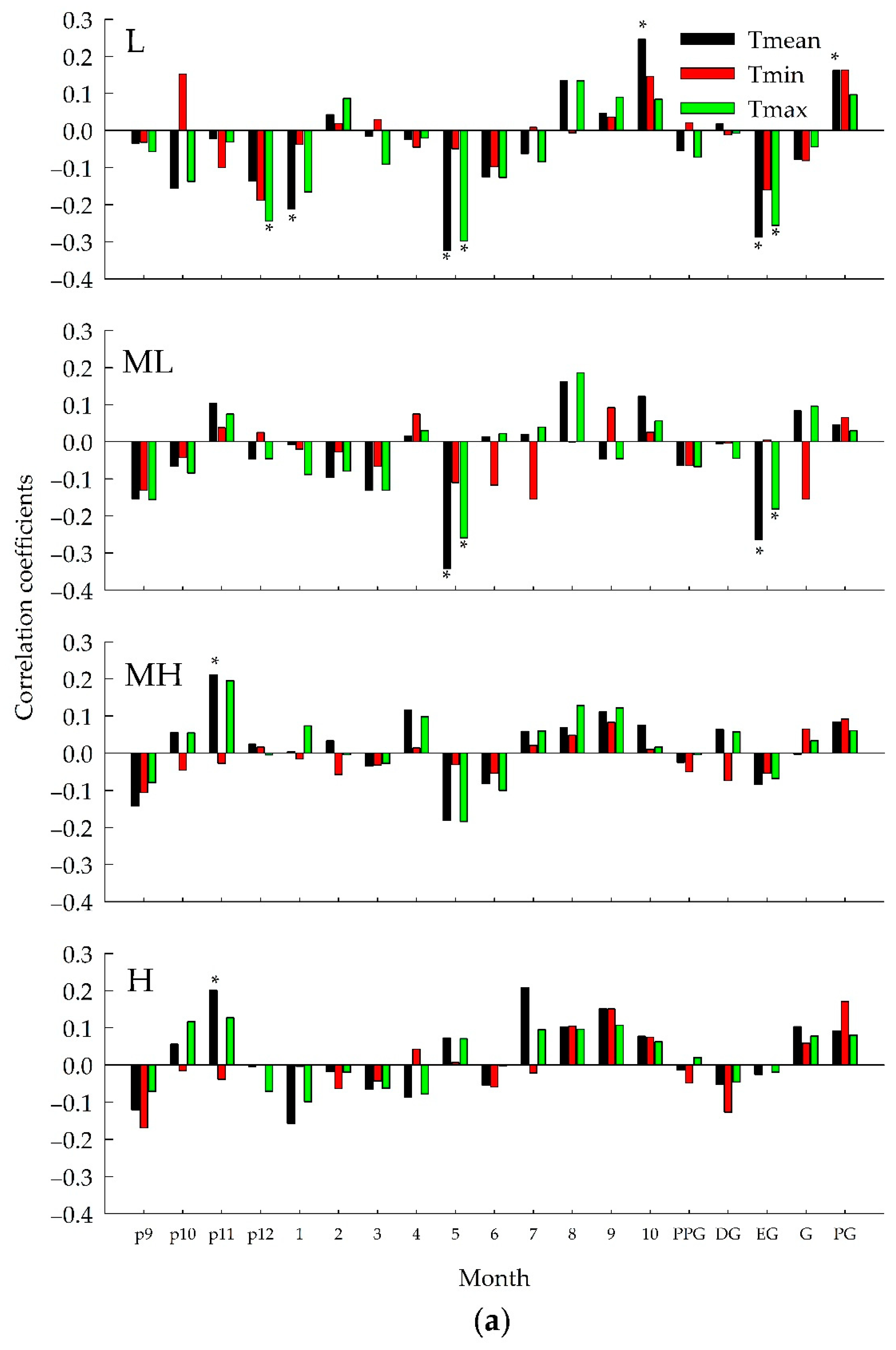
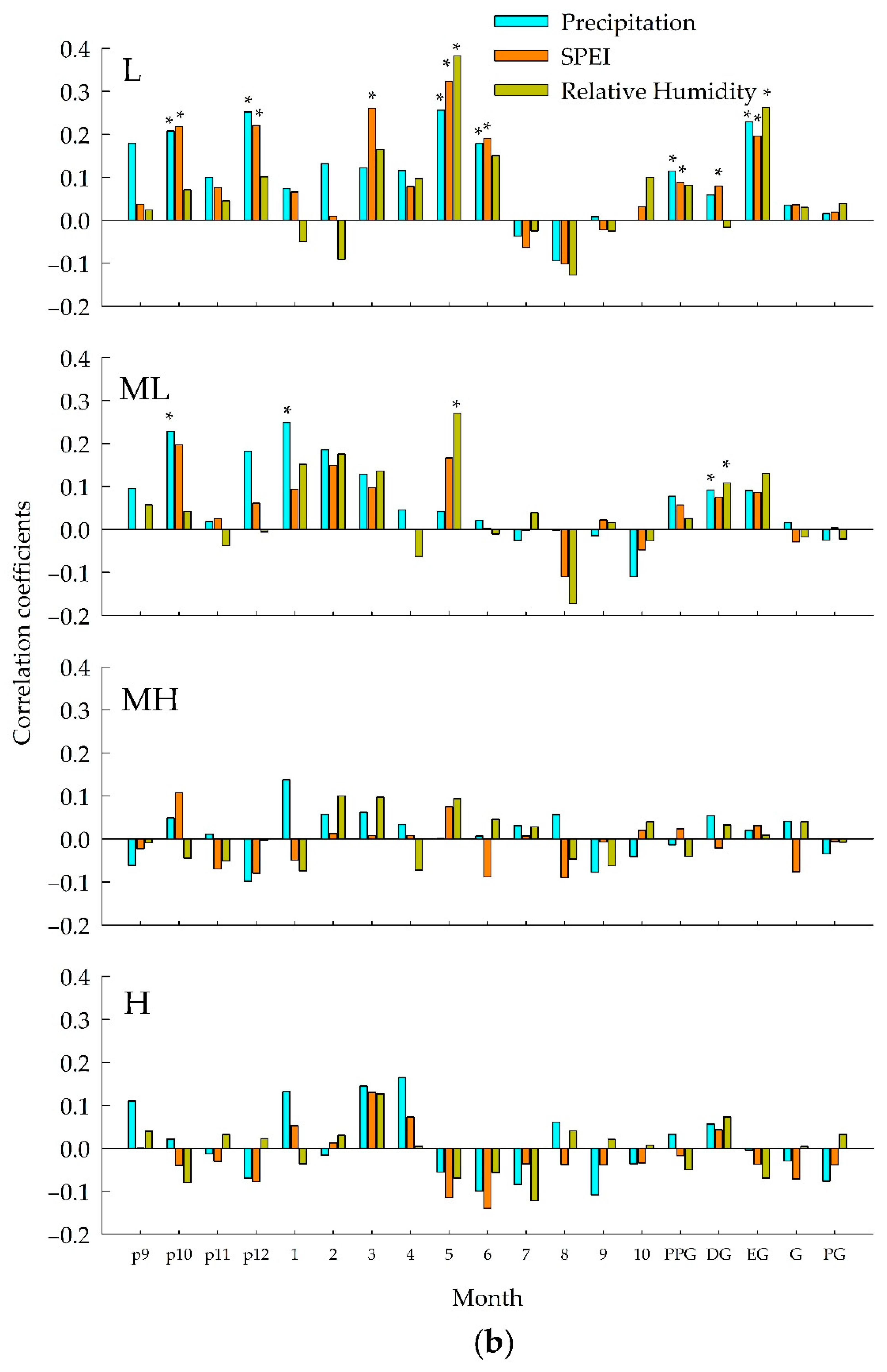
3.2. Relationship between Ring width Index and Climate Variables
3.3. Dynamic Relationships between Radial Growth and Climatic Change
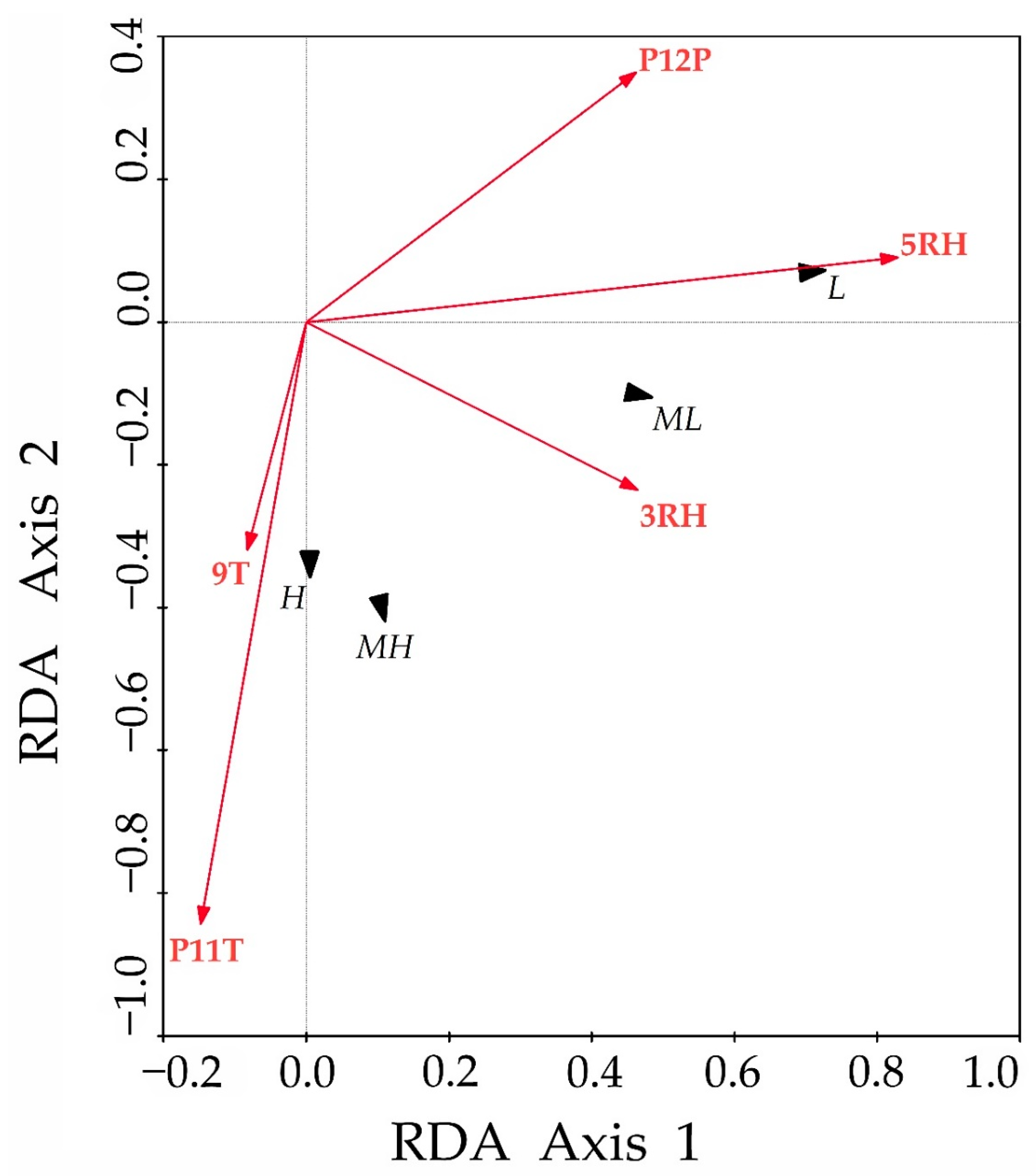
3.4. Redundancy Analysis between Climatic Factors and Residual Chronologies
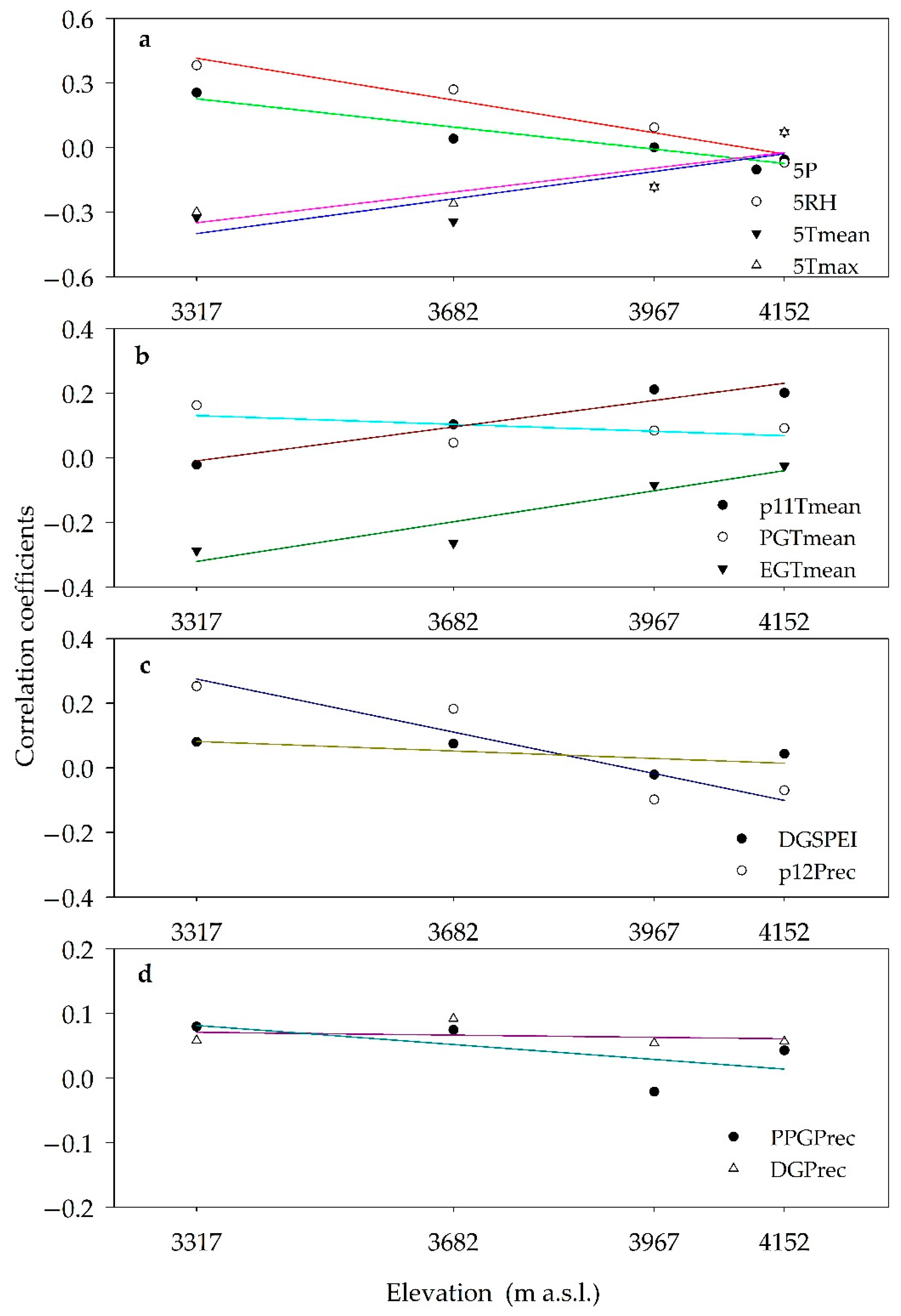
3.5. Altitudinal Trends of Growth Responses
4. Discussion
4.1. Climate-Growth Relationships along the Altitudinal Gradient
4.2. Consistent Effects among All Sites
4.3. Temporal Stability in Climate-Growth Relationships
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Maes, S.L.; Perring, M.P.; Vanhellemont, M.; Depauw, L.; Verheyen, K. Environmental drivers interactively affect individual tree growth across temperate European forests. Glob. Chang. Biol. 2019, 25, 201–217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boisvenue, C.; Running, S.W. Impacts of climate change on natural forest productivity-evidence since the middle of the 20th century. Glob. Chang. Biol. 2006, 12, 862–882. [Google Scholar] [CrossRef]
- Hessburg, P.F.; Miller, C.L.; Povak, N.A.; Taylor, A.H.; Safford, H.D. Climate, Environment, and Disturbance History Govern Resilience of Western North American Forests. Front. Ecol. Evol. 2019, 7, 239. [Google Scholar] [CrossRef] [Green Version]
- Galván, J.D.; Büntgen, U.; Ginzler, C.; Grudd, H.; Camarero, J.J. Drought-induced weakening of growth-temperature associations in high-elevation Iberian pines. Glob. Planet. Chang. 2015, 124, 95–106. [Google Scholar] [CrossRef] [Green Version]
- Shi, C.; Shen, M.; Wu, X.; Cheng, X.; Li, X.; Fan, T.; Li, Z.; Zhang, Y.; Fan, Z.; Shi, F. Growth response of alpine treeline forests to a warmer and drier climate on the southeastern Tibetan Plateau. Agric. For. Meteorol. 2018, 264, 73–79. [Google Scholar] [CrossRef]
- Lu, X.; Liang, E.; Wang, Y.; Babst, F.; Leavitt, S.W.; Camarero, J.J. Past the climate optimum: Recruitment is declining at the world’s highest juniper shrublines on the Tibetan Plateau. Ecology 2019, 100, e02557. [Google Scholar] [CrossRef] [Green Version]
- Arekhi, M.; Yesil, A.; Ozkan, U.Y.; Balik Sanli, F. Detecting treeline dynamics in response to climate warming using forest stand maps and Landsat data in a temperate forest. For. Ecosyst. 2018, 5, 311–324. [Google Scholar] [CrossRef] [Green Version]
- Marchand, W.; Girardin, M.P.; Hartmann, H.; Gauthier, S.; Bergeron, Y. Taxonomy, together with ontogeny and growing conditions, drives needleleaf species’ sensitivity to climate in boreal North America. Glob. Chang. Biol. 2019, 25, 2793–2809. [Google Scholar] [CrossRef]
- Alvarez, C.; Veblen, T.T.; Christie, D.A.; González-Reyes, A. Relationships between climate variability and radial growth of Nothofagus pumilio near altitudinal treeline in the Andes of northern Patagonia, Chile. For. Ecol. Manag. 2015, 342, 112–121. [Google Scholar] [CrossRef]
- Huang, R.; Zhu, H.F.; Liang, E.Y.; Liu, B.; Shi, J.F.; Zhang, R.B.; Yuan, Y.J.; Grießinger, J. A tree ring-based winter temperature reconstruction for the southeastern Tibetan Plateau since 1340 CE. Clim. Dyn. 2019, 53, 3221–3233. [Google Scholar] [CrossRef]
- Yu, D.; Wang, Q.; Wang, Y.; Zhou, W.; Hong, D.; Fang, X.; Jiang, S.; Dai, L. Climatic effects on radial growth of major tree species on Changbai Mountain. Ann. For. Sci. 2011, 68, 921–933. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Yin, D.C.; Sun, M.; Wang, H.; Tian, K.; Xiao, D.R.; Zhang, W.G. Variations of climate-growth response of major conifers at upper distributional limits in Shika Snow Mountain, Northwestern Yunnan Plateau, China. Forests 2017, 8, 377. [Google Scholar] [CrossRef] [Green Version]
- Lamarche, V.C. Frequency-Dependent Relationships between Tree-Ring Series along an Ecological Gradient and Some Dendroclimatic Implications; Citeseer: Princeton, NJ, USA, 1974. [Google Scholar]
- Dittmar, C.; Zech, W.; Elling, W. Growth variations of Common beech (Fagus sylvatica L.) under different climatic and environmental conditions in Europe-a dendroecological study. For. Ecol. Manag. 2003, 173, 63–78. [Google Scholar] [CrossRef]
- Leal, S.; Melvin, T.M.; Grabner, M.; Wimmer, R.; Briffa, K.R. Tree-ring growth variability in the Austrian Alps: The influence of site, altitude, tree species and climate. Boreas 2007, 36, 426–440. [Google Scholar] [CrossRef]
- Liu, L.S.; Shao, X.M.; Liang, E.Y. Climate Signals from Tree Ring Chronologies of the Upper and Lower Treelines in the Dulan Region of the Northeastern Qinghai-Tibetan Plateau. J. Integr. Plant Biol. 2010, 48, 278–285. [Google Scholar] [CrossRef]
- Liang, E.Y.; Wang, Y.F.; Xu, Y.; Liu, B.M.; Shao, X. Growth variation in Abies georgei var. smithii along altitudinal gradients in the Sygera Mountains, southeastern Tibetan Plateau. Trees 2010, 24, 363–373. [Google Scholar] [CrossRef]
- Fan, Z.X.; Bräuning, A.; Thomas, A.; Li, J.B.; Cao, K.F. Spatial and temporal temperature trends on the Yunnan Plateau (Southwest China) during 1961–2004. Int. J. Climatol. 2011, 31, 2078–2090. [Google Scholar] [CrossRef]
- Bräuning, A. Dendrochronology for the last 1400 years in eastern Tibet. GeoJournal 1994, 34, 75–95. [Google Scholar] [CrossRef]
- Zhang, Q.B.; Cheng, G.D.; Yao, T.D.; Kang, X.C.; Huang, J.G. A 2326-year tree-ring record of climate variability on the Northeastern Qinghai-Tibetan Plateau. Geophys. Res. Lett. 2003, 30, HLS 2-1. [Google Scholar] [CrossRef] [Green Version]
- Fan, Z.X.; Bräuning, A.; Cao, K.F. Annual temperature reconstruction in the central Hengduan Mountains, China, as deduced from tree rings. Dendrochronologia 2008, 26, 97–107. [Google Scholar] [CrossRef]
- Fan, Z.X.; Bräuning, A.; Cao, K.F. Tree-ring based drought reconstruction in the central Hengduan Mountains region (China) since A.D. 1655. Int. J. Climatol. 2010, 28, 1879–1887. [Google Scholar] [CrossRef]
- Fan, Z.X.; Bräuning, A.; Cao, K.F.; Zhu, S.D. Growth-climate responses of high-elevation conifers in the central Hengduan Mountains, southwestern China. For. Ecol. Manag. 2009, 258, 306–313. [Google Scholar] [CrossRef]
- Li, Z.S.; Zhang, Q.B.; Ma, K. Tree-ring reconstruction of summer temperature for A.D. 1475-2003 in the central Hengduan Mountains, Northwestern Yunnan, China. Clim. Chang. 2012, 110, 455–467. [Google Scholar] [CrossRef]
- Panthi, S.; Bräuning, A.; Zhou, Z.K.; Fan, Z.X. Growth response of Abies georgei to climate increases with elevation in the central Hengduan Mountains, southwestern China. Dendrochronologia 2018, 47, 1–9. [Google Scholar] [CrossRef]
- Guo, G.; Li, Z.S.; Zhang, Q.B.; Ma, K.P.; Mu, C. Dendroclimatological studies of Picea likiangensis and Tsuga dumosa in Lijiang, China. IAWA J. 2009, 30, 435–441. [Google Scholar] [CrossRef] [Green Version]
- Bi, Y.F.; Xu, J.C.; Gebrekirstos, A.; Guo, L.; Zhao, M.X.; Liang, E.Y.; Yang, X.F. Assessing drought variability since 1650 AD from tree-rings on the Jade Dragon Snow Mountain, southwest China. Int. J. Climatol. 2016, 35, 4057–4065. [Google Scholar] [CrossRef]
- Zhang, Y.; Cao, R.J.; Yin, J.; Tian, K.; Zhang, W.G.; Yin, D.C. Radial growth response of major conifers to climate change on Haba Snow Mountain, Southwestern China. Dendrochronologia 2020, 60, 125682. [Google Scholar] [CrossRef]
- Yin, D.C.; Xu, D.R.; Tian, K.; Xiao, D.R.; Zhang, W.G.; Sun, D.C.; Sun, H.; Zhang, Y. Radial growth response of Abies georgei to climate at the Upper timberlines in Central Hengduan Mountains, Southwestern China. Forests 2018, 9, 606. [Google Scholar] [CrossRef] [Green Version]
- Liu, G.; Zhang, W.; He, M.Y. The preliminary study of quaternary glacier development in Haba snow mountain. Yunnan Geogr. Environ. Res. 2012, 24, 104–110. [Google Scholar]
- Su, H.; Wang, P. Climate vertical zoning of Haba natural reserve. Yunnan Geogr. Environ. Res. 2011, 23, 47–51. [Google Scholar]
- Stokes, M.A.; Smiley, T.L. An Introduction to Tree-Ring Dating; University of Arizona Press: Tucson, AZ, USA, 1996. [Google Scholar]
- Larsson. CooRecorder and CDendro Programs of the CooRecorder/CDendro Package Version 9.3. 2019. Available online: http://www.cybis.se/forfun/dendro/ (accessed on 11 July 2017).
- Holmes, R.L. Computer-assisted quality control in tree-ring Dating and Measurement. Tree-Ring Bull. 1983, 43, 69–75. [Google Scholar]
- Vicente-Serrano, S.M.; Beguería, S.; López-Moreno, J.I. A multiscalar drought index sensitive to global warming: The standardized precipitation evapotranspiration index. J. Clim. 2010, 23, 1696–1718. [Google Scholar] [CrossRef] [Green Version]
- Oberhuber, W. Influence of climate on radial growth of Pinus cembra within the alpine timberline ecotone. Tree Physiol. 2004, 24, 291–301. [Google Scholar] [CrossRef]
- Biondi, F.; Waikul, K. DENDROCLIM2002: A C++ program for statistical calibration of climate signals in tree-ring chronologies. Comput. Geosci. 2004, 30, 303–311. [Google Scholar] [CrossRef]
- Ter Braak, C.J.; Smilauer, P. CANOCO Reference Manual and CanoDraw for Windows User’s Guide: Software for Canonical Community Ordination (Version 4.5); Microcomputer Power: Ithaca, NY, USA, 2002. [Google Scholar]
- Gaire, N.P.; Fan, Z.X.; Braeuning, A.; Panthi, S.; Rana, P.; Shrestha, A.; Bhuju, D.R. Abies spectabilis shows stable growth relations to temperature, but changing response to moisture conditions along an elevation gradient in the central Himalaya. Dendrochronologia 2020, 60, 125675. [Google Scholar] [CrossRef]
- Rai, S.; Dawadi, B.; Wang, Y.F.; Lu, X.M.; Sigdel, S.R. Growth response of Abies spectabilis to climate along an elevation gradient of the Manang valley in the central Himalayas. J. For. Res. 2020, 31, 2245–2254. [Google Scholar] [CrossRef] [Green Version]
- Denmead, O.T.; Shaw, R.H. Availability of soil water to plants as affected by soil moisture content and meteorological conditions 1. Agron. J. 1962, 54, 385–390. [Google Scholar] [CrossRef]
- Grießinger, J.; Bräuning, A. Late Holocene variations in monsoon intensity in the Tibetan Himalayan Region-Evidence from tree rings. J. Geol. Soc. India. 2006, 68, 485–493. [Google Scholar]
- Li, J.B.; Shi, J.F.; Zhang, D.D.; Yang, B.; Fang, K.Y.; Yue, P.H. Moisture increase in response to high-altitude warming evidenced by tree-rings on the southeastern Tibetan Plateau. Clim. Dyn. 2017, 48, 649–660. [Google Scholar] [CrossRef] [Green Version]
- Liang, E.Y.; Dawadi, B.; Pederson, N.; Eckstein, D. Is the growth of birch at the upper timberline in the Himalayas limited by moisture or by temperature? Ecology 2014, 95, 2453–2465. [Google Scholar] [CrossRef] [Green Version]
- Dawadi, B.; Liang, E.Y.; Tian, L.D.; Devkota, L.P.; Yao, T.D. Pre-monsoon precipitation signal in tree rings of timberline Betula utilis in the central Himalayas. Quat. Int. 2013, 283, 72–77. [Google Scholar] [CrossRef]
- Kharal, D.K.; Thapa, U.K.; George, S.S.; Meilby, H.; Rayamajhi, S.; Bhuju, D.R. Tree-climate relations along an elevational transect in Manang Valley, central Nepal. Dendrochronologia 2017, 41, 57–64. [Google Scholar] [CrossRef]
- Sohar, K.; Altman, J.; Lehečková, E.; Doležal, J. Growth-climate relationships of Himalayan conifers along elevational and latitudinal gradients. Int. J. Climatol. 2017, 37, 2593–2605. [Google Scholar] [CrossRef]
- Wang, H.; Zhou, J.; Qin, X.H.; Zhang, Y. Radial Growth Responses of Pinus densata to Climate Change in Haba Snow Mountain, Southwest China. For. Resour. Manag. 2019, 2, 67–72. (In Chinese) [Google Scholar]
- Liang, E.Y.; Shao, X.M.; Xu, Y. Tree-ring evidence of recent abnormal warming on the southeast Tibetan Plateau. Theor. Appl. Climatol. 2009, 98, 9–18. [Google Scholar] [CrossRef]
- Cannell, M.G.R. Growth control in woody plants. Tree Physiol. 1997, 17, 489. [Google Scholar] [CrossRef] [Green Version]
- Büntgen, U.; Frank, D.C.; Kaczka, R.J.; Verstege, A.; Zwijacz-Kozica, T.; Esper, J. Growth responses to climate in a multi-species tree-ring network in the Western Carpathian Tatra Mountains, Poland and Slovakia. Tree Physiol. 2007, 27, 689–702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gricar, J.; Zupancic, M.; Cufar, K.; Oven, P. Regular cambial activity and xylem and phloem formation in locally heated and cooled stem portions of Norway spruce. Wood Sci. Technol. 2007, 41, 463–475. [Google Scholar] [CrossRef]
- Takahashi, K.; Okuhara, I.; Tokumitsu, Y.; Yasue, K. Responses to climate by tree-ring widths and maximum latewood densities of two Abies species at upper and lower altitudinal distribution limits in central Japan. Trees-Struct. Funct. 2011, 25, 745–753. [Google Scholar] [CrossRef]
- Zhang, Y.; Yin, D.C.; Zhang, W.G.; Yue, H.T.; Du, J.C.D.; Li, Q.P.; Yang, R.; Tian, K. Response of radial growth of two conifers to temperature and precipitation in Potatso National Park, Southwest China. Acta Ecol. Sin. 2018, 38, 5383–5392. (In Chinese) [Google Scholar]
- Qin, L.; Yuan, Y.; Zhang, R.; Wei, W.; Yu, S.; Fan, Z.; Chen, F.; Zhang, Y.; Shang, H. Tree-ring response to snow cover and reconstruction of century annual maximum snow depth for Northern Tianshan Mountains, China. Geochronometria 2016, 43, 9–17. [Google Scholar] [CrossRef] [Green Version]
- Villalba, R.; Boninsegna, J.; Veblen, T.T.; Schmelter, A.; Rubulis, S. Recent trends in tree-ring records from high elevation sites in the Andes of northern Patagonia. Clim. Chang. 1997, 36, 425–454. [Google Scholar] [CrossRef]
- Vaganov, E.A.; Hughes, M.K.; Kirdyanov, A.V.; Schweingruber, F.H.; Silkin, P.P. Influence of snowfall and melt timing on tree growth in Subarctic Eurasia. Nature 1999, 400, 149–151. [Google Scholar] [CrossRef]
- Yu, D.; Wang, G.G.; Dai, L.; Wang, Q. Dendroclimatic analysis of Betula ermanii forests at their upper limit of distribution in Changbai Mountain, Northeast China. For. Ecol. Manag. 2007, 240, 105–113. [Google Scholar] [CrossRef]
- Takahashi, K.; Tokumitsu, Y.; Yasue, K. Climatic factors affecting the tree-ring width of Betula ermanii at the timberline on Mount Norikura, central Japan. Ecol. Res. 2005, 20, 445–451. [Google Scholar] [CrossRef] [Green Version]
- Payette, S.; Delwalde, A.; Morneau, C.; Lavole, C. Patterns of tree stem decline along a snow-drift gradient at treeline: A case study using stem analysis. Can. J. Bot. 1998, 74, 1671–1683. [Google Scholar] [CrossRef]
- Liang, E.Y.; Shao, X.M.; Hu, Y.X.; Lin, J.X. Dendroclimatic evaluation of climate-growth relationships of Meyer spruce (Picea meyeri) on a sandy substrate in semi-arid grassland, north China. Trees-Struct Funct. 2001, 15, 230–235. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sites | Altitude/m a.s.l. | Longitude/E | Latitude/N | No. (Tree/Core) |
|---|---|---|---|---|
| Low altitude (L) | 3317 | 100°04′54.78″ | 27°20′19.86″ | 26/52 |
| Middle–Low altitude (ML) | 3682 | 100°04′19.62″ | 27°20′25.37″ | 33/60 |
| Middle–High altitude (MH) | 3967 | 100°05′31.25″ | 27°20′39.71″ | 35/62 |
| High altitude (H) | 4152 | 100°05′51.48″ | 27°20′48.90″ | 34/56 |
| Residual Chronologies | L | ML | MH | H |
|---|---|---|---|---|
| No. (tree/radii) | 26/51 | 30/52 | 34/57 | 34/56 |
| chronology length | 1933–2016 | 1923–2016 | 1831–2016 | 1737–2016 |
| Mean sensitivity | 0.21 | 0.14 | 0.15 | 0.11 |
| Statistics of common interval analysis (1950–2016) | ||||
| Variance in first eigenvector/% | 49.33% | 32.18% | 34.79% | 36.20% |
| Standard deviation | 0.18 | 0.13 | 0.12 | 0.10 |
| Signal-to-noise ratio | 6.77 | 6.25 | 22.20 | 26.81 |
| Expressed population signal | 0.87 | 0.86 | 0.96 | 0.96 |
| L | ML | MH | H | |
|---|---|---|---|---|
| L | 1.000 | |||
| ML | 0.367 ** | 1.000 | ||
| MH | 0.076 | 0.348 ** | 1.000 | |
| H | 0.027 | 0.363 ** | 0.494 ** | 1.000 |
| Sampling Site | Interval Period | Climatic Variable | Significant Year |
|---|---|---|---|
| L | 1953–2016 | Previous October precipitation | 1953–1970 (+), 1974–1990 (+), 2012 (+), 2014–2016 (+) |
| Previous December precipitation | 1953–1985 (+), 1992–1993 (+), 1995 (+), 1999–2016 (+) | ||
| Current May precipitation | 1953–2016 (+) | ||
| Previous October SPEI | 1953–1963 (+), 1968–1969 (+), 1980 (+), 1993–2009 (+), 2014–2016 (+) | ||
| Previous December SPEI | 1953–1990 (+), 2002 (+), 2007 (+), 2009 (+), 2011 (+), 2013–2016 (+) | ||
| Current March SPEI | 1953–1965 (+), 1967 (+), 1969 (+), 1986–1987 (+), 1999 (+), 2002–2016 (+) | ||
| Current May SPEI | 1953–2016 (+) | ||
| Current May relative humidity | 1953–2016 (+) | ||
| Current May temperature | 1953–2016 (−) | ||
| Current October temperature | 1953–1967 (+), 1972–1976 (+), 1998 (+), 2000–2001 (+), 2003–2013 (+), 2015–2016 (+) | ||
| ML | 1953–2016 | Previous October precipitation | 1953–1963 (+), 1966–1967 (+), 1980–1986 (+) 1990–1994 (+), 2006–2007 (+), 2009–2016 (+) |
| Current January precipitation | 1953–1969 (+), 1973–1975 (+), 1989–1990 (+), 2003 (+), 2006–2016 (+) | ||
| Current May relative humidity | 1953–1970 (+), 1972–1975 (+), 1978 (+), 1980–1985 (+), 2004 (+), 2007–2016 (+) | ||
| Current May temperature | 1953–2016 (−) | ||
| MH | 1953–2016 | Previous November temperature | 1953–1967 (+), 1974 (+), 1980–1981 (+), 1983–1996 (+), 2000 (+), 2003 (+), 2011–2016 (+) |
| H | 1953–2016 | Previous November temperature | 1953–1965 (+), 2014–2016 (+) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, M.; Li, J.; Cao, R.; Tian, K.; Zhang, W.; Yin, D.; Zhang, Y. Climate-Growth Relations of Abies georgei along an Altitudinal Gradient in Haba Snow Mountain, Southwestern China. Forests 2021, 12, 1569. https://0-doi-org.brum.beds.ac.uk/10.3390/f12111569
Sun M, Li J, Cao R, Tian K, Zhang W, Yin D, Zhang Y. Climate-Growth Relations of Abies georgei along an Altitudinal Gradient in Haba Snow Mountain, Southwestern China. Forests. 2021; 12(11):1569. https://0-doi-org.brum.beds.ac.uk/10.3390/f12111569
Chicago/Turabian StyleSun, Mei, Jianing Li, Renjie Cao, Kun Tian, Weiguo Zhang, Dingcai Yin, and Yun Zhang. 2021. "Climate-Growth Relations of Abies georgei along an Altitudinal Gradient in Haba Snow Mountain, Southwestern China" Forests 12, no. 11: 1569. https://0-doi-org.brum.beds.ac.uk/10.3390/f12111569