
Predicting Potential Habitat of a Plant Species with Small Populations under Climate Change: Ostryarehderiana

 , , ,
, , , 
Abstract
:1. Introduction
2. Materials and Methods
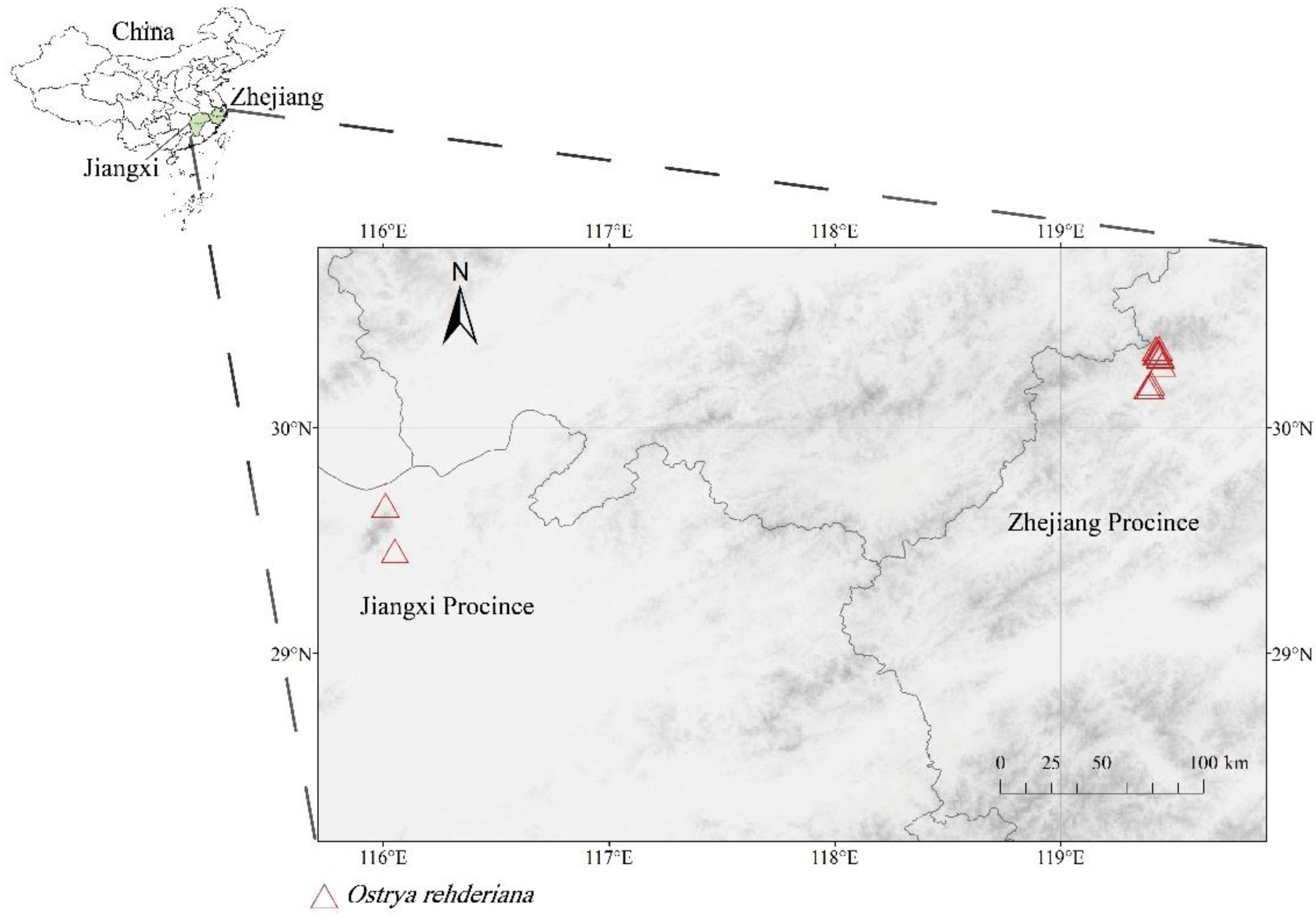
2.1. Data Collection
2.2. Model Setting and Evaluation
2.3. Model Predictions
3. Results
3.1. Prediction Accuracy of O. rehderiana Suitability
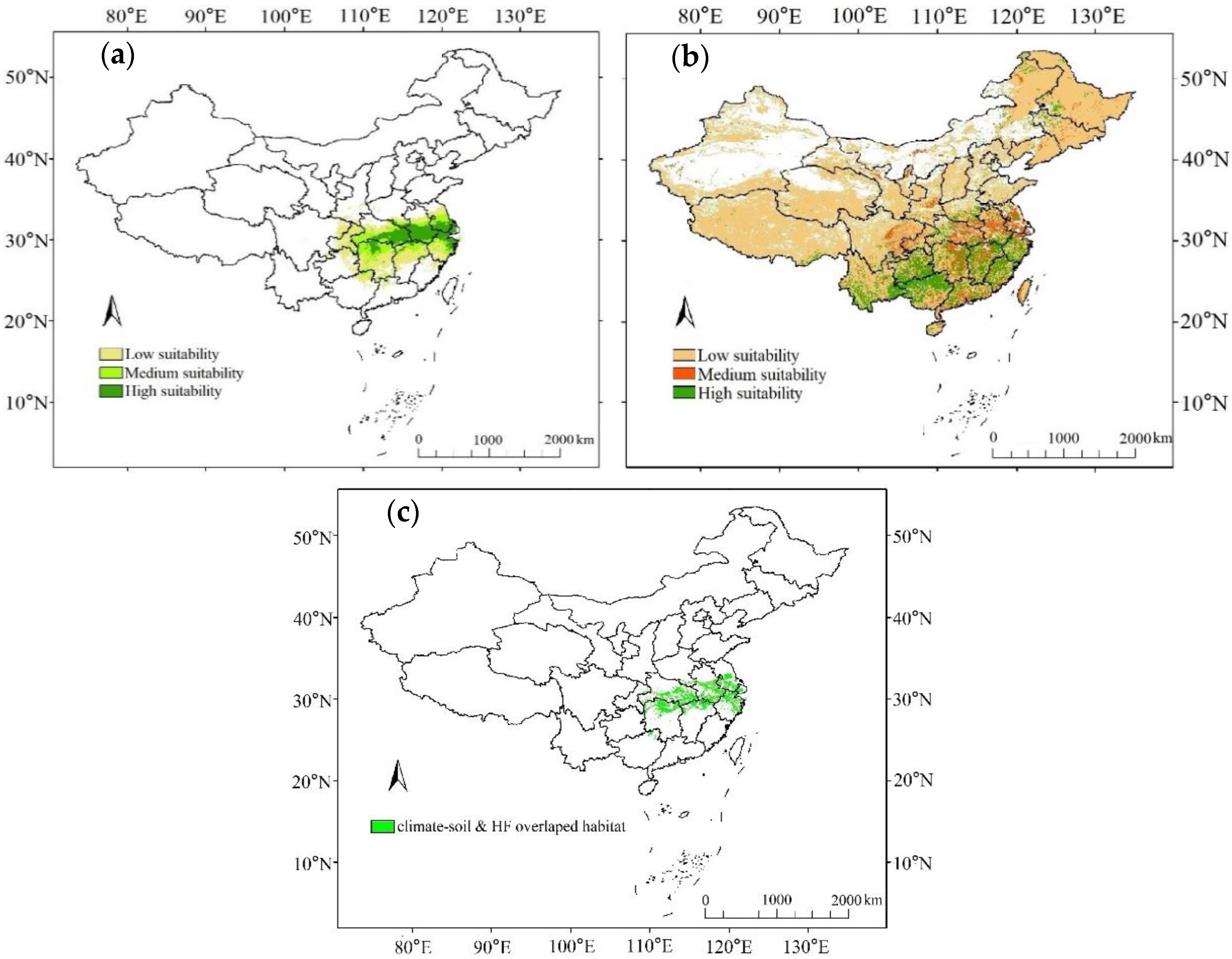
3.2. Predicted O. rehderiana Contemporary Suitable Habitats
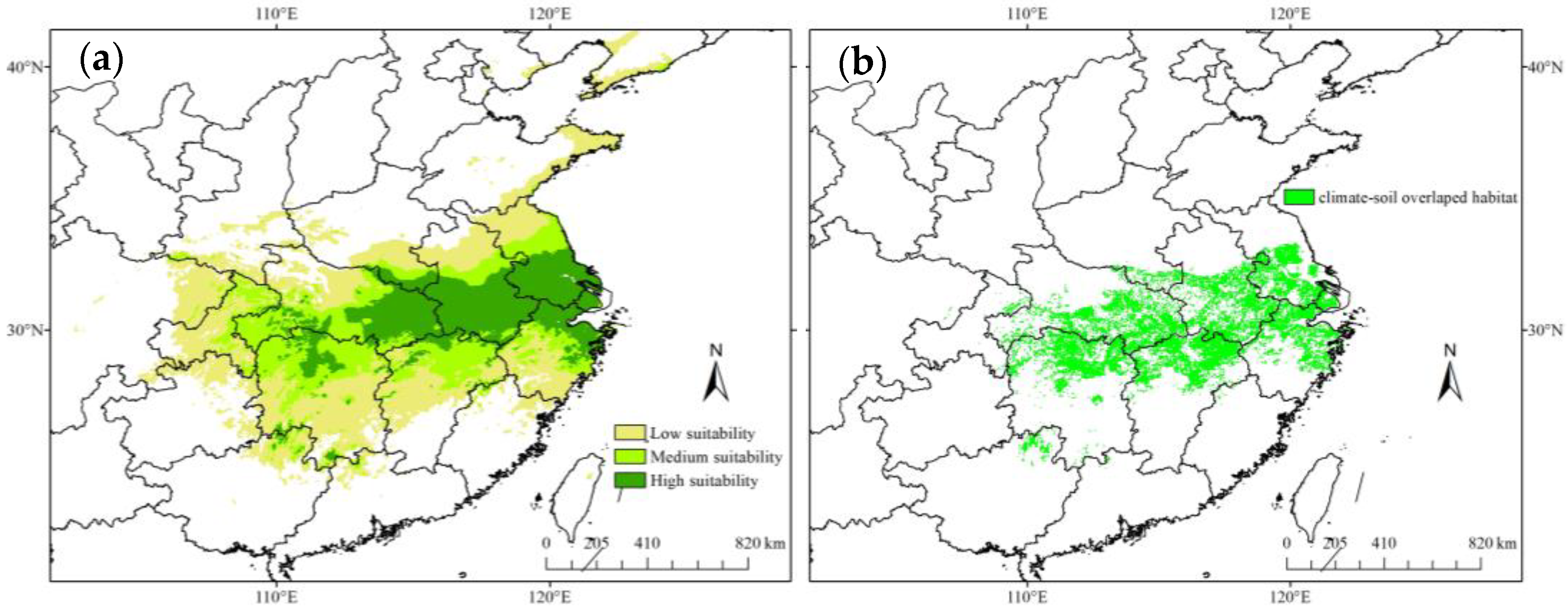
3.3. O. rehderiana Potential Distribution under Different Climate Change Scenarios
4. Discussion
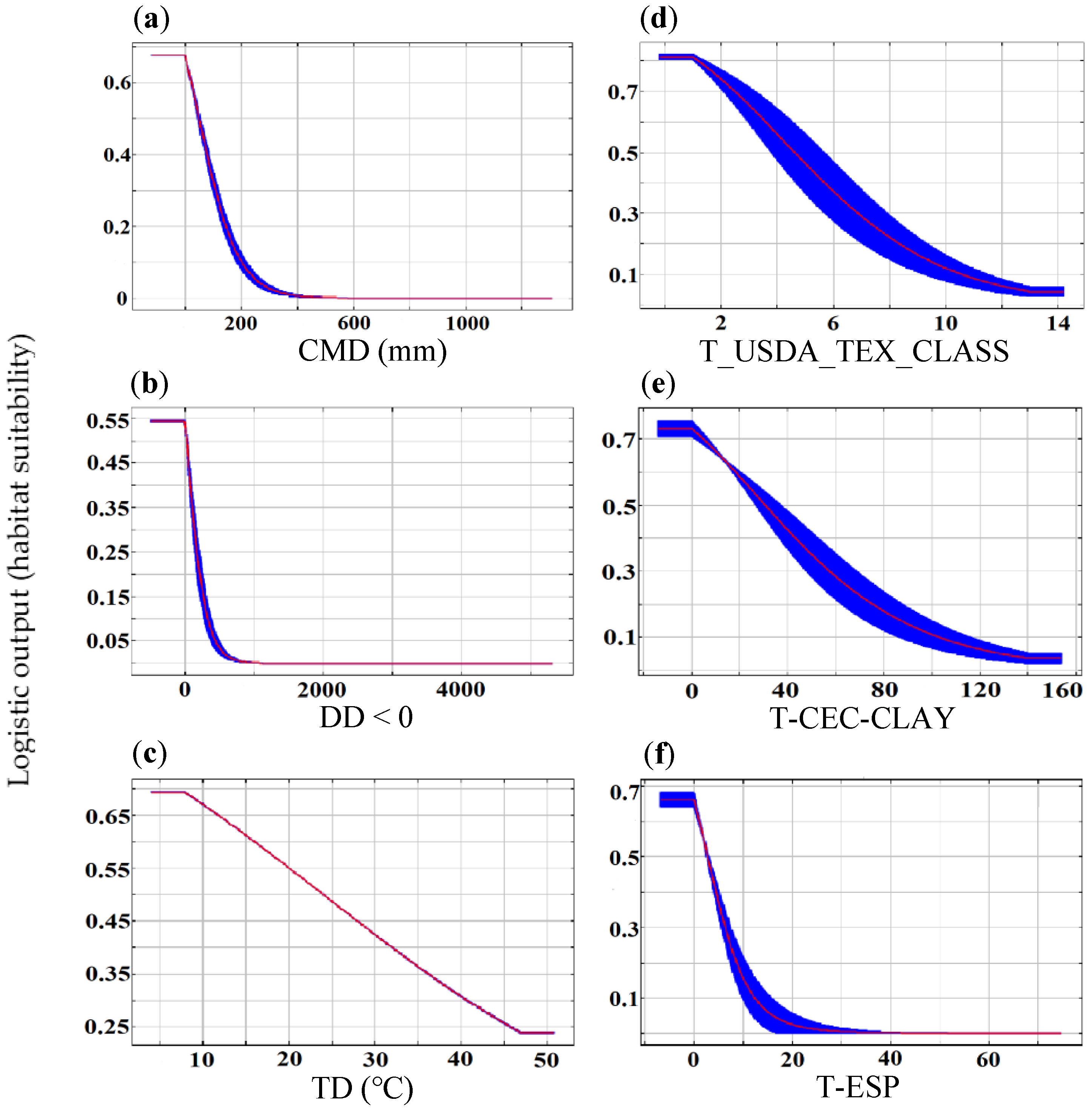
4.1. Key Environmental Factors Determining the O. rehderiana Distribution
4.2. O. rehderiana Predicted Changes to Climatic Suitable Habitat
4.3. Implications for O. rehderiana Conservation
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Cortini, F.; Comeau, P.G.; Wang, T.; Hibbs, D.; Bluhm, A. Climate effects on red alder growth in the Pacific Northwest of North America. For. Ecol. Manag. 2012, 277, 98–106. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.; Coops, N.C.; Wang, T.; Wang, G. A process-based approach to estimate Chinese fir (Cunninghamia lanceolata) distribution and productivity in southern China under climate change. Forests 2015, 6, 360–379. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.; Hamann, A.; Spittlehouse, D.; Carroll, C. Locally downscaled and spatially customizable climate data for historical and future periods for North America. PLoS ONE 2016, 11, e0156720. [Google Scholar] [CrossRef] [PubMed]
- Davis, M.B.; Shaw, R.G. Range shifts and adaptive responses to Quaternary climate change. Science 2001, 292, 673–679. [Google Scholar] [CrossRef] [Green Version]
- Provan, J.; Bennett, K.D. Phylogeographic insights into cryptic glacial refugia. Trends Ecol. Evol. 2008, 23, 564–571. [Google Scholar] [CrossRef]
- Li, Y.-Y.; Guan, S.-M.; Yang, S.-Z.; Luo, Y.; Chen, X.-Y. Genetic decline and inbreeding depression in an extremely rare tree. Conserv. Genet. 2012, 13, 343–347. [Google Scholar] [CrossRef]
- Sun, W. Words from the guest editor-in-chief. Plant Divers. 2016, 38, 207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, Y.; Chen, G.; Grumbine, R.E.; Dao, Z.; Sun, W.; Guo, H. Conserving plant species with extremely small populations (PSESP) in China. Biodivers. Conserv. 2013, 22, 803–809. [Google Scholar] [CrossRef]
- Liu, X.; He, Y.; Xiao, Y.; Wang, Y.; Jiang, Y.; Jiang, Y. Soil seed burial and competition with surrounding plants determine the emergence and development of seedling of an endangered species Horsfieldia hainanensis Merr. in China. Sci. Rep. 2019, 9, 17970. [Google Scholar] [CrossRef]
- Bertrand, R.; Perez, V.; Gégout, J.C. Disregarding the edaphic dimension in species distribution models leads to the omission of crucial spatial information under climate change: The case of Quercus pubescens in F rance. Glob. Chang. Biol. 2012, 18, 2648–2660. [Google Scholar] [CrossRef]
- Li, S.; Wu, J.; Gong, J.; Li, S. Human footprint in Tibet: Assessing the spatial layout and effectiveness of nature reserves. Sci. Total Environ. 2018, 621, 18–29. [Google Scholar] [CrossRef]
- Joly, C.A.; Metzger, J.P.; Tabarelli, M. Experiences from the B razilian A tlantic F orest: Ecological findings and conservation initiatives. New Phytol. 2014, 204, 459–473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chardon, N.I.; Rixen, C.; Wipf, S.; Doak, D.F. Human trampling disturbance exerts different ecological effects at contrasting elevational range limits. J. Appl. Ecol. 2019, 56, 1389–1399. [Google Scholar] [CrossRef]
- Di Marco, M.; Venter, O.; Possingham, H.P.; Watson, J.E. Changes in human footprint drive changes in species extinction risk. Nat. Commun. 2018, 9, 4621. [Google Scholar] [CrossRef] [PubMed]
- Blach Overgaard, A.; Svenning, J.C.; Dransfield, J.; Greve, M.; Balslev, H. Determinants of palm species distributions across Africa: The relative roles of climate, nonclimatic environmental factors, and spatial constraints. Ecography 2010, 33, 380–391. [Google Scholar] [CrossRef]
- Figueiredo, F.O.; Zuquim, G.; Tuomisto, H.; Moulatlet, G.M.; Balslev, H.; Costa, F.R. Beyond climate control on species range: The importance of soil data to predict distribution of Amazonian plant species. J. Biogeogr. 2018, 45, 190–200. [Google Scholar] [CrossRef]
- Zuquim, G.; Tuomisto, H.; Jones, M.M.; Prado, J.; Figueiredo, F.O.; Moulatlet, G.M.; Costa, F.R.; Quesada, C.A.; Emilio, T. Predicting environmental gradients with fern species composition in Brazilian Amazonia. J. Veg. Sci. 2014, 25, 1195–1207. [Google Scholar] [CrossRef]
- Deng, S.; Wu, Y.; Wu, K.; Fang, L.; Li, L.; Zeng, S. Breeding characteristics and artificial propagation of 14 species of Wild Plant with Extremely Small Populations (WPESP) in China. Biodivers. Sci. 2020, 28, 385–400. [Google Scholar] [CrossRef]
- Kanglin, G. Current situation and propagation of rare tree species-Ostrya rederiana. J. Zhejiang For. Coll. 1988, 1, 93–95. [Google Scholar]
- Yang, Y.; Ma, T.; Wang, Z.; Lu, Z.; Li, Y.; Fu, C.; Chen, X.; Zhao, M.; Olson, M.S.; Liu, J. Genomic effects of population collapse in a critically endangered ironwood tree Ostrya rehderiana. Nat. Commun. 2018, 9, 5449. [Google Scholar] [CrossRef]
- Feng, L.; Sun, J.; Shi, Y.; Wang, G.; Wang, T. Predicting Suitable Habitats of Camptotheca acuminata Considering Both Climatic and Soil Variables. Forests 2020, 11, 891. [Google Scholar] [CrossRef]
- Zhang, L.; Liu, S.; Sun, P.; Wang, T.; Wang, G.; Zhang, X.; Wang, L. Consensus forecasting of species distributions: The effects of niche model performance and niche properties. PLoS ONE 2015, 10, e0120056. [Google Scholar] [CrossRef] [Green Version]
- Elith, J.; Phillips, S.J.; Hastie, T.; Dudík, M.; Chee, Y.E.; Yates, C.J. A statistical explanation of MaxEnt for ecologists. Divers. Distrib. 2011, 17, 43–57. [Google Scholar] [CrossRef]
- Phillips, S.J.; Dudík, M. Modeling of species distributions with Maxent: New extensions and a comprehensive evaluation. Ecography 2008, 31, 161–175. [Google Scholar] [CrossRef]
- Di Febbraro, M.; Carotenuto, F.; Castiglione, S.; Russo, D.; Loy, A.; Maiorano, L.; Raia, P. Does the jack of all trades fare best? Survival and niche width in Late Pleistocene megafauna. J. Biogeogr. 2017, 44, 2828–2838. [Google Scholar] [CrossRef] [Green Version]
- Kukwa, M.; Kolanowska, M. Glacial refugia and the prediction of future habitat coverage of the South American lichen species Ochrolechia austroamericana. Sci. Rep. 2016, 6, 38779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, T.; Wang, G.; Innes, J.L.; Seely, B.; Chen, B. ClimateAP: An application for dynamic local downscaling of historical and future climate data in Asia Pacific. Front. Agric. Sci. Eng. 2017, 4, 448–458. [Google Scholar] [CrossRef] [Green Version]
- Fischedick, M.; Roy, J.; Acquaye, A.; Allwood, J.; Ceron, J.-P.; Geng, Y.; Kheshgi, H.; Lanza, A.; Perczyk, D.; Price, L. Industry In: Climate Change 2014: Mitigation of Climate Change. In Contribution of Working Group III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Technical Report; Gambridge University Press: Cambridge, UK; New York, NY, USA, 2014. [Google Scholar]
- Fischer, G.; Van Velthuizen, H.; Shah, M.; Nachtergaele, F. Global Agroecological Assessment for Agriculture in the 21st Century; Food and Agriculture Organization of the United Nations (FAO): Rome, Italy; International Institute for Applied Systems Analysis (IIASA): Laxenburg, 2002. [Google Scholar]
- Venter, O.; Sanderson, E.W.; Magrach, A.; Allan, J.R.; Beher, J.; Jones, K.R.; Possingham, H.P.; Laurance, W.F.; Wood, P.; Fekete, B.M. Sixteen years of change in the global terrestrial human footprint and implications for biodiversity conservation. Nat. Commun. 2016, 7, 12558. [Google Scholar] [CrossRef] [Green Version]
- Zhan, P.; Wang, F.; Xia, P.; Zhao, G.; Wei, M.; Wei, F.; Han, R. Assessment of suitable cultivation region for Panax notoginseng under different climatic conditions using MaxEnt model and high-performance liquid chromatography in China. Ind. Crops Prod. 2022, 176, 114416. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef] [Green Version]
- Fielding, A.H.; Bell, J.F. A review of methods for the assessment of prediction errors in conservation presence/absence models. Environ. Conserv. 1997, 24, 38–49. [Google Scholar] [CrossRef]
- Elith, J.; Graham, C.H.; Anderson, R.P.; Dudík, M.; Ferrier, S.; Guisan, A.; Hijmans, R.J.; Huettmann, F.; Leathwick, J.R.; Lehmann, A. Novel methods improve prediction of species’ distributions from occurrence data. Ecography 2006, 29, 129–151. [Google Scholar] [CrossRef] [Green Version]
- Guo, J.; Liu, X.P.; Zhang, Q.; Zhang, D.F.; Xie, C.X.; Liu, X. Prediction for the potential distribution area of Codonopsis pilosula at global scale based on Maxent model. J. Appl. Ecol. 2017, 28, 992–1000. [Google Scholar] [CrossRef]
- Qin, A.; Liu, B.; Guo, Q.; Bussmann, R.W.; Ma, F.; Jian, Z.; Xu, G.; Pei, S. Maxent modeling for predicting impacts of climate change on the potential distribution of Thuja sutchuenensis Franch., an extremely endangered conifer from southwestern China. Glob. Ecol. Conserv. 2017, 10, 139–146. [Google Scholar] [CrossRef]
- Yi, Y.-j.; Zhou, Y.; Cai, Y.-p.; Yang, W.; Li, Z.-w.; Zhao, X. The influence of climate change on an endangered riparian plant species: The root of riparian Homonoia. Ecol. Indic. 2018, 92, 40–50. [Google Scholar] [CrossRef]
- Stanton, J.C.; Pearson, R.G.; Horning, N.; Ersts, P.; Reşit Akçakaya, H. Combining static and dynamic variables in species distribution models under climate change. Methods Ecol. Evol. 2012, 3, 349–357. [Google Scholar] [CrossRef]
- Albrecht, M.A.; Guerrant, E.O.; Maschinski, J.; Kennedy, K.L. A long-term view of rare plant reintroduction. Biol. Conserv. 2011, 11, 2557–2558. [Google Scholar] [CrossRef]
- Shao, H.; TIAN, J.-Q.; Guo, K.; SUN, O.J. Effects of sample size and species traits on performance of bioclim in predicting geographical distribution of tree species-a case study with 12 deciduous Quercus species indigenous to China. Chin. J. Plant Ecol. 2009, 33, 870–877. [Google Scholar] [CrossRef]
- Jiang, Y.; Wang, T.; De Bie, C.; Skidmore, A.; Liu, X.; Song, S.; Zhang, L.; Wang, J.; Shao, X. Satellite-derived vegetation indices contribute significantly to the prediction of epiphyllous liverworts. Ecol. Indic. 2014, 38, 72–80. [Google Scholar] [CrossRef]
- Bakoumé, C.; Shahbudin, N.; Yacob, S.; Siang, C.S.; Thambi, M.N.A. Improved method for estimating soil moisture deficit in oil palm (Elaeis guineensis Jacq.) areas with limited climatic data. J. Agric. Sci. 2013, 5, 57. [Google Scholar] [CrossRef] [Green Version]
- Sun, J.; Feng, L.; Wang, T.; Tian, X.; He, X.; Xia, H.; Wang, W. Predicting the Potential Habitat of Three Endangered Species of Carpinus Genus under Climate Change and Human Activity. Forests 2021, 12, 1216. [Google Scholar] [CrossRef]
- Feng, L.; Sun, J.; Wang, T.; Tian, X.; Wang, W.; Guo, J.; Feng, H.; Guo, H.; Deng, H.; Wang, G. Predicting suitable habitats of Ginkgo biloba L. fruit forests in China. Clim. Risk Manag. 2021, 100364. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, T.; El-Kassaby, Y.A. Contributions of dynamic environmental signals during life-cycle transitions to early life-history traits in lodgepole pine (Pinus contorta Dougl.). Biogeosciences 2016, 13, 2945–2958. [Google Scholar] [CrossRef] [Green Version]
- Yu, F.; Wang, T.; Groen, T.A.; Skidmore, A.K.; Yang, X.; Ma, K.; Wu, Z. Climate and land use changes will degrade the distribution of Rhododendrons in China. Sci. Total Environ. 2019, 659, 515–528. [Google Scholar] [CrossRef] [PubMed]
- Castro Franco, M.; Domenech, M.B.; Borda, M.R.; Costa, J.L. A Spatial Dataset of Topsoil Texture for the Southern Argentine Pampas. Geoderma Regional. 2018, 12, 18–27. [Google Scholar] [CrossRef]
- Li, N.; Zare, E.; Huang, J.; Triantafilis, J. Mapping soil cation-exchange capacity using Bayesian modeling and proximal sensors at the field scale. Soil Sci. Soc. Am. J. 2018, 82, 1203–1216. [Google Scholar] [CrossRef]
- Panta, S.; Doyle, R.; Hardie, M.; Lane, P.; Flowers, T.; Haros, G.; Shabala, S. Can highly saline irrigation water improve sodicity and alkalinity in sodic clayey subsoils? J. Soils Sediments 2018, 18, 3290–3302. [Google Scholar] [CrossRef]
- Abolmaali, S.M.-R.; Tarkesh, M.; Bashari, H. MaxEnt modeling for predicting suitable habitats and identifying the effects of climate change on a threatened species, Daphne mucronata, in central Iran. Ecol. Inform. 2018, 43, 116–123. [Google Scholar] [CrossRef]
- Gallardo, B.; Aldridge, D.C. Evaluating the combined threat of climate change and biological invasions on endangered species. Biol. Conserv. 2013, 160, 225–233. [Google Scholar] [CrossRef]
- Boisvert-Marsh, L.; de Blois, S. Unravelling potential northward migration pathways for tree species under climate change. J. Biogeogr. 2021, 48, 1088–1100. [Google Scholar] [CrossRef]
- Tong, X.; Wang, K.; Chen, Z.; Wang, L.; Xiang, T. Endangerment of Ostrya rehderiana Chun and its relationship with rhizosphere soil microflora. Agron. J. 2021, 113, 746–759. [Google Scholar] [CrossRef]
- Gu, D.; Lu, S.; Ba, C.; Li, Y. Technique of callus induction and bud seedling of Ostrya rehderiana Chun. J. Zhejiang Univ. 2013, 40, 216–220. [Google Scholar]
- Li, Y.; Bi, H.; Liu, B.; Guo, X.; Hao, G.; He, Q.; Ma, T. The complete chloroplast genome of Ostrya rehderiana. Mitochondr. DNA Part A 2016, 27, 4536–4537. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Jingmin, J. Germplasm and conservation of rare and endangered tree species endemic to east China. Plant Sci. Today 2014, 1, 183–187. [Google Scholar] [CrossRef] [Green Version]
- Carroll, C.; Roberts, D.R.; Michalak, J.L.; Lawler, J.J.; Nielsen, S.E.; Stralberg, D.; Hamann, A.; Mcrae, B.H.; Wang, T. Scale-dependent complementarity of climatic velocity and environmental diversity for identifying priority areas for conservation under climate change. Glob. Chang. Biol. 2017, 23, 4508–4520. [Google Scholar] [CrossRef]
- Martin, T.G.; Kehoe, L.; Mantyka-Pringle, C.; Chades, I.; Wilson, S.; Bloom, R.G.; Davis, S.K.; Fisher, R.; Keith, J.; Mehl, K. Prioritizing recovery funding to maximize conservation of endangered species. Conserv. Lett. 2018, 11, e12604. [Google Scholar] [CrossRef]
- Zhang, L.; Xu, W.-h.; Ouyang, Z.-y.; Zhu, C.-q. Determination of priority nature conservation areas and human disturbances in the Yangtze River Basin, China. J. Nat. Conserv. 2014, 22, 326–336. [Google Scholar] [CrossRef]
- Ma, B.; Li, X.; Chang, S.X. Capping material type affects rhizosphere bacteria community structure in the cover soil in oil sands reclamation. J. Soils Sediments 2017, 17, 2516–2523. [Google Scholar] [CrossRef]
- Ren, H.; Zhang, Q.; Lu, H.; Liu, H.; Guo, Q.; Wang, J.; Jian, S.; Bao, H.o. Wild plant species with extremely small populations require conservation and reintroduction in China. Ambio 2012, 41, 913–917. [Google Scholar] [CrossRef] [Green Version]
- Vandenbergh, M.P.; Ackerly, B.A.; Forster, F.E. Micro-offsets and macro-transformation: An inconvenient view of climate change justice. Harv. Environ. Law Rev. 2009, 33, 303. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Abbreviation | Variables Description | Abbreviation | Variables Description |
|---|---|---|---|
| MAT | Mean Annual Temperature | DD > 5 | Degree-days above 5 °C, Growing Degree-days |
| MWMT | Mean Warmest Month Temperature | DD < 0 | Degree-days below 0 °C, Chilling Degree-days |
| MCMT | Mean Coldest Month Temperature | NFFD | The Number of Frost-free Days |
| TD | Temperature Difference between MWMT and MCMT, or Continentality | PAS | Precipitation as Snow (mm) between August in Previous Year and July in Current Year |
| MAP | Mean Annual Precipitation | EMT | Extreme Minimum Temperature over 30 years |
| EXT | Extreme Maximum Temperature over 30 years | Eref | Hargreaves reference evaporation |
| AHM | Annual Heat:Moisture index (MAT + 10)/(MAP/1000) | CMD | Hargreaves Climatic Moisture Deficit |
| DD < 18 | Degree-days below 18 °C | DD > 18 | Degree-days above 18 °C |
| T_USDA_TEX_CLASS | Topsoil USDA Texture Classification | T-CEC-CLAY | Topsoil CEC (clay) |
| T-GRAVEL | Topsoil Gravel Content | T-CEC-SOIL | Topsoil CEC (soil) |
| T-SAND | Topsoil Sand Fraction | T-BS | Topsoil Base Saturation |
| T-SILT | Topsoil Silt Fraction | T-TEB | Topsoil TEB |
| T-CLAY | Topsoil Clay Fraction | T-CACO3 | Topsoil Calcium Carbonate |
| T_BULK_DENSITY | Topsoil Bulk Density | T-CASO4 | Topsoil Gypsum |
| T-OC | Topsoil Organic Carbon | T-ESP | Topsoil Sodicity (ESP) |
| T-PH-H2O | Topsoil pH (H2O) | T-ECE | Topsoil Salinity (Elco) |
| T_REF_BULK_DENSITY | Topsoil Reference Bulk Density | HF | Human Footprint Index |
| Category | Variable | Contribution (%) |
|---|---|---|
| Climate | CMD | 45.3 |
| DD < 0 | 39.0 | |
| TD | 14.7 | |
| NFFD | 0.6 | |
| MWMT | 0.2 | |
| MAP | 0.1 | |
| Soil + HF | T_USDA_TEX_CLASS | 76.1 |
| T-CEC-CLAY | 10.3 | |
| T-ESP | 6.3 | |
| T-SAND | 2.5 | |
| T-GRAVEL | 1.1 | |
| T-CACO3 | 1.0 | |
| T-OC | 0.7 | |
| HF | 0.7 | |
| T_BULK_DENSITY | 0.5 | |
| T-SILT | 0.4 | |
| T-CASO4 | 0.2 | |
| T_REF_BULK_DENSITY | 0.1 |
| Classes | Low-Suitable | Medium-Suitable | High-Suitable |
|---|---|---|---|
| Climatic habitats | 45.3 | 22.5 | 23.4 |
| Soil + HF habitats | 537.8 | 47.0 | 76.2 |
| 2050s—RCP4.5 | 43.3 | 23.5 | 24.3 |
| Scenarios | Soil and HF Habitats | Climate-Soil and HF Overlap Habitats | Overlap Habitats as a Percentage of Climate | Overlap Habitats as a Percentage of Soil + HF |
|---|---|---|---|---|
| (×105 km2) | (×105 km2) | (%) | (%) | |
| Current climatic habitats | 45.8 | 123.1 | 55 | 19 |
| 2050s-RCP4.5 | 47.8 | 123.1 | 57 | 21 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Feng, L.; Sun, J.; El-Kassaby, Y.A.; Yang, X.; Tian, X.; Wang, T. Predicting Potential Habitat of a Plant Species with Small Populations under Climate Change: Ostryarehderiana. Forests 2022, 13, 129. https://0-doi-org.brum.beds.ac.uk/10.3390/f13010129
Feng L, Sun J, El-Kassaby YA, Yang X, Tian X, Wang T. Predicting Potential Habitat of a Plant Species with Small Populations under Climate Change: Ostryarehderiana. Forests. 2022; 13(1):129. https://0-doi-org.brum.beds.ac.uk/10.3390/f13010129
Chicago/Turabian StyleFeng, Lei, Jiejie Sun, Yousry A. El-Kassaby, Xianyu Yang, Xiangni Tian, and Tongli Wang. 2022. "Predicting Potential Habitat of a Plant Species with Small Populations under Climate Change: Ostryarehderiana" Forests 13, no. 1: 129. https://0-doi-org.brum.beds.ac.uk/10.3390/f13010129