
Tree Species and Stand Density: The Effects on Soil Organic Matter Contents, Decomposability and Susceptibility to Microbial Priming



Abstract
:1. Introduction
2. Materials and Methods
2.1. Research Sites and Experimental Setup
2.2. Soil Sample Preparation and Analysis
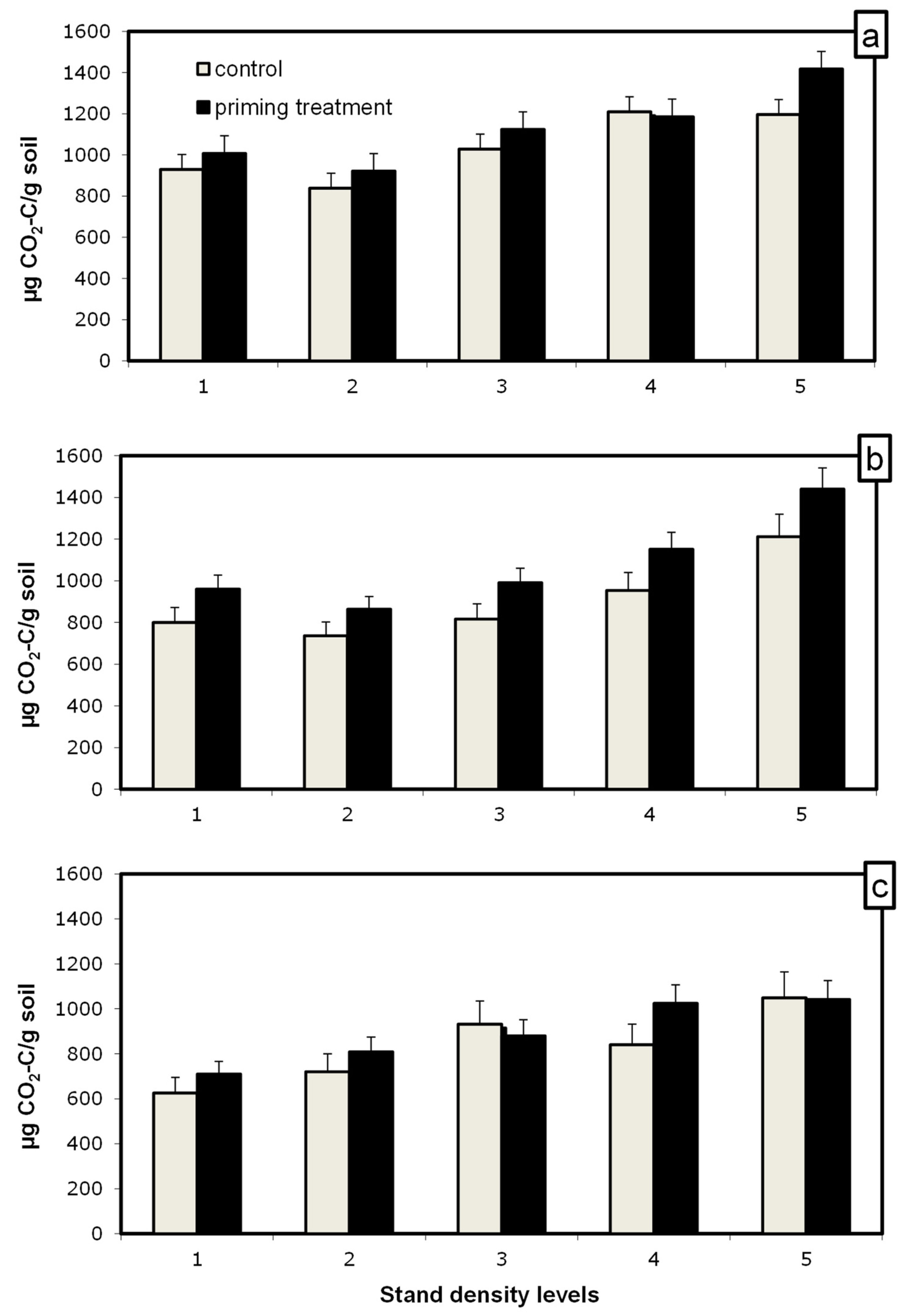
2.3. Determining the Priming of Soil Organic Matter and C Decomposability
2.4. Priming Calculation
2.5. Statistical Data Analysis
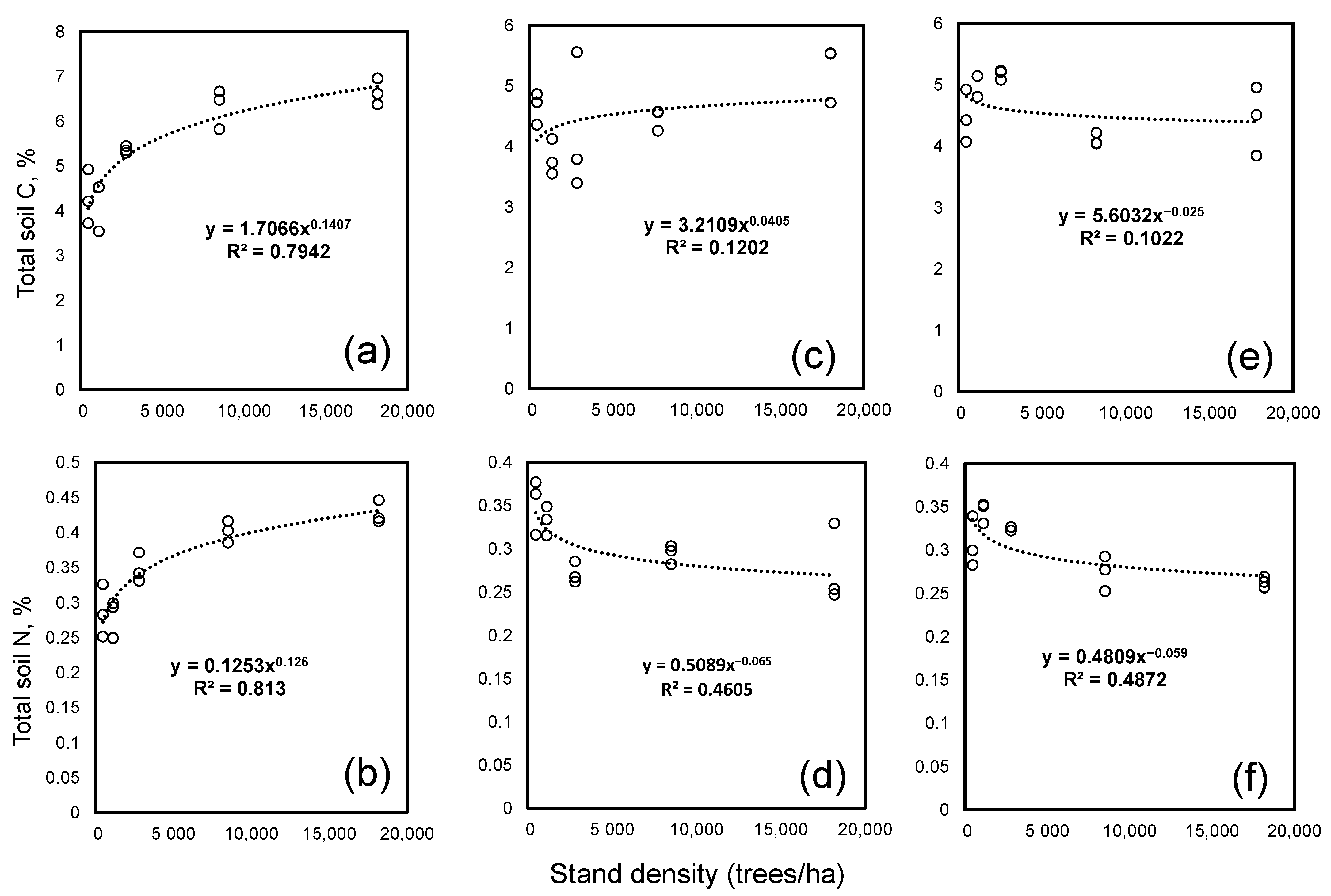
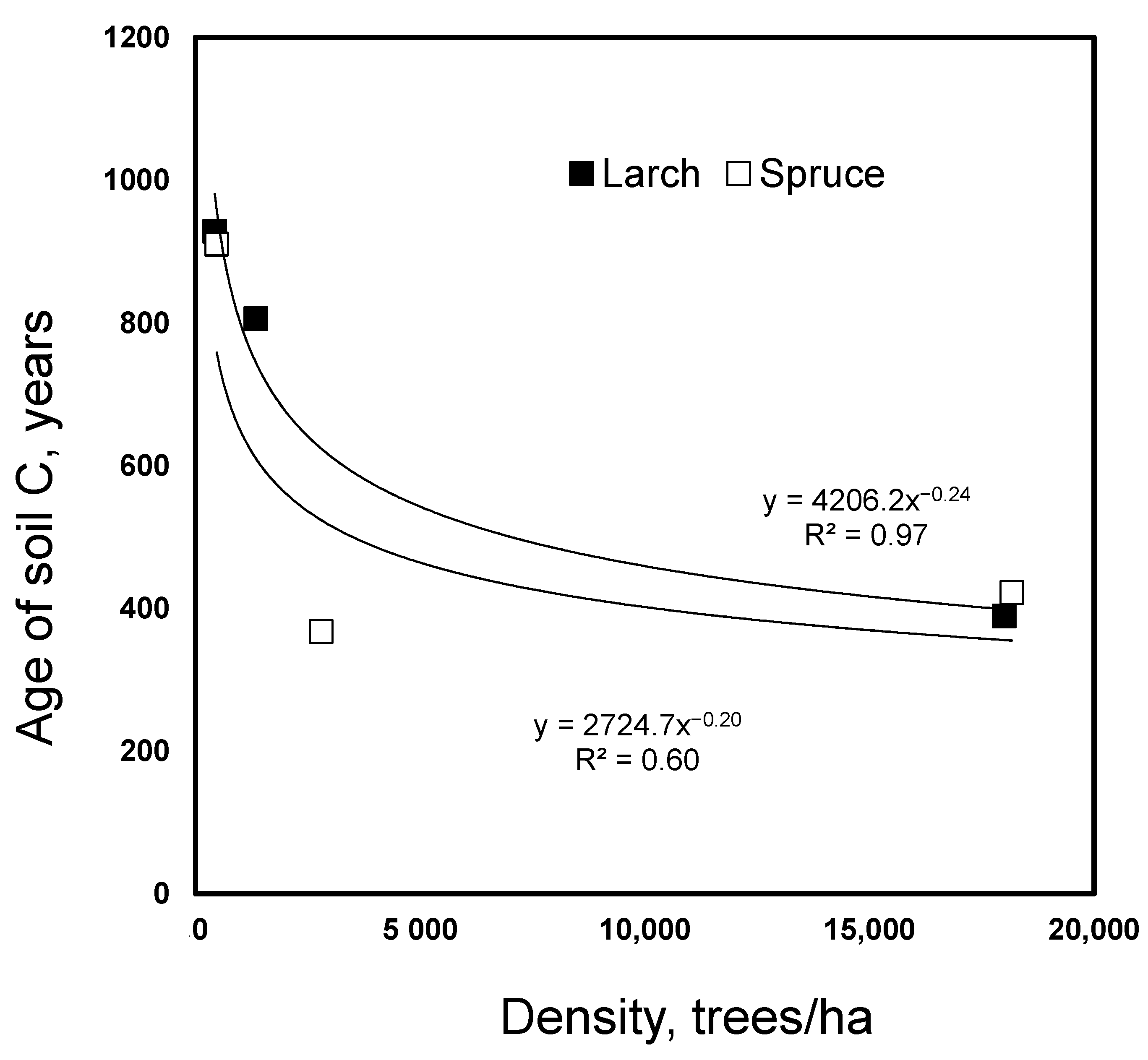
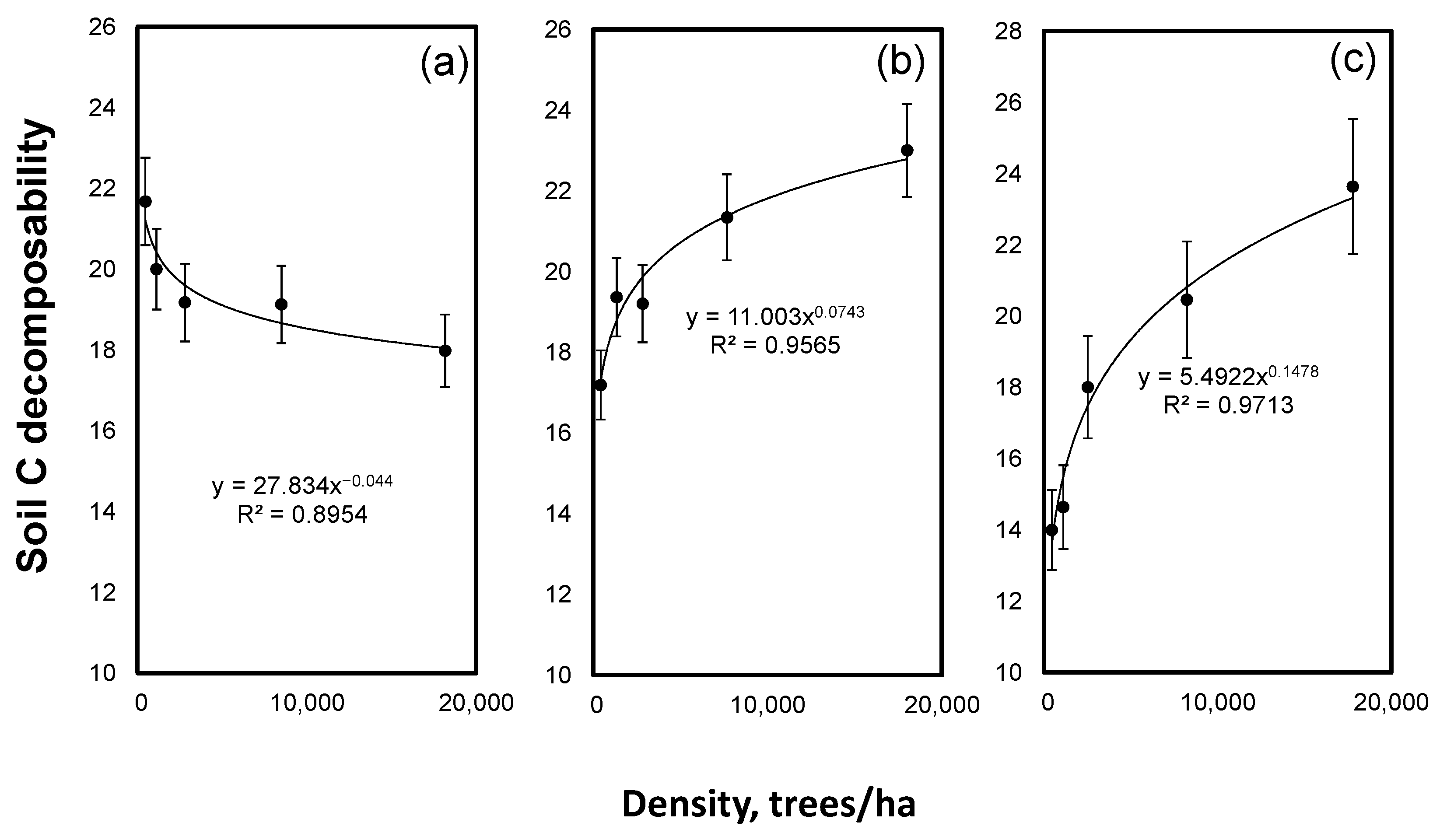
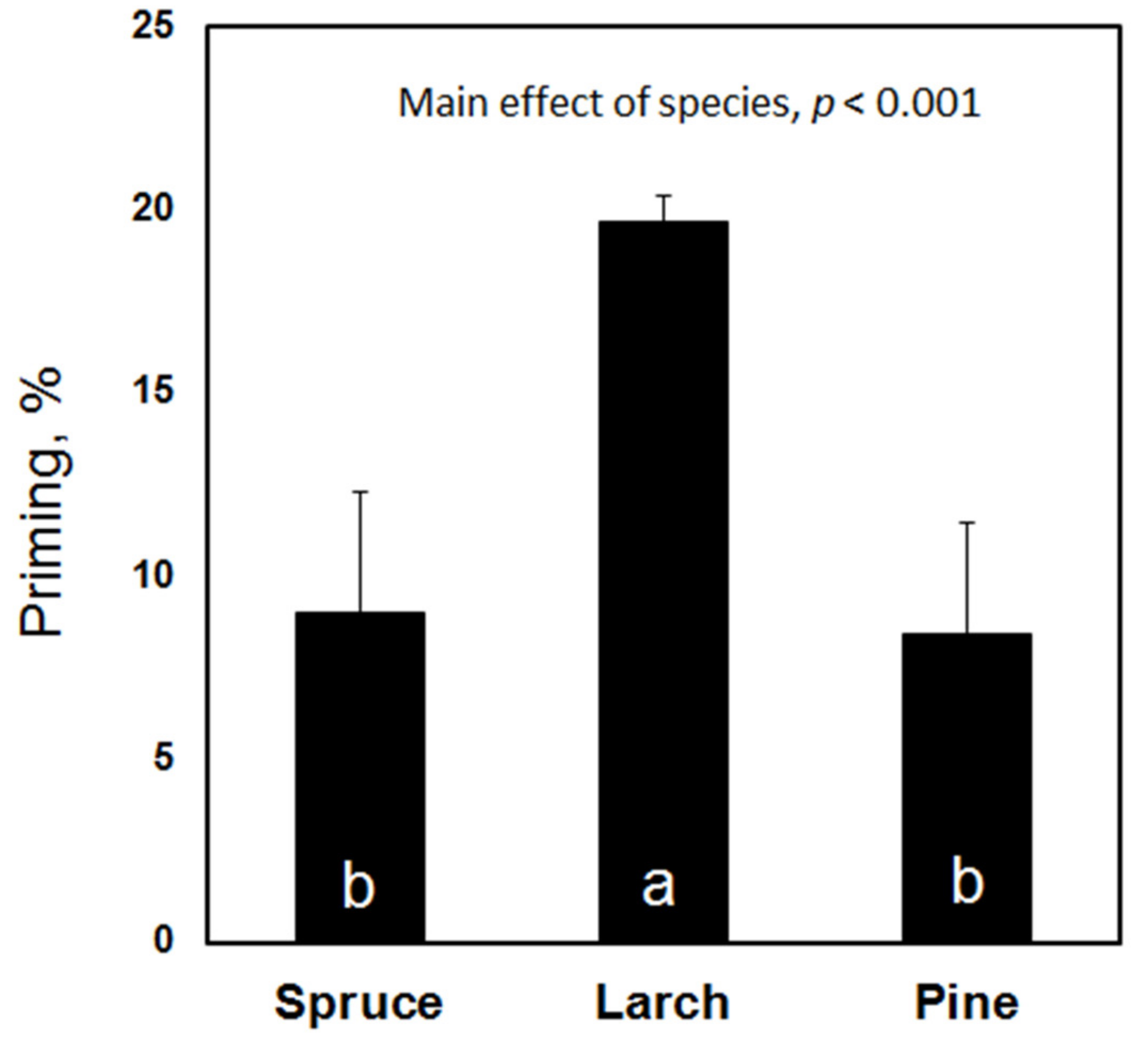
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Mayer, M.; Prescott, C.E.; Abaker, W.E.A.; Augusto, L.; Cécillon, L.; Ferreira, G.W.D.; James, J.; Jandl, R.; Katzensteiner, K.; Laclau, J.-P.; et al. Tamm Review: Influence of forest management activities on soil organic carbon stocks: A knowledge synthesis. For. Ecol. Manag. 2020, 466, 118–127. [Google Scholar] [CrossRef]
- Laganiere, J.; Angers, D.A.; Pare, D. Carbon accumulation in agricultural soils after afforestation: A meta-analysis. Glob. Chang. Biol. 2010, 16, 439–453. [Google Scholar] [CrossRef]
- Sun, X.; Sun, H.; Wang, W.; Razaq, M. Effects of stand density on soil organic carbon storage in the top and deep soil layers of Fraxinus mandshurica plantations. Austrian J. For. Sci. 2019, 136, 27–44. [Google Scholar]
- Farooq, T.; Ma, X.; Rashid, M.; Wu, W.; Xu, J.; Tarin, M.; He, Z.; Wu, P. Impact of stand density on soil quality in Chinese fir (Cunninghamia lanceolate) monoculture. Appl. Ecol. Environ. Res. 2019, 17, 3553–3566. [Google Scholar] [CrossRef]
- Truax, B.; Fortier, J.; Gagnon, D.; Lambert, F. Planting density and site effects on stem dimensions, stand productivity, biomass partitioning, carbon stocks and soil nutrient supply in hybrid poplar plantations. Forests 2018, 9, 293. [Google Scholar] [CrossRef] [Green Version]
- Hernández, J.; del Pino, A.; Vance, E.D.; Califra, Á.; Del Giorgio, F.; Martínez, L.; González-Barrios, P. Eucalyptus and Pinus stand density effects on soil carbon sequestration. For. Ecol. Manag. 2016, 368, 28–38. [Google Scholar] [CrossRef]
- González, I.G.; Corbí, J.G.; Cancio, A.F.; Ballesta, R.J.; Cascón, M.G. Soil carbon stocks and soil solution chemistry in Quercus ilex stands in Mainland Spain. Eur. J. For. Res. 2012, 131, 1653–1667. [Google Scholar] [CrossRef]
- Noh, N.J.; Kim, C.; Bae, S.W.; Lee, W.K.; Yoon, T.K.; Muraoka, H.; Son, Y. Carbon and nitrogen dynamics in a Pinus densiflora forest with low and high stand densities. J. Plant Ecol. 2013, 6, 368–379. [Google Scholar] [CrossRef] [Green Version]
- Cécillon, L.; Soucémarianadin, L.N.; Berthelot, A.; Duverger, M.; De Boisseson, J.M.; Gosselin, F.; Guenet, B.; Barthès, B.; De Danieli, S.; Barrier, R.; et al. piCaSo: Pilotage sylvicole et contrôle pédologique des stocks de carbone des sols forestiers. Rep. ADEME 2017, 103. Available online: https://0-www-scopus-com.brum.beds.ac.uk/record/display.uri?eid=2-s2.0-85082819757&origin=inward&txGid=f5b2dbd15c3d3d529b5291c5fe536486 (accessed on 30 January 2022).
- Vesterdal, L.; Dalsgaard, M.; Felby, C.; Raulund-Rasmussen, K.; Jørgensen, B.B. Effects of thinning and soil properties on accumulation of carbon, nitrogen and phosphorus in the forest floor of Norway spruce stands. For. Ecol. Manag. 1995, 77, 1–10. [Google Scholar] [CrossRef]
- Skovsgaard, J.P.; Stupak, I.; Vesterdal, L. Distribution of biomass and carbon in even-aged stands of Norway spruce (Picea abies (L.) Karst.): A case study on spacing and thinning effects in northern Denmark. Scand. J. For. Res. 2006, 21, 470–488. [Google Scholar] [CrossRef]
- Jandl, R.; Lindner, M.; Vesterdal, L.; Bauwens, B.; Baritz, R.; Hagedorn, F.; Johnson, D.W.; Minkkinen, K.; Byrne, K.A. How strongly can forest management influence soil carbon sequestration? Geoderma 2007, 137, 253–268. [Google Scholar] [CrossRef]
- Hoover, C.M. Management impacts on forest floor and soil organic carbon in northern temperate forests of the US. Carbon Balance Manag. 2011, 6, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Powers, M.; Kolka, R.; Palik, B.; McDonald, R.; Jurgensen, M. Long-term management impacts on carbon storage in Lake States forests. For. Ecol. Manag. 2011, 262, 424–431. [Google Scholar] [CrossRef]
- Jurgensen, M.; Tarpey, R.; Pickens, J.; Kolka, R.; Palik, B. Long-term Effect of Silvicultural Thinnings on Soil Carbon and Nitrogen Pools. Soil Sci. Soc. Am. J. 2012, 76, 1418–1425. [Google Scholar] [CrossRef]
- Cheng, X.; Yu, M.; Wu, T. Effect of forest structural change on carbon storage in a coastal Metasequoia glyptostroboides stand. Sci. World J. 2013, 2013, 830509. [Google Scholar] [CrossRef]
- Ruiz-Peinado, R.; Bravo-Oviedo, A.; López-Senespleda, E.; Montero, G.; Río, M. Do thinnings influence biomass and soil carbon stocks in Mediterranean maritime pinewoods? Eur. J. For. Res. 2013, 132, 253–262. [Google Scholar] [CrossRef]
- Zhou, D.; Zhao, S.; Liu, S.; Oeding, J. A meta-analysis on the impacts of partial cutting on forest structure and carbon storage. Biogeosciences 2013, 10, 3691–3703. [Google Scholar] [CrossRef] [Green Version]
- Achat, D.L.; Fortin, M.; Landmann, G.; Ringeval, B.; Augusto, L. Forest soil carbon is threatened by intensive biomass harvesting. Sci. Rep. 2015, 5, 15991. [Google Scholar] [CrossRef]
- Noormets, A.; Epron, D.; Domec, J.C.; McNulty, S.G.; Fox, T.; Sun, G.; King, J.S. Effects of forest management on productivity and carbon sequestration: A review and hypothesis. For. Ecol. Manag. 2015, 355, 124–140. [Google Scholar] [CrossRef] [Green Version]
- Strukelj, M.; Brais, S.; Paré, D. Nine-year changes in carbon dynamics following different intensities of harvesting in boreal aspen stands. Eur. J. For. Res. 2015, 134, 737–754. [Google Scholar] [CrossRef]
- Kim, S.; Han, S.H.; Li, G.; Yoon, T.K.; Lee, S.-T.; Kim, C.; Son, Y. Effects of thinning intensity on nutrient concentration and enzyme activity in Larix kaempferi forest soils. J. Ecol. Environ. 2016, 40, 2. [Google Scholar] [CrossRef] [Green Version]
- Mattson, K.G.; Smith, H.C. Detrital organic matter and soil CO2 efflux in forests regenerating from cutting in West Virginia. Soil Biol. Biochem. 1993, 25, 1241–1248. [Google Scholar] [CrossRef]
- Chiti, T.; Perugini, L.; Vespertino, D.; Valentini, R. Effect of selective logging on soil organic carbon dynamics in tropical forests in central and western Africa. Plant Soil 2015, 399, 283–294. [Google Scholar] [CrossRef]
- Moreno-Fernández, D.; Díaz-Pinés, E.; Barbeito, I.; Sánchez-González, M.; Montes, F.; Rubio, A.; Cañellas, I. Temporal carbon dynamics over the rotation period of two alternative management systems in Mediterranean mountain Scots pine forests. For. Ecol. Manag. 2015, 348, 186–195. [Google Scholar] [CrossRef]
- Mushinski, R.M.; Gentry, T.J.; Boutton, T.W. Forest organic matter removal leads to long-term reductions in bacterial and fungal abundance. Appl. Soil Ecol. 2019, 137, 106–110. [Google Scholar] [CrossRef]
- Gross, C.D.; James, J.N.; Turnblom, E.C.; Harrison, R.B. Thinning Treatments Reduce Deep Soil Carbon and Nitrogen Stocks in a Coastal Pacific Northwest Forest. Forests 2018, 9, 238. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Guan, D.; Li, W.; Sun, D.; Jin, C.; Yuan, F.; Wang, A.; Wu, J. The effects of forest thinning on soil carbon stocks and dynamics: A meta-analysis. For. Ecol. Manag. 2018, 429, 36–43. [Google Scholar] [CrossRef]
- Cepáková, Š.; Tošner, Z.; Frouz, J. The effect of tree species on seasonal fluctuations in water-soluble and hot water-extractable organic matter at post-mining sites. Geoderma 2016, 275, 19–27. [Google Scholar] [CrossRef]
- Mueller, K.E.; Hobbie, S.; Chorover, J.; Reich, P.; Eisenhauer, N.; Castellano, M.; Chadwick, O.A.; Dobies, T.; Hale, C.M.; Jagodzinski, A.; et al. Effects of litter traits, soil biota, and soil chemistry on soil carbon stocks at a common garden with 14 tree species. Biogeochemistry 2015, 123, 313–327. [Google Scholar] [CrossRef]
- Menyailo, O.V.; Hungate, B.A.; Zech, W. Tree species mediated soil chemical changes in a Siberian artificial afforestation experiment. Plant Soil 2002, 242, 171–182. [Google Scholar] [CrossRef]
- Menyailo, O.V.; Hungate, B.A.; Zech, W. The effect of single tree species on soil microbial activities related to C and N cycling in the Siberian artificial afforestation experiment. Plant Soil 2002, 242, 183–196. [Google Scholar] [CrossRef]
- Marschner, B.; Brodowski, S.; Dreves, A.; Gleixner, G.; Gude, A.; Grootes, P.M.; Hamer, U.; Heim, A.; Jandl, G.; Ji, R.; et al. How relevant is recalcitrance for the stabilization of organic matter in soils? J. Plant Nutr. Soil Sci. 2008, 171, 91–110. [Google Scholar] [CrossRef] [Green Version]
- Vesterdal, L.; Elberling, B.; Christiansen, J.R.; Callesen, I.; Schmidt, I.K. Soil respiration and rates of soil carbon turnover differ among six common European tree species. For. Ecol. Manag. 2012, 264, 185–196. [Google Scholar] [CrossRef]
- Plante, A.F.; Conant, R.T.; Carlson, J.; Greenwood, R.; Shulman, J.M.; Haddix, M.L.; Paul, E.A. Decomposition temperature sensitivity of isolated soil organic matter fractions. Soil Biol. Biochem. 2010, 42, 1991–1996. [Google Scholar] [CrossRef]
- Paterson, E.; Thornton, B.; Midwood, A.J.; Osborne, S.M.; Sim, A.; Millard, P. Atmospheric CO2 enrichment and nutrient additions to planted soil increase mineralisation of soil organic matter. Soil Biol. Biochem. 2008, 40, 2434–2440. [Google Scholar] [CrossRef]
- Taneva, L.; Gonzalez-Meler, M.A. Decomposition kinetics of soil C of different age from a forest exposed to 8 years of elevated atmospheric CO2 concentration. Soil. Biol. Biochem. 2008, 40, 2670–2677. [Google Scholar] [CrossRef]
- Wutzler, T.; Reichstein, M. Colimitation of decomposition by substrates and decomposers—A comparison of model formulations. Biogeosciences 2008, 5, 749–759. [Google Scholar] [CrossRef] [Green Version]
- Chapin, F.S.; McFarland, J.; McGuire, A.D.; Euskirchen, E.S.; Ruess, R.W.; Kielland, K. The changing global C cycle: Linking plant-soil C dynamics to global consequences. J. Ecol. 2009, 97, 840–850. [Google Scholar]
- Sobachkin, R.S.; Sobachkin, D.S.; Buzykin, A.I. The influence of stand density on growth of tree conifer species. In Trees and Soil Interactions: Implications to Global Change; Binkley, D., Menyailo, O.V., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2005; pp. 247–255. [Google Scholar]
- Hurlbert, S.H. Pseudoreplication and the design of ecological field experiments. Ecol. Monogr. 1984, 54, 187–211. [Google Scholar] [CrossRef] [Green Version]
- Hamer, U.; Marschner, B. Priming effects in different soil types induced by fructose, alanine, oxalic acid and catechol additions. Soil Biol. Biochem. 2005, 37, 445–454. [Google Scholar] [CrossRef]
- Liu, X.-J.A.; Sun, J.; Mau, R.L.; Finley, B.K.; Compson, Z.G.; van Gestel, N.; Brown, J.R.; Schwartz, E.; Dijkstra, P.; Hungate, B.A. Labile carbon input determines the direction and magnitude of the priming effect. Appl. Soil Ecol. 2017, 109, 7–13. [Google Scholar] [CrossRef] [Green Version]
- Qiao, N.; Schaefer, D.; Blagodatskaya, E.; Zou, X.; Xu, X.; Kuzyakov, Y. Labile carbon retention compensates for CO2 released by priming in forest soils. Glob. Chang. Biol. 2014, 20, 1943–1954. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Xu, W.; Hu, G.; Dai, W.; Jiang, P.; Bai, E. The priming effect of soluble carbon inputs in organic and mineral soils from a temperate forest. Oecologia 2015, 178, 1239–1250. [Google Scholar] [CrossRef]
- StatSoft. Statistica for Windows (Computer Program Manual); StatSoft: Tulsa, OK, USA, 1997. [Google Scholar]
- Menyailo, O.V.; Abraham, W.R.; Conrad, R. Tree species affect atmospheric CH4 oxidation without altering community composition of soil methanotrophs. Soil Biol. Biochem. 2010, 42, 101–107. [Google Scholar] [CrossRef]
- Menyailo, O.V.; Hungate, B.A. Interactive effects of tree species and soil moisture on methane consumption. Soil Biol. Biochem 2003, 35, 625–628. [Google Scholar] [CrossRef]
- Angst, G.; Mueller, K.E.; Eissenstat, D.M.; Trumbore, S.; Freeman, K.H.; Hobbie, S.E.; Chorover, J.; Oleksyn, J.; Reich, P.B.; Mueller, C.W. Soil organic carbon stability in forests: Distinct effects of tree species identity and traits. Glob. Chang. Biol 2019, 25, 1529–1546. [Google Scholar] [CrossRef]
- Vesterdal, L.; Schmidt, I.K.; Callesen, I.; Nilsson, L.O.; Gundersen, P. Carbon and nitrogen in forest floor and mineral soil under six common European tree species. For. Ecol. Manag. 2008, 255, 35–48. [Google Scholar] [CrossRef]
- Litton, C.M.; Ryan, M.G.; Knight, D.H. Effects of tree density and stand age on carbon allocation patterns in postfire lodgepole pine. Ecol. Appl. 2004, 14, 460–475. [Google Scholar] [CrossRef] [Green Version]
- Berger, T.W.; Neubauer, C.; Glatzel, G. Factors controlling soil carbon and nitrogen stores in pure stands of Norway spruce (Picea abies) and mixed species stands in Austria. For. Ecol. Manag. 2002, 159, 3–14. [Google Scholar] [CrossRef]
- Hagen-Thorn, A.; Callesen, I.; Armolaitis, K.; Nihlgård, B. The impact of six European tree species on the chemistry of mineral topsoil in forest plantations on former agricultural land. For. Ecol. Manag. 2004, 195, 373–384. [Google Scholar] [CrossRef]
- Hansson, K.; Olsson, B.A.; Olsson, M.; Johansson, U.; Berggren Kleja, D. Differences in soil properties in adjacent stands. For. Ecol. Manag. 2011, 262, 522–530. [Google Scholar] [CrossRef]
- Pastor, J.; Post, W.M. Response of northern forests to CO2-induced climate change. Nature 1988, 334, 55–58. [Google Scholar] [CrossRef]
- Fekete, I.; Lajtha, K.; Kotroczó, Z.; Várbíró, G.; Varga, C.; Tóth, J.A.; Demeter, I.; Veperdi, G.; Berki, I. Long-term effects of climate change on carbon storage and tree species composition in a dry deciduous forest. Glob. Chang. Biol. 2017, 23, 3154–3168. [Google Scholar] [CrossRef]
- Krankina, O.N.; Dixon, R.K.; Kirilenko, A.P.; Kobak, K.I. Global Climate Change Adaptation: Examples from Russian Boreal Forests. Clim. Chang. 1997, 36, 197–215. [Google Scholar] [CrossRef]
- Kharuk, V.I.; Dvinskaya, M.L.; Ranson, K.J.; Im, S.T. Expansion of Evergreen Conifers to the Larch-Dominated Zone and Climatic Trends. Russ. J. Ecol. 2005, 36, 164–170. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Our Numbering | Initial Density at Planting (Trees/ha) | Current Density (Trees/ha) | ||
|---|---|---|---|---|
| Spruce | Larch | Scots Pine | ||
| 1 | 500 | 470 | 426 | 457 |
| 2 | 2500 | 1118 | 1342 | 1124 |
| 3 | 8000 | 2793 | 2838 | 2523 |
| 4 | 40,000 | 9495 | 7667 | 8236 |
| 5 | 125,000 | 18,170 | 18,000 | 17,795 |
| Soil Chemical Properties | Two-Way ANOVA | One-Way ANOVA, Density Effect Within Different Tree Species | ||||
|---|---|---|---|---|---|---|
| Tree Species | Density | Tree Species × Density Interactions | Spruce | Larch | Scots Pine | |
| Soil C contents | <0.001 | <0.001 | <0.001 | <0.001 | 0.109 | 0.019 |
| Soil N contents | <0.001 | <0.001 | <0.001 | <0.001 | 0.016 | <0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Menyailo, O.V.; Sobachkin, R.S.; Makarov, M.I.; Cheng, C.-H. Tree Species and Stand Density: The Effects on Soil Organic Matter Contents, Decomposability and Susceptibility to Microbial Priming. Forests 2022, 13, 284. https://0-doi-org.brum.beds.ac.uk/10.3390/f13020284
Menyailo OV, Sobachkin RS, Makarov MI, Cheng C-H. Tree Species and Stand Density: The Effects on Soil Organic Matter Contents, Decomposability and Susceptibility to Microbial Priming. Forests. 2022; 13(2):284. https://0-doi-org.brum.beds.ac.uk/10.3390/f13020284
Chicago/Turabian StyleMenyailo, Oleg V., Roman S. Sobachkin, Mikhail I. Makarov, and Chih-Hsin Cheng. 2022. "Tree Species and Stand Density: The Effects on Soil Organic Matter Contents, Decomposability and Susceptibility to Microbial Priming" Forests 13, no. 2: 284. https://0-doi-org.brum.beds.ac.uk/10.3390/f13020284