
Identification of Sieve Element Occlusion Gene (SEOs) Family in Rubber Trees (Hevea brasiliensis Muell. Arg.) Provides Insights to the Mechanism of Laticifer Plugging


Abstract
:1. Introduction
2. Materials and Methods
2.1. Identification of Rubber Tree SEO Genes
2.2. Sequence Alignment and Phylogenetic Analysis of HbSEOs
2.3. Basic Informations and Sequence Analysis of the HbSEOs
2.4. RNA Isolation and Real-Time Quantitative Analysis
3. Results
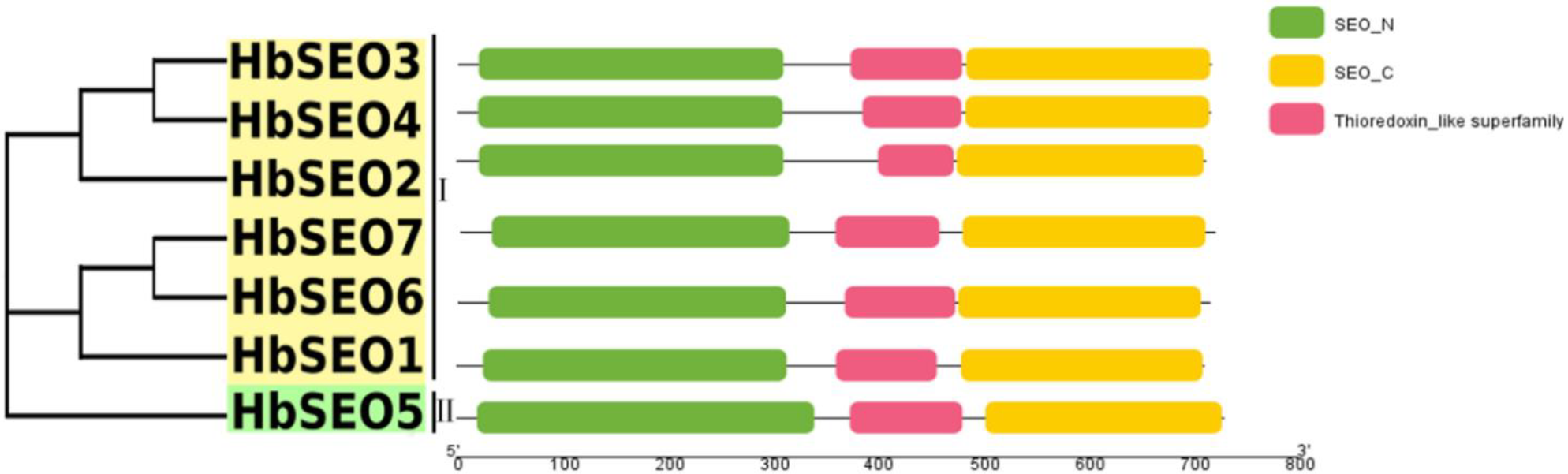
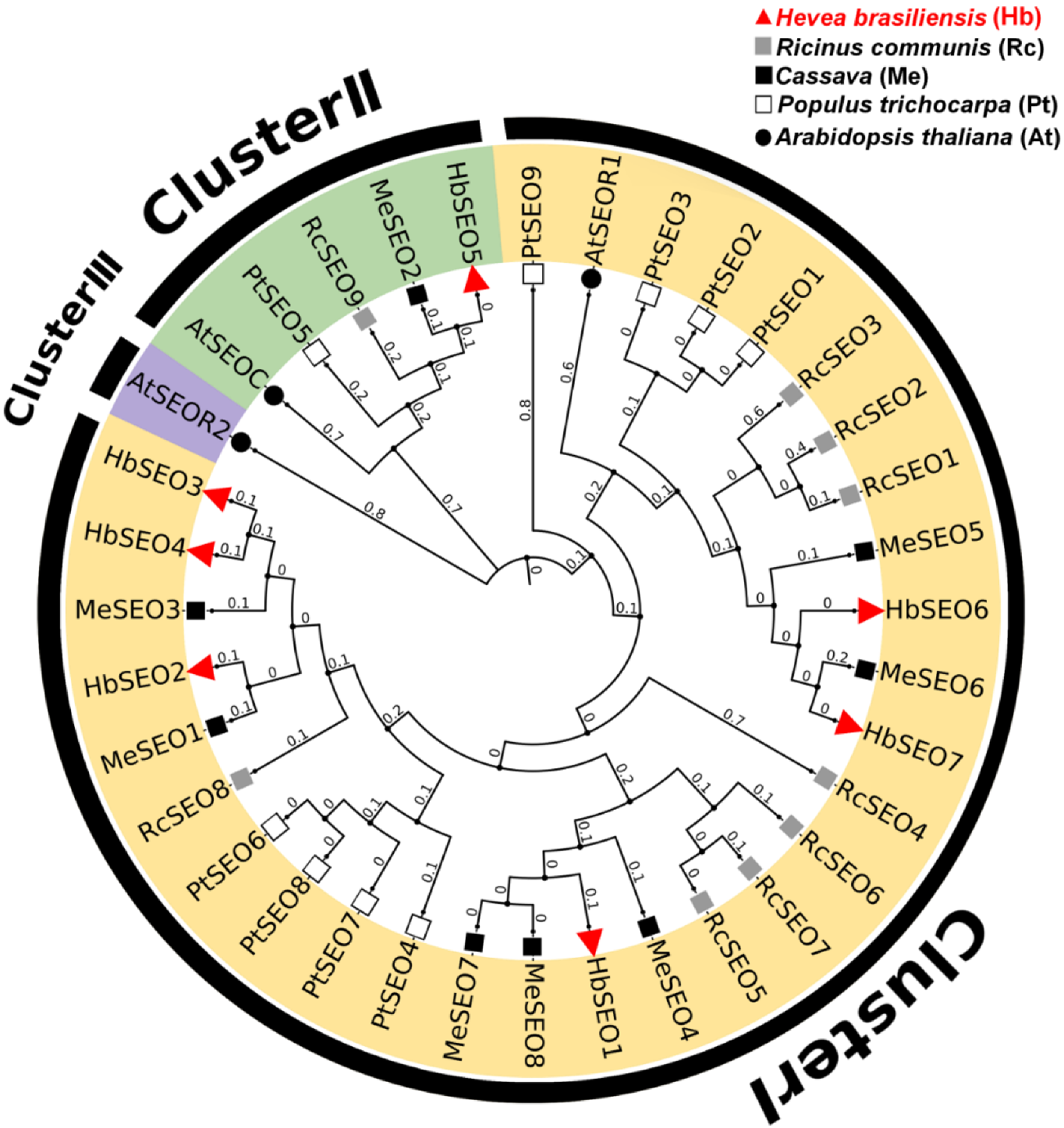
3.1. Identification, Classification, and Protein Property Analysis of HbSEOs
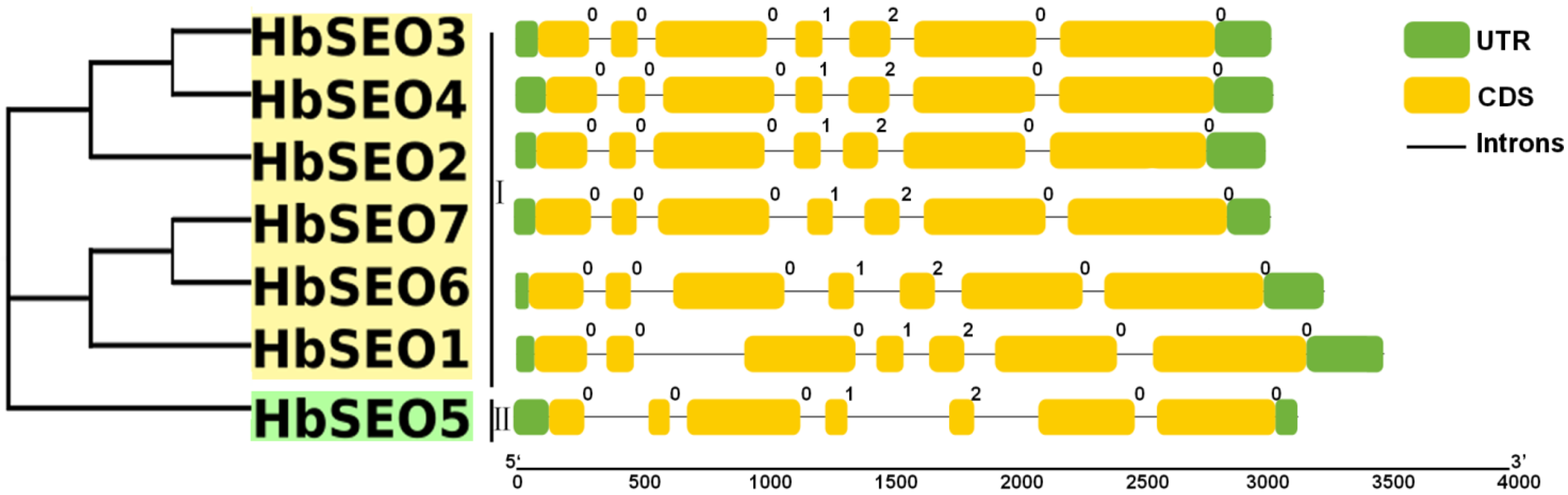
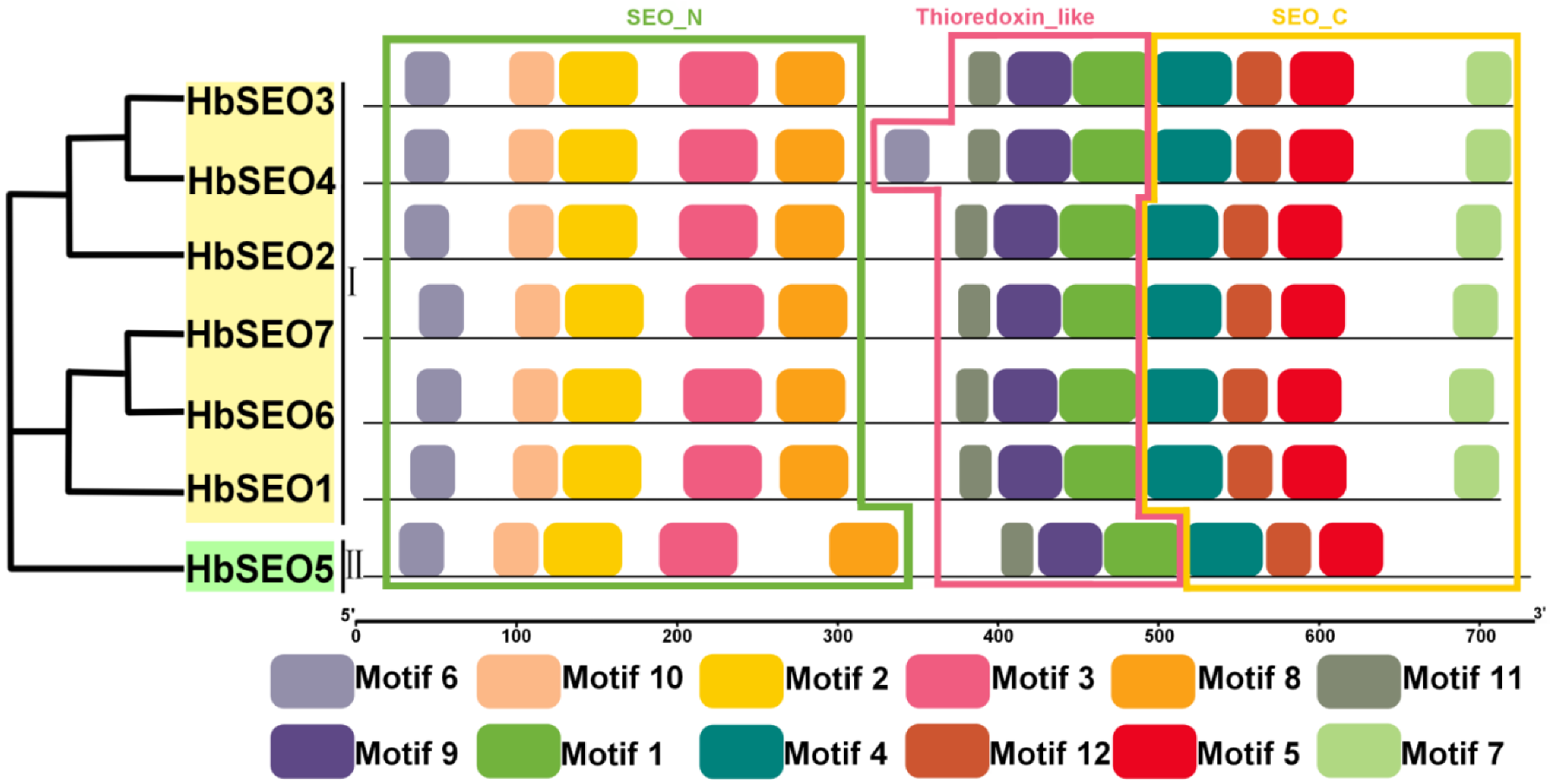
3.2. Gene Structures and Conserved Motifs of HbSEOs
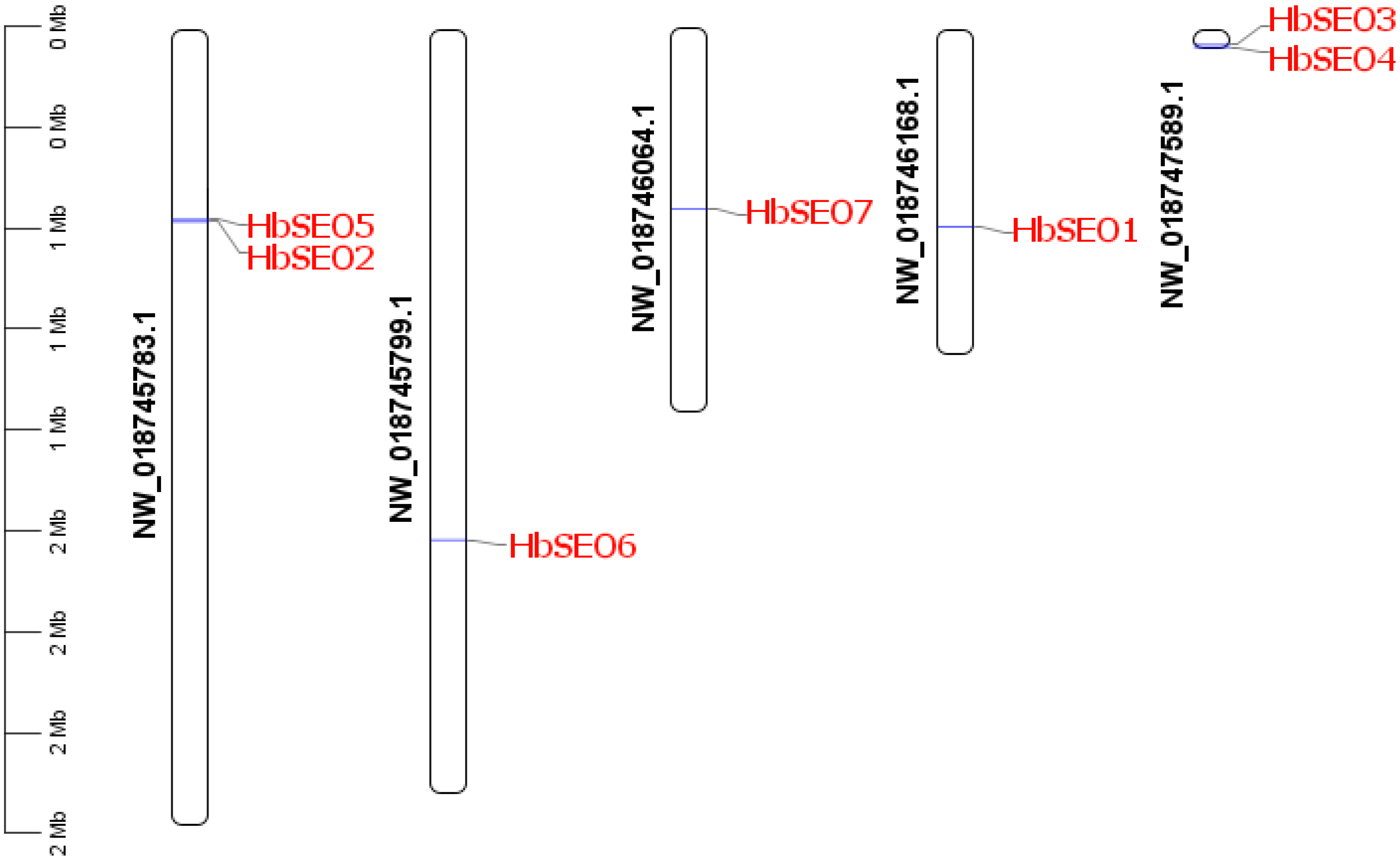
3.3. Chromosome Localization
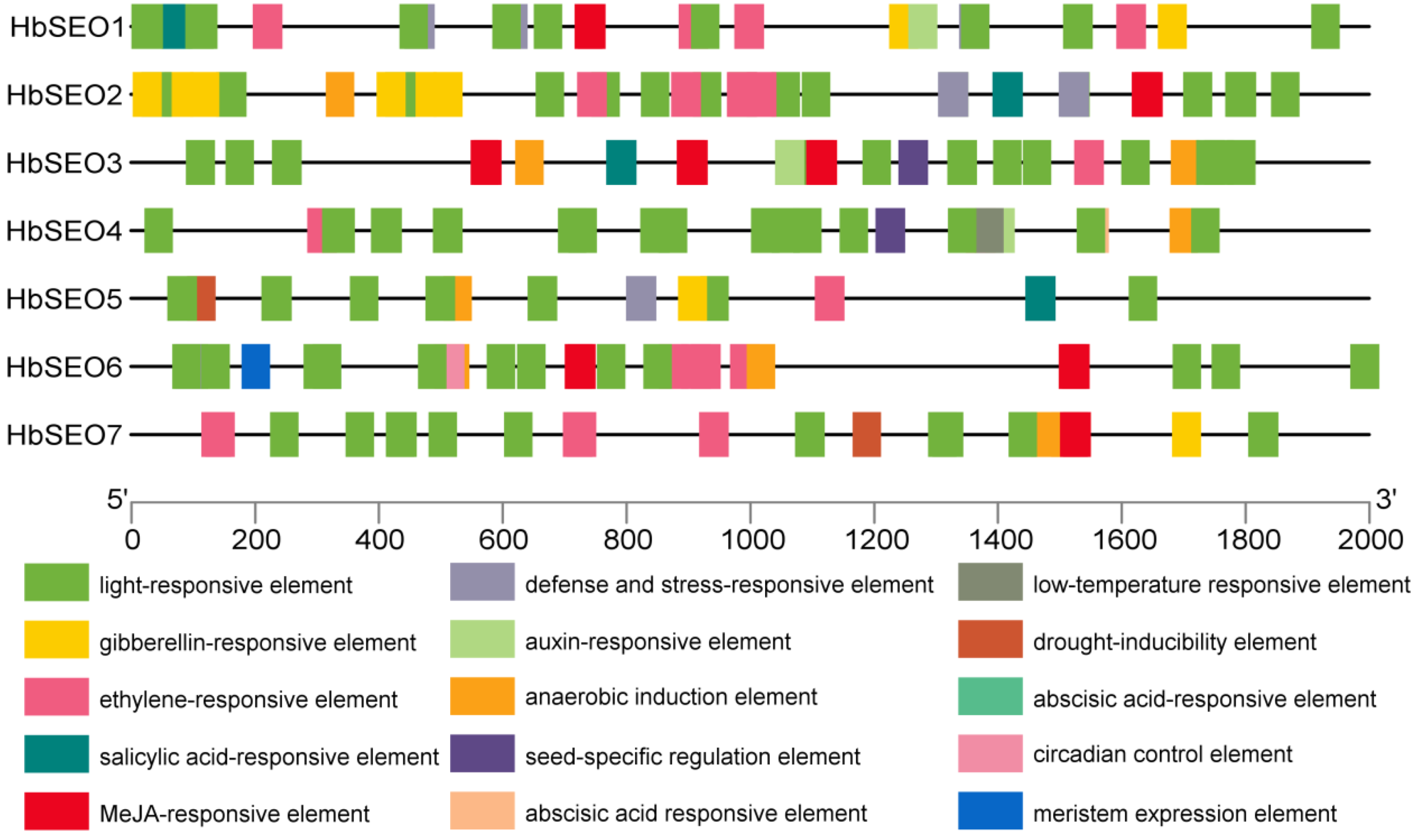
3.4. Analysis of Cis-Acting Elements in Promoter Region
3.5. Expression Analysis of HbSEOs
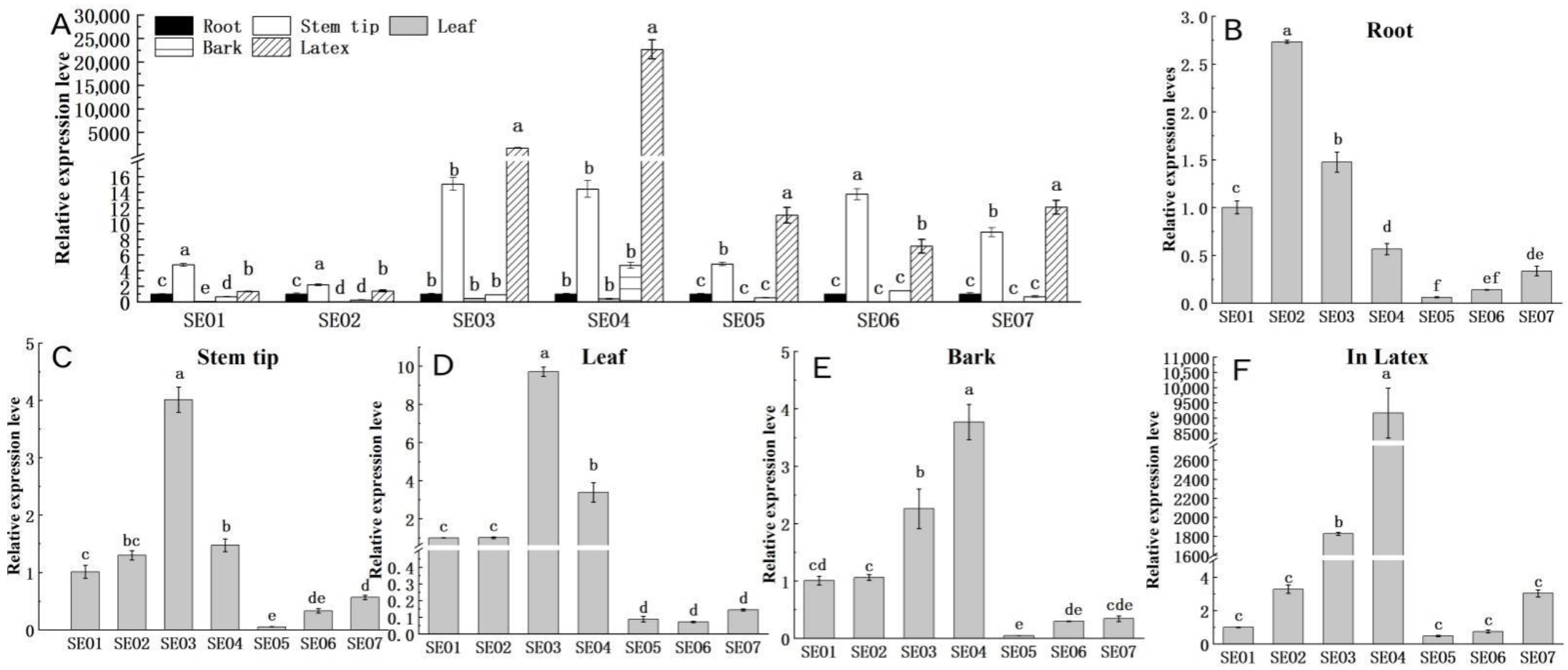
3.5.1. Tissue-Specific Expressions of HbSEOs
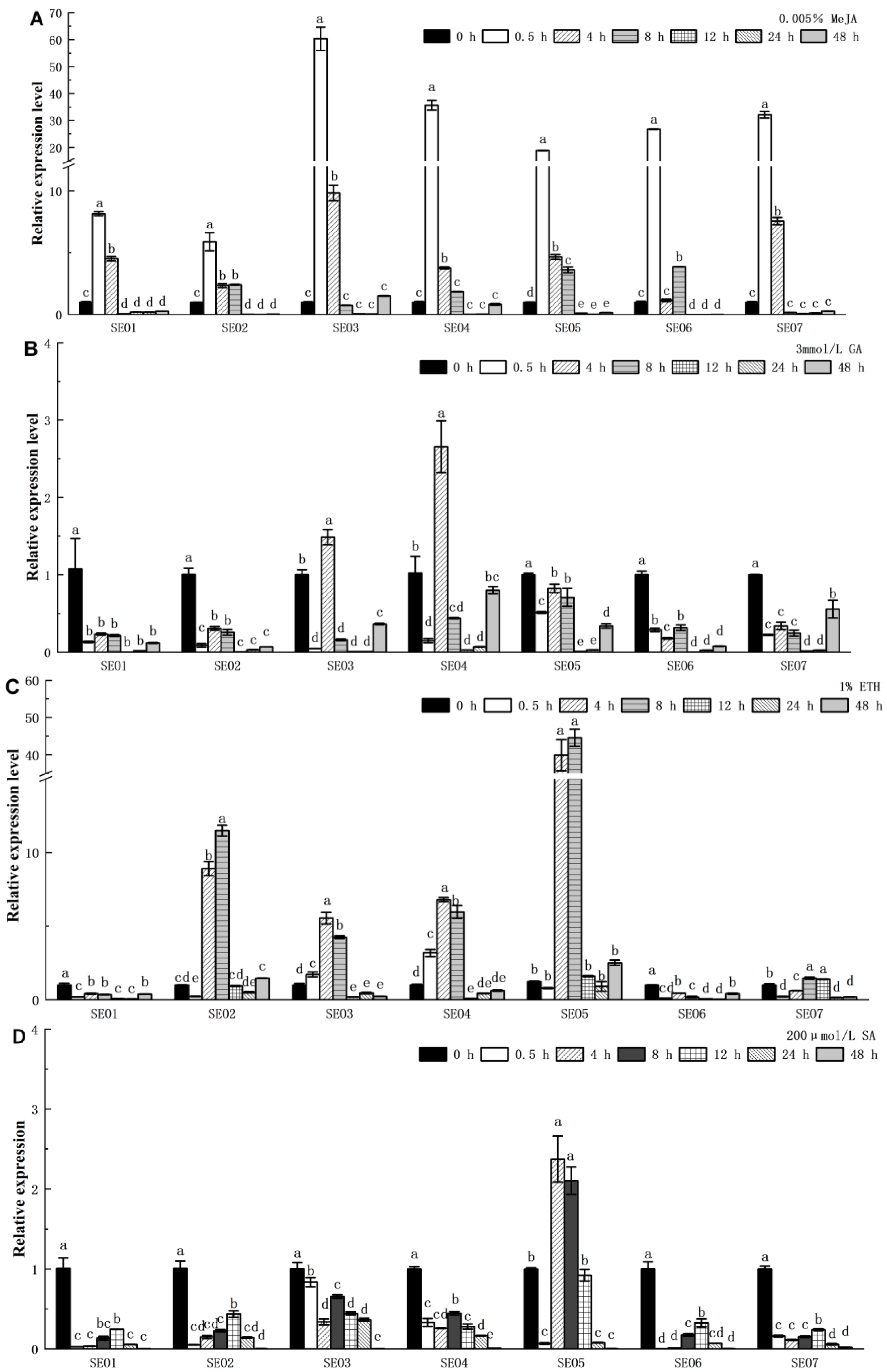
3.5.2. Expression Patterns of HbSEOs Subjected to Various Hormone Treatments
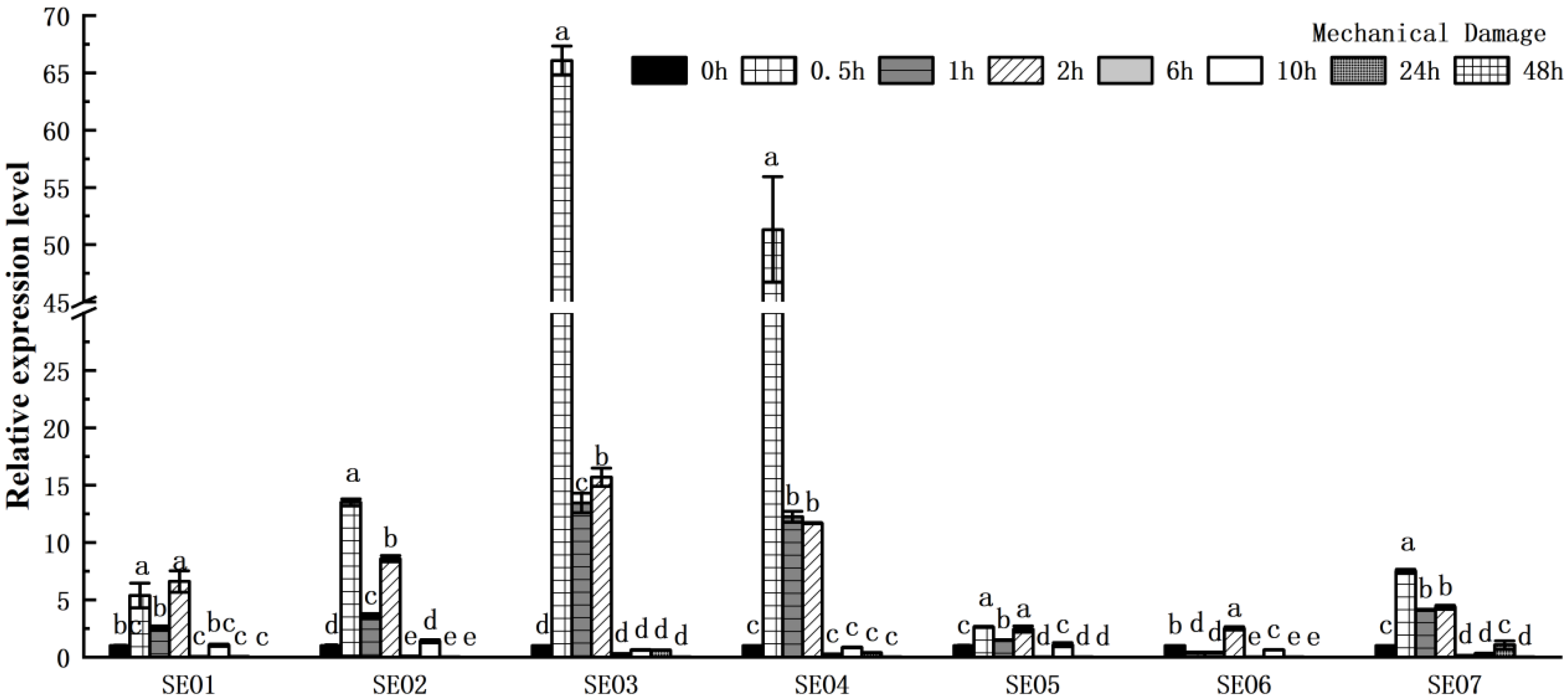
3.5.3. The Responses of HbSEOs to Mechanical Injury
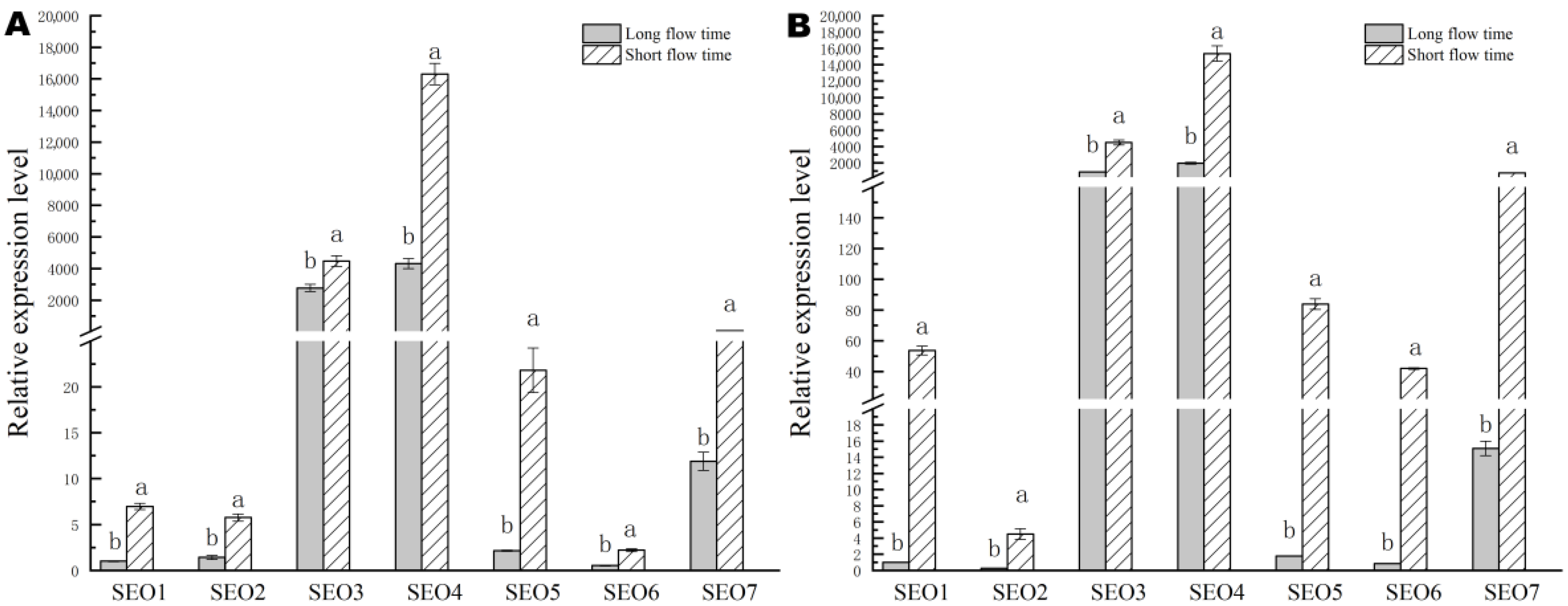
3.5.4. Variation of HbSEO Genes Expressions in the Rubber Trees with Different Latex Flow Durations
4. Discussion
4.1. Genome-Wide Identification of HbSEO Gene Family and Their Classification
4.2. Association of HbSEOs with Laticifer Plugging
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hao, B.; Wu, J. Biology of laticifer in Hevea and latex. Chin. J. Trop. Crops 2004, 4, 1–7. [Google Scholar]
- Wei, O.C.; Razak, S. Rubber tree cultivation and improvement in malaysia: Anatomical and morphological studies on Hevea brasiliensis and Hevea camargoana. J. Agric. Crops 2021, 7, 27–32. [Google Scholar]
- Kochian, L.V.; Pineros, M.A.; Liu, J.; Magalhaes, J.V. Plant adaptation to acid soils: The molecular basis for crop aluminum resistance. Annu. Rev. Plant Biol. 2015, 66, 571–598. [Google Scholar] [CrossRef]
- Diezel, C.; Allmann, S.; Baldwin, I.T. Mechanisms of optimal defense patterns in Nicotiana attenuata: Flowering attenuates herbivory-elicited ethylene and jasmonate signaling. J. Integr. Plant Biol. 2011, 12, 971–983. [Google Scholar] [CrossRef]
- Wu, J.L.; Hao, B.Z. Ultrastructure of P-protein in Hevea brasiliensis during sieve-tube development and after wounding. Protoplasma 1990, 3, 186–192. [Google Scholar] [CrossRef]
- D’Auzac, J.; Jacob, J.L.; Prévôt, J.C. The regulation of cis-polyisoprene production (natural rubber) from Hevea brasiliensis. Recent Res. Dev. Plant Physiol. 1997, 1, 273–332. [Google Scholar]
- Hao, B.; Wu, J.; Meng, C. Laticifer wound plugging in Hevea brasiliensis: The role of a protein-network with rubber particle aggregations in stopping latex flow and protecting wounded laticifers. J. Rubber Res. 2004, 7, 281–299. [Google Scholar]
- Ruping, B.; Ernst, A.M.; Jekat, S.B. Molecular and phylogenetic characterization of the sieve element occlusion gene family in Fabaceae and non-Fabaceae plants. BMC Plant Biol. 2010, 10, 219. [Google Scholar] [CrossRef] [Green Version]
- Knoblauch, M.; Peters, W.S. Forisomes, a novel type of Ca2+-dependent contractile protein motor. Cell Motil. Cytoskelet. 2004, 58, 137–142. [Google Scholar] [CrossRef]
- Will, T.; Van Bel, A.J.E. Physical and chemical interactions between aphids and plants. J. Exp. Bot. 2006, 57, 729–737. [Google Scholar] [CrossRef] [Green Version]
- Peters, W.S.; Knoblauch, M.; Warmann, S.A.; Schnetter, R.; Shen, A.Q.; Pickard, W.F. Tailed forisomes of canavalia gladiata: A new model to study Ca2+-driven protein contractility. Ann. Bot. 2007, 100, 101–109. [Google Scholar] [CrossRef] [Green Version]
- Peters, W.S.; Knoblauch, M.; Warmann, S.A. Anisotropic contraction in forisomes: Simple models won’t fit. Cell Motil. Cytoskelet. 2010, 65, 368–378. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y. TBtools: An integrative toolkit developed for Interactive analyses of big biological data. Mol. Plant 2020, 13, 1194–1210. [Google Scholar] [CrossRef]
- Hua, Y.W.; Huang, T.D.; Huang, H.S. Micropropagation of self-rooting juvenile clones by secondary somatic embryogenesis in Hevea brasiliensis. Plant Breed. 2010, 129, 202–207. [Google Scholar] [CrossRef]
- Tang, C.; Huang, D.; Yang, J. The sucrose transporter HbSUT3 plays an active role in sucrose loading to laticifer and rubber productivity in exploited trees of Hevea brasiliensis (para rubber tree). Plant Cell Environ. 2010, 33, 1708–1720. [Google Scholar] [CrossRef]
- Noll, G.A.; Rüping, B.; Ernst, A.M. The promoters of forisome genes MtSEO2 and MtSEO3 direct gene expression to immature sieve elements in Medicago truncatula and Nicotiana tabacum. Plant Mol. Biol. Rep. 2009, 4, 526–533. [Google Scholar] [CrossRef]
- Ernst, A.M.; Rüping, B.; Jekat, S.B. The sieve element occlusion gene family in dicotyledonous plants. Plant Signal. Behav. 2011, 6, 151–153. [Google Scholar] [CrossRef] [Green Version]
- Pélissier, H.C.; Peters, W.S.; Ray, C. GFP tagging of sieve element occlusion (SEO) proteins results in green fluorescent forisomes. Plant Cell Physiol. 2008, 49, 1699–1710. [Google Scholar] [CrossRef]
- Bowers, J.E.; Chapman, B.A.; Rong, J. Unravelling angiosperm genome evolution by phylogenetic analysis of chromosomal duplication events. Nature 2003, 422, 433–438. [Google Scholar] [CrossRef]
- Blanc, G. Functional divergence of duplicated genes formed by polyploidy during Arabidopsis evolution. Plant Cell 2004, 7, 1679–1691. [Google Scholar] [CrossRef] [Green Version]
- Tuskan, G.; Difazio, S.; Jansson, S. The genome of black cottonwood, populus trichocarpa (Torr. & Gray). Science 2006, 313, 1596–1604. [Google Scholar] [CrossRef] [Green Version]
- Johnson, A.R.; Moghe, G.D.; Frank, M.H. Growing a glue factory: Open questions in laticifer development. Curr. Opin. Plant Biol. 2021, 64, 102096. [Google Scholar] [CrossRef]
- Ehlers, K.; Knoblauch, M.; van Bel, A.J.E. Ultrastructural features of well-preserved and injured sieve elements: Minute clamps keep the phloem transport conduits free for mass flow. Protoplasma 2000, 214, 80–92. [Google Scholar] [CrossRef]
- Michael, K.; van Bel Aart, J.E. Sieve tubes in action. Plant Cell 1998, 10, 35–50. [Google Scholar] [CrossRef] [Green Version]
- Ernst, A.M.; Jekat, S.B.; Zielonka, S. PNAS Plus: Sieve element occlusion (SEO) genes encode structural phloem proteins involved in wound sealing of the phloem. Proc. Natl. Acad. Sci. USA 2012, 109, E1980–E1990. [Google Scholar] [CrossRef] [Green Version]
- Tungngoen, K.; Viboonjun, U.; Kongsawadworakul, P. Hormonal treatment of the bark of rubber trees (Hevea brasiliensis) increases latex yield through latex dilution in relation with the differential expression of two aquaporin genes. J. Plant Physiol. 2010, 168, 253–262. [Google Scholar] [CrossRef]
- Wilson, R.N.; Somerville, H. Gibberellin Is Required for Flowering in Arabidopsis thaliana under Short Days. Plant Physiol. 1992, 100, 403–408. [Google Scholar] [CrossRef] [Green Version]
- Hao, B. The effect of stimulating tapping with etherl on the structure of the barj of Hevea brasilensis Muell. Agric. Chin. J. Trop. Crops 1980, 1, 61–65. [Google Scholar]
- Haiyan, Z.; An, F. Progress in study on the mechanisms to increase latex yield of Hevea brasiliensis by ethephon stimula. Sci. Silvae Sin. 2010, 46, 120–125. [Google Scholar] [CrossRef]
- Yamaguchi, T.; Kurihara, Y.; Makita, Y. Regulatory potential of bHLH-type transcription factors on the road to rubber biosynthesis in Hevea brasiliensis. Plants 2020, 9, 674. [Google Scholar] [CrossRef]
- D’Auzac, J. Factors involved in the stopping of the latex flow after tapping. In Physiol of the Rubber Tree Latex; CRC Press: Boca Raton, FL, USA, 1989. [Google Scholar]
- Yeang, H.Y. The kinetics of latex flow from the rubber tree in relation to latex vessel plugging and turgor pressure. J. Rubber Res. 2005, 8, 160–181. [Google Scholar]
- Shi, M. Effect on the induction of the secondary laticifer differentiation by the transportation of exogenous JA in Hevea brasiliensis. Chin. J. Trop. Crops 2012, 33, 1647–1653. [Google Scholar] [CrossRef]
- Hao, B.; Wu, J. Laticifer differentiation in Hevea brasiliensis: Induction by exogenous jasmonic acid and linolenic acid. Ann. Bot. 2000, 85, 37–43. [Google Scholar] [CrossRef] [Green Version]
- An, F.; Rookes, J.; Xie, G. Ethephon increases rubber tree latex yield by regulating aquaporins and alleviating the tapping-induced local increase in latex total solid content. J. Plant Growth Regul. 2016, 35, 701–709. [Google Scholar] [CrossRef]
- An, F.; Cai, X.; Rookes, J. Latex dilution reaction during the tapping flow course of Hevea brasiliensis and the effect of ethrel stimulation. Braz. J. Bot. 2015, 38, 211–221. [Google Scholar] [CrossRef]
- Priyadarshan, P.M. Biology of Hevea Rubber; Commonwealth Agricultural Bureau International: Wallingford, UK, 2011. [Google Scholar]
- Hao, B.; Wu, J.; Tan, H. Laticifer plugging of Hevea brasiliensis after severing. Chin. J. Trop. Crops 1996, 17, 1–6. [Google Scholar]
- An, F.; Cai, X.; Rookes, J. Regulation of HbPIP2;3, a latex-abundant water transporter, is associated with latex dilution and yield in the rubber tree (Hevea brasiliensis Muell. Arg.). PLoS ONE 2015, 10, e0125595. [Google Scholar] [CrossRef] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Accession Number (NCBI) | Rename | Forward Primer | Reverse Primer |
|---|---|---|---|
| XM_021833957.1 | HbSEO1 | AGGCTTGGGAGTTGTCGAGC | GCAGGACGTCAATGTTAACCCTC |
| XM_021811739.1 | HbSEO2 | GGCTTCCTGTTGTGGACCGAT | GGATTCACTACTTTCCCTTGCGG |
| XM_021804083.1 | HbSEO3 | GTGAGGCTGGAGAGCATGTG | TTCCATGACTGAAGGGCATC |
| XM_021804082.1 | HbSEO4 | ACGCGTTGTGTTTTGTTCATCAG | TGTGCATGGGAAGCTCCTTGT |
| XM_021811738.1 | HbSEO5 | GGAAGAAAGGCAACGGAATGCT | ACTCTCCATGGGACGCTTACAC |
| XM_021813449.1 | HbSEO6 | TAATCCAGGGATTGGCGCTG | GAAGGGTGGCACGATTCAGG |
| XM_021828364.1 | HbSEO7 | ACCCGTCAAGTTGAGACCAC | AGCACCAACTTAGCATCCCAGGAG |
| AB268099 | 18S rRNA | GCTCGAAGACGATCAGATACC | TTCAGCCTTGCGACCATAC |
| Gene ID | Gene Name | Genomics (bp) | CDS (bp) | Amino Acids | MW (kDa) | PI | Instability Index | α-Helix | β-Turn | Random Coil | Sub-Cellular Location |
|---|---|---|---|---|---|---|---|---|---|---|---|
| XP_021689649.1 | HbSEO1 | 2518 | 2136 | 711 | 80.94 | 6.12 | 39.85 | 54.29% | 4.50% | 30.38% | cytoplasm |
| XP_021667431.1 | HbSEO2 | 2463 | 2139 | 712 | 80.95 | 6.12 | 44.81 | 54.07% | 4.21% | 31.46% | mitochondria |
| XP_021659775.1 | HbSEO3 | 2476 | 2157 | 718 | 82.14 | 6.14 | 40.56 | 53.34% | 3.06% | 32.59% | cytoplasm |
| XP_021659774.1 | HbSEO4 | 2758 | 2157 | 718 | 81.98 | 6.13 | 37.54 | 56.69% | 4.04% | 29.81% | mitochondria |
| XP_021667430.1 | HbSEO5 | 2485 | 2193 | 730 | 83.26 | 6.29 | 47.99 | 53.97% | 4.11% | 31.78% | cytoplasm |
| XP_021669141.1 | HbSEO6 | 2448 | 2151 | 716 | 82.06 | 6.29 | 39.8 | 54.47% | 4.33% | 30.59% | microbody |
| XP_021684056.1 | HbSEO7 | 2419 | 2157 | 718 | 82.36 | 6.27 | 35.64 | 55.01% | 4.32% | 30.92% | microbody |
| Long Latex Flow Durations | Short Latex Flow Durations | ||||||
|---|---|---|---|---|---|---|---|
| Tree Number | Girth (cm) | Average Latex Flow Duration | Average Yield (mL∙tree−1∙tapping−1) | Tree Number | Girth (cm) | Average Latex Flow Duration | Average Yield (mL∙tree−1∙tapping−1) |
| 250 | 65 | 6 h 16 min | 394 | 59 | 64 | 1 h 57 min | 67 |
| 287 | 65 | 4 h 49 min | 212 | 293 | 64 | 1 h 48 min | 11 |
| 317 | 65 | 4 h 04 min | 192 | 301 | 67 | 2 h 07 min | 51 |
| 33 | 65 | 4 h 31 min | 342 | 271 | 63 | 1 h 48 min | 27 |
| 77 | 66 | 4 h 05 min | 206 | 333 | 65 | 1 h 54 min | 14 |
| Mean | 65.2 A | 4 h 45 min A | 269.2 A | 64.6 A | 1 h 55 min B | 34 B | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, X.; Cheng, L.; Peng, W.; Xie, G.; Liu, Z.; An, F. Identification of Sieve Element Occlusion Gene (SEOs) Family in Rubber Trees (Hevea brasiliensis Muell. Arg.) Provides Insights to the Mechanism of Laticifer Plugging. Forests 2022, 13, 433. https://0-doi-org.brum.beds.ac.uk/10.3390/f13030433
Wang X, Cheng L, Peng W, Xie G, Liu Z, An F. Identification of Sieve Element Occlusion Gene (SEOs) Family in Rubber Trees (Hevea brasiliensis Muell. Arg.) Provides Insights to the Mechanism of Laticifer Plugging. Forests. 2022; 13(3):433. https://0-doi-org.brum.beds.ac.uk/10.3390/f13030433
Chicago/Turabian StyleWang, Xuan, Linlin Cheng, Wentao Peng, Guishui Xie, Zifan Liu, and Feng An. 2022. "Identification of Sieve Element Occlusion Gene (SEOs) Family in Rubber Trees (Hevea brasiliensis Muell. Arg.) Provides Insights to the Mechanism of Laticifer Plugging" Forests 13, no. 3: 433. https://0-doi-org.brum.beds.ac.uk/10.3390/f13030433