
Effect of Two Exogenous Organic Acids on the Excitation Effect of Soil Organic Carbon in Beijing, China


Abstract
:1. Introduction
2. Materials and Methods
2.1. Soil Sample Collection and Processing
2.2. Indoor Analysis
2.2.1. Soil CO2 Cumulative Release Amount Determination Method
2.2.2. Soil Microbial Diversity Determination Method
2.3. Fitting the Equations for the Soil Organic Carbon Mineralization Process
2.4. Data Analysis
3. Results
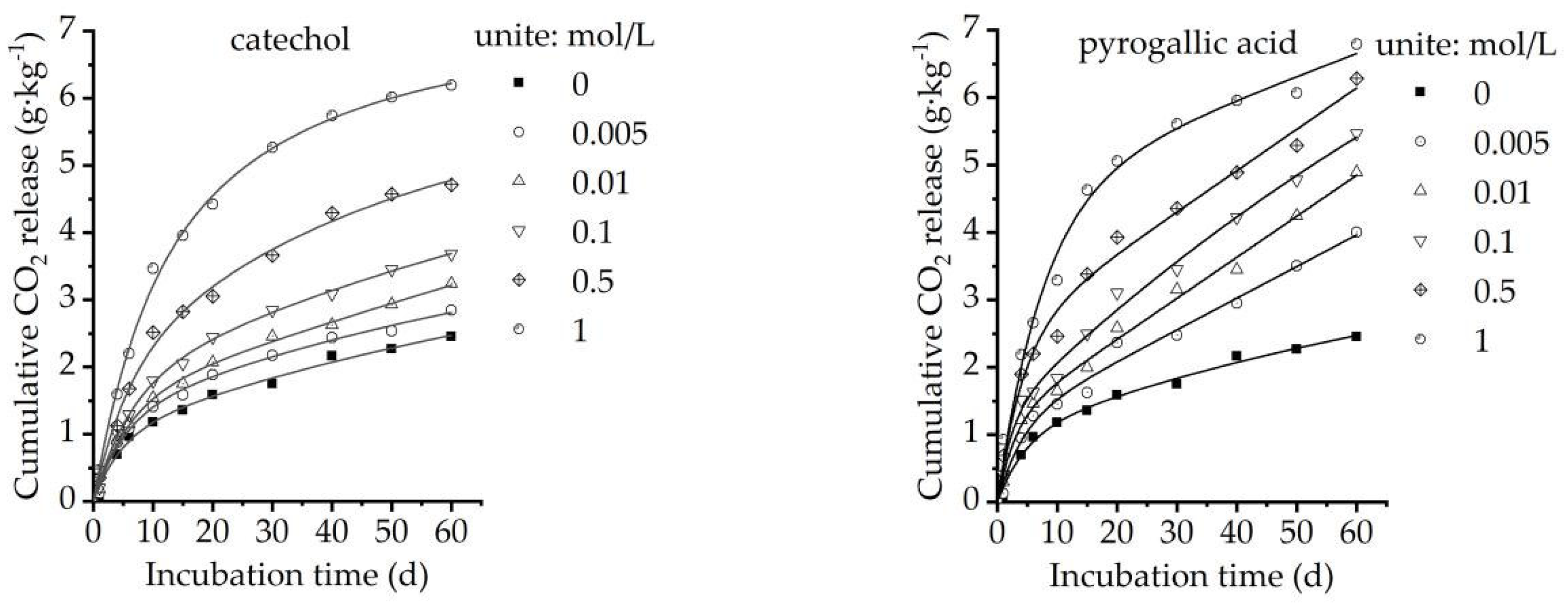
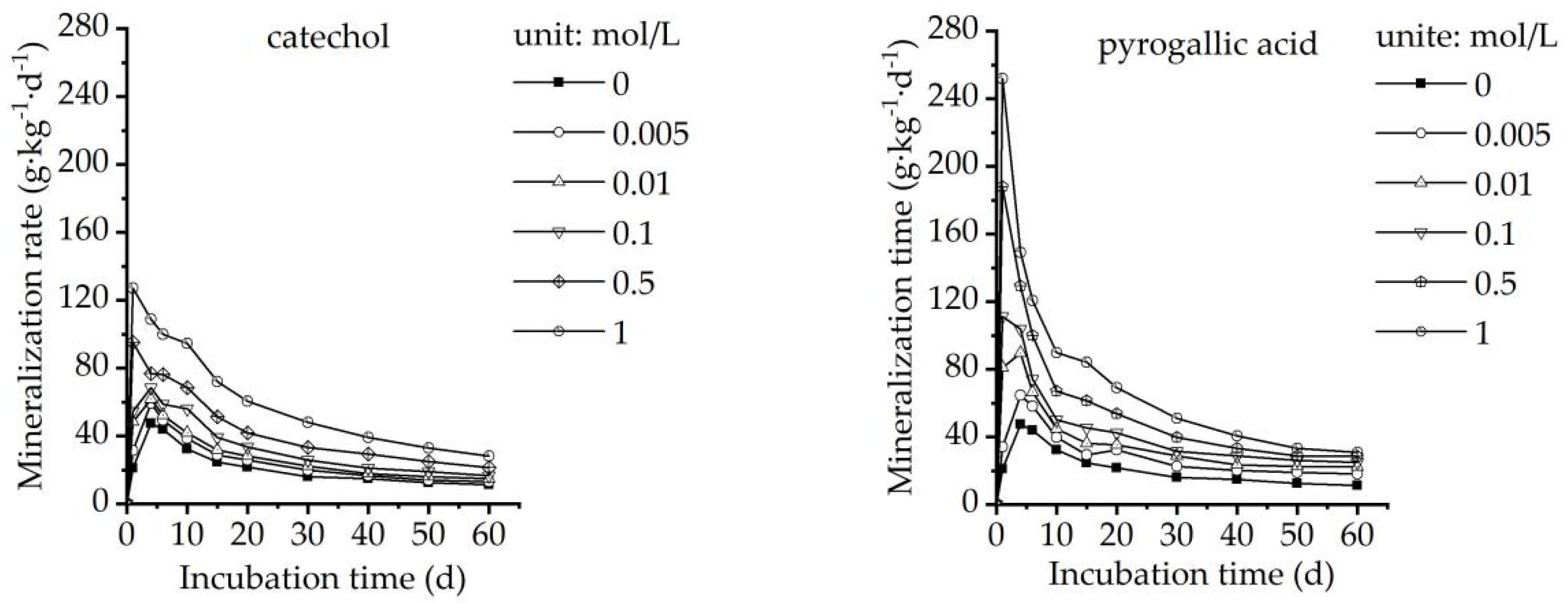
3.1. Effect of Organic Acids on Soil Organic Carbon Mineralization
3.2. Effect of Organic Acids on the Active Carbon Content of the Soil
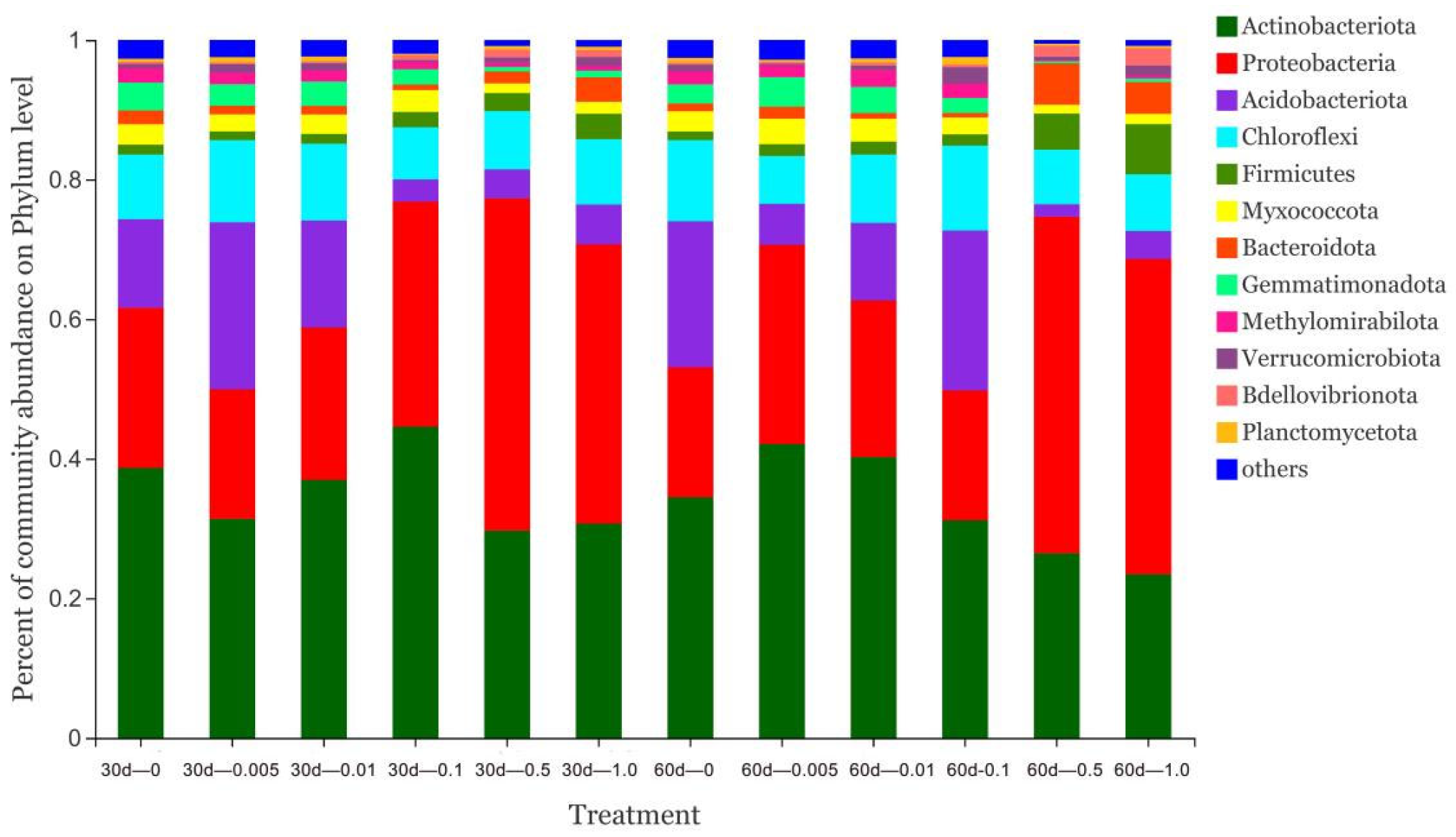
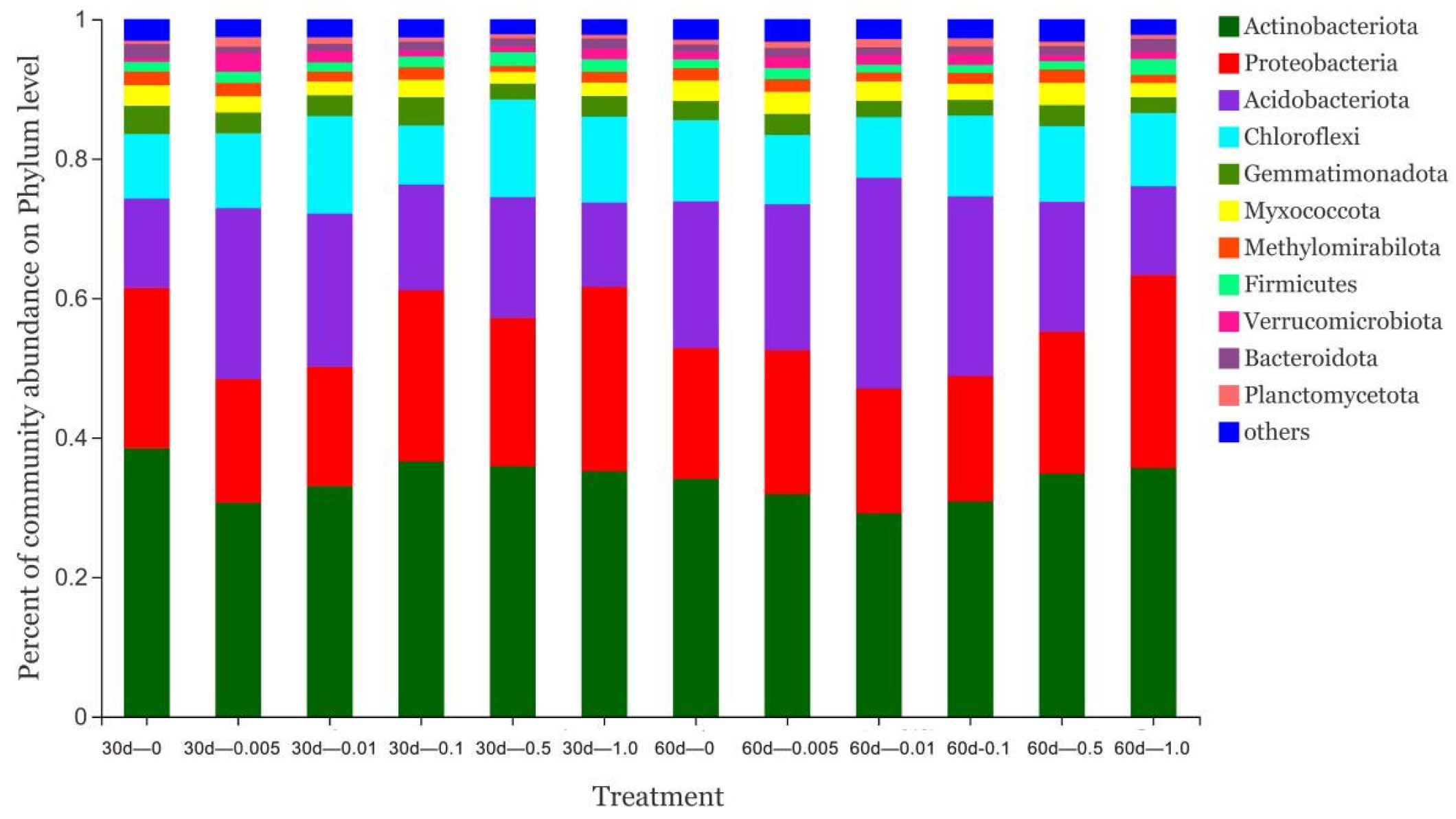
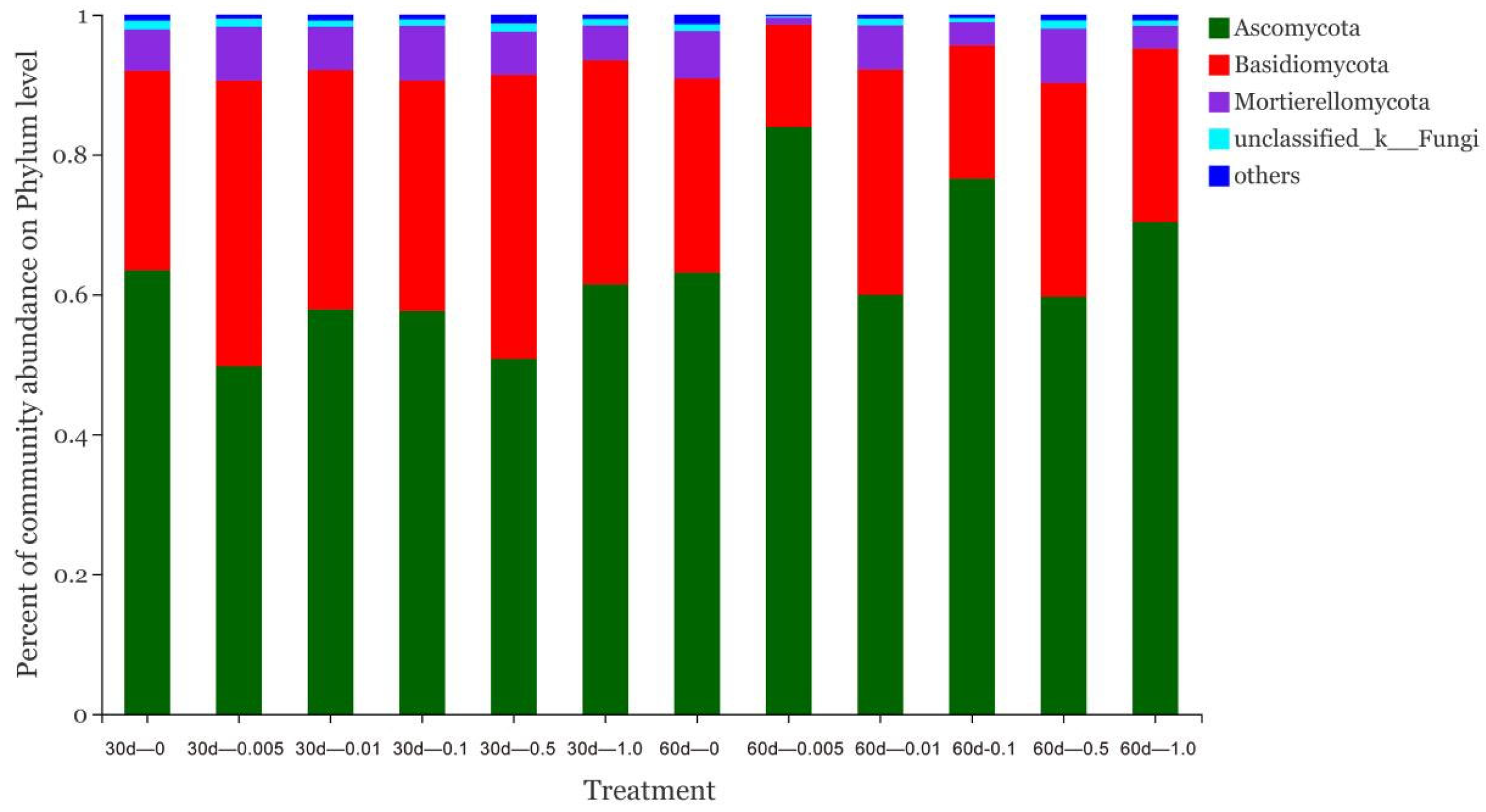
3.3. Effect of Catechol on Soil Microorganisms
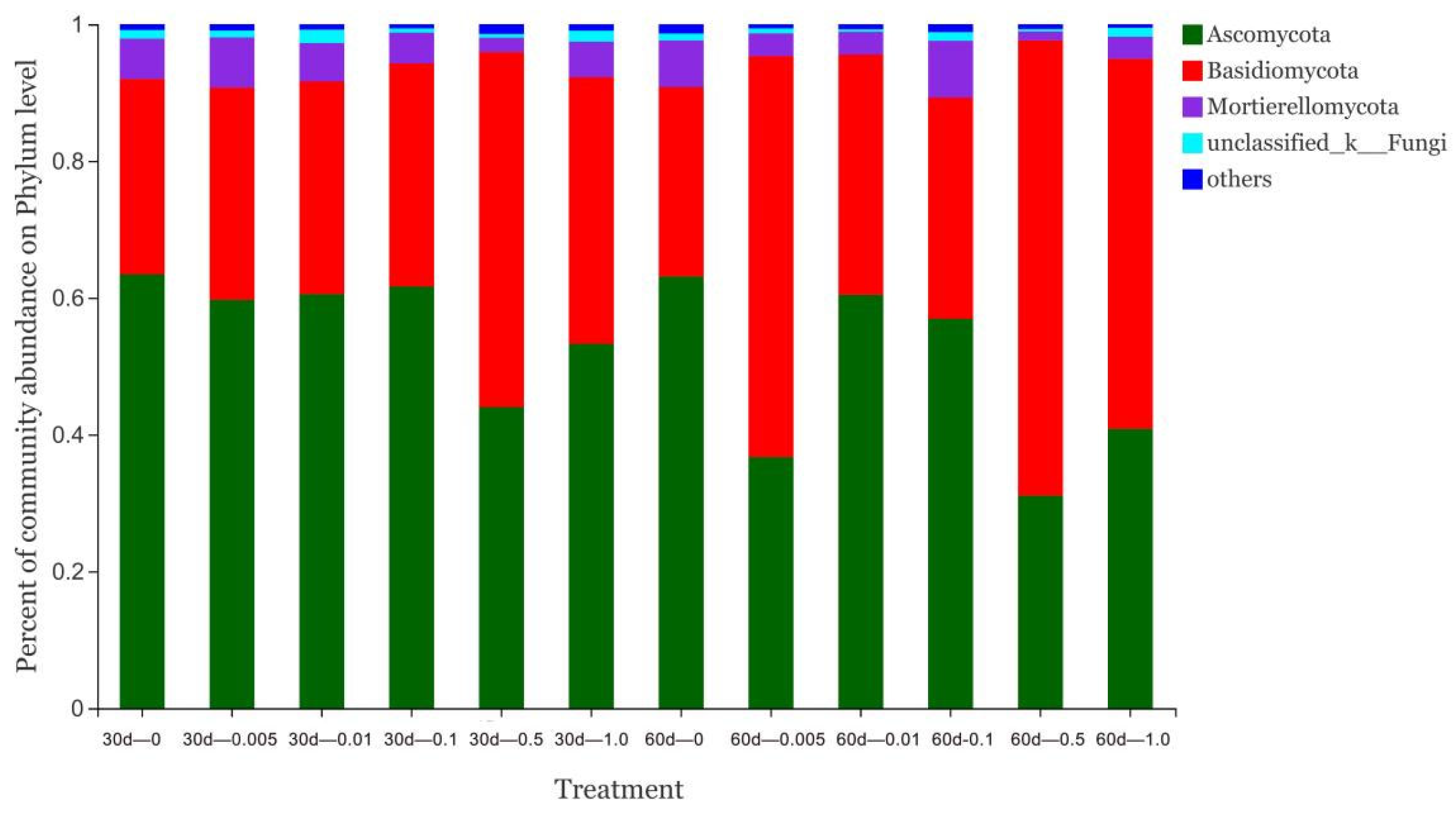
3.4. Effect of Pyrogallic Acid on Soil Microorganisms
3.5. Correlation Analysis between Cumulative CO2 Release and Microbial Diversity
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lal, R. Sequestering carbon in soils of agro-ecosystems—ScienceDirect. Food Policy 2011, 36, S33–S39. [Google Scholar] [CrossRef]
- Zhang, Z.; Cai, X.Z.; Tang, C.D.; Guo, J.F. Priming effect of dissolved organic matter in the surface soil of a Cunninghamia lanceolata plantation. Acta Ecol. Sin. 2017, 37, 7660–7667. [Google Scholar]
- Arunrat, N.; Kongsurakan, P.; Sereenonchai, S.; Hatano, R. Soil Organic Carbon in Sandy Paddy Fields of Northeast Thailand: A Review. Agronomy 2020, 10, 1061. [Google Scholar] [CrossRef]
- Arunrat, N.; Pumijumnong, N.; Sereenonchai, S.; Hatano, R. Factors Controlling Soil Organic Carbon Sequestration of Highland Agricultural Areas in the Mae Chaem Basin, Northern Thailand. Agronomy 2020, 10, 305. [Google Scholar] [CrossRef] [Green Version]
- Keiluweit, M.; Bougoure, J.J.; Nico, P.S.; Pett-Ridge, J.; Kleber, M. Mineral protection of soil carbon counteracted by root exudates. Nat. Clim. Chang. 2015, 5, 588–595. [Google Scholar] [CrossRef]
- Hamer, U.; Marschner, B. Priming effects of sugars, amino acids, organic acids and catechol on the mineralization of lignin and peat. J. Plant Nutr. Soil Sci. 2002, 165, 261–268. [Google Scholar] [CrossRef]
- Hamer, U.; Marschner, B. Priming effects in different soil types induced by fructose, alanine, oxalic acid and catechol additions. Soil Biol. Biochem. 2005, 37, 445–454. [Google Scholar] [CrossRef]
- Mallik, A.U. Effect of phenolic compounds on selected soil properties. For. Ecol. Manag. 1997, 92, 11–18. [Google Scholar]
- Qu, X.H.; Wang, J.G. Effect of amendments with different phenolic acids on soil microbial biomass, activity, and community diversity. Appl. Soil Ecol. 2008, 39, 172–179. [Google Scholar] [CrossRef]
- Ma, Y.H.; Wang, X.F.; Wei, M. Accumulation of phenolic acids in continuously cropped cucumber soil and their effects on soil microbes and enzyme activities. Chin. J. Appl. Ecol. 2005, 16, 2149–2153. [Google Scholar]
- Hu, K.; Tao, J.P.; Huang, K.; Hu, J.; Wang, W. Effects of simulated root exudate carbon inputs on the dynamics of microbial communities during litter decomposition. Chin. J. Appl. Environ. Biol. 2020, 26, 417–424. [Google Scholar]
- Bodelier, P.L.E.; Wijlhuizen, A.G.; Blom, C.W.; Laanbroek, H.J. Effects of photoperiod on growth of and denitrification by Pseudomonas chlororaphis in the root zone of Glyceria maxima, studied in a gnotobiotic microcosm. Plant Soil 1997, 190, 91–103. [Google Scholar] [CrossRef]
- Rouatt, J.W.; Katznelson, H.; Payne, T.M.B. Statistical Evaluation of the Rhizosphere Effect. Soil Sci. Soc. 1960, 24, 271–273. [Google Scholar] [CrossRef]
- Song, J.F.; Cui, X.Y. Advanc in Study of Low Molecular Mass Organic Acids in Forest Soils. Sci. Silvae Sin. 2008, 6, 121–127. [Google Scholar]
- Sun, Y.; Xu, X.L.; Yakov, K. Mechanisms of rhizosphere priming effects and their ecological significance. Chin. J. Plant Ecol. 2014, 38, 62–75. [Google Scholar]
- Bengtson, P.; Barker, J.; Grayston, S.J. Evidence of a strong coupling between root exudation, C and N availability, and stimulated SOM decomposition caused by rhizosphere priming effects. Ecol. Evol. 2012, 2, 1843–1852. [Google Scholar] [CrossRef]
- Graaff, M.A.D.; Classen, A.T.; Castro, H.F.; Schadt, C.W. Labile soil carbon inputs mediate the soil microbial community composition and plant residue decomposition rates. New Phytol. 2010, 188, 1055–1064. [Google Scholar] [CrossRef] [Green Version]
- Cheng, W.; Johnson, D.W.; Fu, S. Rhizosphere Effects on Decomposition. Soil Sci. Soc. Am. J. 2003, 67, 1418. [Google Scholar] [CrossRef]
- Mondini, C.; Sinicco, T.; Cayuela, M.L.; Sanchez, M.M.A. A simple automated system for measuring soil respiration by gas chromatography. Talanta 2010, 81, 849–855. [Google Scholar] [CrossRef]
- Updegraff, K.; Pastor, J.; Bridgham, S.D.; Johnston, C.A. Environmental and substrate controls over carbon and nitrogen mineralization in northern wetlands. Ecol. Appl. 1995, 5, 151–163. [Google Scholar] [CrossRef]
- Wedin, D.A.; Pastor, J. Nitrogen mineralization dynamics in grass monocultures. Oecologia 1993, 96, 186–192. [Google Scholar] [CrossRef] [PubMed]
- Murwira, H.K.; Kirchmann, H.; Swift, M.J. The effect of moisture on the decomposition rate of cattle manure. Plant Soil 1990, 122, 197–199. [Google Scholar] [CrossRef]
- Cordovil, C.M.D.S.; Coutinho, J.; Goss, M.; Cabral, F. Potentially mineralisable nitrogen from organic materials applied to a sandy soil: Fitting the one-pool exponential model. Soil Use Manag. 2010, 21, 65–72. [Google Scholar] [CrossRef]
- Ribeiro, H.M.; Fangueiro, D.; Alves, F.; Vasconcelos, E.; Coutinho, J.; Bol, R.; Cabral, F. Carbon-mineralization kinetics in an organically managed Cambic Arenosol amended with organic fertilizers. J. Plant Nutr. Soil Sci. 2010, 173, 39–45. [Google Scholar] [CrossRef]
- Kuzyakov, Y.; Bol, R. Sources and mechanisms of priming effect induced in two grassland soils amended with slurry and sugar. Soil Biol. Biochem. 2006, 38, 747–758. [Google Scholar] [CrossRef]
- Fontaine, S.; Bardoux, G.; Abbadie, L.; Mariotti, A. Carbon input to soil may decrease soil carbon content. Ecol. Lett. 2004, 7, 314–320. [Google Scholar] [CrossRef]
- Nottingham, A.T.; Griffiths, H.; Chamberlain, P.M.; Stott, A.W.; Tanner, E.V.J. Soil priming by sugar and leaf-litter substrates: A link to microbial groups. Appl. Soil Ecol. 2009, 42, 183–190. [Google Scholar] [CrossRef]
- Collins, H.P.; Rasmussen, P.E.; Douglas, C.L. Crop rotation and residue management effects on soil carbon and microbialdynamics. Soil Sci. Soc. Am. J. 1992, 56, 783–788. [Google Scholar] [CrossRef]
- Duong, T.T.T.; Baumann, K.; Marschner, P. Frequent addition of wheat straw residues to soil enhances carbon mineralization rate. Soil Biol. Biochem. 2009, 41, 1475–1482. [Google Scholar] [CrossRef]
- Zhang, P.; Li, H.; Jia, Z.K.; Lu, W.T.; Zhang, H.; Yang, B.P. Effects of Straw Returning on Soil Organic Carbon and Carbon Mineralization in Semi-arid Areas of Southern Ningxia, China. J. Agro-Environ. Sci. 2011, 30, 2518–2525. [Google Scholar]
- Lu, W.T.; Jia, Z.K.; Zhang, P.; Wang, W.; Hou, X.Q.; Yang, B.P. Effects of Straw Returning on Soil Labile Organic Carbon and Enzyme Activity in Semi-arid Areas of Southern Ningxia, China. J. Agro-Environ. Sci. 2011, 30, 522–528. [Google Scholar]
- Qiao, N.; Schaefer, D.; Blagodatskaya, E.; Zou, X.M.; Xu, X.L.; Kuzyakov, Y. Labile carbon retention compensates for CO2 released by priming in forest soils. Glob. Chang. Biol. 2014, 20, 1943–1954. [Google Scholar] [CrossRef]
- Yan, D.Z.; Wang, D.J.; Yang, L.Z. Long-term effect of chemical fertilizer, straw, and manure on labile organic matter fractions in a paddy soil. Biol. Fertil. Soils 2007, 44, 93–101. [Google Scholar] [CrossRef]
- Nadelhoffer, K.J.; Boone, R.D.; Bowden, R.D.; Canary, J.D.; Kaye, J.; Micks, P.; Ricca, A.; Aitkenhead, J.A.; Lajtha, K.; Mcdowell, W.H. The DIRT Experiment: Litter and Root influences on Forest Soil Organic Matter Stocks and Function. In Forests in Time: The Environmental Consequences of 1000 Years of Change in New England; Foster, D.R., Aber, J.D., Eds.; Yale University Press: New Haven, CT, USA, 2004; pp. 300–315. [Google Scholar]
- Baath, E.; Anderson, T.H. Comparison of soil fungal /bacterial ratios in a pH gradient using physiological and PLFA-based techniques. Soil Biol. Biochem. 2003, 35, 955–963. [Google Scholar] [CrossRef]
- Deng, H.; Li, X.F.; Cheng, W.D.; Zhu, Y.G. Resistance and resilience of Cu-polluted soil after Cu perturbation, tested by a wide range of soil microbial parameters. FEMS Microbiol. Ecol. 2009, 70, 293–304. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Yang, J.P.; Yang, S.H.; Yang, Z.C.; Lv, Y.M. Effect of a 10 °C-elevated temperature under different water contents on the microbial community in a tea orchard soil. Eur. J. Soil Biol. 2014, 62, 113–120. [Google Scholar] [CrossRef]
- Zheng, Y.X.; Cao, J.L.; Yang, Z.J.; Lin, Y.F.; Huang, M.Q.; Yang, Y.S. Seasonal Variation of Soil Microbial Community Structure in Castanopsis Carlesii Natural Forest and Orange Orchard. Ecol. Environ. Sci. 2019, 28, 1991–1998. [Google Scholar]
- Fierer, N.; Lauber, C.L.; Ramirez, K.S.; Zaneveld, J.; Bradford, M.A.; Knight, R. Comparative metagenomic, phylogenetic and physiological analyses of soil microbial communities across nitrogen gradients. ISME J. 2012, 6, 1007–1017. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.J.; Sui, Y.Y.; Yu, Z.H.; Shi, Y.; Chu, H.Y.; Jin, J.; Liu, X.B.; Wang, G.H. High throughput sequencing analysis of biogeographical distribution of bacterial communities in the black soils of northeast China. Soil Biol. Biochem. 2014, 70, 113–122. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Catechol | Pyrogallic Acid | |||||||
|---|---|---|---|---|---|---|---|---|
| Concentrations (mol/L) | Equations | C0 (g/kg) | k0 | R2 | Equations | C0 (g/kg) | k0 | R2 |
| 0 | Y = 2.35 (1 − e−0.063x) | 2.35 | 0.063 | 0.969 | Y = 2.35 (1 − e−0.063x) | 2.35 | 0.063 | 0.969 |
| 0.005 | Y = 2.66 (1 − e−0.069x) | 2.66 | 0.069 | 0.975 | Y = 3.62 (1 − e−0.039x) | 3.62 | 0.039 | 0.984 |
| 0.01 | Y = 3.05 (1 − e−0.063x) | 3.05 | 0.063 | 0.977 | Y = 4.47 (1 − e−0.041x) | 4.47 | 0.041 | 0.983 |
| 0.1 | Y = 3.58 (1 − e−0.063x) | 3.58 | 0.063 | 0.982 | Y = 4.99 (1 − e−0.050x) | 4.99 | 0.050 | 0.973 |
| 0.5 | Y = 4.65 (1 − e−0.064x) | 4.65 | 0.064 | 0.985 | Y = 5.89 (1 − e−0.062x) | 5.89 | 0.062 | 0.977 |
| 1.0 | Y = 6.12 (1 − e−0.072x) | 6.12 | 0.072 | 0.995 | Y = 6.43 (1 − e−0.082x) | 6.43 | 0.082 | 0.985 |
| Concentrations (mol/L) | Ace | Chao1 | Shannon | ||||||
|---|---|---|---|---|---|---|---|---|---|
| 30 d | 60 d | Differences | 30 d | 60 d | Differences | 30 d | 60 d | Differences | |
| 0 | 3014.88 | 2980.23 | −34.65 | 2966.15 | 2950.20 | −15.96 | 6.39 | 6.38 | −0.02 |
| 0.005 | 3018.22 | 2873.40 | −144.82 | 3009.06 | 2839.56 | −169.50 | 6.34 | 6.18 | −0.16 |
| 0.01 | 2936.62 | 2751.88 | −184.74 | 2931.71 | 2754.11 | −177.60 | 6.31 | 6.24 | −0.07 |
| 0.1 | 2819.87 | 2671.37 | −148.50 | 2884.99 | 2636.00 | −158.99 | 5.99 | 5.66 | −0.13 |
| 0.5 | 2264.41 | 2039.64 | −224.78 | 2657.18 | 2092.78 | −564.40 | 5.32 | 5.20 | −0.11 |
| 1 | 2476.23 | 2209.31 | −266.92 | 2610.06 | 2200.85 | −409.21 | 5.70 | 5.56 | −0.15 |
| Concentrations (mol/L) | Ace | Chao1 | Shannon | ||||||
|---|---|---|---|---|---|---|---|---|---|
| 30 d | 60 d | Differences | 30 d | 60 d | Differences | 30 d | 60 d | Differences | |
| 0 | 537.61 | 508.54 | −29.07 | 526.86 | 505.93 | −20.93 | 3.28 | 3.17 | −0.11 |
| 0.005 | 562.19 | 498.95 | −63.24 | 542.97 | 490.12 | −52.85 | 3.14 | 2.42 | −0.72 |
| 0.01 | 570.56 | 539.98 | −30.58 | 565.63 | 485.41 | −80.22 | 3.30 | 2.98 | −0.32 |
| 0.1 | 527.94 | 493.98 | −33.96 | 522.01 | 471.69 | −50.32 | 2.86 | 2.61 | −0.25 |
| 0.5 | 436.42 | 264.28 | −172.14 | 441.62 | 242.29 | −199.34 | 2.82 | 1.97 | −0.85 |
| 1 | 401.75 | 243.61 | −158.14 | 498.88 | 305.04 | −193.83 | 2.77 | 2.53 | −0.24 |
| Concentrations (mol/L) | Ace | Chao1 | Shannon | ||||||
|---|---|---|---|---|---|---|---|---|---|
| 30 d | 60 d | Differences | 30 d | 60 d | Differences | 30 d | 60 d | Differences | |
| 0 | 3174.11 | 3173.50 | −0.61 | 3074.97 | 3125.72 | 50.74 | 6.442 | 6.426 | −0.016 |
| 0.005 | 562.19 | 498.95 | −63.24 | 542.97 | 490.12 | −52.85 | 3.14 | 2.42 | −0.72 |
| 0.01 | 3141.12 | 3416.40 | 275.27 | 3143.80 | 3462.27 | 318.47 | 6.311 | 6.459 | 0.148 |
| 0.1 | 3225.59 | 3105.74 | −119.85 | 3230.44 | 3082.39 | −148.05 | 6.361 | 6.365 | 0.005 |
| 0.5 | 2801.83 | 3331.09 | 529.26 | 2779.22 | 3331.06 | 551.84 | 6.101 | 6.476 | 0.375 |
| 1 | 3259.09 | 2963.23 | −295.86 | 3252.48 | 3045.26 | −207.23 | 6.276 | 6.169 | −0.106 |
| Concentrations (mol/L) | Ace | Chao1 | Shannon | ||||||
|---|---|---|---|---|---|---|---|---|---|
| 30 d | 60 d | Differences | 30 d | 60 d | Differences | 30 d | 60 d | Differences | |
| 0 | 537.71 | 504.10 | −33.61 | 531.35 | 512.40 | −18.95 | 3.287 | 3.181 | −0.106 |
| 0.005 | 493.03 | 471.99 | −21.04 | 479.00 | 501.11 | 22.11 | 2.971 | 1.344 | −1.627 |
| 0.01 | 534.50 | 524.47 | −10.03 | 522.92 | 556.33 | 33.41 | 3.038 | 3.043 | 0.006 |
| 0.1 | 511.95 | 536.28 | 24.32 | 514.74 | 492.69 | −22.04 | 2.999 | 2.672 | −0.327 |
| 0.5 | 416.10 | 477.52 | 61.42 | 453.25 | 506.71 | 53.46 | 2.814 | 3.237 | 0.423 |
| 1 | 449.74 | 478.46 | 28.72 | 430.74 | 475.06 | 44.32 | 2.965 | 2.980 | 0.015 |
| Bacteria | Ace Indices | Chao1 Indices | Shannon Indices |
| CO2 accumulative delivery amount (g/kg) | −0.857 ** | −0.880 ** | −0.772 ** |
| Fungi | Ace Indices | Chao1 Indices | Shannon Indices |
| CO2 accumulative delivery amount (g/kg) | −0.889 ** | −0.743 ** | −0.660 ** |
| Bacteria | Ace Indices | Chao1 Indices | Shannon Indices |
| CO2 accumulative delivery amount (g/kg) | −0.062 | 0.095 | −0.281 |
| Fungi | Ace Indices | Chao1 Indices | Shannon Indices |
| CO2 accumulative delivery amount (g/kg) | −0.370 | −0.362 | −0.073 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiao, Y.; Yu, Y.; Wang, Y.; Wang, X.; Wang, Y.; Dai, W.; Luan, Y. Effect of Two Exogenous Organic Acids on the Excitation Effect of Soil Organic Carbon in Beijing, China. Forests 2022, 13, 487. https://0-doi-org.brum.beds.ac.uk/10.3390/f13030487
Xiao Y, Yu Y, Wang Y, Wang X, Wang Y, Dai W, Luan Y. Effect of Two Exogenous Organic Acids on the Excitation Effect of Soil Organic Carbon in Beijing, China. Forests. 2022; 13(3):487. https://0-doi-org.brum.beds.ac.uk/10.3390/f13030487
Chicago/Turabian StyleXiao, Yongli, Yanni Yu, Yue Wang, Xuqin Wang, Yuanyuan Wang, Wei Dai, and Yaning Luan. 2022. "Effect of Two Exogenous Organic Acids on the Excitation Effect of Soil Organic Carbon in Beijing, China" Forests 13, no. 3: 487. https://0-doi-org.brum.beds.ac.uk/10.3390/f13030487