
Effect of Simulated Combined N and P on Soil Acidity within Soil Aggregates in Natural and Planted Korean Pine Forest in Northeast China


 ,
,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Experimental Design
2.3. Soil Sampling and Processing
2.4. Chemical Analysis
2.5. Effective Cation Exchange Capacity, Base Saturation%, Al:Ca Ratio, and Effective Acidity Calculated by the Following Formula
3. Statistical Analysis
4. Results
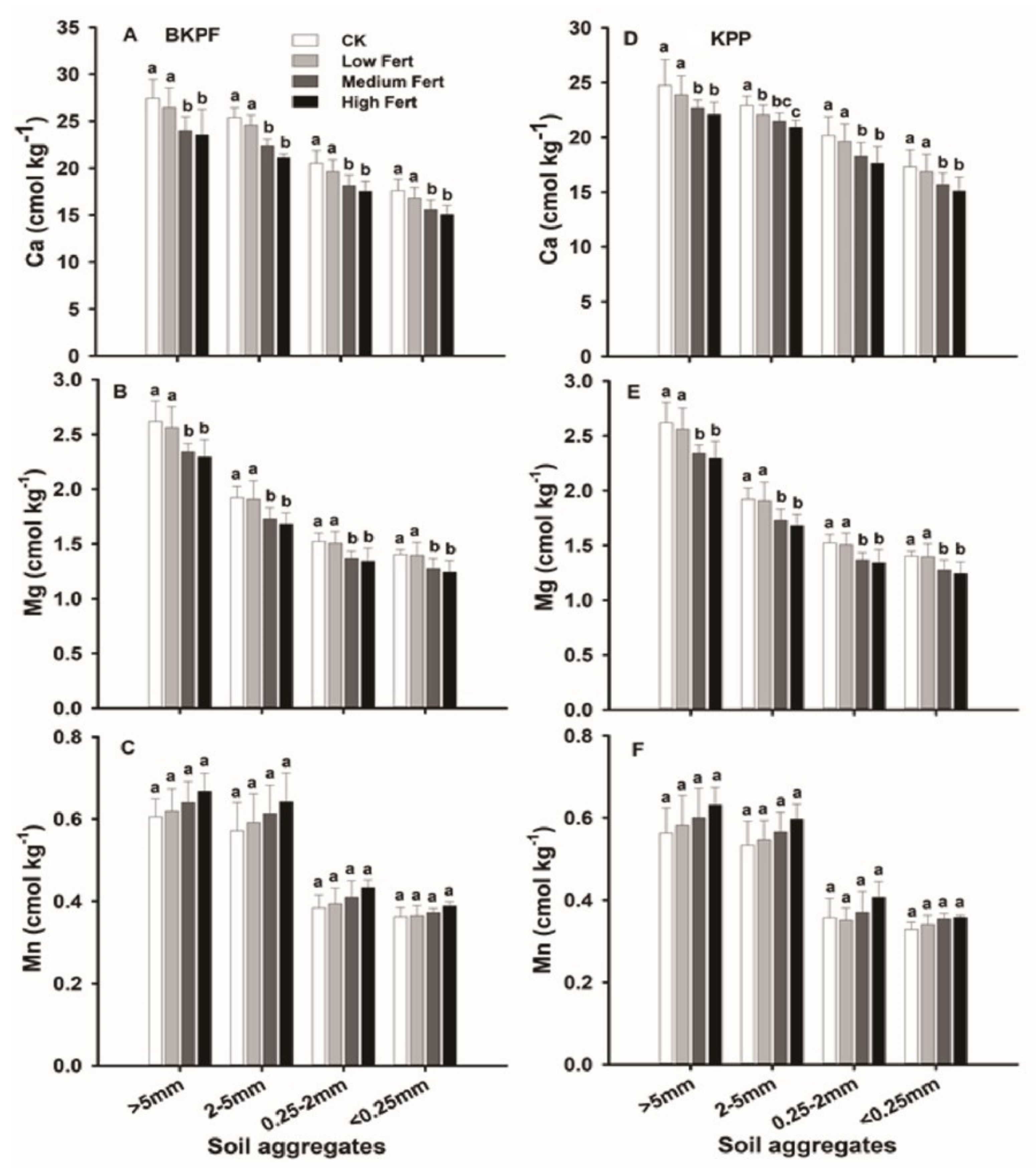
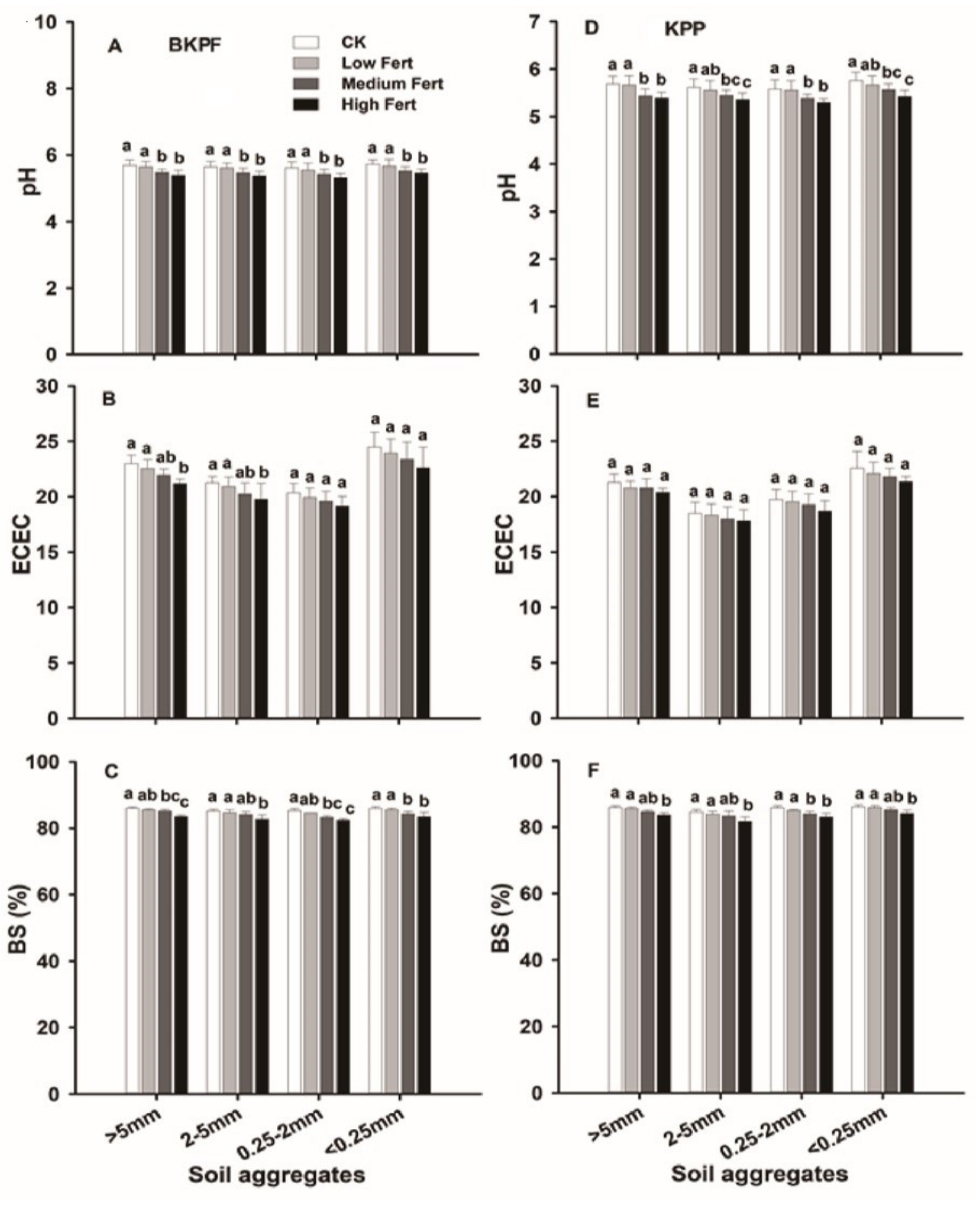
4.1. Impact of Simulated Nitrogen and Phosphorus Deposition on Soil Properties
4.2. Impact of Soil Aggregates on Soil Properties
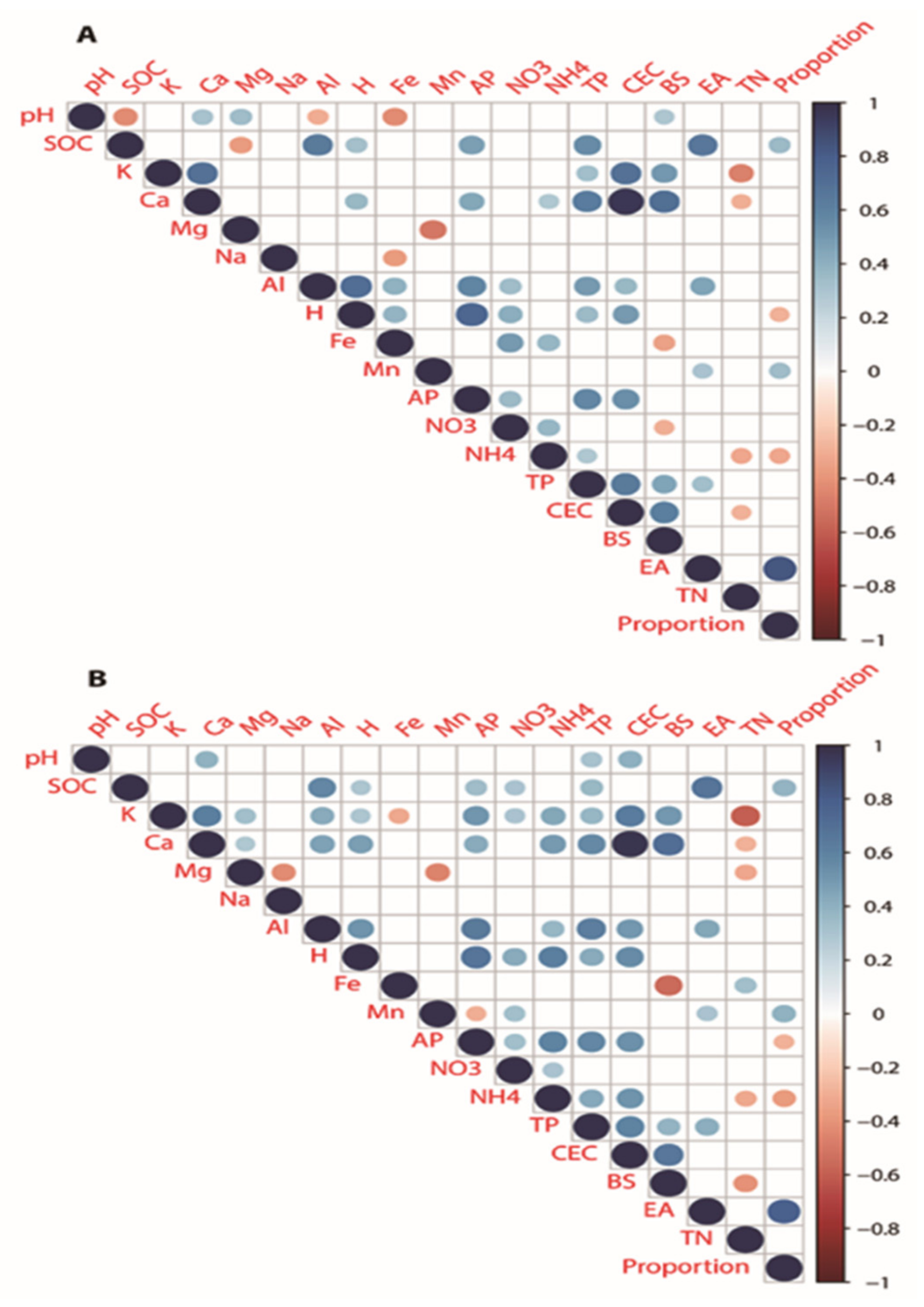
4.3. Correlation of Soil Properties
4.4. Interrelation of Soil Properties with the Soil Related Environmental Parameters
5. Discussion
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Elser, J.J.; Bracken, M.E.; Cleland, E.E.; Gruner, D.S.; Harpole, W.S.; Hillebrand, H.; Ngai, J.T.; Seabloom, E.W.; Shurin, J.B.; Smith, J.E. Global analysis of nitrogen and phosphorus limitation of primary producers in freshwater, marine and terrestrial ecosystems. Ecol. Lett. 2007, 10, 1135–1142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vitousek, P.M.; Porder, S.; Houlton, B.Z.; Chadwick, O.A. Terrestrial phosphorus limitation: Mechanisms, implications, and nitrogen–phosphorus interactions. Ecol. Appl. 2010, 20, 5–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cape, J.N.; Freer-Smith, P.H.; Paterson, I.S.; Parkinson, J.A.; Wolfenden, J.J.T. The nutritional status of Picea abies (L.) Karst. across Europe, and implications for ‘forest decline’. Trees 1990, 4, 211–224. [Google Scholar] [CrossRef]
- Vadas, P.; Sims, J. Soil Fertility: Phosphorus in Soils. J. Environ. Qual. 2014, 28, 1870–1877. [Google Scholar] [CrossRef]
- Hou, E.; Luo, Y.; Kuang, Y.; Chen, C.; Lu, X.; Jiang, L.; Luo, X.; Wen, D. Global meta-analysis shows pervasive phosphorus limitation of aboveground plant production in natural terrestrial ecosystems. Nat. Commun. 2020, 11, 637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- LeBauer, D.S.; Treseder, K.K. Nitrogen limitation of net primary productivity in terrestrial ecosystems is globally distributed. Ecology 2008, 89, 371–379. [Google Scholar] [CrossRef] [Green Version]
- Vitousek, P.M.; Howarth, R.W. Nitrogen limitation on land and in the sea: How can it occur? Biogeochemistry 1991, 13, 87–115. [Google Scholar] [CrossRef]
- Yang, Y.; Ji, C.; Ma, W.; Wang, S.; Wang, S.; Han, W.; Mohammat, A.; Robinson, D.; Smith, P. Significant soil acidification across northern China’s grasslands during 1980s–2000s. Glob. Change Biol. 2012, 18, 2292–2300. [Google Scholar] [CrossRef]
- Bowman, W.D.; Cleveland, C.C.; Halada, Ĺ.; Hreško, J.; Baron, J.S.J.N.G. Negative impact of nitrogen deposition on soil buffering capacity. Nat. Geosci. 2008, 1, 767–770. [Google Scholar] [CrossRef]
- Wang, R.; Dungait, J.A.; Buss, H.L.; Yang, S.; Zhang, Y.; Xu, Z.; Jiang, Y.J. Base cations and micronutrients in soil aggregates as affected by enhanced nitrogen and water inputs in a semi-arid steppe grassland. Sci. Total Environ. 2017, 575, 564–572. [Google Scholar] [CrossRef] [Green Version]
- Malhi, S.; Nyborg, M.; Harapiak, J.T. Effects of long-term N fertilizer-induced acidification and liming on micronutrients in soil and in bromegrass hay. Soil Tillage Res. 1998, 48, 91–101. [Google Scholar] [CrossRef]
- Yang, K.; Zhu, J.; Gu, J.; Yu, L.; Wang, Z. Changes in soil phosphorus fractions after 9 years of continuous nitrogen addition in a Larix gmelinii plantation. Ann. For. Sci. 2015, 72, 435–442. [Google Scholar] [CrossRef]
- Ulrich, B. Natural and anthropogenic components of soil acidification. Z. Pflanz. Bodenkd. 1986, 149, 702–717. [Google Scholar] [CrossRef]
- Małek, S.; Astel, A. The effect of stand age on throughfall chemistry in spruce stands in the Potok Dupniański Catchment in the Silesian Beskid Mountains, Southern Poland. Sci. World J. 2007, 7, 181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Małek, S.; Astel, A. Throughfall chemistry in a spruce chronosequence in southern Poland. Environ. Pollut. 2008, 155, 517–527. [Google Scholar] [CrossRef] [PubMed]
- Reuss, J.O.; Johnson, D.W. Acid Deposition and the Acidification of Soils and Waters; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2012; Volume 59. [Google Scholar]
- Rhoades, C.C. Single-tree influences on soil properties in agroforestry: Lessons from natural forest and savanna ecosystems. Agrofor. Syst. 1996, 35, 71–94. [Google Scholar] [CrossRef]
- Binkley, D.; Giardina, C. Why do tree species affect soils? The warp and woof of tree-soil interactions. In Plant-Induced Soil Changes: Processes and Feedbacks; Springer: Berlin/Heidelberg, Germany, 1998; pp. 89–106. [Google Scholar]
- Bardgett, R.D.; Bowman, W.D.; Kaufmann, R.; Schmidt, S.K. A temporal approach to linking aboveground and belowground ecology. Trends Ecol. Evol. 2005, 20, 634–641. [Google Scholar] [CrossRef] [PubMed]
- West, J.B.; Hobbie, S.E.; Reich, P.B. Effects of plant species diversity, atmospheric [CO2], and N addition on gross rates of inorganic N release from soil organic matter. Glob. Change Biol. 2006, 12, 1400–1408. [Google Scholar] [CrossRef]
- Shi, B.; Jin, G. Variability of soil respiration at different spatial scales in temperate forests. Biol. Fertil. Soils 2016, 52, 561–571. [Google Scholar] [CrossRef]
- Penuelas, J.; Janssens, I.A.; Ciais, P.; Obersteiner, M.; Sardans, J. Anthropogenic global shifts in biospheric N and P concentrations and ratios and their impacts on biodiversity, ecosystem productivity, food security, and human health. Glob. Change Biol. 2020, 26, 1962–1985. [Google Scholar] [CrossRef] [Green Version]
- Jia, Y.; Yu, G.; He, N.; Zhan, X.; Fang, H.; Sheng, W.; Zuo, Y.; Zhang, D.; Wang, Q. Spatial and decadal variations in inorganic nitrogen wet deposition in China induced by human activity. Sci. Rep. 2014, 4, 3763. [Google Scholar] [CrossRef] [Green Version]
- Pan, Y.; Wang, Y.; Tang, G.; Wu, D. Wet and dry deposition of atmospheric nitrogen at ten sites in Northern China. Atmos. Chem. Phys. 2012, 12, 6515–6535. [Google Scholar] [CrossRef] [Green Version]
- Harris, G. Ecological Stoichiometry: Biology of Elements from Molecules to the Biosphere; $29.95; Sterner, R.W., Elser, J.J., Eds.; Princeton University Press: Princeton, NJ, USA, 2002; Oxford University Press: Oxford, UK, 2003; ISSN 0-691-07491-7. [Google Scholar]
- Goll, D.S. Coupled Cycling of Carbon, Nitrogen, and Phosphorus; CRC Press: Boca Raton, FL, USA, 2016; Available online: https://opus.bibliothek.uni-augsburg.de/opus4/frontdoor/index/index/docId/67024 (accessed on 12 February 2022).
- Gress, S.E.; Nichols, T.D.; Northcraft, C.C.; Peterjohn, W.T. Nutrient limitation in soils exhibiting differing nitrogen availabilities: What lies beyond nitrogen saturation? Ecology 2007, 88, 119–130. [Google Scholar] [CrossRef] [Green Version]
- Marklein, A.R.; Houlton, B.Z. Nitrogen inputs accelerate phosphorus cycling rates across a wide variety of terrestrial ecosystems. New Phytol. 2012, 193, 696–704. [Google Scholar] [CrossRef]
- Weand, M.P.; Arthur, M.A.; Lovett, G.M.; McCulley, R.L.; Weathers, K.C. Effects of tree species and N additions on forest floor microbial communities and extracellular enzyme activities. Soil Biol. Biochem. 2010, 42, 2161–2173. [Google Scholar] [CrossRef]
- Sinsabaugh, R.L.; Gallo, M.E.; Lauber, C.; Waldrop, M.P.; Zak, D.R. Extracellular enzyme activities and soil organic matter dynamics for northern hardwood forests receiving simulated nitrogen deposition. Biogeochemistry 2005, 75, 201–215. [Google Scholar] [CrossRef] [Green Version]
- Nave, L.; Vance, E.; Swanston, C.; Curtis, P. Impacts of elevated N inputs on north temperate forest soil C storage, C/N, and net N-mineralization. Geoderma 2009, 153, 231–240. [Google Scholar] [CrossRef]
- Cusack, D.F.; Silver, W.L.; Torn, M.S.; Burton, S.D.; Firestone, M.K. Changes in microbial community characteristics and soil organic matter with nitrogen additions in two tropical forests. Ecology 2011, 92, 621–632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Futa, B.; Mocek-Płóciniak, A. The influence of uncontrolled grass burning on biochemical qualities of soil. J. Res. Appl. Agric. Eng. 2016, 61, 98–100. [Google Scholar]
- Wei, C.; Gao, M.; Shao, J.; Xie, D.; Pan, G. Soil aggregate and its response to land management practices. China Particuology 2006, 4, 211–219. [Google Scholar] [CrossRef]
- Ranatunga, T.D.; Reddy, S.S.; Taylor, R.W. Phosphorus distribution in soil aggregate size fractions in a poultry litter applied soil and potential environmental impacts. Geoderma 2013, 192, 446–452. [Google Scholar] [CrossRef]
- Wei, K.; Chen, Z.; Zhu, A.; Zhang, J.; Chen, L.J.S.; Research, T. Application of 31P NMR spectroscopy in determining phosphatase activities and P composition in soil aggregates influenced by tillage and residue management practices. Soil Tillage Res. 2014, 138, 35–43. [Google Scholar] [CrossRef]
- Zhang, S.; Li, Q.; Lü, Y.; Zhang, X.; Liang, W. Contributions of soil biota to C sequestration varied with aggregate fractions under different tillage systems. Soil Biol. Biochem. 2013, 62, 147–156. [Google Scholar] [CrossRef]
- Wang, R.; Dungait, J.A.; Creamer, C.A.; Cai, J.; Li, B.; Xu, Z.; Zhang, Y.; Ma, Y.; Jiang, Y. Carbon and Nitrogen Dynamics in Soil Aggregates under Long-Term Nitrogen and Water Addition in a Temperate Steppe. Soil Sci. Soc. Am. J. 2015, 79, 527–535. [Google Scholar] [CrossRef]
- Wang, R.; Creamer, C.A.; Wang, X.; He, P.; Xu, Z.; Jiang, Y. The effects of a 9-year nitrogen and water addition on soil aggregate phosphorus and sulfur availability in a semi-arid grassland. Ecol. Indic. 2016, 61, 806–814. [Google Scholar] [CrossRef] [Green Version]
- Wei, T.; Simko, V.; Levy, M.; Xie, Y.; Jin, Y.; Zemla, J. Package ‘corrplot’. Statistician 2017, 56, e24. [Google Scholar]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’hara, R.; Simpson, G.L.; Solymos, P.; Stevens, M.H.H.; Wagner, H. Package ‘vegan’. Community Ecol. Package Version 2013, 2, 1–295. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing. 2013. Available online: https://www.scirp.org/(S(i43dyn45teexjx455qlt3d2q))/reference/ReferencesPapers.aspx?ReferenceID=1787696 (accessed on 12 February 2022).
- Mao, Q.; Lu, X.; Zhou, K.; Chen, H.; Zhu, X.; Mori, T.; Mo, J. Effects of long-term nitrogen and phosphorus additions on soil acidification in an N-rich tropical forest. Geoderma 2017, 285, 57–63. [Google Scholar] [CrossRef]
- Zarif, N.; Khan, A.; Wang, Q. Linking Soil Acidity to P Fractions and Exchangeable Base Cations under Increased N and P Fertilization of Mono and Mixed Plantations in Northeast China. Forests 2020, 11, 1274. [Google Scholar] [CrossRef]
- Dossa, E.; Khouma, M.; Diedhiou, I.; Sene, M.; Kizito, F.; Badiane, A.; Samba, S.; Dick, R. Carbon, nitrogen and phosphorus mineralization potential of semiarid Sahelian soils amended with native shrub residues. Geoderma 2009, 148, 251–260. [Google Scholar] [CrossRef]
- Cronan, C.S.; Grigal, D.F. Use of calcium/aluminum ratios as indicators of stress in forest ecosystems. J. Environ. Qual. 1995, 24, 209–226. [Google Scholar] [CrossRef]
- Mori, T.; Ohta, S.; Ishizuka, S.; Konda, R.; Wicaksono, A.; Heriyanto, J.; Hardjono, A.J.S.S.; Nutrition, P. Effects of phosphorus addition on N2O and NO emissions from soils of an Acacia mangium plantation. Soil Sci. Plant Nutr. 2010, 56, 782–788. [Google Scholar] [CrossRef] [Green Version]
- Katou, H. A pH-dependence implicit formulation of cation-and anion-exchange capacities of variable-charge soils. Soil Sci. Soc. Am. J. 2002, 66, 1218–1224. [Google Scholar] [CrossRef]
- Yin, J.; Wang, R.; Liu, H.; Feng, X.; Xu, Z.; Jiang, Y. Nitrogen addition alters elemental stoichiometry within soil aggregates in a temperate steppe. Solid Earth 2016, 7, 1565–1575. [Google Scholar] [CrossRef]
- Lu, X.; Mao, Q.; Gilliam, F.S.; Luo, Y.; Mo, J. Nitrogen deposition contributes to soil acidification in tropical ecosystems. Glob. Change Biol. 2014, 20, 3790–3801. [Google Scholar] [CrossRef]
- Lucas, R.; Klaminder, J.; Futter, M.; Bishop, K.H.; Egnell, G.; Laudon, H.; Högberg, P. A meta-analysis of the effects of nitrogen additions on base cations: Implications for plants, soils, and streams. For. Ecol. Manag. 2011, 262, 95–104. [Google Scholar] [CrossRef]
- Matschonat, G.; Matzner, E. Soil chemical properties affecting NH4+ sorption in forest soils. Z. Pflanz. Bodenkd. 1996, 159, 505–511. [Google Scholar] [CrossRef]
- Smith, S.; Read, D. Mycorrhizal Symbiosis, 3rd ed.; Academic Press: London, UK, 2008. [Google Scholar]
- Tafazoli, M.; Hojjati, S.M.; Jalilvand, H.; Lamersdorf, N. Simulated Nitrogen Deposition Reduces the Concentration of Soil Base Cations in Acer velutinum Bioss. Plantation, North of Iran. J. Soil Sci. Plant Nutr. 2019, 19, 440–449. [Google Scholar] [CrossRef]
- Uehara, G.; Gillman, G. The Mineralogy, Chemistry, and Physics of Tropical Soils with Variable Charge Clays; Westview Press, Inc.: Boulder, CO, USA, 1981. [Google Scholar]
- Sverdrup, H.; Rosen, K. Long-term base cation mass balances for Swedish forests and the concept of sustainability. For. Ecol. Manag. 1998, 110, 221–236. [Google Scholar] [CrossRef]
- Watmough, S.A.; Dillon, P. Base cation and nitrogen budgets for a mixed hardwood catchment in south-central Ontario. Ecosystems 2003, 6, 675–693. [Google Scholar] [CrossRef]
- Huntington, T.G. Assessment of calcium status in Maine forests: Review and future projection. Can. J. For. Res. 2005, 35, 1109–1121. [Google Scholar] [CrossRef]
- Horswill, P.; O’Sullivan, O.; Phoenix, G.K.; Lee, J.A.; Leake, J.R. Base cation depletion, eutrophication and acidification of species-rich grasslands in response to long-term simulated nitrogen deposition. Environ. Pollut. 2008, 155, 336–349. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Mao, Q.; Mo, J.; Gilliam, F.S.; Zhou, G.; Luo, Y.; Zhang, W.; Huang, J. Divergent responses of soil buffering capacity to long-term N deposition in three typical tropical forests with different land-use history. Environ. Sci. Technol. 2015, 49, 4072–4080. [Google Scholar] [CrossRef] [PubMed]
- Mo, J.; Zhang, W.; Zhu, W.; Gundersen, P.; Fang, Y.; Li, D.; Wang, H. Nitrogen addition reduces soil respiration in a mature tropical forest in southern China. Glob. Change Biol. 2008, 14, 403–412. [Google Scholar] [CrossRef]
- Lovett, G.M.; Arthur, M.A.; Weathers, K.C.; Fitzhugh, R.D.; Templer, P.H. Nitrogen addition increases carbon storage in soils, but not in trees, in an eastern US deciduous forest. Ecosystems 2013, 16, 980–1001. [Google Scholar] [CrossRef]
- Xian-Kai, L.; Jiang-Ming, M.; Gundersern, P.; Wei-Xing, Z.; Guo-Yi, Z.; De-Jun, L.; Zhang, X. Effect of simulated N deposition on soil exchangeable cations in three forest types of subtropical China. Pedosphere 2009, 19, 189–198. [Google Scholar]
- Parfitt, R.L.; Giltrap, D.J.; Whitton, J.S. Contribution of organic matter and clay minerals to the cation exchange capacity of soils. Commun. Soil Sci. Plant Anal. 1995, 26, 1343–1355. [Google Scholar] [CrossRef]
- Šimanský, V.; Polláková, N. Soil organic matter and sorption capacity under different soil management practices in a productive vineyard. Arch. Agron. Soil Sci. 2014, 60, 1145–1154. [Google Scholar] [CrossRef]
- Gundersen, P.; Schmidt, I.K.; Raulund-Rasmussen, K. Leaching of nitrate from temperate forests effects of air pollution and forest management. Environ. Rev. 2006, 14, 1–57. [Google Scholar] [CrossRef]
- Finzi, A.C.; Canham, C.D.; Van Breemen, N. Canopy tree–soil interactions within temperate forests: Species effects on pH and cations. Ecol. Appl. 1998, 8, 447–454. [Google Scholar]
- Rigueiro-Rodríguez, A.; Mosquera-Losada, M.; Fernández-Núñez, E. Afforestation of agricultural land with Pinus radiata D. Don and Betula alba L. in nw Spain: Effects on soil pH, understorey production and floristic diversity eleven years after establishment. Land Degrad. Dev. 2012, 23, 227–241. [Google Scholar] [CrossRef]
- Sollins, P.; McCorison, F. Nitrogen and carbon solution chemistry of an old growth coniferous forest watershed before and after cutting. Water Resour. Res. 1981, 17, 1409–1418. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site Characteristics of Forest Stands | |||||||
|---|---|---|---|---|---|---|---|
| Forest Stand | Main Species Composition | Land Use History | Age | Stand Density | Mean DBH | Canopy Closure | Soil Density |
| Forest Type | (Year) | (Trees/hm−2) | (cm) | (g/m−3) | |||
| Broad-leaved Korean pine forest (BKPF) | Pinus koraiensis, Betula costata, Tilia amurensis, Acer ukurunduense, Abies nephrolepis, Ulmus laciniata, Acer tegmentosum, and Fraxinus Mandshurica | Natural forest | >230 | 1175 | 21.1 | 0.75 | 0.99 |
| Planted Korean pine plantation (KPP) | Pinus koraiensis | Reforestation in 1954 after clear-cutting of primary mixed broad-leaved Korean pine forest | 65 | 1475 | 26.4 | 0.7 | 0.80 |
| Soil Aggregates (mm) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Bulk Soil | >5 mm | 2–5 mm | 0.25–2 mm | >0.25 mm | ||||||
| BKP | KPP | BKPF | KPP | BKPF | KPP | BKPF | KPP | BKPF | KPP | |
| pH | 5.70 | 5.65 | 5.65 | 5.61 | 5.57 | 5.60 | 5.60 | 5.58 | 5.72 | 5.61 |
| SOC (g kg−1) | 63.2 | 60.3 | 59.23 | 57.3 | 55.4 | 52.54 | 48.4 | 46.5 | 44.4 | 42.3 |
| AP (mg kg−1) | 40.2 | 36.2 | 24.20 | 21.5 | 22.3 | 20.10 | 15.2 | 14.1 | 13.5 | 13.0 |
| TN (g kg−1) | 4.70 | 4.10 | 3.50 | 3.30 | 2.90 | 2.50 | 2.50 | 2.30 | 1.90 | 1.80 |
| TP (g kg−1) | 3.10 | 2.90 | 2.10 | 1.90 | 1.60 | 1.45 | 1.76 | 1.35 | 1.20 | 1.05 |
| C:N (ratio) | 13.4 | 14.7 | 16.9 | 17.3 | 19.1 | 21.01 | 19.3 | 20.2 | 23.3 | 23.5 |
| NH4–N (mg kg−1) | 40.6 | 38.2 | 36.8 | 34.2 | 34.1 | 33.06 | 24.5 | 23.1 | 24.3 | 22.4 |
| NO3–N (mg kg−1) | 6.40 | 5.30 | 7.43 | 6.20 | 6.50 | 6.10 | 4.20 | 3.90 | 3.80 | 3.60 |
| Source of Variations | df | pH | SOC | C/N | K+ | Ca2+ | Mg2+ | Na+ | Al3+ | H+ | Fe3+ | Mn2+ |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Forest types (FT) | 1 | 0.799 | 0.204 | 0.153 | 0.009 | 0.001 | 0.127 | 0.184 | 0.001 | 0.001 | 0.316 | 0.027 |
| Soil aggregates (SA) | 3 | 0.385 | 0.001 | 0.066 | 0.010 | 0.001 | 0.422 | 0.509 | 0.203 | 0.001 | 0.004 | 0.001 |
| Fertilization (Fert) | 3 | 0.001 | 0.001 | 0.295 | 0.137 | 0.001 | 0.008 | 0.896 | 0.002 | 0.001 | 0.001 | 0.081 |
| FT × SA | 3 | 0.997 | 0.998 | 0.992 | 0.966 | 0.131 | 0.907 | 1.000 | 0.997 | 0.508 | 0.944 | 0.997 |
| FT × Fert | 3 | 1.000 | 0.998 | 1.000 | 1.000 | 0.778 | 0.893 | 1.000 | 0.845 | 0.965 | 0.900 | 1.000 |
| SA × Fert | 9 | 1.000 | 1.000 | 1.000 | 1.000 | 1.000 | 1.000 | 1.000 | 1.000 | 0.999 | 1.000 | 1.000 |
| FT × SA × Fert | 9 | 1.000 | 1.000 | 1.000 | 1.000 | 1.000 | 1.000 | 1.000 | 1.000 | 1.000 | 1.000 | 1.000 |
| Source of variations | df | Ap | NO3 | NH4 | TP | Al:Ca | ECEC | BS % | E.A | TN | Proportion % | |
| Forest types (FT) | 1 | 0.933 | 0.001 | 0.108 | 0.039 | 0.001 | 0.001 | 0.833 | 0.001 | 0.196 | 0.397 | |
| Soil aggregates (SA) | 3 | 0.018 | 0.169 | 0.001 | 0.003 | 0.001 | 0.001 | 0.002 | 0.001 | 0.529 | 0.001 | |
| Fertilization (Fert) | 3 | 0.039 | 0.001 | 0.449 | 0.163 | 0.001 | 0.085 | 0.001 | 0.001 | 0.911 | 0.956 | |
| FT × SA | 3 | 0.966 | 0.943 | 0.928 | 0.994 | 0.791 | 0.288 | 0.255 | 0.561 | 1.000 | 0.105 | |
| FT × Fert | 3 | 0.935 | 0.881 | 1.000 | 1.000 | 0.281 | 0.918 | 0.998 | 0.841 | 1.000 | 0.955 | |
| SA × Fert | 9 | 1.000 | 0.998 | 1.000 | 1.000 | 0.99 | 1.000 | 0.999 | 0.990 | 1.000 | 0.826 | |
| FT × SA × Fert | 9 | 1.000 | 1.000 | 1.000 | 1.000 | 0.999 | 1.000 | 1.000 | 1.000 | 1.000 | 0.976 | |
| Aggregate Size | Al3+ (cmol kg−1) | H+ (cmol kg−1) | Fe3+ (cmol kg−1) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| C | Low NP | Medium NP | High NP | C | Low NP | Medium NP | High NP | C | Low NP | Medium NP | High NP | |
| BKFP | ||||||||||||
| >5 mm | 4.99 | 5.11 | 5.51 | 5.72 | 0.64 | 0.66 | 0.68 | 0.72 | 2.91 | 3.01 | 3.26 | 3.37 |
| 2–5 mm | 4.72 | 4.78 | 5.43 | 5.60 | 0.53 | 0.56 | 0.58 | 0.61 | 2.77 | 2.70 | 3.19 | 3.32 |
| 0.25–2 mm | 3.39 | 3.43 | 3.62 | 3.65 | 0.38 | 0.40 | 0.42 | 0.45 | 2.35 | 2.30 | 2.57 | 2.67 |
| <0.25 mm | 3.02 | 3.18 | 3.52 | 3.58 | 0.32 | 0.33 | 0.35 | 0.38 | 2.14 | 2.09 | 2.33 | 2.43 |
| KPP | ||||||||||||
| >5 mm | 4.36 | 4.47 | 4.73 | 4.88 | 0.59 | 0.60 | 0.61 | 0.64 | 2.8 | 2.9 | 3.1 | 3.2 |
| 2–5 mm | 4.15 | 4.19 | 4.58 | 4.64 | 0.48 | 0.48 | 0.52 | 0.55 | 2.7 | 2.8 | 3.0 | 3.1 |
| 0.25–2 mm | 2.95 | 2.97 | 3.13 | 3.21 | 0.36 | 0.37 | 0.39 | 0.41 | 2.3 | 2.4 | 2.6 | 2.7 |
| <0.25 mm | 2.75 | 2.77 | 2.97 | 3.02 | 0.30 | 0.31 | 0.32 | 0.34 | 2.1 | 2.2 | 2.4 | 2.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jamil, M.A.; Hussain, A.; Duan, W.; Chen, L.; Khan, K.; Abid, K.; Li, C.; Guo, Q.; Zarif, N.; Qu, M.; et al. Effect of Simulated Combined N and P on Soil Acidity within Soil Aggregates in Natural and Planted Korean Pine Forest in Northeast China. Forests 2022, 13, 529. https://0-doi-org.brum.beds.ac.uk/10.3390/f13040529
Jamil MA, Hussain A, Duan W, Chen L, Khan K, Abid K, Li C, Guo Q, Zarif N, Qu M, et al. Effect of Simulated Combined N and P on Soil Acidity within Soil Aggregates in Natural and Planted Korean Pine Forest in Northeast China. Forests. 2022; 13(4):529. https://0-doi-org.brum.beds.ac.uk/10.3390/f13040529
Chicago/Turabian StyleJamil, Muhammad Atif, Anwaar Hussain, Wenbiao Duan, Lixin Chen, Kashif Khan, Kulsoom Abid, Changzhun Li, Qiwen Guo, Nowsherwan Zarif, Meixue Qu, and et al. 2022. "Effect of Simulated Combined N and P on Soil Acidity within Soil Aggregates in Natural and Planted Korean Pine Forest in Northeast China" Forests 13, no. 4: 529. https://0-doi-org.brum.beds.ac.uk/10.3390/f13040529