
Stimulation, Reduction and Compensation Growth, and Variable Phenological Responses to Spring and/or Summer–Autumn Warming in Corylus Taxa and Cornus sanguinea L.

 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
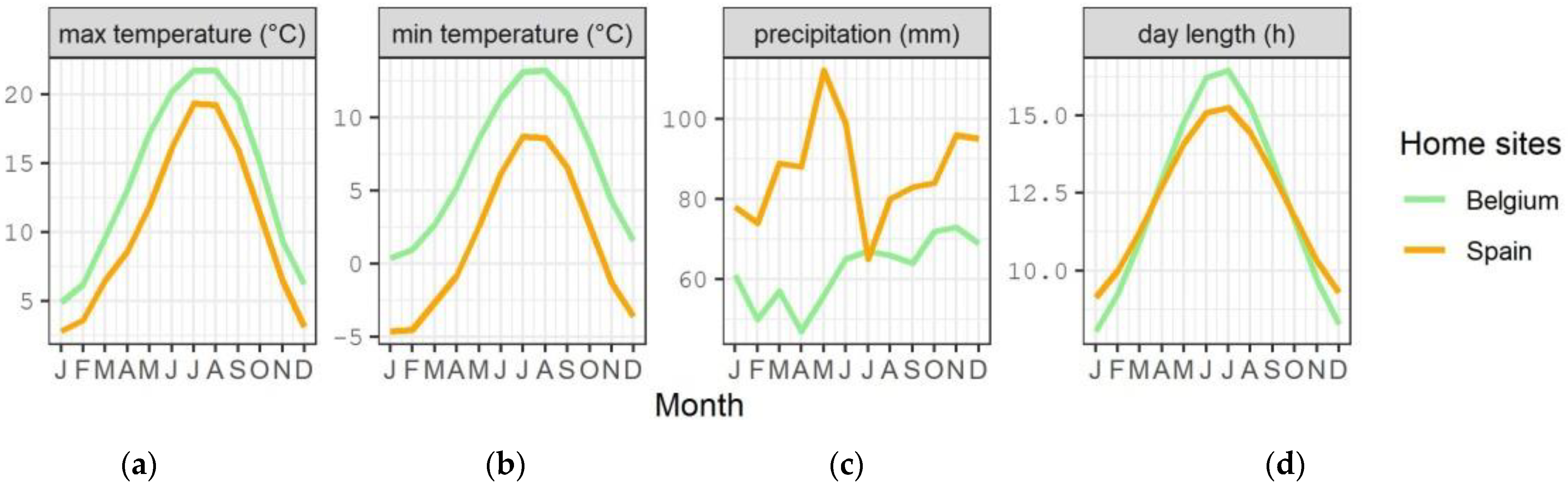
2.1. Seed Collection and Germination
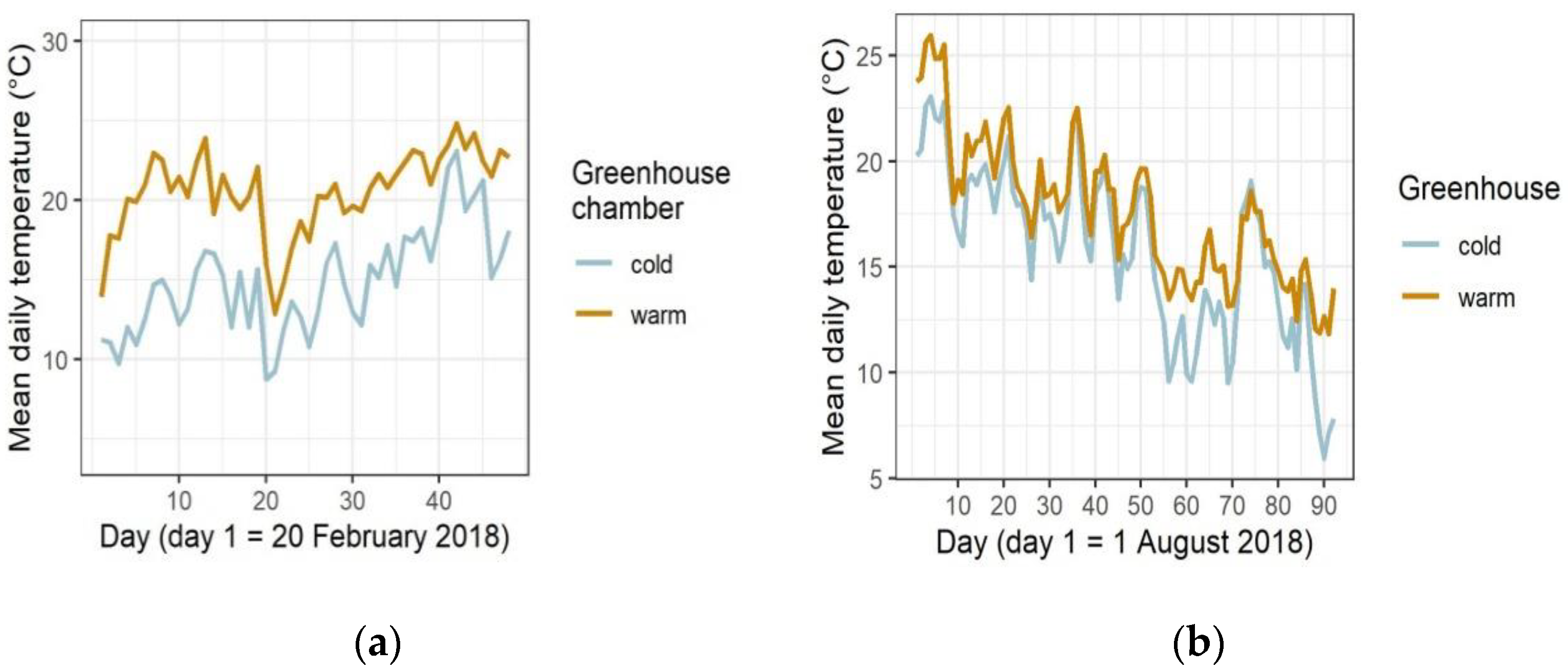
2.2. Temperature Treatments
2.3. Measurements and Phenological Observations
2.4. Statistical Analysis
3. Results
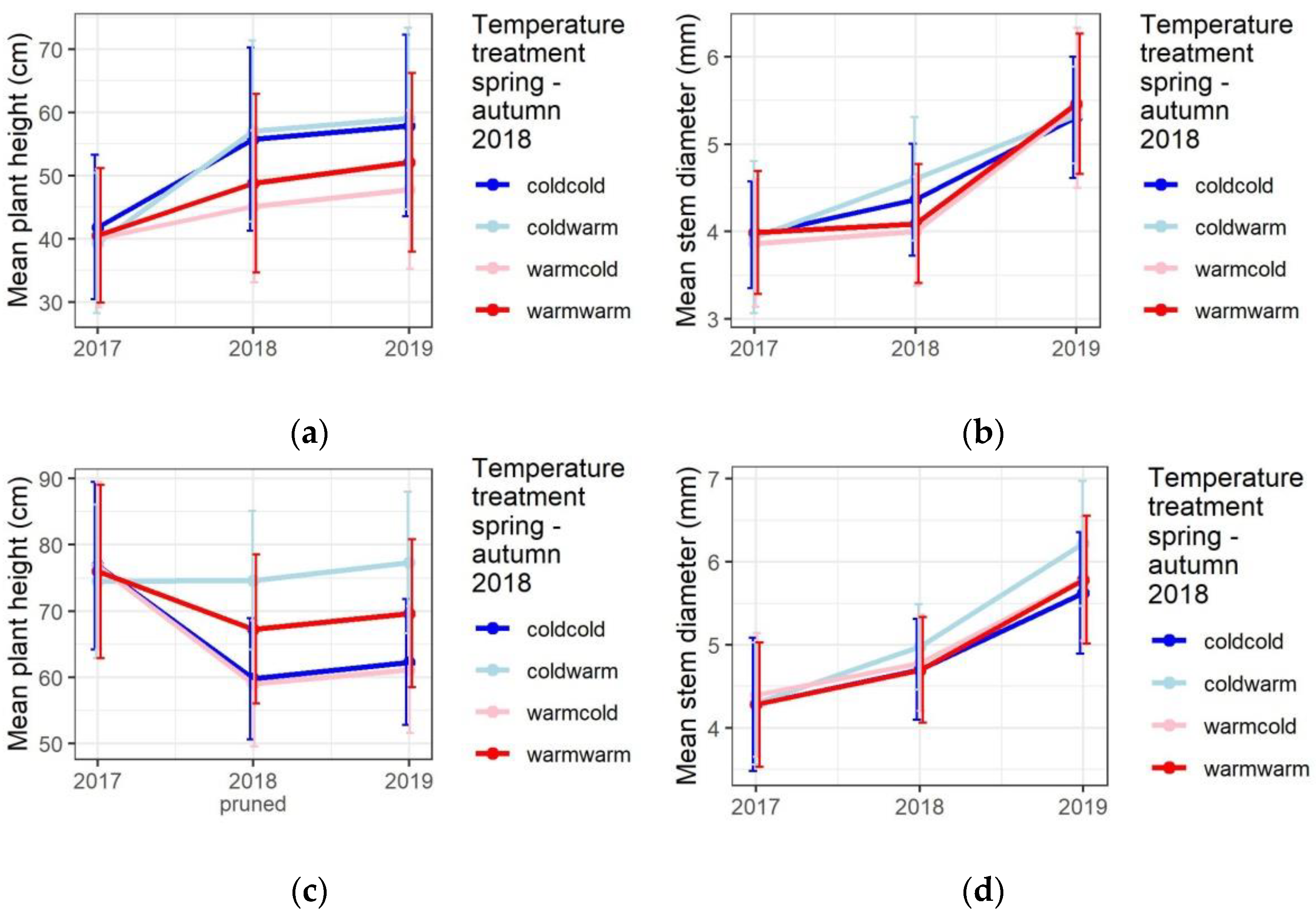
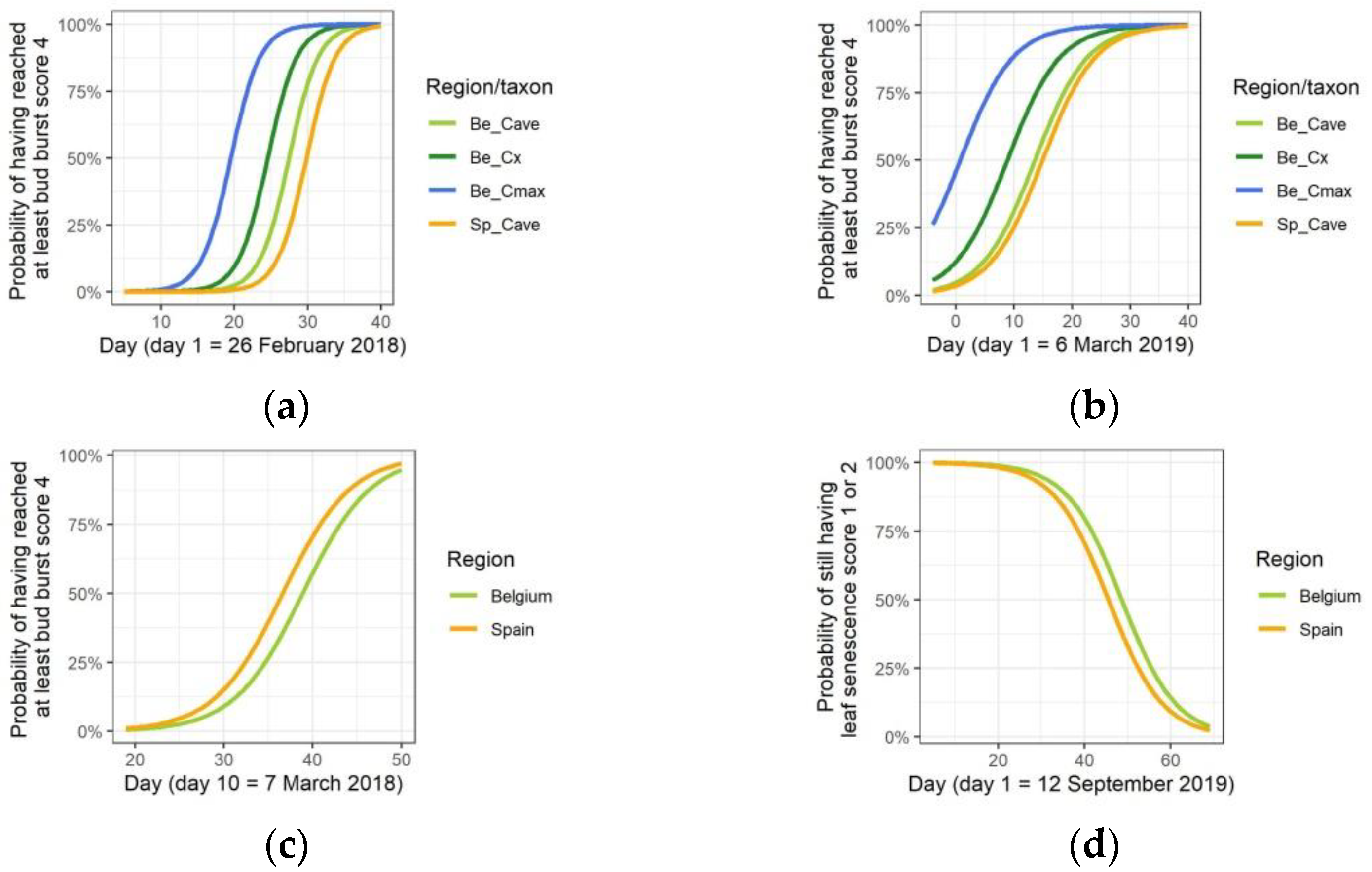
3.1. Temperature Treatments Affect the Growth
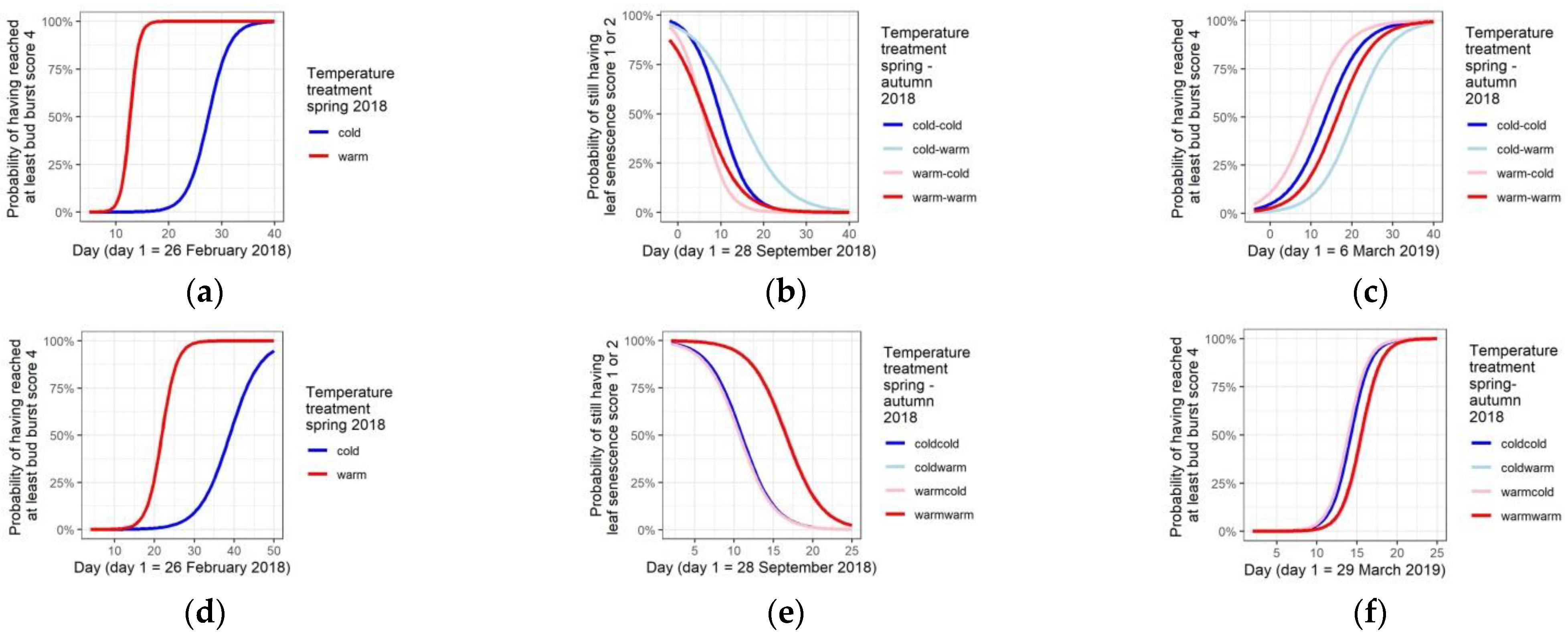
3.2. Temperature Treatments Affect Phenology
3.3. Growth Differentiation among the Taxa and Provenances in the Common Garden
3.4. Phenological Differentiation among the Taxa and Provenances in the Common Garden
4. Discussion
4.1. Responses to Spring and Summer–Autumn Warming
4.2. Common Garden
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Peñuelas, J.; Ciais, P.; Canadell, J.G.; Janssens, I.A.; Fernández-Martínez, M.; Carnicer, J.; Obersteiner, M.; Piao, S.; Vautard, R.; Sardans, J. Shifting from a fertilization-dominated to a warming-dominated period. Nat. Ecol. Evol. 2017, 1, 1438–1445. [Google Scholar] [CrossRef] [Green Version]
- IPCC. Climate Change 2022: Impacts, Adaptation, and Vulnerability. Contribution of Working Group II to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Pörtner, H.-O., Roberts, D.C., Tignor, M., Poloczanska, E.S., Mintenbeck, K., Alegría, A., Craig, M., Langsdorf, S., Löschke, S., Möller, V., et al., Eds.; Cambridge University Press: Cambridge, UK, 2022. [Google Scholar]
- Wang, D.; Wang, H.; Wang, P.; Ling, T.; Tao, W.; Yang, Z. Warming Treatment Methodology Affected the Response of Plant Ecophysiological Traits to Temperature Increases: A Quantitive Meta-Analysis. Front. Plant Sci. 2019, 10, 957. [Google Scholar] [CrossRef] [PubMed]
- Chung, H.; Muraoka, H.; Nakamura, M.; Han, S.; Muller, O.; Son, Y. Experimental warming studies on tree species and forest ecosystems: A literature review. J. Plant Res. 2013, 126, 447–460. [Google Scholar] [CrossRef] [PubMed]
- Zohner, C.M.; Renner, S.S. Ongoing seasonally uneven climate warming leads to earlier autumn growth cessation in deciduous trees. Oecologia 2019, 189, 549–561. [Google Scholar] [CrossRef] [PubMed]
- Malyshev, A.V. Warming Events Advance or Delay Spring Phenology by Affecting Bud Dormancy Depth in Trees. Front. Plant Sci. 2020, 11, 856. [Google Scholar] [CrossRef]
- Beil, I.; Kreyling, J.; Meyer, C.; Lemcke, N.; Malyshev, A.V. Late to bed, late to rise-Warmer autumn temperatures delay spring phenology by delaying dormancy. Glob. Change Biol. 2021, 27, 5806–5817. [Google Scholar] [CrossRef] [PubMed]
- Piao, S.; Ciais, P.; Friedlingstein, P.; Peylin, P.; Reichstein, M.; Luyssaert, S.; Margolis, H.; Fang, J.; Barr, A.; Chen, A.; et al. Net carbon dioxide losses of northern ecosystems in response to autumn warming. Nature 2008, 451, 49–52. [Google Scholar] [CrossRef]
- Sittaro, F.; Paquette, A.; Messier, C.; Nock, C.A. Tree range expansion in eastern North America fails to keep pace with climate warming at northern range limits. Glob. Change Biol. 2017, 23, 3292–3301. [Google Scholar] [CrossRef]
- Polgar, C.A.; Primack, R.B. Leaf-out phenology of temperate woody plants: From trees to ecosystems. New Phytol. 2011, 191, 926–941. [Google Scholar] [CrossRef]
- Piao, S.; Liu, Q.; Chen, A.; Janssens, I.A.; Fu, Y.; Dai, J.; Liu, L.; Lian, X.; Shen, M.; Zhu, X. Plant phenology and global climate change: Current progresses and challenges. Glob. Change Biol. 2019, 25, 1922–1940. [Google Scholar] [CrossRef]
- Fu, Y.S.; Campioli, M.; Vitasse, Y.; De Boeck, H.J.; Van den Berge, J.; AbdElgawad, H.; Asard, H.; Piao, S.; Deckmyn, G.; Janssens, I.A. Variation in leaf flushing date influences autumnal senescence and next year’s flushing date in two temperate tree species. Proc. Natl. Acad. Sci. USA 2014, 111, 7355–7360. [Google Scholar] [CrossRef] [Green Version]
- Penuelas, J.; Rutishauser, T.; Filella, I. Ecology. Phenology feedbacks on climate change. Science 2009, 324, 887–888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vitasse, Y.; Delzon, S.; Bresson, C.C.; Michalet, R.; Kremer, A. Altitudinal differentiation in growth and phenology among populations of temperate-zone tree species growing in a common garden. Can. J. For. Res.-Rev. Can. Rech. For. 2009, 39, 1259–1269. [Google Scholar] [CrossRef] [Green Version]
- An, N.; Turp, M.T.; Türkeş, M.; Kurnaz, M.L. Mid-Term Impact of Climate Change on Hazelnut Yield. Agriculture 2020, 10, 159. [Google Scholar] [CrossRef]
- Saxe, H.; Cannell, M.G.R.; Johnsen, B.; Ryan, M.G.; Vourlitis, G. Tree and forest functioning in response to global warming. New Phytol. 2001, 149, 369–399. [Google Scholar] [CrossRef] [PubMed]
- Arend, M.; Brem, A.; Kuster, T.M.; Gunthardt-Goerg, M.S. Seasonal photosynthetic responses of European oaks to drought and elevated daytime temperature. Plant Biol. 2013, 15, 169–176. [Google Scholar] [CrossRef]
- Way, D.A.; Oren, R. Differential responses to changes in growth temperature between trees from different functional groups and biomes: A review and synthesis of data. Tree Physiol. 2010, 30, 669–688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reich, P.B.; Sendall, K.M.; Rice, K.; Rich, R.L.; Stefanski, A.; Hobbie, S.E.; Montgomery, R.A. Geographic range predicts photosynthetic and growth response to warming in co-occurring tree species. Nat. Clim. Change 2015, 5, 148–152. [Google Scholar] [CrossRef]
- Fu, Y.H.; Piao, S.; Delpierre, N.; Hao, F.; Hänninen, H.; Liu, Y.; Sun, W.; Janssens, I.A.; Campioli, M. Larger temperature response of autumn leaf senescence than spring leaf-out phenology. Glob. Change Biol. 2018, 24, 2159–2168. [Google Scholar] [CrossRef]
- Yan, T.; Fu, Y.; Campioli, M.; Peñuelas, J.; Wang, X. Divergent responses of phenology and growth to summer and autumnal warming. Glob. Change Biol. 2021, 27, 2905–2913. [Google Scholar] [CrossRef]
- Popescu, I.; Caudullo, G.; de Rigo, D. Cornus sanguinea in Europe: Distribution, habitat, usage and threats. In European Atlas of Forest Tree Species; Publications Office of the European Union: Luxembourg, 2016. [Google Scholar]
- Enescu, C.; Durrant, T.; de Rigo, D.; Caudullo, G. Corylus avellana in Europe: Distribution, habitat, usage and threats. In European Atlas of Forest Tree Species; Publication Office of the European Union: Luxembourg, 2016. [Google Scholar]
- Uyttenbroeck, R.; De Vos, B.; Vander Mijnsbrugge, K. Verspreiding en Standplaats van Inheemse Bomen en Struiken in Vlaanderen. Onderzoek naar de Relatie Tussen Voorkomen, Bodem en Omgevingskenmerken; INBO.R.2014.1388322; Instituut voor Natuur-en Bosonderzoek: Brussel, Belgium, 2014. [Google Scholar]
- Ferreira, J.J.; Garcia-González, C.; Tous, J.; Rovira, M. Genetic diversity revealed by morphological traits and ISSR markers in hazelnut germplasm from northern Spain. Plant Breed. 2010, 129, 435–441. [Google Scholar] [CrossRef]
- Campa, A.; Trabanco, N.; Pérez-Vega, E.; Rovira, M.; Ferreira, J.J. Genetic relationship between cultivated and wild hazelnuts (Corylus avellana L.) collected in northern Spain. Plant Breed. 2011, 130, 360–366. [Google Scholar] [CrossRef]
- Gönenç, S.; Tanrivermis, H.; Bülbül, M. Economic assessment of hazelnut production and the importance of supply management approaches in Turkey. J. Agric. Rural Dev. Trop. Subtrop. 2006, 107, 19–32. [Google Scholar]
- Čokeša, V.; Pavlović, B.; Stajic, S.; Poduška, Z.; Jović, Đ. Corylus L.: Its diversity, geographical distribution and morpho-anatomical characteristics with special reference to the systematic classification and phylogenics of Turkish hazel (Corylus colurna L.). Sustain. For. Collect. 2020, 81–82, 1–17. [Google Scholar] [CrossRef]
- Palmé, A.E. Chloroplast DNA variation, postglacial recolonization and hybridization in hazel, Corylus avellana. Mol. Ecol. 2002, 11, 1769–1779. [Google Scholar] [CrossRef] [PubMed]
- Kollmann, J.; Grubb, P.J. Biological Flora of Central Europe: Cornus sanguinea L. Flora 2001, 196, 161–179. [Google Scholar] [CrossRef]
- Vander Mijnsbrugge, K.; Malanguis, J.M.; Moreels, S.; Turcsán, A.; Van der Schueren, N.; Notivol Paino, E. Direct Phenological Responses but Later Growth Stimulation upon Spring and Summer/Autumn Warming of Prunus spinosa L. in a Common Garden Environment. Forests 2022, 13, 23. [Google Scholar] [CrossRef]
- Vander Mijnsbrugge, K.; Depypere, L.; Michiels, B.; De Cuyper, B. Genetic and temporal plastic variation in bud burst, bud set and flower opening responses of local versus non-local provenances of Prunus spinosa in a provenance trial. Basic Appl. Ecol. 2016, 17, 262–272. [Google Scholar] [CrossRef]
- R_Core_Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Bates, D.; Machler, M.; Bolker, B.M.; Walker, S.C. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Christensen, R.H.B. Ordinal: Regression Models for Ordinal Data. R Package Version 2015.6-28. 2015. Available online: http://www.cran.r-project.org/package=ordinal/ (accessed on 1 April 2019).
- Vander Mijnsbrugge, K.; Malanguis, J.M.; Moreels, S.; Lauwers, A.; Thomaes, A.; De Keersmaeker, L.; Vandekerkhove, K. Growth Recovery and Phenological Responses of Juvenile Beech (Fagus sylvatica L.) Exposed to Spring Warming and Late Spring Frost. Forests 2021, 12, 1604. [Google Scholar] [CrossRef]
- Turcsán, A.; Steppe, K.; Sárközi, E.; Erdélyi, É.; Missoorten, M.; Mees, G.; Vander Mijnsbrugge, K. Early summer drought stress during the first growing year stimulates extra shoot growth in oak seedlings (Quercus petraea). Front. Plant Sci. 2016, 7, 193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spiess, N.; Oufir, M.; Matusikova, I.; Stierschneider, M.; Kopecky, D.; Homolka, A.; Burg, K.; Fluch, S.; Hausman, J.F.; Wilhelm, E. Ecophysiological and transcriptomic responses of oak (Quercus robur) to long-term drought exposure and rewatering. Environ. Exp. Bot. 2012, 77, 117–126. [Google Scholar] [CrossRef]
- Gill, A.L.; Gallinat, A.S.; Sanders-DeMott, R.; Rigden, A.J.; Short Gianotti, D.J.; Mantooth, J.A.; Templer, P.H. Changes in autumn senescence in northern hemisphere deciduous trees: A meta-analysis of autumn phenology studies. Ann. Bot. 2015, 116, 875–888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strømme, C.B.; Julkunen-Tiitto, R.; Olsen, J.E.; Nybakken, L.; Tognetti, R. High daytime temperature delays autumnal bud formation in Populus tremula under field conditions. Tree Physiol. 2017, 37, 71–81. [Google Scholar] [CrossRef]
- McKown, A.D.; Guy, R.D.; Klapste, J.; Geraldes, A.; Friedmann, M.; Cronk, Q.C.B.; El-Kassaby, Y.A.; Mansfield, S.D.; Douglas, C.J. Geographical and environmental gradients shape phenotypic trait variation and genetic structure in Populus trichocarpa. New Phytol. 2014, 201, 1263–1276. [Google Scholar] [CrossRef]
- Wu, C.; Wang, X.; Wang, H.; Ciais, P.; Peñuelas, J.; Myneni, R.B.; Desai, A.R.; Gough, C.M.; Gonsamo, A.; Black, A.T.; et al. Contrasting responses of autumn-leaf senescence to daytime and night-time warming. Nat. Clim. Change 2018, 8, 1092–1096. [Google Scholar] [CrossRef] [Green Version]
- Jeong, S.J.; Ho, C.H.; Gim, H.J.; Brown, M.E. Phenology shifts at start vs. end of growing season in temperate vegetation over the Northern Hemisphere for the period 1982–2008. Glob. Change Biol. 2011, 17, 2385–2399. [Google Scholar] [CrossRef]
- Fridley, J.D. Extended leaf phenology and the autumn niche in deciduous forest invasions. Nature 2012, 485, 359–362. [Google Scholar] [CrossRef]
- Salminen, H.; Jalkanen, R.; Lindholm, M. Summer temperature affects the ratio of radial and height growth of Scots pine in northern Finland. Ann. For. Sci. 2009, 66, 810. [Google Scholar] [CrossRef]
- Lin, Y.-S.; Medlyn, B.E.; Ellsworth, D.S. Temperature responses of leaf net photosynthesis: The role of component processes. Tree Physiol. 2012, 32, 219–231. [Google Scholar] [CrossRef] [Green Version]
- Hill, M.O.; Mountford, J.O.; Roy, D.B.; Bince, R.G.H. Ellenberg’s Indicator Values for British Plants; Institute of Terrestrial Ecology: Norwich, UK, 1999; p. 46. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Country, Region, Village | Abb. | Latitude | Longitude | Altitude (m a.s.l.) | n°mpl | n° | n°t cc/cw/wc/ww |
|---|---|---|---|---|---|---|---|---|
| Corylus avellana | Belgium, Flanders | Be_Cave | 50.947929 | 3.765214 | 24 | 10 | 92 | 26/19/18/29 |
| Spain, Pyrenees, Linas De Broto | Sp_Cave | 42.630049 | −0.169068 | 1270 | 9 | 80 | 26/14/18/22 | |
| Intermediate forms | Belgium, Flanders | Be_Cx | 50.965736 | 3.693461 | 10 | 7 | 61 | 17/13/13/18 |
| Corylus maxima | Belgium, Flanders | Be_Cmax | 50.992723 | 3.775170 | 19 | 2 | 20 | 6/4/4/6 |
| Cornus sanguinea | Belgium, Flanders, Kriephoek | Be | 50.953324 | 3.663467 | 10 | 21 | 205 | 61/42/44/58 |
| Spain, Pyrenees, Linas De Broto | Sp | 42.630049 | −0.169068 | 1270 | 12 | 117 | 34/24/24/35 |
| Genus | Year | Bud Burst | Leaf Senescence |
|---|---|---|---|
| Corylus | 2018 | 26/2, 1/3, 5/3, 8/3, 12/3, 15/3, 19/3, 26/3, 3/4, 9/4 | 28/9, 17/10 |
| 2019 | 6/3, 14/3, 4/4 | 16/9, 15/10, 19/11 | |
| 2020 | 9/3, 27/3, 8/4 | 14/9, 2/10, 23/10 | |
| Cornus | 2018 | 26/2, 1/3, 5/3, 8/3, 12/3, 15/3, 19/3, 26/3, 3/4, 9/4 | 28/9, 16/10 |
| 2019 | 5/4, 10/4, 17/4 | 12/9, 15/10, 19/11 | |
| 2020 | 9/3, 16/3, 27/3, 6/4 | 21/9, 12/10, 6/11 |
| Corylus Taxa | Cornus sanguinea | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Height Increment | Diameter Increment | Height Increment | Diameter Increment | |||||||
| Year | Variable | Estimate | p-Value | Estimate | p-Value | Variable | Estimate | p-Value | Estimate | p-Value |
| 2018 | (Intercept) | 12.41 | <0.001 *** | 2.20 | <0.001 *** | (Intercept) | 17.68 | <0.001 *** | 2.76 | <0.001 *** |
| Tcw | 3.84 | 0.004 ** | 0.23 | <0.001 *** | Tcw | 15.10 | <0.001 *** | 0.27 | <0.001 *** | |
| Twc | −8.78 | <0.001 *** | −0.32 | <0.001 *** | Twc | −1.04 | 0.487 | 0.03 | 0.698 | |
| Tww | −4.85 | <0.001 *** | −0.30 | <0.001 *** | Tww | 7.71 | <0.001 *** | 0.00 | 0.951 | |
| H17/D17 | 0.02 | 0.630 | −0.47 | <0.001 *** | H17/D17 | 0.28 | <0.001 *** | −0.54 | <0.001 *** | |
| Be_Cx | 2.55 | 0.096 | 0.34 | 0.025 * | Sp | 1.95 | 0.265 | −0.10 | 0.111 | |
| Be_Cmax | 2.85 | 0.204 | 0.53 | 0.026 * | ||||||
| Sp_Cave | −1.03 | 0.454 | −0.15 | 0.279 | ||||||
| 2019 | (Intercept) | 5.02 | <0.001 *** | 2.40 | <0.001 *** | (Intercept) | 4.18 | <0.001 *** | 2.78 | <0.001 *** |
| Tcw | −0.43 | 0.469 | −0.10 | 0.312 | Tcw | 0.01 | 0.979 | 0.43 | <0.001 *** | |
| Twc | −0.31 | 0.604 | 0.35 | 0.001 ** | Twc | −0.57 | 0.178 | 0.12 | 0.245 | |
| Tww | 0.23 | 0.661 | 0.34 | <0.001 *** | Tww | 0.31 | 0.437 | 0.17 | 0.073 | |
| H18/D18 | −0.05 | 0.007 ** | −0.34 | <0.001 *** | H18/D18 | −0.02 | 0.190 | −0.39 | <0.001 *** | |
| Be_Cx | 0.60 | 0.344 | 0.02 | 0.885 | Sp | −1.05 | 0.047 * | −0.13 | 0.143 | |
| Be_Cmax | 1.43 | 0.131 | 0.32 | 0.124 | ||||||
| Sp_Cave | −0.15 | 0.797 | −0.01 | 0.951 | ||||||
| 2020 | (Intercept) | 7.25 | <0.001 *** | 4.31 | <0.001 *** | (Intercept) | 59.3 | <0.001 *** | 4.78 | <0.001 *** |
| Tcw | −0.66 | 0.257 | −0.16 | 0.294 | Tcw | −1.07 | 0.721 | 0.36 | 0.048 * | |
| Twc | −0.12 | 0.832 | −0.22 | 0.135 | Twc | 1.05 | 0.694 | 0.05 | 0.771 | |
| Tww | 0.23 | 0.664 | −0.07 | 0.604 | Tww | −3.11 | 0.215 | 0.09 | 0.552 | |
| H19/D19 | −0.08 | <0.001 *** | −0.17 | 0.035 * | H19/D19 | 0.09 | 0.340 | −0.34 | <0.001 *** | |
| Be_Cx | 0.16 | 0.803 | 0.08 | 0.696 | Sp | −10.47 | <0.001 *** | −0.24 | 0.184 | |
| Be_Cmax | −0.36 | 0.716 | −0.08 | 0.796 | ||||||
| Sp_Cave | −0.81 | 0.184 | −0.25 | 0.169 | ||||||
| Corylus Taxa | Cornus sanguinea | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Bud Burst | Leaf Senescence | Bud Burst | Leaf Senescence | |||||||||
| Year | Variable | Estimate | p-Value | Variable | Estimate | p-Value | Variable | Estimate | p-Value | Variable | Estimate | p-Value |
| 2018 | D | −0.50 | <0.001 *** | D | 0.30 | <0.001 *** | D | −0.26 | <0.001 *** | D | 0.47 | <0.001 *** |
| Tw | 1.39 | <0.001 *** | Tcw | 0.25 | 0.540 | Tw | 1.55 | <0.001 *** | Tcw | −2.30 | 0.002 ** | |
| Twc | 0.88 | 0.040 * | Twc | 0.24 | 0.692 | |||||||
| Tww | 1.47 | <0.001 *** | Tww | −2.22 | 0.001 ** | |||||||
| D:Tw | −0.69 | <0.001 *** | D:Tcw | −0.11 | <0.001 *** | D:Tw | −0.27 | <0.001 *** | D:Tcw | −0.03 | 0.496 | |
| D:Twc | 0.05 | 0.069 | D:Twc | −0.01 | 0.768 | |||||||
| D:Tww | −0.06 | 0.018 * | D:Tww | −0.03 | 0.438 | |||||||
| H17 | 0.15 | <0.001 *** | H18 | 0.04 | <0.001 *** | H17 | −0.01 | 0.583 | H18 | 0.05 | 0.023 * | |
| Be_Cx | −1.45 | 0.135 | Be_Cx | −0.12 | 0.698 | Sp | −0.59 | 0.017 * | Sp | −0.68 | 0.168 | |
| Be_Cmax | −3.95 | 0.008 ** | Be_Cmax | 0.72 | 0.103 | |||||||
| Sp_Cave | 1.18 | 0.190 | Sp_Cave | 0.63 | 0.026 * | |||||||
| 2019 | D | −0.22 | <0.001 *** | D | 0.14 | <0.001 *** | D | −0.81 | <0.001 *** | D | 0.16 | <0.001 *** |
| Tcw | 1.55 | <0.001 *** | Tcw | 0.35 | 0.191 | Tcw | 1.20 | 0.001 ** | Tcw | 0.26 | 0.316 | |
| Twc | −0.86 | 0.004 ** | Twc | −0.18 | 0.519 | Twc | −0.27 | 0.405 | Twc | 0.41 | 0.077 | |
| Tww | 0.62 | 0.015 * | Tww | 0.30 | 0.227 | Tww | 1.05 | <0.001 *** | Tww | 0.01 | 0.949 | |
| H18 | 0.06 | <0.001 *** | H19 | 0.00 | 0.886 | H18 | −0.02 | 0.111 | H19 | 0.00 | 0.811 | |
| Be_Cx | −1.05 | 0.034 * | Be_Cx | 0.26 | 0.350 | Sp | −1.42 | <0.001 *** | Sp | 0.49 | 0.008 ** | |
| Be_Cmax | −2.83 | <0.001 *** | Be_Cmax | −0.02 | 0.953 | |||||||
| Sp_Cave | 0.29 | 0.522 | Sp_Cave | 0.12 | 0.658 | |||||||
| 2020 | D | −0.40 | <0.001 *** | D | 0.39 | <0.001 *** | D | −0.75 | <0.001 *** | D | 0.13 | <0.001 *** |
| Tcw | −0.57 | 0.174 | Tcw | −0.58 | 0.110 | Tcw | 0.24 | 0.622 | Tcw | −0.11 | 0.744 | |
| Twc | −0.03 | 0.941 | Twc | 0.34 | 0.344 | Twc | −0.37 | 0.388 | Twc | 0.29 | 0.373 | |
| Tww | 0.01 | 0.981 | Tww | −0.07 | 0.821 | Tww | −0.09 | 0.813 | Tww | 0.04 | 0.885 | |
| H19 | 0.04 | 0.001 ** | H20 | 0.05 | <0.001 *** | H19 | 0.007 | 0.652 | H20 | −0.006 | 0.382 | |
| Be_Cx | −1.18 | 0.020 * | Be_Cx | 0.85 | 0.010* | Sp | −2.08 | <0.001 *** | Sp | −0.47 | 0.093 | |
| Be_Cmax | −2.24 | 0.003 ** | Be_Cmax | 0.34 | 0.482 | |||||||
| Sp_Cave | 0.18 | 0.688 | Sp_Cave | −0.01 | 0.983 | |||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vander Mijnsbrugge, K.; Malanguis, J.M.; Moreels, S.; Turcsán, A.; Paino, E.N. Stimulation, Reduction and Compensation Growth, and Variable Phenological Responses to Spring and/or Summer–Autumn Warming in Corylus Taxa and Cornus sanguinea L. Forests 2022, 13, 654. https://0-doi-org.brum.beds.ac.uk/10.3390/f13050654
Vander Mijnsbrugge K, Malanguis JM, Moreels S, Turcsán A, Paino EN. Stimulation, Reduction and Compensation Growth, and Variable Phenological Responses to Spring and/or Summer–Autumn Warming in Corylus Taxa and Cornus sanguinea L. Forests. 2022; 13(5):654. https://0-doi-org.brum.beds.ac.uk/10.3390/f13050654
Chicago/Turabian StyleVander Mijnsbrugge, Kristine, Jessa May Malanguis, Stefaan Moreels, Arion Turcsán, and Eduardo Notivol Paino. 2022. "Stimulation, Reduction and Compensation Growth, and Variable Phenological Responses to Spring and/or Summer–Autumn Warming in Corylus Taxa and Cornus sanguinea L." Forests 13, no. 5: 654. https://0-doi-org.brum.beds.ac.uk/10.3390/f13050654