
Changes in Soil Ectomycorrhizal Fungi Community in Oak Forests along the Urban–Rural Gradient

1
School of Water Conservancy and Environment, University of Jinan, Jinan 250022, China
2
Office of Academic Research, Shandong Academy of Forestry Sciences, Jinan 250000, China
*
Author to whom correspondence should be addressed.
Forests 2022, 13(5), 675; https://0-doi-org.brum.beds.ac.uk/10.3390/f13050675
Submission received: 21 March 2022
/
Revised: 15 April 2022
/
Accepted: 25 April 2022
/
Published: 27 April 2022
(This article belongs to the Special Issue Plant Adaptation to Extreme Environments in Drylands)
Abstract
:The ectomycorrhizal fungi communities of forests are closely correlated with forest health and ecosystem functions. To investigate the structure and composition of ectomycorrhizal fungi communities in oak forest soil and their driving factors along the urban–rural gradient, we set up a Quercus acutissima forest transect and collected samples from the center to the edge of Jinan city (urban, suburban, rural). The results showed that the ectomycorrhizal fungal community composition at the phyla level mainly included Basidiomycota and Ascomycota in three sites. At the genus level, the community compositions of ectomycorrhizal fungi, along the urban–rural gradient, exhibited significant differences. Inocybe, Russula, Scleroderma, Tomentella, Amanita and Tuber were the dominant genera in these Quercus acutissima forests. Additionally, the diversity of ectomycorrhizal fungi was the highest in rural Quercus acutissima forest, followed by urban and suburban areas. Key ectomycorrhizal fungi species, such as Tuber, Russula and Sordariales, were identified among three forests. We also found that pH, soil organic matter and ammonium nitrogen were the main driving factors of the differences in ectomycorrhizal fungi community composition and diversity along the urban–rural gradient. Overall, the differences in composition and diversity in urban–rural gradient forest were driven by the differences in soil physicochemical properties resulting from the forest location.
1. Introduction
Ectomycorrhizal (ECM) fungi are one of the taxa of fungi that are among the most abundant and widespread in forests [1]. There are about 20,000~25,000 ECM fungi in the world, and they can associate with 6000 species of trees and shrubs mainly distributed in tropical and temperate forests [2,3,4]. In particular, the tree species from Fagaceae, Betulaceae, Pinaceae and Legumes are the dominant symbiotic plants of ECM fungi [1]. These plants provide carbon synthesized by photosynthesis for the fungi, while ECM fungi supply nutrients to the host plants, such as nitrogen and phosphorus [5,6]. Moreover, ECM fungi play critical roles in global carbon cycles by secreting extracellular enzymes and organic acids to facilitate the decomposition of soil organic matter [7]. In addition, previous studies have confirmed that ECM fungi play a significant role in the enhancement of plant tolerance to adverse environments [8] and the sustainment of forest ecosystem diversity and functions [1,9]. Generally, ECM diversity and community structure are considered to be the important indicators to evaluate the health and stability of forest ecosystems [10]. In this sense, studies on ECM fungi diversity and its driving factors can provide us with a better understanding of ecological processes such as geochemical element cycling, plant nutrient acquisition, sustainable forest development and global environmental change [1,7].
The diversity and richness of ECM fungi are affected by different soil conditions [11,12]. It has been found that higher levels of organic matter and more phosphorus available in soil usually inhibit ECM fungi occurrence [13,14]. Lower pH has been found to be conducive to increases in ECM diversity and richness [15]. ECM communities are significantly different in the cold temperate zone, warm temperate zone and Mediterranean climate zone, indicating that climatic conditions are important driving factors for ECM composition and structure [11]. Furthermore, on the basis of their specificity to host plants, some ECM fungi are able to associate with specific tree species, which is another important factor affecting ECM fungi structure and diversity in different forest ecosystems [1]. Therefore, understanding the influence of environmental factors on the ECM community is critical for the exploitation and protection of ECM fungi function in forest ecosystems.
Oak (Quercus Spp.) is one of the dominant deciduous tree genera in boreal forests around the world, and it has great ecological and economic value for environmental protection, seedling establishment and wood yield [16]. Its roots rely on obligate symbiosis with ECM fungi. It cannot survive and establish itself without ECM fungi [17]. Oak roots can improve nutrient absorption and plant adaptability once they are associated with ECM fungi, especially in adverse conditions [16,17]. Thus, the composition and diversity of ECM fungi in oak forests have been paid much attention in previous studies. He et al. [18] observed that the compositions of ECM fungal communities varied with the stand ages of oak forests, but their diversities were broadly similar across the 20-, 30- and 40-year-old Quercus Mongolica forests. Furthermore, the ECM fungal community structure of oak forest soil was obviously different in different habitats. The soil fungal biodiversity had a great impact on hosts, especially at the nursery stage, where it could influence the quality of the plating material [19]. Jin et al. [20] found that the ECM fungi in oak nurseries were mainly composed of Boletales while Agaricales dominated in the Guizhou wild oak forest. Moreover, previous studies have shown that ECM fungal diversity and richness in oak forests significantly change with seasonal shifts and that they generally present the highest abundance in summer [20,21]. Although the changes in ECM fungal communities have been extensively studied, there is little information on the composition of ECM fungal communities and its driving factors in Quercus acutissima forests along the urban–rural gradient. As an effective ecological research method, the rural–urban gradient method has been widely used to study ecological problems in various regions of the world [22,23]. Here, we investigated the ECM community in Quercus acutissima forests along the urban–rural gradient in Jinan city, northern China. The aims were to: (1) explore the changes in ECM community composition and diversity along the urban–rural gradient and (2) reveal the effects of soil physicochemical properties on ECM community compositions and diversity. Our results provide practical guidance for urban and suburban forest ecosystem management.
2. Materials and Methods
2.1. Sample Site
Jinan city, the capital of Shandong Province, is located between 34°46′–37°32′ N and 116°13′–117°58′ E, in the middle and lower reaches of the Yellow River. The terrain in the south is higher than that in the north in this area. Jinan has a typical temperate monsoon climate characterized by distinct seasons, a mean annual temperature of 13.8 °C and mean precipitation of 650–700 mm. Jinan city has abundant forest resources, dominated by Quercus acutissima, Platycladus orientalis, Robinia pseudoacacia and Pinus thunbergii Parl [24]. The urbanization level in Jinan reached 73.46% as of 2021 [25]. The typical urban areas, ecologically sensitive areas and southern mountain control areas were designed in Jinan along with the development of the economy. Here, we focused on the Quercus acutissima forest, which is a typical type of vegetation in Jinan. The study areas were Quancheng Park (36°38′ N, 117°38′ E) in the urban area, Liubu Forest Farm (36°27′ N, 117°11′ E) in the suburbs and Yaoxiang Forest Park (36°12′ N, 117°4′ E) in the countryside. The geographic information for the study areas is shown in Figure 1.
2.2. Sample Collection and Treatment Process
In October 2020, three independent sample plots (20 m × 20 m each; the distances were more than 1000 m from each other) were set up in the urban, suburban and rural Quercus acutissima forests of Jinan city, respectively. Five healthy trees with similar tree heights and diameters at breast height were randomly selected in each plot. The distances between the trees were more than 10 m. Humus such as dead branches and leaves was removed from the soil surface, and then four soil cores were collected from the east, west, south and north directions around each tree with 3.5 cm diameter soil augers at 20 cm depths. All the soil samples were evenly combined into one in each plot and a total of nine composite soil samples were thus obtained from these urban, suburban and rural Quercus acutissima forests (three replicated samples per forest × three forests). Afterward, the collected samples were stored in cryogenic ice boxes and quickly transported back to the laboratory. The fresh soil samples were sieved through a 2 mm sieve after roots and residues were removed. After being mixed evenly, samples were divided into two parts: one was stored at 4 °C for the determination of soil physicochemical properties and the other was frozen at −80 °C until molecular testing of ECM fungi.
2.3. Soil Physicochemical Properties Determination
The soil pH (water: soil = 1: 2.5) was measured using an UltraBasic pH meter (Denver Instruments, UB-10, Bohemia, NY, USA) and conductivity (EC) (water: soil = 1:5) was measured using a portable conductivity meter. Microbial biomass carbon (MBC) and microbial biomass nitrogen (MBN) in the soil samples were determined by chloroform fumigation. [26]. Soil organic matter (SOM) was determined using the H2SO4-K2CrO7 oxidation method [27]. Total nitrogen (TN) was analyzed using an automatic Kjeldahl apparatus (KDY-9820, Tongrunyuan, Beijing, China) [27]. Soil ammonium nitrogen (NH4+-N) and nitrate nitrogen (NO3−-N) in the KCl extraction were measured using a UV-visible spectrophotometer (UV-5200PC, Yuanxi, Shanghai, China) [28]. The ratio of C/N in the soil was calculated based on soil organic carbon and total nitrogen contents. Soil available phosphorus (AP) and available potassium (AK) were determined with atomic absorption spectrometers (SHIMADIV AA-7000, Shimadzu, Tokyo, Japan) after being extracted with 0.5 mol/L NaHCO3 and 1 mol/L CH3COONH4 (pH = 7), respectively [29].
2.4. Soil DNA Extraction, PCR Amplifification and Sequencing
DNA extraction, PCR amplification and high-throughput sequencing of the fungal ITS sequences of the soil samples were completed by Hangzhou LC-Bio Technology Co., Ltd. DNA extraction was performed using an OMEGA Soil DNA Kit (OMEGA, Norcross, GA, USA) according to the manufacturer’s instructions. The ITS1 and ITS2 regions were used to identify the fungal species, and the analysis was carried out with ITS1FI2 (5′-GTGARTCATCGAATCTTTG-3′), ITS2 (5′-TCCTCCGCTTATTGATATGC-3′), F (5′-GAACCWGCGGARGGATCA-3′) and R (5′-GCTGCGTTCTTCATCGATGC-3′) primers. PCR amplification was performed in a reaction mixture with a total volume of 25 μL containing 25 ng of template DNA, 12.5 μL PCR Premix, 2.5 μL of each primer and PCR-grade water to adjust the volume. The PCR conditions to amplify the ITS fragments consisted of an initial denaturation at 98 °C for 30 s; 32 cycles of denaturation at 98 °C for 10 s, annealing at 54 °C for 30 s and extension at 72 °C for 45 s; and then final extension at 72 °C for 10 min. The PCR products were confirmed with 2% agarose gel electrophoresis. The PCR products were purified with AMPure XT beads (Beckman Coulter Genomics, Danvers, MA, USA) and quantified with Qubit (Invitrogen, Carlsbad, CA, USA). The amplicon pools were prepared for sequencing and the size and quantity of the amplicon library were assessed with an Agilent 2100 Bioanalyzer (Agilent, Palo Alto, CA, USA) and the Library Quantification Kit for Illumina (Kapa Biosciences, Woburn, MA, USA), respectively. The libraries were sequenced on a NovaSeq PE250 platform. Samples were sequenced on an Illumina NovaSeq platform according to the manufacturer’s recommendations, provided by LC-Bio. QIIME software was used to divide the OTU of Illumina Miseq sequencing data. The RDP database and UNITE database (https://unite.ut.ee/ (accessed on 1 March 2021)) were applied to classify and annotate ECM fungi, and Mothur software was used to calculate the Chao1 index, Shannon index and Simpson index.
2.5. Data Analysis
IBM SPSS Statistics 23.0 (SPSS Inc., Chicago, IL, USA) was used for one-way analysis of variance (ANOVA) and multiple comparison to evaluate the significant differences in the different forest soils (p < 0.05). Origin 2021 (OriginLab Inc., Northampton, MA, USA) was used to draw a histogram of community relative abundance at the order and genus levels (others < 0.01). Redundancy analysis (RDA) was performed with CANOCO 5.0 to examine the habitat differences in key ECM fungi species. We utilized linear discriminant analysis effect size (LEfSe) on the website of LC-Bio Technology Co., Ltd., to identify the ECM fungi that might explain the differences among the three forests along the urban–rural gradient (LDA > 4, p < 0.05). In addition, Pearson’s correlation analysis was applied with Origin 2021 to examine the relative effects of ECM fungi characteristics (diversity and richness) and soil factors (pH, SOM, TN, NH4+-N, NO3−-N, AP, AK, EC).
3. Results
3.1. Differences in Soil Physicochemical Properties
There were significant differences in the soil physicochemical properties among the three sampling areas along the urban–rural gradient in Jinan (Table 1). Specifically, the pH value and the contents of NH4+-N, AK and AP in the urban forest soil were significantly higher than those in the suburban and rural forests (p < 0.05), whereas TN and NO3−-N in the urban forest were clearly lower than those in suburban and rural forests, and the difference between suburban and rural forests was not significant. In addition, the content of SOM in the suburban forest soil was 2.50 and 2.19 times higher than that in urban and rural forests, respectively. No significant differences in C/N among any of the sampling sites were observed.
3.2. MBC and MBN Contents in the Quercus acutissima Forest Soil
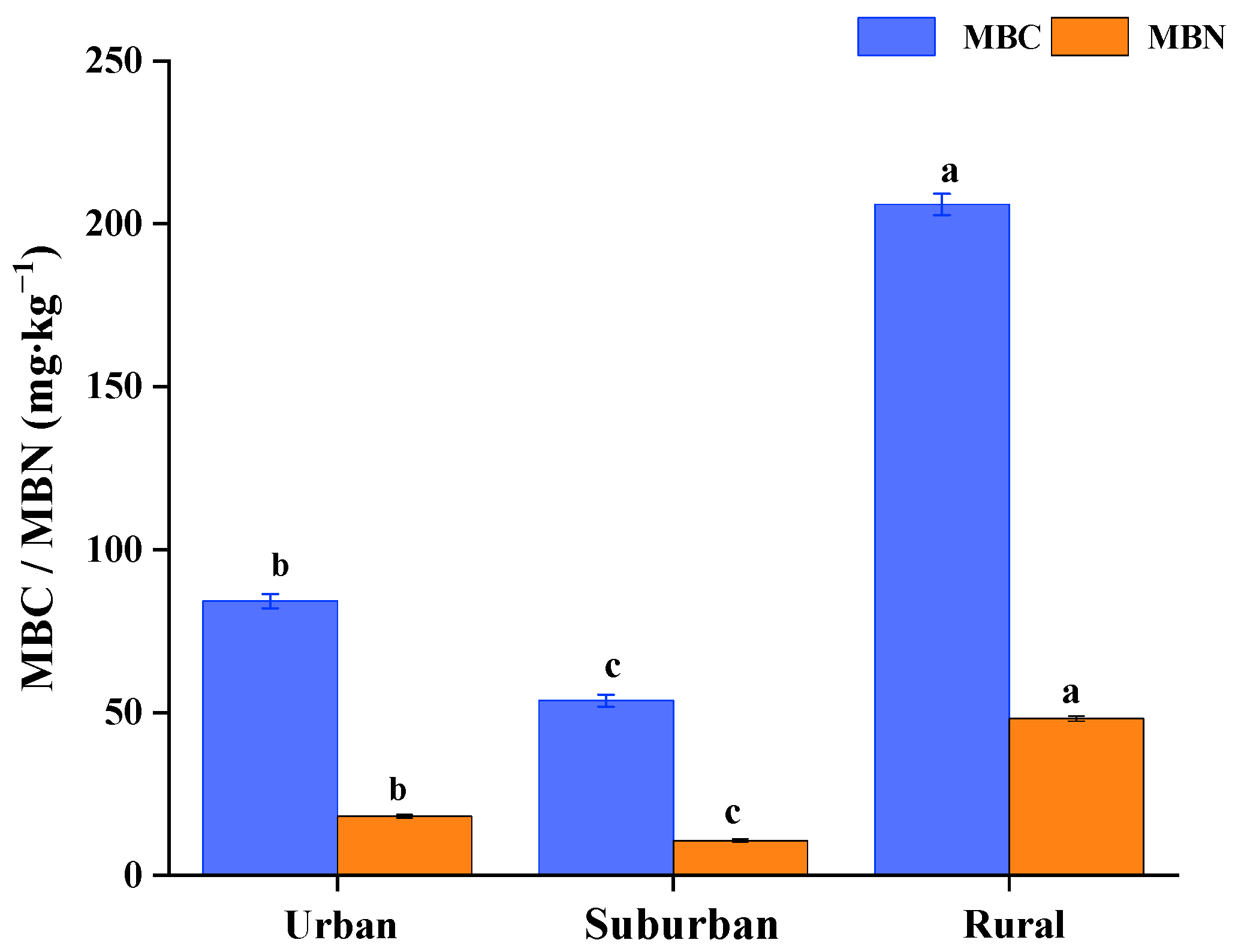
Both the MBC and MBN in the forest soil exhibited significant differences in Quercus acutissima forests along the urban–rural gradient (Figure 2). Generally, the average content of MBC in urban, suburban and rural soil was 84.21, 53.68 and 205.96 mg∙kg−1, respectively, showing a descending order of rural > urban > suburban. With the same trend as the MBC, the average content of MBN in urban, suburban and rural forest soil was 18.18, 10.78 and 48.15 mg∙kg−1, respectively.
3.3. The Change in ECM Fungi Diversity
All the Good’s coverage estimators were greater than 99%, which suggested that there was an overall good sampling (Table 2). The ECM fungal community diversity in rural forest soil was the highest, with a Shannon index of 2.76 and a Simpson index of 0.92, followed by urban forest (2.54 and 0.89) and suburban forest (2.12 and 0.83). However, Chao1 values were broadly similar in the forest soil of urban, suburban and rural forests, which tat indicated habitat changes had little influence on the ECM fungi richness along the urban–rural gradient. This also showed that the effects of the urban–rural gradient on ECM fungi diversity and abundance were significantly different.
3.4. ECM Fungal Community Compositions Vary with Urban–Rural Gradient
A total of 282 ECM fungal OTUs were retrieved from the ITS sequence of soil samples collected from Quercus acutissima forests in Jinan along the urban–rural gradient. These ECM fungi were distributed in 3 phyla, 5 classes, 11 orders and 26 genera. At the phyla level, the most dominant were Basidiomycota (81.11%), followed by Ascomycota (13.83%) and Mucoromycota (1.06%). Agaricomycetes were the most dominant guild at the class level, accounting for up to 86.53%, 99.94% and 72.69% of the total community in urban, suburban and rural sampling sites, respectively. Sordariomycetes were the second class, accounting for 0.56%, 0.12% and 27.57% in urban, suburban and rural forests, respectively. Pezizomycetes, Eurotiomycetes and Endogonomycetes were occasionally detected, implying they were the minor classes.
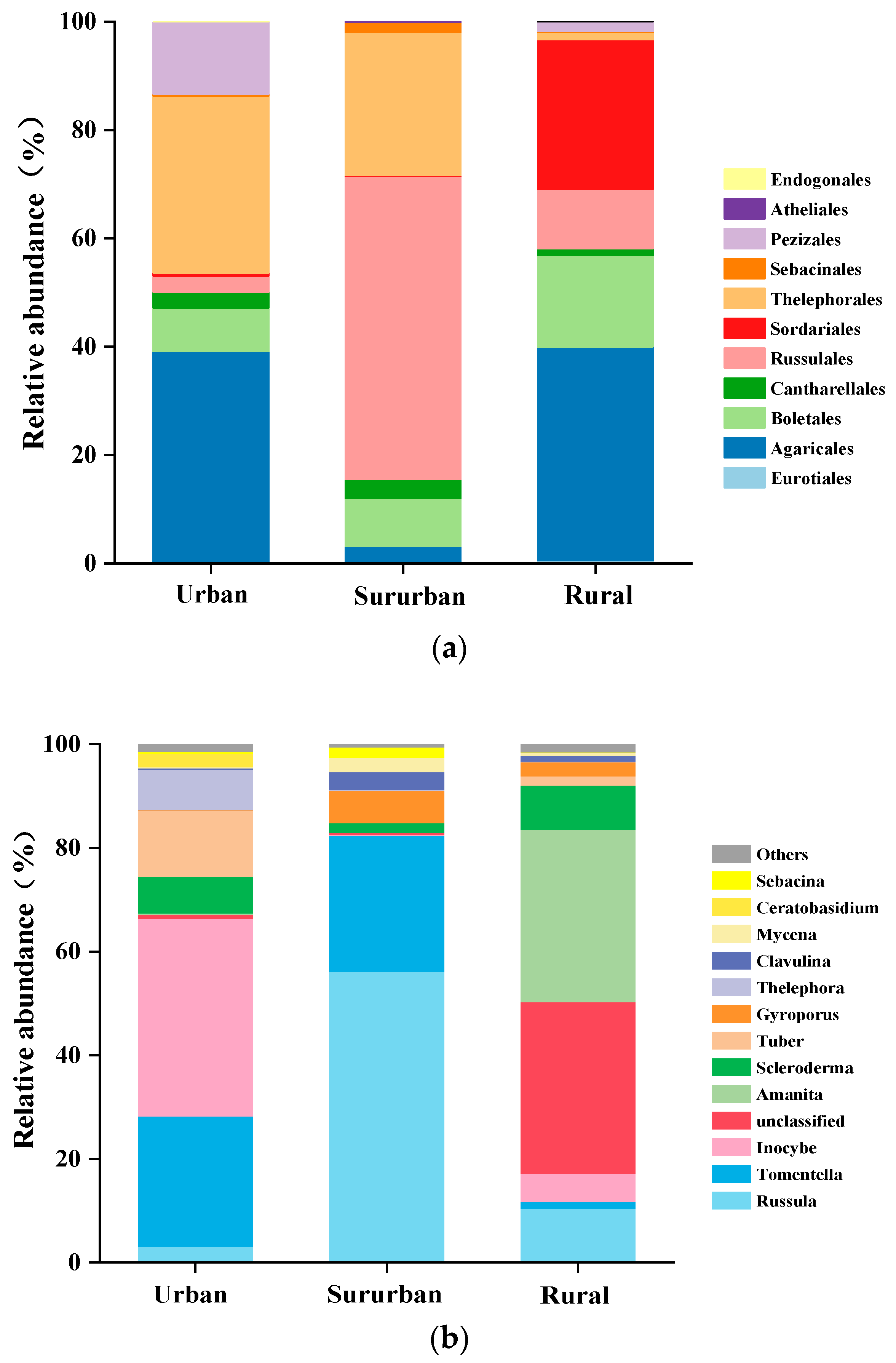
Major differences were observed in ECM fungal community compositions along the urban–rural gradient (Figure 3). At the order level, Agaricales had the highest relative abundance in urban and rural Quercus acutissima forests (Figure 3a), which indicates that it can survive in broad habitats and plays crucial roles in sustaining ecosystem balance [30]. Russulales were extensively found in these Quercus acutissima forests along the urban–rural gradient, especially in suburban areas where the highest proportion was 56.02%, implying that Russulales may play an important role in the growth and health of oak trees in the sensitive areas disturbed by human activities. The top 13 genera of ECM fungi in Quercus acutissima forest soil are shown in Figure 3b. Inocybe was the dominant genus in the urban site and its relative abundance was 37.87%, followed by 5.49% for the rural site and 2.84% for the suburban site. However, the relative abundance of Russula was highest in the suburban area but lowest in the urban area. High proportions of Amanita were observed in the soil samples from the rural area, whereas Amanita was almost absent in urban and suburban areas. Tomentella was also one of the representative genera in the soil of Quercus acutissima forest. The order of its relative abundance in the study sites along the urban–rural gradient was suburban > urban > rural. Overall, the dominant ECM fungi in Quercus acutissima forest soil showed remarkable differences along the urban–rural gradient.
3.5. LEfSe Analysis of ECM Fungi Community
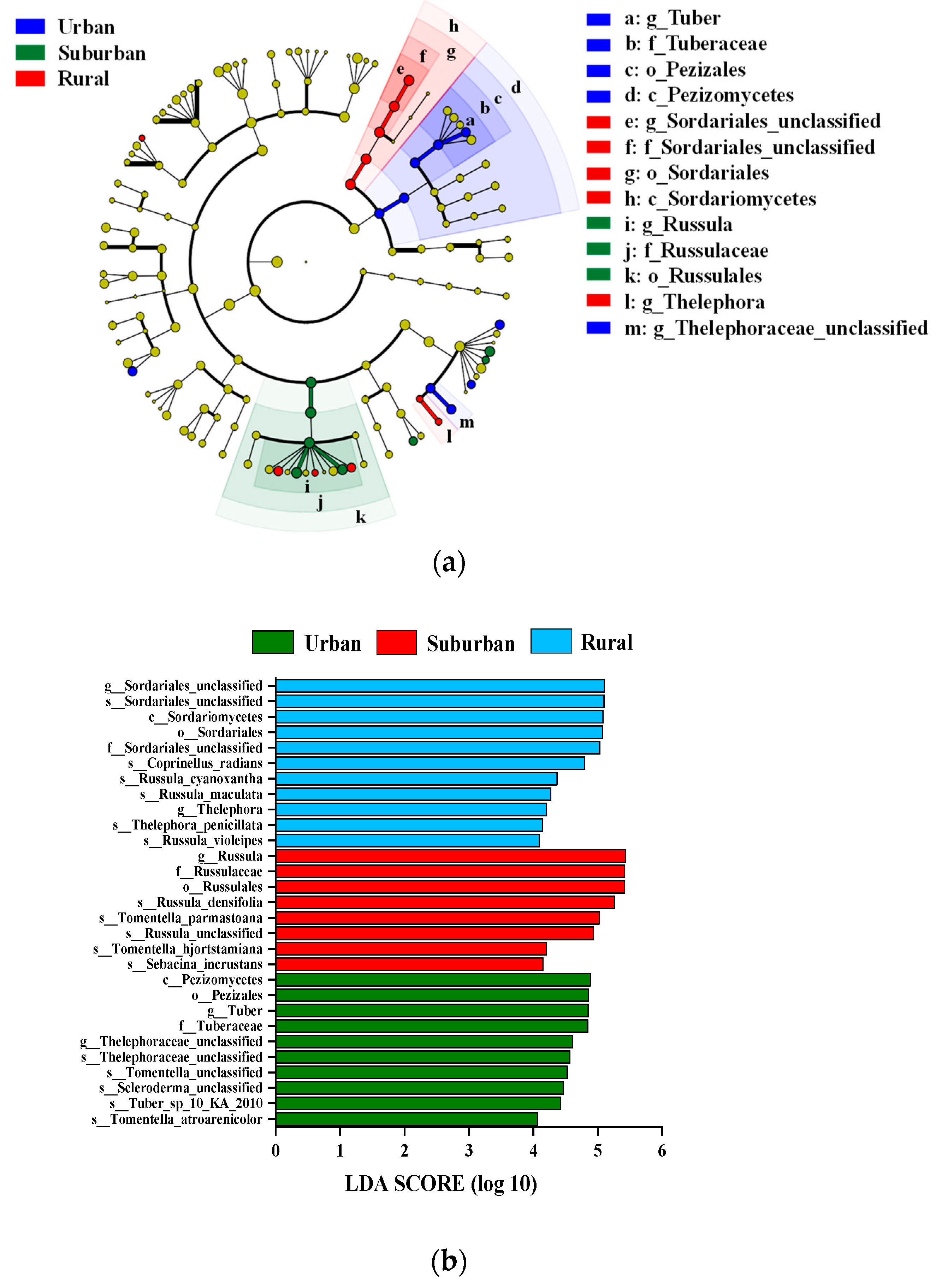
As shown in Figure 4, remarkable differences in key ECM fungi taxa were observed among the urban, suburban and rural Quercus acutissima forest soils. Specifically, Tuber and an unclassified genus from Thelephoraceae were two distinct ECM fungi members explaining habitat variations that showed significant differences in relative abundance in the urban soil. Meanwhile, Russula was the taxon of ECM fungi present in the suburban soil. In addition, there were two characteristic groups of ECM fungi with significant differences in relative abundance in the rural site compared to the urban and suburban sites. They were an unclassified genus from Sordariales and an unclassified genus from Thelephoraceae, respectively. The LDA value also further indicated that Tuber, Russula and Sordariales were the key groups in responses to more pronounced habitat shifts in the urban, suburban and rural Quercus acutissima forest soils, respectively.
3.6. Driving Factors of ECM Fungal Composition
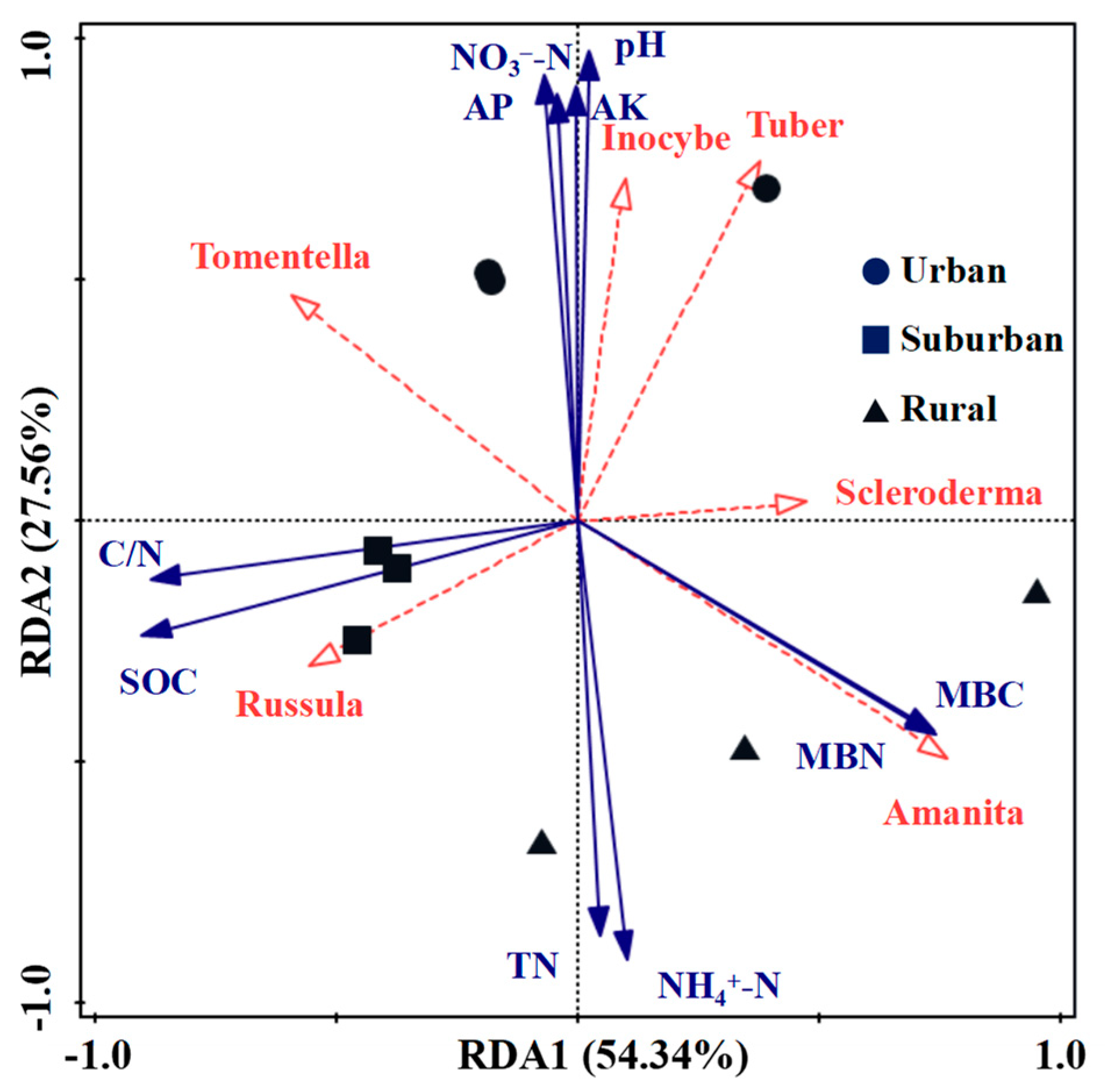
Redundancy analysis confirmed that soil physicochemical properties had the greatest effect on dominant ECM fungi at the genus level (top six relative abundances) (Figure 5). The first axis and second axis explain the variations of 54.34% and 27.56%, respectively. The dominant Russula exhibited a strongly positive relationship with SOC and C/N, while Scleroderma was negatively correlated with SOC and C/N, indicating that soil carbon could drive much more variation in Russula and Scleroderma. Inocybe and Tuber showed a remarkably positive relationship with NO3−-N, AK, AP and pH, but a strongly negative relationship with NH4+-N and TN. In addition, Tomentella weakly positively correlated with SOC, NO3−-N, AK and AP. Amanita significantly positively correlated with MBC and MBN, but weakly related with NH4+-N and TN. The three sampling sites for urban, suburban and rural forests were distributed in different quadrants, and the three sampling plots in the same area showed obvious clustering phenomena, implying that the ECM fungal community composition differed significantly in the Quercus acutissima forests along the urban–rural gradient (Figure 3).
3.7. Relationships of ECM Fungal Diversity with Soil Factors
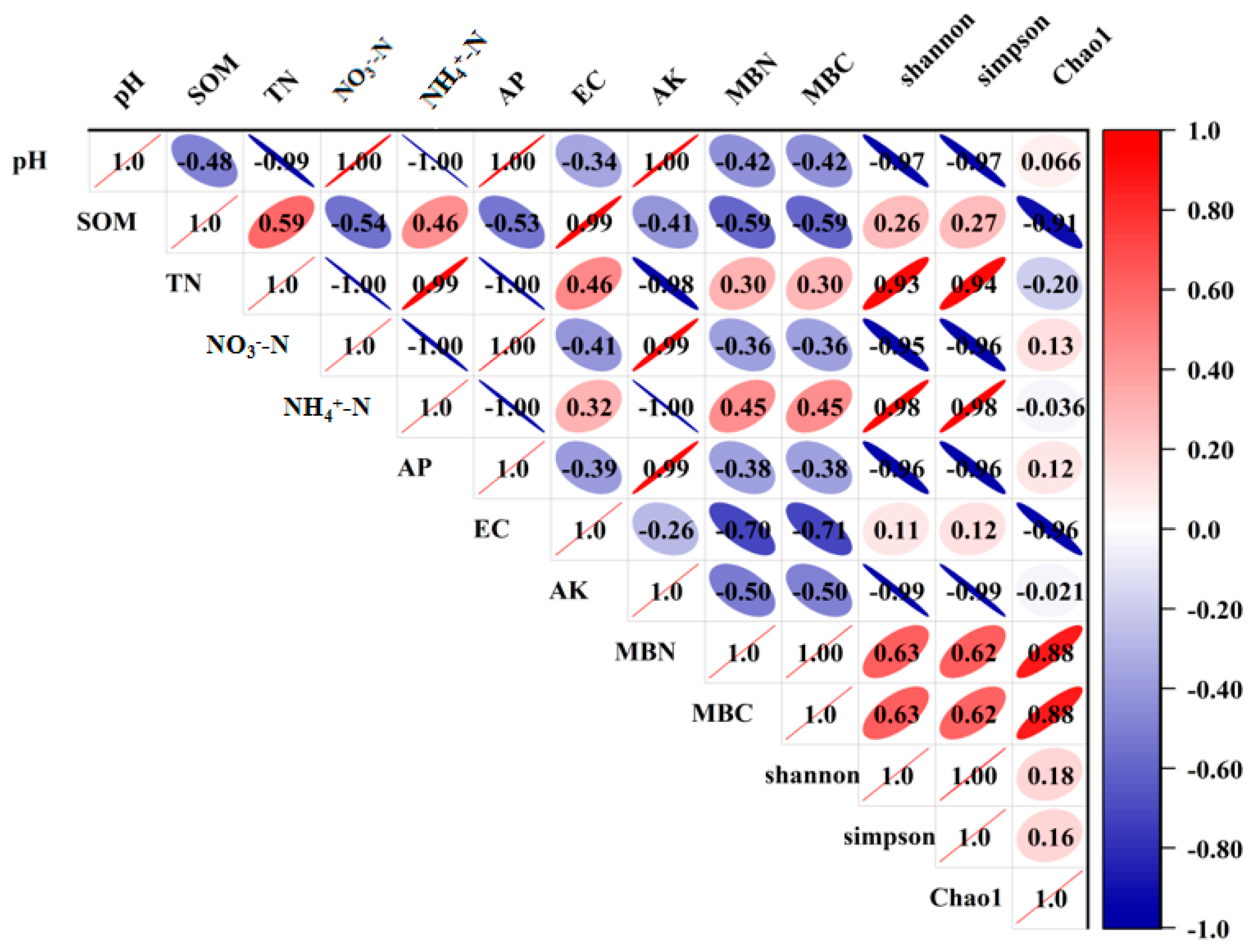
Among all the soil factors examined (Figure 6), the diversity indices (Shannon index and Simpson index) and richness index (Chao1 index) were strongly positively related to TN, NH4+-N, MBC and MBN, but negatively related to NO3−-N, AP and AK, which was consistent with the research conclusions of Erlandson et al. [11] and He et al. [18]. Moreover, NH4+-N was negatively correlated with AP and AK, while NO3−-N was positively correlated with AP and AK. It is worth noting that SOM and TN showed positive relationships with ECM fungal diversity, but negative relationships with ECM fungal richness, suggesting that the higher SOM and TN contents in soil were probably associated with the higher ECM fungi diversity.
4. Discussion
4.1. Diversity and Composition of ECM Fungal Community in the Quercus acutissima Forest Soil along the Urban–Rural Gradient
ECM fungi are one of the most important functional phyla in the forest soil ecosystem [31,32,33]. Quercus acutissima is a typical tree species associated with ECM fungi and can form a huge mycelium network to participate in the nutrient cycle and energy metabolism of the host [21]. ECM fungal community composition and diversity are commonly used to evaluate the stability of forest ecosystems because their changes are strongly related to soil health [10,34]. In this study, ECM fungal diversity exhibited more pronounced shifts along the urban–rural gradient. It was obvious that there were minimum levels of MBC and MBN in the suburban forest soil, indicating that microorganism growth was significantly inhibited in this ecotone. The contents of TN, NH4+-N and SOM were higher in the soil of the suburban Quercus acutissima forest than in urban and rural forests, whereas the Shannon index, Simpson index and MBC and MBN contents of ECM fungi were the lowest, implying that the ECM fungi were not apt to occur in more fertile soil [35]. This finding could have been related to the higher contents of carbon and nitrogen in the suburban forest soil, where ECM fungi were not necessary for the host to obtain more nutrients. Moreover, the decrease in ECM fungi diversity also resulted in a weaker decomposition effect on the soil organic matter or litter [25,26]. Additionally, the ECM fungal diversity and soil microbial biomass in the urban Quercus acutissima forest were significantly lower than in rural forest, which implied that human activities in urban areas likely had a negative influence on ECM fungal diversity and even on other kinds of microbial diversity [36]. Previous studies have also shown that lower abundance and diversity of soil microorganisms can result in the limitation of community function. Thus, conservation of ECM diversity is of great significance for maintaining the function of urban forest ecosystems [1].
Key species are generally critical indicators of environment conditions and community functions [36]. This study of ECM fungal community composition in forests could provide a basis for the exploration of the ectomycorrhizal symbiotic mechanism. Earlier studies of ECM fungal communities were only based on morphological and anatomical observations [1,37]. In recent years, use of molecular biology methods has provided new insights into the below-ground ECM fungal community and a more precise approach to the study of ECM fungal diversity [1]. Here, the ECM fungal community compositions in Quercus acutisana forest soil were investigated in different habitats (urban, suburban and rural forests) in Jinan city. Some genera, such as Inocybe, Russula and Tomentella, were found to be dominant, which was similar to the results reported in other oak forests [18]. It was confirmed that these taxa may have important ecological significance in terms of improving the environmental adaptability of trees due to their high genetic diversity and probable functional diversity [38]. Inocybe, Russula, Tomentella, Amanita and Scleroderma belonging to Basidiomycetes and Tuber from Ascomycetes were identified as the key ECM fungi taxa. They were closely associated with the community function; thus, changes in these ECM fungi’s relative abundance could potentially alter forest structure and function [20]. Interestingly, Inocybe frequently demonstrated a relatively high abundance in urban forest soil, but it was also occasionally observed in suburban and rural forests soil. Similarly, Jin [20] also found that the relative abundance of Inocybe in artificial forests was significantly higher in comparison to that growing in natural environment. Our result may be attributable to the presence of simple tree species in urban artificial forests, the conditions of which are favorable for Inocybe colonization. Russula is a widely distributed ECM fungi genus that can associate and be symbiotic with Quercus, Pine, Abies and Spruce throughout the world, especially in boreal temperate forests [39]. In our study, Russula showed a high relative abundance in the Quercus acutissima forest ecosystem along the urban–rural gradient, notably accounting for up to 56.01% in the suburbs. Amanita was the most abundant ECM fungi in the rural Quercus acutissima forest. Previous studies have confirmed that both Russula and Amanita are late successional species [18,39,40]. In the current study, the suburban and rural Quercus acutisana forests were older than the urban forest, which might have been the reason why the relative abundances of Russula and Amanita were higher in suburban and rural forest ecosystems. Yang et al. [41] found that Scleroderma and Tomentella had a strong Cd tolerance under the stress of Cd and benomyl. Both were also prevalent in this study, indicating that they play an essential role in resisting environmental stress, as well as maintaining plant growth and metabolism. In addition, some ECM fungi with lower abundances, such as Elaphomyces, Helvella, Boletus, Amphinema and Hydnum, were only observed in the rural Quercus acutissima forest, while Pisolithus was occasionally identified in the suburban forest. The functions of these rare or specific ECM fungi in different habitats need to be further studied.
4.2. Influencing Factors of ECM Fungal Community Composition in Quercus acutissima Forests along the Urban–Rural Gradient
The changes in ECM fungal community composition and diversity are often used to predict ecosystem health [1,20]. Here, there was an obvious correlation between soil physicochemical properties and ECM fungal community structure in the Quercus acutissima forests along the urban–rural gradient. Soil physicochemical properties can affect the growth and distribution of EMC fungi, thereby resulting in different EMC fungal communities [1,42]. In particular, pH exerts an important role in soil nutrient transformation and cycling by influencing the microbial community composition and activity [43]. It has been reported that most EMC fungi can survive in weakly acidic conditions, while their growth is inhibited in alkaline conditions [21]. Studies have found that the seedling roots associated with mycorrhizal fungi not only perform normal respiration to produce CO2, but also secrete organic acids and H+, leading to soil acidification in forests [44,45]. In this study, the soil pH ranged from 4.50 to 5.00 in the suburban and rural forests, while it was about 7.80 in the urban forest. The ECM fungal Shannon and Simpson indices in the rural forest were higher than those in the urban forest, suggesting that soil pH was significantly negatively correlated with diversity indices. This conclusion is consistent with the previous results obtained by Kyaschenko, who also stated that the presence of a large number of ECM fungi likely reduces soil pH values and that the diversity of a fungal community decreases as the pH values increase [46].
SOM is one of the main sources of metabolic substances and energy for the colonization of mycorrhizal fungal [47]. In turn, the interaction between fungal communities can also regulate the SOM accumulation and soil fertility [47,48]. Remarkably higher SOM content was detected in the suburban Quercus acutissima forest soil in Jinan. A possible reason was that the proportion of other mixed tree species was larger in the Quercus acutissima forests, which probably enhanced the input of organic matter from the aboveground litter layer into the soil and provided sufficient sources during SOM retention. In contrast, the Quercus acutissima forest in the urban area was strongly disturbed by human activities, which resulted in a low level of litter input and affected the formation and accumulation of soil SOM. Nitrogen is widely recognized as a key limiting nutrition in controlling species composition and diversity in forest ecosystems [49]. The soil nitrate nitrogen content in the Quercus acutissima forest was significantly higher than that of ammonium nitrogen. The NO3−-N has a negative charge and the adsorption with soil is weak [50]. Thus, NO3−-N is likely to be leached from soil, which is not conducive to fertilizer conservation [50]. However, the content of NH4+-N was more than twice that of NO3−-N in the suburban and rural Quercus acutissima forest soils, which could have effectively prevented soil nitrogen loss and maintained soil fertility [50]. Hao et al. [51] found that most EMC fungi have a preference for NH4+-N in the growth process. Similar results were found in this study: ECM fungal diversity was significantly positively correlated with NH4+-N and negatively correlated with NO3−-N, indicating that most ECM fungi also preferred NH4+-N in the Quercus acutissima forest soil. However, a number of studies have reported that excessive nitrogen deposition (mainly nitrate) also inhibits the growth of EMC fungi in temperate broad-leaved oak forests and leads to a decrease in their diversity [52]. Moreover, EMC fungi have different preferences and adaptability in different soils [34]. In the present study, there were different correlations between soil properties and key ECM fungi taxa. For example, Russula and Scleroderma were extremely sensitive to SOM content and Russula was suitable for growing in suburban forests with high SOM content. However, Scleroderma preferred to colonize in urban and rural forests with relatively low SOM content. It also had a wide adaptive capacity in the environment and its abundance was not affected by the contents of nitrogen, phosphorus and potassium in soil. Scleroderma was dominant in low-nutrient-level soil, which means that this genus may have a unique function in nutrient uptake. In comparison with Scleroderma, Inocybe is generally considered as a later successional ECM fungus [53], and it showed a very significant positive correlation with AP, AK and NO3−-N contents in the soil, suggesting that the growth of Inocybe requires abundant nutrient supply and a stable ecosystem. These results indicate that the occurrence of ECM fungi in soil is affected by different soil physicochemical properties, which may be related to ECM fungal species and their ecological adaptability [31]. In turn, ECM fungi also greatly alter the chemical form and availability of soil nutrients by secreting functional substances, thus playing a regulatory role in the host rhizosphere environment [20]. Hence, ECM fungi in forests along the urban–rural gradient strongly change with the various soil properties. The effects of soil physicochemical properties on soil health and ECM fungi should be considered in the cultivation of Quercus acutissima forests. As far as ECM fungi protection is concerned, it is necessary to detect in a timely manner the changes in soil physicochemical properties in forests. In order to obtain detailed information on ECM fungi along the urban–rural gradient, further studies need to be implemented including focuses on sampling sites, frequency and the driving force of human activities.
5. Conclusions
Soil ECM fungi are fundamental components of forest ecosystems. The three Quercus acutissima forests demonstrated significant shifts in ECM fungal communities and diversity in different habitats, suggesting that different forest management strategies may be needed for microbial biodiversity conservation along the urban–rural gradient. Moreover, the distribution of ECM fungi in the oak forest soil implies that the dominance of ECM fungi differs depending on the spatial location. Our results provide basic information on forest protection and management along the urban–rural gradient in Jinan city. More abundant information about the location-specific relationship between environmental factors and microbial community is urgently needed for effective forest soil management.
Author Contributions
Conceptualization, H.S. and W.S.; methodology, H.S.; software, H.S. and K.J.; validation, B.Y., G.Q. and H.W.; formal analysis, H.S.; investigation, H.S. and B.Y.; resources, H.S.; data curation, H.S.; writing—original draft preparation, H.S.; writing—review and editing, H.S.; visualization, H.S.; supervision, H.W.; project administration, H.W.; funding acquisition, H.W. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Natural Science Foundation of China (41877424; 31870606), the Research Leader Studio Project (2021GXRC094), the Key R & D project of Shandong Province (2021LZGC005-02-02) and the Fundamental Research Funds for the Central Universities, CHD (300102351505).
Acknowledgments
The authors acknowledge Hangzhou LC-Bio Technology Co., Ltd. (Hangzhou, China), for providing the sequencing platform and technical support.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Kumar, J.; Atri, N.S. Studies on ectomycorrhiza: An appraisal. Bot. Rev. 2018, 84, 108–155. [Google Scholar] [CrossRef]
- Roy-Bolduc, A.; Laliberté, E.; Hijri, M. High richness of ectomycorrhizal fungi and low host specificity in costal sand dune ecosystem revealed by network analysis. Ecol. Evol. 2016, 6, 349–362. [Google Scholar] [CrossRef] [PubMed]
- Brundrett, M.C. Mycorrhizal associations and other means of nutrition of vascular plants: Understanding the global diversity of host plants by resolving conflicting information and developing reliable means of diagnosis. Plant Soil 2009, 320, 37–77. [Google Scholar] [CrossRef]
- Rinaldi, A.C.; Comadini, O.; Kuyper, T.W. Ectomycorrhizal fungal diversity: Separating the wheat from the chaff. Fungal Diversi. 2008, 33, 1–45. [Google Scholar]
- Simard, S.W.; Durall, D.M. Mycorrhizal networks: A review of their extent, function, and importance. Can. J. Bot. 2004, 82, 1140–1165. [Google Scholar] [CrossRef]
- Tedersoo, L.; Smith, M.E.; May, T.W. Ectomycorrhizal lifestyle in fungi: Global diversity, distribution, and evolution of phylogenetic lineages. Mycorrhiza 2010, 20, 217–263. [Google Scholar] [CrossRef]
- Finlay, R.D. Ecological aspects of mycorrhizal symbiosis: With special emphasis on the functional diversity of interactions involving the extraradical mycelium. J. Exp. Bot. 2008, 59, 1115–1128. [Google Scholar] [CrossRef]
- Sebastiana, M.; Martins, J.; Figueiredo, A.; Monteiro, F.; Sardans, J. Oak protein profile alterations upon root colonization by an ectomycorrhizal fungus. Mycorrhiza 2017, 27, 109–128. [Google Scholar] [CrossRef] [Green Version]
- Rudawska, M.; Pietras, M.; Smutek, I.; Strzeliński, P.; Leski, T. Ectomycorrhizal fungal assemblages of Abies alba Mill. outside its native range in Poland. Mycorrhiza 2016, 26, 57–65. [Google Scholar] [CrossRef] [Green Version]
- Delgado-Baquerizo, M.; Reich, P.B.; Trivedi, C.; Eldridge, D.J. Multiple elements of soil biodiversity drive ecosystem functions across biomes. Nat. Ecol. Evol. 2020, 4, 210–220. [Google Scholar] [CrossRef]
- Erlandson, R.S.; Savage, J.A.; Cavender-Bares, J.M.; Peay, K.G. Soil moisture and chemistry influence diversity of ectomycorrhizal fungal communities associating with willow along a hydrologic gradient. FEMS Microbiol. Ecol. 2016, 92. [Google Scholar] [CrossRef] [Green Version]
- Garcia, M.; Jane, O.; Smith, E.; Daniel, E.; Luoma, L.; Melanie, L.; Jones, D. Ectomycorrhizal communities of ponderosa pine and lodgepole pine in the south-central Oregon pumice zone. Mycorrhiza 2016, 26, 275–286. [Google Scholar] [CrossRef]
- Corrales, A.; Arnold, A.E.; Ferrer, A.; Turner, B.L.; Dalling, J.W. Variationin ectomycorrhizal fungal communities associated with Oreomunnea mexicana (Juglandaceae) in a Neotropical montane forest. Mycorrhiza 2016, 26, 1–17. [Google Scholar] [CrossRef]
- Cox, F.; Barsoum, N.; Lilleskov, E.A.; Bidartondo, M.I. Nitrogen availability is a primary determinant of conifer mycorrhizas across complex environmental gradients. Ecol. Lett. 2010, 13, 1103–1113. [Google Scholar] [CrossRef]
- Benucci, G.M.N.; Bonito, G.M. The Truffle Microbiome: Species and geography effects on bacteria associated with fruiting bodies of hypogeous Pezizales. Microb. Ecol. 2016, 72, 4–8. [Google Scholar] [CrossRef]
- Szuba, A. Ectomycorrhiza of Populus. Forest Ecol. Manag. 2015, 347, 156–169. [Google Scholar] [CrossRef]
- Walker, J.; Miller, O.; Horton, J. Hyperdiversity of ectomycorrhizal fungus assemblages on oak seedlings in mixed forests in the southern Appalachian Mountains. Mol. Ecol. 2010, 14, 829–838. [Google Scholar] [CrossRef]
- He, F.; Yang, B.S.; Wang, H.; Yan, Q.L.; Cao, Y.N.; He, X.H. Changes in composition and diversity of fungal communities along Quercus mongolica forests developments in Northeast China. Appl. Soil Ecol. 2016, 100, 162–171. [Google Scholar] [CrossRef]
- Behnke-Borowczyk, J.; Kowalkowski, W.; Kartawik, N.; Baranowska, M.; Barzdajn, W. Soil fungal communities in nurseries producing Abies alba. Balt For. 2020, 26, 426. [Google Scholar] [CrossRef]
- Jin, W.; Yang, Y.Z.; Sun, H.J.; Yuan, Z.L. Diversity of Ectomycorrhizal Fungi a Seed Collecting Forest of Quercus virginiana. Sci. Silvae Sin. 2020, 56, 1001–7488. [Google Scholar]
- Voříšková, J.; Brabcová, V.; Cajthaml, T.; Baldrian, P. Seasonal dynamics of fungal communities in a temperate Oak forest soil. New Phytol. 2014, 201, 269–278. [Google Scholar] [CrossRef]
- Salvati, L.; Ranalli, F.; Gitas, I. Landscape fragmentation and the agro-forest ecosystem along a rural-to-urban gradient: An exploratory study. Int. J. Sustain. Dev. World Ecol. 2014, 21, 160–167. [Google Scholar] [CrossRef]
- Cardou, F.; Aubin, I.; Bergeron, A.; Shipley, B. Functional markers to predict forest ecosystem properties along a rural-to-urban gradient. J. Veg. Sci. 2020, 31, 416–428. [Google Scholar] [CrossRef]
- Bu, F.Q.; Yan, H.; Fan, M.J. Suburban communities in ecologically sensitive areas of vegetation Optimization—Case study of jinan city, the southern mountains. Shandong For. Sci. Technol. 2014, 44, 16–20. [Google Scholar]
- The 7th National Census of Jinan. Available online: http://m.iqilu.com/pcarticle/4882711?ivk_sa=1024320u (accessed on 16 June 2021).
- Vance, E.D.; Brookes, P.C.; Jenkinson, D.S. An extraction method for measuring soil microbial biomass C. Soil Biol. Biochem. 1987, 19, 703–707. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, M.; Huang, S.Y.; Zha, X. Assessing spatial variability of soil organic carbon and total nitrogen in eroded hilly region of subtropical China. PLoS ONE 2020, 15, e0244322. [Google Scholar] [CrossRef]
- Gao, G.F.; Li, P.F.; Zhong, J.X.; Shen, Z.J.; Chen, J.; Li, Y.T.; Isabwe, A.; Zhu, X.Y.; Ding, Q.S.; Zhang, S.; et al. Spartina alterniflflora invasion alters soil bacterial communities and enhances soil N2O emissions by stimulating soil denitrifification in mangrove wetland. Sci. Total Environ. 2019, 653, 231–240. [Google Scholar] [CrossRef]
- Foster, J.C. Soil sampling, handling, storage and analysis. In Methods in Applied Soil Microbiology and Biochemistry; Academic Press: New York, NY, USA, 1995; pp. 49–121. [Google Scholar]
- Richard, F.; Roy, M.; Shahin, O. Ectomycorrhizal communities in a Mediterranean forest ecosystem dominated by Quercus ilex: Seasonal dynamics and response to drought in the surface organic horizon. Ann. Forest Sci. 2011, 68, 57–68. [Google Scholar] [CrossRef] [Green Version]
- Garcia, K.; Doidy, J.; Zimmermann, S.D.; Wipf, D.; Courty, P.E. Take a trip through the plant and fungal transportome of mycorrhiza. Trends. Plant Sci. 2016, 21, 937–950. [Google Scholar] [CrossRef] [PubMed]
- Van Dam, N.M.; Bouwmeester, H.J. Metabolomics in the rhizosphere: Tapping into belowground chemical communication. Trends. Plant Sci. 2016, 21, 256–265. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Gao, M.H. Community structure and driving factors for rhizosphere ectomycorrhizal fungi of Betula platyphylla in Daqing Mountain. Chin. J. Ecol. 2021, 40, 1244–1252. [Google Scholar]
- Treseder, K.K. A meta-analysis of mycorrhizal responses to nitrogen, phosphorus, and atmospheric CO2 in field studies. New Phytol. 2004, 164, 347–355. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, D.J.E.; Pouyat, R.; Szlavecz, K.; Setälä, H.; Kotze, D.J.; Yesilonis, I.; Sarel, C. Urbanization erodes ectomycorrhizal fungal diversity and may cause microbial communities to converge. Nat. Ecol. Evol. 2017, 1, 439–447. [Google Scholar] [CrossRef]
- Ashley, S.; Hannes, P.; Steven, A.D.; Baho, D.L.; Mercè, B. Fundamentals of microbial community resistance and resilience. Front. Microbiol. 2012, 3, 417. [Google Scholar] [CrossRef] [Green Version]
- Tedersoo, L.; Suvi, T.; Larsson, E.; Kõljalg, U. Diversity and community structure of ectomycorrhizal fungi in a wooded meadow. Mycol. Res. 2006, 110, 734–748. [Google Scholar] [CrossRef]
- Yang, R.H.; Li, Y.; Wu, Y.Y.; Tang, L.H.; Shang, J.J.; Bao, D.P. Genome-based analysis of lignocellulose-degrading enzyme systems in different Lentinus edodes strains. Acta Edulis Fungi 2018, 25, 15–22. [Google Scholar]
- Wieg, B.D.; Durall, D.M.; Simard, S.W. Ectomycorrhizal fungal succession in mixed temperate forests. New Phytol. 2007, 176, 437–447. [Google Scholar]
- Redecker, D.; Szaro, T.M.; Bowman, R.J.; Bruns, T.D. Small genets of Lactarius xanthogalactus, Russula cremoricolor and Amanita francheti in late-stage ectomycorrhizal successions. Mol. Ecol. 2001, 10, 1025–1034. [Google Scholar] [CrossRef] [Green Version]
- Yang, B.S.; He, F.; Zhao, X.H.; Wang, H.; Xu, X.H.; He, X.H.; Zhu, Y.D. Composition and function of soil fungal community during the establishment of Quercus acutissima seedlings in a Cd-contaminated soil. J. Environ. Manag. 2019, 246, 150–156. [Google Scholar] [CrossRef]
- Jiang, Z.Y.; Zou, W.Q.; Yang, L.; Li, W.Z.; Zhang, H.D.; Chen, X.W.; Wang, X.W. Root exudation rate and rhizosphere effect of different mycorrhizal associations of tree species in typical black soil area. Chin. J. Ecol. 2021, 33, 2810–2816. [Google Scholar] [CrossRef]
- Liu, X.L.; Dou, L.; Ding, X.H.; Sun, T.; Zhang, H.J. Influences of different afforestation systems on the soil properties of limestone mountains in the mid-eastern region of China. Catena 2021, 201, 105198. [Google Scholar] [CrossRef]
- Fang, F.; Wu, C.Z.; Hong, W.; Fang, H.L.; Song, P. Study on relationship between plant rhizosphere and non-rhizosphere soil enzymes and microorganisms. Subtrop. Agric. Res. 2016, 3, 209–215. [Google Scholar]
- Wang, X.; Liu, J.; Long, D.; Han, Q.; Huang, J. The ectomycorrhizal fungal communities associated with Quercus liaotungensis in different habitats across northern China. Mycorrhiza 2017, 27, 441–449. [Google Scholar] [CrossRef] [PubMed]
- Kyaschenko, J.; Clemmensen, K.E.; Karltun, E.; Lindahl, B.D. Below-ground organic matter accumulation along a boreal forest fertility gradient relates to guild interaction within fungal communities. Ecol. Lett. 2017, 20, 1546–1555. [Google Scholar] [CrossRef]
- Zhou, Y.; Hartemink, A.E.; Shi, Z.; Liang, Z.; Lu, Y. Land use and climate change effects on soil organic carbon in North and Northeast China. Sci. Total Environ. 2019, 647, 1230–1238. [Google Scholar] [CrossRef]
- Dong, W.Y.; Zhang, X.Y.; Liu, X.Y.; Fu, X.L.; Chen, F.S.; Wang, H.M.; Sun, X.M.; Wen, X.M. Responses of soil microbial communities and enzyme activities to nitrogen and phosphorus additions in Chinese fir plantations of subtropical China. Biogeosciences 2016, 12, 5537–5546. [Google Scholar] [CrossRef] [Green Version]
- Dong, S.K.; Wen, L.; Li, Y.Y.; Wang, X.X.; Zhu, L.; Li, X.Y. 2012. Soil-quality effects of grassland degradation and restoration on the Qinghai-Tibetan Plateau. Soil Sci. Soc. Am. J. 2012, 76, 2256–2264. [Google Scholar] [CrossRef]
- Wu, Z.Z.; Cheng, H.G.; Wang, J.T.; Cheng, Q.D. Effects of biochar addition ratio and freezing-thawing on nitrogen leaching from ditched soil. J. Agro-Environ. Sci. 2019, 39, 1295–1302. [Google Scholar]
- Hao, L.F.; Hao, W.Y.; Wang, X.F.; Liu, H.; Liu, T.Y. Response of four ectomycorrhizal fungi to nitrogen sources. South. For. Sci. 2020, 48, 20–24. [Google Scholar]
- Simkin, S.M.; Allen, E.B.; Bowman, W.D.; Clark, C.M. Conditional vulnerability of plant diversity to atmospheric nitrogen deposition across the United States. Proc. Natl. Acad. Sci. USA 2016, 113, 4086–4091. [Google Scholar] [CrossRef] [Green Version]
- Olchowik, J.; Hilszczańska, D.; Bzdyk, R.M.; Studnicki, M.; Malewski, T.; Borowski, Z. Effect of deadwood on ectomycorrhizal colonisation of old-growth oak forests. Forests 2019, 10, 480. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Maps of the sampling site and study area location.

Figure 2.
MBC and MBN contents in Quercus acutissima forest soil along the urban–rural gradient. Different letters in each column indicate significant differences in soil samples (p < 0.05).
Figure 2.
MBC and MBN contents in Quercus acutissima forest soil along the urban–rural gradient. Different letters in each column indicate significant differences in soil samples (p < 0.05).

Figure 3.
The relative abundance of ECM fungi at the order (a) and genus (b) levels in the soil of Quercus acutissima forests along the urban–rural gradient.
Figure 3.
The relative abundance of ECM fungi at the order (a) and genus (b) levels in the soil of Quercus acutissima forests along the urban–rural gradient.

Figure 4.
Cladogram (a) and LDA distribution histogram (b) for ECM fungi in the soil of Quercus acutissima forests along the urban–rural gradient.
Figure 4.
Cladogram (a) and LDA distribution histogram (b) for ECM fungi in the soil of Quercus acutissima forests along the urban–rural gradient.

Figure 5.
RDA of dominant ECM fungi and soil physicochemical properties in Quercus acutissima forests. The solid and dashed arrows represent ECM fungi and soil properties.
Figure 5.
RDA of dominant ECM fungi and soil physicochemical properties in Quercus acutissima forests. The solid and dashed arrows represent ECM fungi and soil properties.

Figure 6.
Pearson correlation heat map of soil physicochemical properties and ECM fungal diversity. Red is positive, blue is negative; the darker color means the correlation is stronger.
Figure 6.
Pearson correlation heat map of soil physicochemical properties and ECM fungal diversity. Red is positive, blue is negative; the darker color means the correlation is stronger.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Soil physicochemical properties in Quercus acutissima forests along the urban–rural gradient.
Table 1.
Soil physicochemical properties in Quercus acutissima forests along the urban–rural gradient.
| Sampling Site | pH | SOM (g∙kg−1) | TN (g∙kg−1) | NH4+-N (mg∙kg−1) | NO3−-N (mg∙kg−1) | AP (g∙kg−1) | AK (g∙kg−1) | C/N |
|---|---|---|---|---|---|---|---|---|
| Urban | 7.87 ± 0.03 a | 10.88 ± 4.13 b | 2.67 ± 0.09 b | 7.28 ± 0.03 b | 11.65 ± 1.59 a | 17.92 ± 1.11 a | 297.61 ± 9.38 a | 3.26 ± 1.28 a |
| Suburban | 4.88 ± 0.11 b | 27.16 ± 9.88 a | 2.92 ± 0.03 a | 8.59 ± 0.19 a | 3.81 ± 0.68 b | 5.22 ± 0.06 b | 242.02 ± 15.24 b | 5.44 ± 1.05 a |
| Rural | 4.50 ± 0.31 b | 12.40 ± 8.40 b | 2.89 ± 0.01 a | 8.81 ± 0.13 a | 3.48 ± 0.20 b | 4.42 ± 0.10 b | 228.14 ± 9.80 b | 2.50 ± 1.70 a |
Different letters in each column indicate significant differences in soil samples (p < 0.05). The values in the table represent the mean ± standard error.
Table 2.
Community diversity indices of ECM fungi in Quercus acutissima forest soil along the urban–rural gradient.
Table 2.
Community diversity indices of ECM fungi in Quercus acutissima forest soil along the urban–rural gradient.
| Sampling Site | Shannon Index | Simpson Index | Chao1 Index | Good’s Coverage |
|---|---|---|---|---|
| Urban | 2.54 ± 0.06 b | 0.89 ± 0.01 b | 58.00 ± 1.76 a | 0.992± 0.002 a |
| Suburban | 2.12 ± 0.04 c | 0.83 ± 0.01 c | 63.00 ± 10.50 a | 0.993± 0.001 a |
| Rural | 2.76 ± 0.02 a | 0.92 ± 0.00 a | 65.72 ± 2.97 a | 0.993± 0.001 a |
Different letters in each column indicate significant differences in soil samples (p < 0.05). The values in the table represent the mean ± standard error.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Shen, H.; Yang, B.; Wang, H.; Sun, W.; Jiao, K.; Qin, G. Changes in Soil Ectomycorrhizal Fungi Community in Oak Forests along the Urban–Rural Gradient. Forests 2022, 13, 675. https://0-doi-org.brum.beds.ac.uk/10.3390/f13050675
AMA Style
Shen H, Yang B, Wang H, Sun W, Jiao K, Qin G. Changes in Soil Ectomycorrhizal Fungi Community in Oak Forests along the Urban–Rural Gradient. Forests. 2022; 13(5):675. https://0-doi-org.brum.beds.ac.uk/10.3390/f13050675
Chicago/Turabian StyleShen, Hongyan, Baoshan Yang, Hui Wang, Wen Sun, Keqin Jiao, and Guanghua Qin. 2022. "Changes in Soil Ectomycorrhizal Fungi Community in Oak Forests along the Urban–Rural Gradient" Forests 13, no. 5: 675. https://0-doi-org.brum.beds.ac.uk/10.3390/f13050675
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.