
Key Strategies Underlying the Adaptation of Mongolian Scots Pine (Pinussylvestris var. mongolica) in Sandy Land under Climate Change: A Review


Abstract
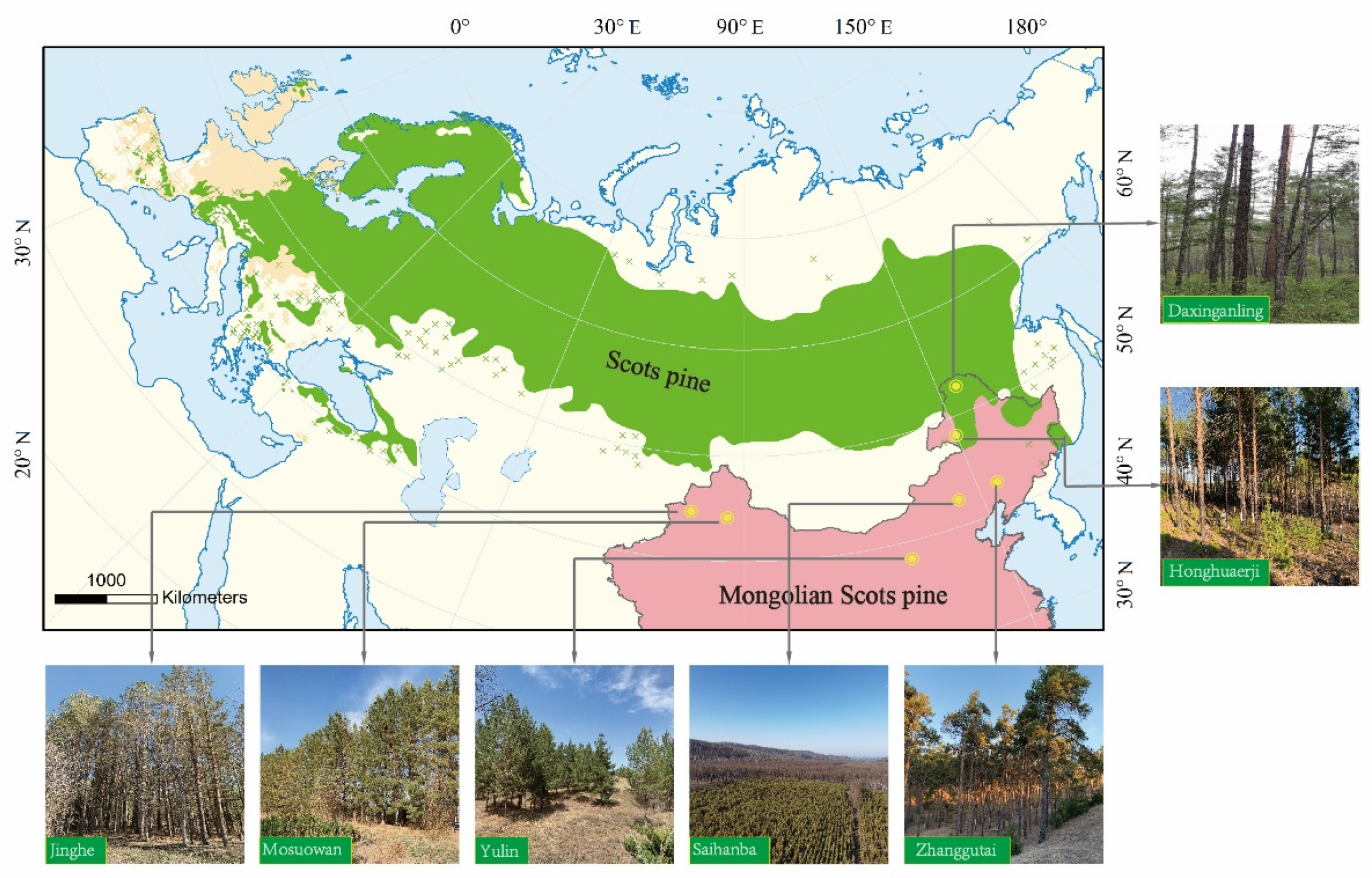
:1. Introduction
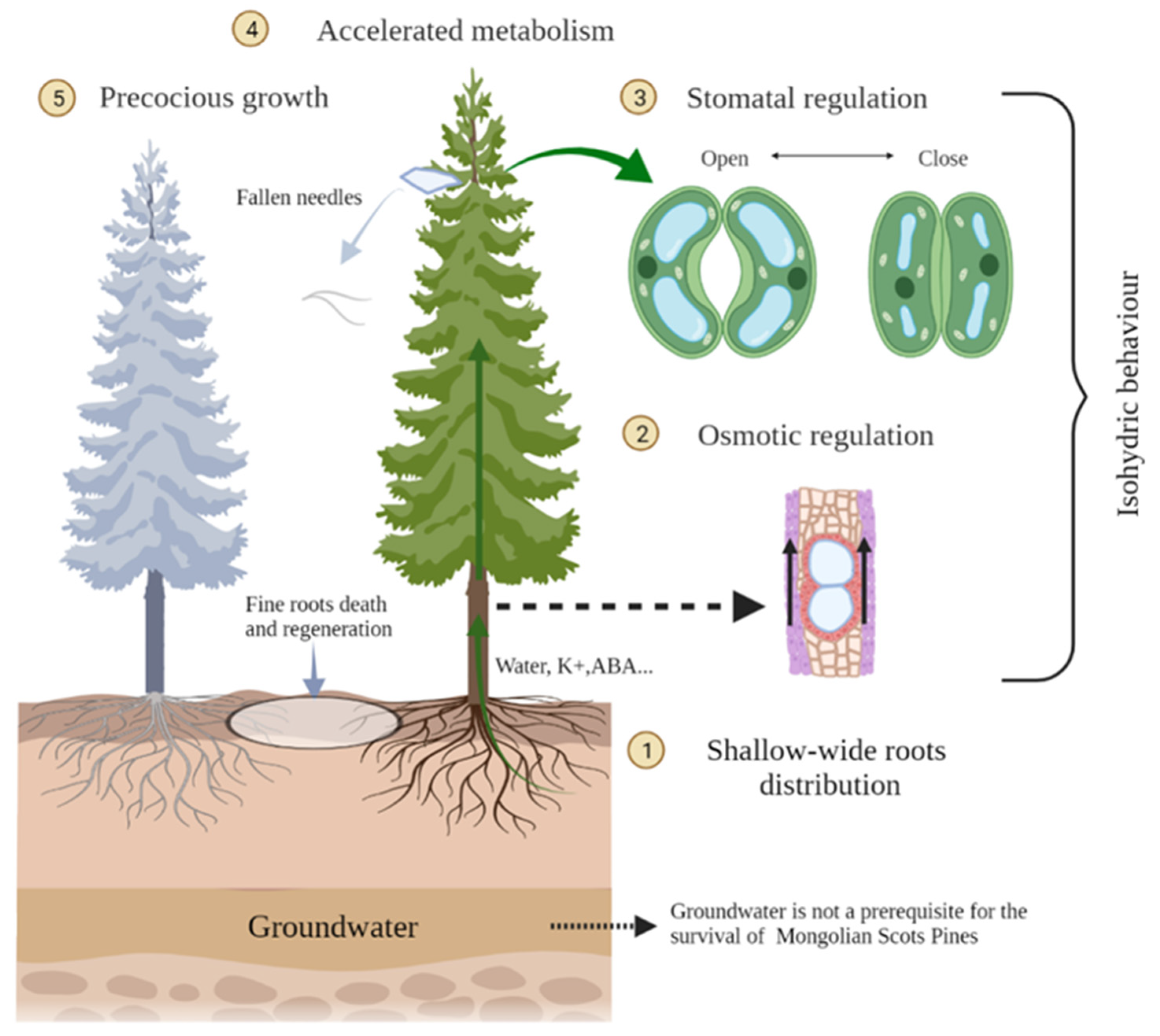
2. Key Strategies
2.1. Opportunistic Strategy of Water Uptake through a Shallow–Wide Root System
2.2. Hydraulic Failure Risk Avoidance Strategy by Biochemical Osmotic Regulation
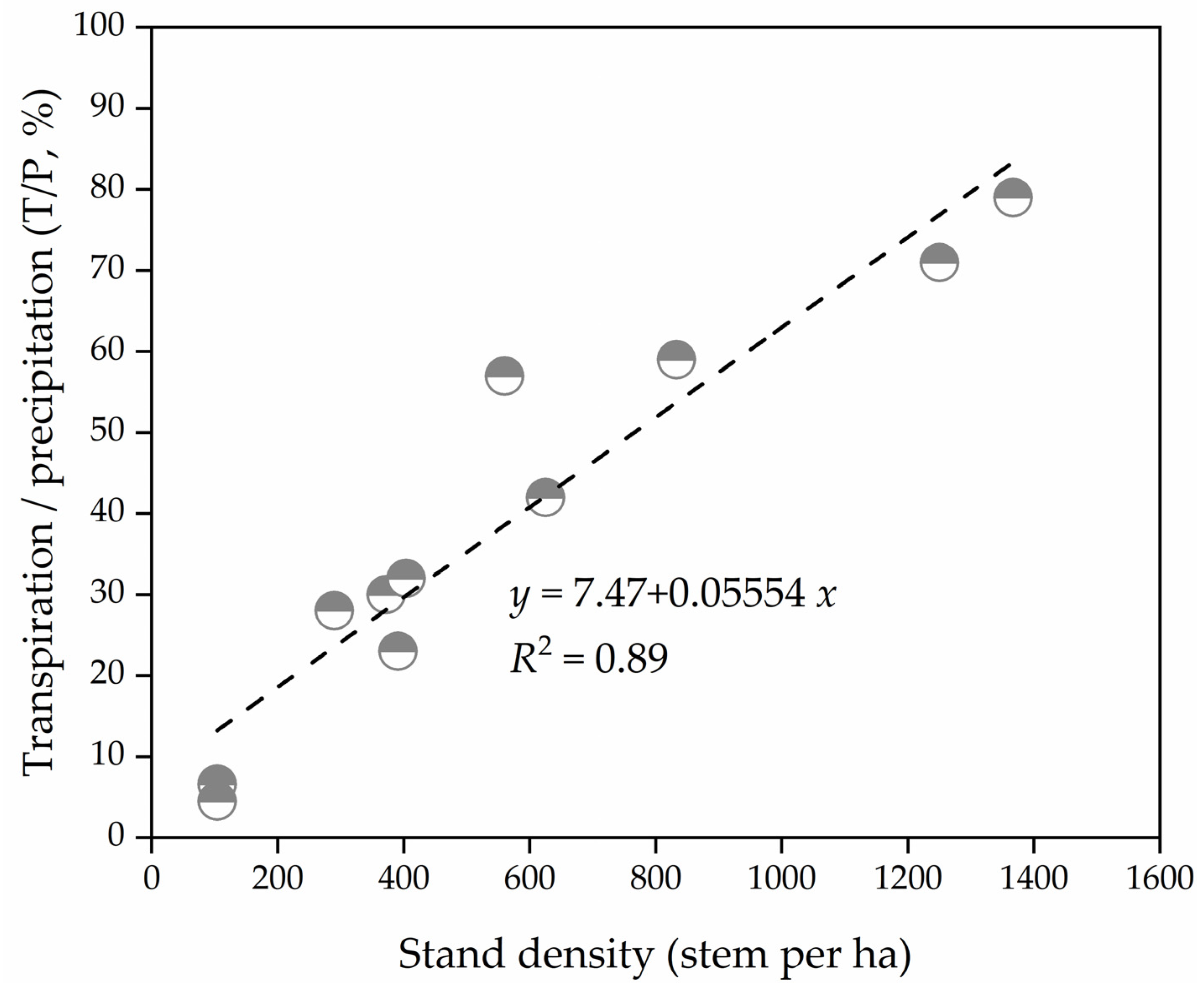
2.3. Water Conservation Strategy by Physiological Stomatal Regulation
2.4. Functional Traits Adjustment Strategy through Accelerated Metabolism
2.5. Rapid Regeneration Strategy through Precocious Growth
3. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Castro, J.; Zamora, R.; Hódar, J.A.; Gómez, J.M. Seedling establishment of a boreal tree species (Pinus sylvestris) at its southernmost distribution limit: Consequences of being in a marginal Mediterranean habitat. J. Hydrol. 2004, 92, 266–277. [Google Scholar] [CrossRef]
- Mason, W.L.; Alía, R. Current and future status of Scots pine (Pinus sylvestris L.) forests in Europe. For. Syst. 2000, 9, 317–335. [Google Scholar]
- Parfenova, E.I.; Kuzmina, N.A.; Kuzmin, S.R.; Tchebakova, N.M. Climate Warming Impacts on Distributions of Scots Pine (Pinus sylvestris L.) Seed Zones and Seed Mass across Russia in the 21st Century. Forests 2021, 12, 1097. [Google Scholar] [CrossRef]
- Jamyansuren, S.; Udval, B.; Batkhuu, N.; Bat-Erdene, J.J.; Michael, F. Result of study on developing forest seed region in Mongolia. Proc. Mong. Acad. Sci. 2019, 59, 18–31. [Google Scholar]
- Zhao, X.L.; Li, W.Y. Pinus sylvestris var. mongolica; Chinese Agricultural Press: Beijing, China, 1963. (In Chinese) [Google Scholar]
- Liu, Y.Y.; Wang, A.Y.; An, Y.N.; Lian, P.Y.; Wu, D.D.; Zhu, J.J.; Meinzer, F.C.; Hao, G.Y. Hydraulics play an important role in causing low growth rate and dieback of aging Pinus sylvestris var. mongolica trees in plantations of Northeast China. Plant Cell Environ. 2018, 41, 1500–1511. [Google Scholar]
- Jiao, S.R. Causes and control measures of the early decline of Pinus sylvestris var. mongolica sand-fixation forest in Zhanggutai, Liaoning Province. Sci. Silvae Sin. 2001, 37, 131–138. [Google Scholar]
- Xu, H.C. Forestry in the Great Khingan Regions in China; Chinese Science Press: Beijing, China, 1998. (In Chinese) [Google Scholar]
- Liu, G.F.; Yang, C.P.; Yang, S.W.; Xia, D.; Zhang, P.G. Adaptative scope of Pinus sylvestris var. mongolica by the introduction as exotic species. J. Northeast For. Univ. 1990, 18, 122–128. (In Chinese) [Google Scholar]
- Li, L.L.; Li, L.G.; Chen, Z.J.; Zhou, Y.B.; Zhang, X.L.; Bai, X.P.; Chang, Y.X.; Xiao, J.Q. Response of radial growth of Pinus sylvestris var. mongolica plantation to the hydrothermal gradient in Liaoning Province. Acta Ecol. Sin. 2015, 35, 4508–4517. [Google Scholar]
- Wang, J.H.; Man, D.Q.; Liu, H.J. Adaptability and developing potential of Pinus sylvestris var. mongolica in an arid area in Gansu province, China. J. Desert Res. 1999, 19, 94–98. (In Chinese) [Google Scholar]
- Liu, F.; Zhang, Y.X.; Ma, Y.B.; Dong, L.L.; Yu, X.C.; Huang, Y.R. Growth rhythm of Pinus sylvestris var. mongolica in the Ulan Buh Desert. J. Desert Res. 2015, 35, 1234–1238. (In Chinese) [Google Scholar]
- LI, M.M.; Ding, G.D.; Gao, G.L.; Zhao, Y.Y.; Yu, M.H.; Wang, D.Y. Suitability of Pinus sylvestris var. mongolica introduced to 10 provinces in northern China. J. Desert Res. 2016, 36, 1021–1028. (In Chinese) [Google Scholar]
- Zhu, J.J.; Zheng, X. Thoughts and prospects on the construction of the Three-North Afforestation Program (TNAP): Based on the general assessment results of the TNAP constructed for 40 years. Chin. J. Ecol. 2019, 38, 1600–1610. (In Chinese) [Google Scholar]
- Song, X.D. Introduction of Pinus sylvestris var. mongolica in sandy land areas in northern China. For. Sci. Technol. Liaoning Prov. 1988, 5, 7–10. (In Chinese) [Google Scholar]
- Jiang, F.Q.; Zeng, D.H.; Yu, Z.Y. Decline of protective forest and its prevention strategies from the viewpoint of restoration ecology: Taking Pinus sylvestris var. mongolica plantation in Zhanggutai as an example. Chin. J. Appl. Ecol. 2006, 17, 2229–2235. (In Chinese) [Google Scholar]
- Song, L.N.; Zhu, J.J.; Zheng, X. Forestation and management scheme based on decline mechanisms of Pinus sylvestris var. mongolica plantations in sandy lands. Acta Ecol. Sin. 2017, 36, 3249–3256. [Google Scholar]
- Dang, H.Z.; Lu, P.; Yang, W.B.; Han, H.; Zhang, J. Drought-induced reductions and limited recovery in the radial growth, transpiration, and canopy stomatal conductance of Mongolian Scots pine (Pinus sylvestris var. mongolica L.): A five-year observation. Forests 2019, 10, 1143. [Google Scholar] [CrossRef] [Green Version]
- Allen, C.D.; Macalady, A.K.; Chenchouni, H.; Bachelet, D.; McDowell, N.; Vennetier, M.; Kitzberger, T.; Rigling, A.; Breshears, D.D.; Hogg, E.H.T.; et al. A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. For. Ecol. Manag. 2010, 259, 660–684. [Google Scholar] [CrossRef] [Green Version]
- Qin, D.H. Climate change science and sustainable development. Prog. Geogr. 2014, 33, 874–883. [Google Scholar]
- Morecroft, M.D.; Duffield, S.; Harley, M.; Pearce-Higgins, J.W.; Stevens, N.; Watts, O.; Whitaker, J. Measuring the success of climate change adaptation and mitigation in terrestrial ecosystems. Science 2019, 366, w9256. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Sun, P.S.; Huettmann, F.; Liu, S.R. Where should China practice forestry in a warming world? Glob. Chang. Biol. 2021, 28, 2461–2475. [Google Scholar] [CrossRef]
- Poyatos, R.; Llorens, P.; Gallart, F. Transpiration of montane Pinus sylvestris L. and Quercus pubescens Willd. Forest stands measured with sap flow sensors in NE Spain. Hydrol. Earth Syst. Sci. 2005, 9, 493–505. [Google Scholar] [CrossRef] [Green Version]
- Allen, C.D.; Breshears, D.D.; Mcdowell, N.G. On underestimation of global vulnerability to tree mortality and forest die-off from hotter drought in the Anthropocene. Ecosphere 2015, 6, 1–55. [Google Scholar] [CrossRef]
- Llorens, P.; Poyatos, R.; Latron, J.; Delgado, J.; Oliveras, I.; Gallart, F. A multi-year study of rainfall and soil water controls on Scots pine transpiration under Mediterranean mountain conditions. Hydrol. Process. 2010, 24, 3053–3064. [Google Scholar] [CrossRef]
- Leo, M.; Oberhuber, W.; Schuster, R.; Grams, T.E.E.; Matyssek, R.; Wieser, G. Evaluating the effect of plant water availability on inner alpine coniferous trees based on sap flow measurements. Eur. J. For. Res. 2013, 133, 691–698. [Google Scholar] [CrossRef]
- Giuggiola, A.; Kuster, T.M.; Saha, S. Drought-induced mortality of Scots pines at the southern limits of its distribution in Europe: Causes and consequences. iForest Biogeosciences For. 2010, 3, 95–97. [Google Scholar] [CrossRef] [Green Version]
- Barbeta, A.; Mejía-Chang, M.; Ogaya, R.; Voltas, J.; Dawson, T.E.; Peñuelas, J. The combined effects of a long-term experimental drought and an extreme drought on the use of plant-water sources in a Mediterranean forest. Glob. Chang. Biol. 2015, 21, 1213–1225. [Google Scholar] [CrossRef] [Green Version]
- Christoforos, P.; Matheny, A.M.; Baltzer, J.L.; Barr, A.G.; Andrew, B.T.; Gil, B.; Matteo, D.; Jason, M.; Alexandre, R.; Oliver, S. Boreal tree hydrodynamics: Asynchronous, diverging, yet complementary. Tree Physiol. 2018, 38, 953–964. [Google Scholar]
- Anderegg, W.R.; Kane, J.M.; Anderegg, L.D. Consequences of widespread tree mortality triggered by drought and temperature stress. Nat. Clim. Chang. 2013, 3, 30–36. [Google Scholar] [CrossRef]
- Popkin, G. Forest fight. Science 2021, 374, 1184–1189. [Google Scholar] [CrossRef]
- Prietzel, J.; Falk, W.; Reger, B.; Uhl, E.; Pretzsch, H.; Zimmermann, L. Half a century of Scots pine forest ecosystem monitoring reveals long-term effects of atmospheric deposition and climate change. Glob. Chang. Biol. 2020, 26, 5796–5815. [Google Scholar] [CrossRef]
- García -Valdés, R.; Estrada, A.; Early, R.; Lehsten, V.; Morin, X. Climate change impacts on long-term forest productivity might be driven by species turnover rather than by changes in tree growth. Glob. Ecol. Biogeogr. 2020, 29, 1360–1372. [Google Scholar] [CrossRef]
- Jiao, S.R. Study on distribution and succession process of the vegetations in sandy land in Zhanggutai, Liaoning province of China. Prot. For. Sci. Technol. 2006, 16, 11–13. [Google Scholar]
- Fu, B.J.; Lv, Y.H.; Gao, G.Y. Major research progresses on the ecosystem service and ecological safety of main terrestrial ecosystems in China. Chin. J. Nat. 2012, 34, 261–272. (In Chinese) [Google Scholar]
- Ryan, M.G. Tree responses to drought. Tree Physiol. 2011, 31, 237–239. [Google Scholar] [CrossRef] [Green Version]
- Novick, K.A.; Ficklin, D.L.; Stoy, P.C.; Williams, C.A.; Bohrer, G.; Oishi, A.C.; Papuga, S.A.; Blanken, P.D.; Noormets, A.; Sulman, B.N. The increasing importance of atmospheric demand for ecosystem water and carbon fluxes. Nat. Clim. Chang. 2016, 6, 1023–1027. [Google Scholar] [CrossRef] [Green Version]
- Li, S.G. A preliminary study on adaptation of Mongolian Scots pine to sandy land. J. Desert Res. 1994, 14, 60–67. [Google Scholar]
- Jiang, F.Q.; Cao, C.Y.; Zeng, D.H. Degradation and Restoration of Ecosystems in Horqin Sand Land; Chinese Forestry Press: Beijing, China, 2002. (In Chinese) [Google Scholar]
- Meng, P.; Zhang, B.X.; Wang, M. Biomass distribution and roots architecture of Pinus densiflora and Pinus sylvestris var. mongolica in Horqin Sandy Land. Chin. J. Ecol. 2018, 37, 2935–2941. (In Chinese) [Google Scholar]
- Wang, K.; Song, L.N.; Lv, L.Y.; Zhang, L.; Qin, Z.Y. Adaptive strategies of fine roots from main afforestation tree species in Horqin Sandy Land. J. Arid. Land Resour. Environ. 2014, 28, 128–131. (In Chinese) [Google Scholar]
- Zhang, J.C.; Wang, J.; Li, A.D.; Er, Y.H. Research on root distribution and growth adaptability of Pinus sylvestris var. mongolica. Prot. For. Sci. Technol. 2000, 3, 46–49. (In Chinese) [Google Scholar]
- Su, F.L.; Liu, M.G.; Guo, C.J.; Zhang, Q. Characteristics of the vertical distribution of the root system of Mongolian scotch pine growing in sandy land and influencing the soil. Soil Water Conserv. China 2006, 52, 20–22. (In Chinese) [Google Scholar]
- Wei, Y.F.; Fang, J.; Zhao, X.Y.; Mei, Y.I. An isotopic model estimate of the relative contribution of potential water pools to water uptake of Pinus sylvestris var. mongolica in Horqin Sandy Land. J. Resour. Ecol. 2012, 3, 308–315. (In Chinese) [Google Scholar]
- Fang, J.; We, Y.F.; Liu, S.; Zhao, X.Y.; Li, S.G. Stable isotopic analysis on water utilization sources of Pinus sylvestris var. mongolica plantations in inter-dune lowland in Horqin Sandy Land. Chin. J. Ecol. 2011, 30, 1894–1900. (In Chinese) [Google Scholar]
- Niu, C.Y.; Liu, Y.; Guo, Y.H.; Tian, Y.H.; Wang, J.L.; Zhang, W. Characteristics of sand-fixation plantations roots and soil moisture in Horqin Sandy Land. J. Arid. Land Resour. Environ. 2015, 29, 106–111. (In Chinese) [Google Scholar]
- Liu, B.Q.; Liu, Z.M.; Qian, J.Q.; Alamusa; Zhang, F.L.; Peng, X.H. Water sources of sand-fixing dominant plants during dry seasons in the southern edge of Horqin Sandy Land, China. Chin. J. Appl. Ecol. 2017, 28, 2093–2101. (In Chinese) [Google Scholar]
- Huxman, T.E.; Snyder, K.A.; Tissue, D.; Leffler, A.J.; Ogle, K.; Pockman, W.T.; Sandquist, D.R.; Potts, D.L.; Schwinning, S. Precipitation pulses and carbon fluxes in semiarid and arid ecosystems. Oecologia 2004, 141, 254–268. [Google Scholar] [CrossRef]
- Chen, L.H.; Li, F.X.; Qiu, X.M.; Zhang, J.X. Eolian Sandy Soil in China; Chinese Science Press: Beijing, China, 1998. (In Chinese) [Google Scholar]
- Song, L.N.; Zhu, J.J.; Li, M.C.; Zhang, J.X. Water use patterns of Pinus sylvestris var. mongolica trees of different ages in semiarid sandy lands of Northeast China. Environ. Exp. Bot. 2016, 129, 94–107. [Google Scholar]
- Song, L.N.; Zhu, J.J.; Li, M.C.; Zhang, J.X.; Lv, L.Y. Sources of water used by Pinus sylvestris var. mongolica trees based on stable isotope measurements in a semiarid sandy region of Northeast China. Agric. Water Manag. 2016, 164, 281–290. [Google Scholar]
- Zhu, J.J.; Kang, H.Z.; Song, L.N.; Yan, Q.L. Seasonal variation of groundwater table in Pinus sylvestris var. mongolica plantations in the southern edge of Horqin Sandy Land. Chin. J. Ecol. 2009, 28, 1767–1772. (In Chinese) [Google Scholar]
- Song, L.N.; Zhu, J.J.; Yan, Q.L.; Li, M.C.; Yu, G.Q. Comparison of intrinsic water use efficiency between different aged Pinus sylvestris var. mongolica wide windbreaks in semiarid sandy land of northern China. Agrofor. Syst. 2015, 89, 477–489. [Google Scholar]
- Ji, K.; Sun, Z.; Fang, Y. Advance on drought resistance of trees. J. Nanjing For. Univ. 2006, 30, 123–128. (In Chinese) [Google Scholar]
- Merlin, M.; Perot, T.; Perret, S.; Korboulewsky, N.; Vallet, P. Effects of stand composition and tree size on resistance and resilience to drought in sessile oak and Scots pine. For. Ecol. Manag. 2015, 339, 22–33. [Google Scholar] [CrossRef] [Green Version]
- Omprakash; Gobu, R.; Bisen, P.; Baghel, M.; Chourasia, K.N. Resistance/Tolerance mechanism under water deficit (Drought) condition in plants. Int. J. Curr. Microbiol. Appl. Sci. 2017, 6, 66–78. [Google Scholar] [CrossRef]
- Zhu, M.Y.; Tian, Y.L.; Guo, L.S. Variation of drought-enduring characteristics of Pinus sylvestris var. mongolica under different climatical moisture. Chin. J. Appl. Ecol. 1996, 7, 250–254. (In Chinese) [Google Scholar]
- Stojnić, S.; Suchocka, M.; Benito-Garzón, M.; Torres-Ruiz, J.M.; Cochard, H.; Bolte, A.; Cocozza, C.; Cvjetković, B.; De Luis, M.; Martinez-Vilalta, J. Variation in xylem vulnerability to embolism in European beech from geographically marginal populations. Tree Physiol. 2017, 38, 173–185. [Google Scholar] [CrossRef] [PubMed]
- Heres, A.M.; Camarero, J.J.; Lopez, B.C.; Martinez-Vilalta, J. Declining hydraulic performances and low carbon investments in tree rings predate Scots pine drought-induced mortality. Trees-Struct. Funct. 2014, 28, 1737–1750. [Google Scholar] [CrossRef]
- Landsberg, J.; Waring, R. Water relations in tree physiology: Where to from here? Tree Physiol. 2017, 37, 18–32. [Google Scholar] [CrossRef]
- Martinez-Vilalta, J.; Pinol, J. Drought-induced mortality and hydraulic architecture in pine populations of the NE Iberian Peninsula. For. Ecol. Manag. 2002, 161, 247–256. [Google Scholar] [CrossRef]
- Cochard, H. Vulnerability of several conifers to air embolism. Tree Physiol. 1992, 11, 73–83. [Google Scholar] [CrossRef]
- Poyatos, R.; Llorens, P.; Pinol, J.; Rubio, C. Response of Scots pine (Pinus sylvestris L.) and pubescent oak (Quercus pubescens Willd.) to soil and atmospheric water deficits under Mediterranean mountain climate. Ann. For. Sci. 2008, 65, 306–318. [Google Scholar] [CrossRef] [Green Version]
- Gazol, A.; Ribas, M.; Gutierrez, E.; Camarero, J.J. Aleppo pine forests from across Spain show drought-induced growth decline and partial recovery. Agric. For. Meteorol. 2017, 232, 186–194. [Google Scholar] [CrossRef]
- Irvine, J.; Perks, M.P.; Grace, J. The response of Pinus sylvestris to drought: Stomatal control of transpiration and hydraulic conductance. Tree Physiol. 1998, 18, 393–402. [Google Scholar] [CrossRef] [PubMed]
- Dang, H.Z.; Zhang, L.Z.; Yang, W.B.; Feng, J.C.; Han, H.; Chen, Y.B. Severe drought strongly reduces water use and its recovery ability of mature Mongolian Scots pine (Pinus sylvestris var. mongolica Litv.) in a semi-arid sandy environment of northern China. J. Arid Land 2019, 11, 880–891. [Google Scholar]
- Sun, Y.R.; Zhu, J.J. Effects of soil water condition on membrane lipid peroxidation and protective enzyme activities of Pinus sylvestris var. mongolica seedlings. Chin. J. Ecol. 2008, 27, 729–734. (In Chinese) [Google Scholar]
- Wang, K.; Sheng, C.; Cao, P.; Song, L.N.; Yu, G.Q. Changes of non-structural carbohydrates of Pinus sylvestris var. mongolica seedlings in the process of drought-induced death. Chin. J. Appl. Ecol. 2018, 29, 3513–3520. (In Chinese) [Google Scholar]
- Chirino, E.; Bellot, J.; Sanchez, J.R. Daily sap flow rate as an indicator of drought avoidance mechanisms in five Mediterranean perennial species in semi-arid southeastern Spain. Trees 2011, 25, 593–606. [Google Scholar] [CrossRef]
- Preisler, Y.; Tatarinov, F.; Rohatyn, S.; Rotenberg, E.; Grünzweig, J.M.; Klein, T.; Yakir, D. Large variations in daily and seasonal patterns of sap flux among Aleppo pine trees in semi-arid forest reflect tree-scale hydraulic adjustments. In Proceedings of the EGU General Assembly Conference, Vienna, Austria, 17–22 April 2015. [Google Scholar]
- Li, D.; Si, J.H.; Zhang, X.Y.; Gao, Y.; Luo, H.; Qin, J.; Gao, G.L. The mechanism of changes in hydraulic properties of Populus euphratica in response to drought stress. Forests 2019, 10, 904. [Google Scholar] [CrossRef] [Green Version]
- Venturas, M.D.; Sperry, J.S.; Hacke, U.G. Plant xylem hydraulics: What we understand, current research, and future challenges. J. Integr. Plant Biol. 2017, 59, 356–389. [Google Scholar] [CrossRef] [Green Version]
- Martínez Vilalta, J.; Poyatos, R.; Aguadé, D.; Retana, J.; Mencuccini, M. A new look at water transport regulation in plants. New Phytol. 2014, 204, 105–115. [Google Scholar] [CrossRef] [Green Version]
- Carminati, A.; Ahmed, M.A.; Zarebanadkouki, M.; Cai, G.C.; Goran, L.; Javaux, M. Stomatal closure prevents the drop in soil water potential around roots. New Phytol. 2020, 226, 1541–1543. [Google Scholar] [CrossRef] [Green Version]
- Zhu, J.J.; Kang, H.Z.; Li, Z.H. Comparison of different types of drought stresses affecting photosynthesis of Mongolian Scots pine seedlings on sandy soils. J. Beijing For. Univ. 2006, 28, 57–63. (In Chinese) [Google Scholar]
- Dang, H.Z.; Han, H.; Chen, S.; Li, M.Y. A fragile soil moisture environment exacerbates the climate change-related impacts on the water use by Mongolian Scots pine (Pinus sylvestris var. mongolica) in northern China: Long-term observations. Agric. Water Manag. 2021, 251, 106857. [Google Scholar]
- Hu, X.S.; Wang, S.J. Review on water stress and drought tolerance in tree species. Sci. Silvae Sin. 1998, 34, 77–89. [Google Scholar]
- Takahashi, F.; Suzuki, T.; Osakabe, Y.; Betsuyaku, S.; Kondo, Y.; Dohmae, N.; Fukuda, H.; Yamaguchi-Shinozaki, K.; Shinozaki, K. A small peptide modulates stomatal control via abscisic acid in long-distance signaling. Nature 2018, 556, 235–238. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.L.; Han, S.J.; Huang, M.R. ABA change and other physiological responses of Pinus sylvestris var. mongolica to water shortage. J. Northeast For. Univ. 2001, 29, 40–43. [Google Scholar]
- Zhu, J.J.; Zeng, D.H.; Kang, H.Z.; Wu, X.Y.; Fan, Z.P. Decline of Pinus sylvestris var. mongolica Plantations in Sandy Land; Chinese Forestry Press: Beijing, China, 2005. (In Chinese) [Google Scholar]
- Chang, X.L.; Zhao, W.Z. Study on needle water physiology and soil moisture condition of the Pinus sylvestris var. mongolica and Populus sinoii plantation. J. Desert Res. 1990, 10, 18–24. (In Chinese) [Google Scholar]
- Jiao, S.R. Influence of aridity on Mongolian Scots pine plantation growth in sandy land in Zhanggutai. Sci. Silvae Sin. 1986, 22, 419–425. [Google Scholar]
- Zhao, W.Z.; Chang, X.L. Study on the relationship between stomatal movement and Mongolia Scots pine transpiration rate. J. Desert Res. 1995, 15, 241–243. (In Chinese) [Google Scholar]
- Bai, X.F.; Han, H.; Zhou, F.Y.; Yang, S.J.; Liu, B.C.; Lei, Z.Y.; Jin, Z. Analysis on the dynamic change of relative water content and needle water conservation of Pinus sylvestris var. mongolica in sandy land. Prot. For. Sci. Technol. 2008, 75, 51–53. (In Chinese) [Google Scholar]
- Li, X.H.; Jiang, D.M.; Luo, Y.M.; Li, R.P. Physiological characteristics of Pinus sylvestris var. mongolica seedlings at different irrigation treatments. Chin. J. Ecol. 2003, 22, 17–20. (In Chinese) [Google Scholar]
- Song, C.S. Species of pinus in the world and introduction in China. J. Beijing For. Univ. 1983, 2, 1–11. (In Chinese) [Google Scholar]
- Luo, D.D.; Wang, C.K.; Jin, Y. Stomatal regulation of plants in response to drought stress. Chin. J. Appl. Ecol. 2019, 30, 4333–4343. (In Chinese) [Google Scholar]
- Brodribb, T.J.; McAdam, S.A.; Jordan, G.J.; Martins, S.C. Conifer species adapt to low-rainfall climates by following two divergent pathways. Proc. Natl. Acad. Sci. USA 2014, 111, 14489–14493. (In Chinese) [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hochberg, U.; Rockwell, F.E.; Holbrook, N.M.; Cochard, H. Iso/anisohydry: A plant–environment interaction rather than a simple hydraulic trait. Trends Plant Sci. 2018, 23, 112–120. [Google Scholar] [CrossRef] [PubMed]
- Leuschner, C.; Schipka, F.; Backes, K. Stomatal regulation and water potential variation in European beech: Challenging the iso/anisohydry concept. Tree Physiol. 2021, 42, 365–378. [Google Scholar] [CrossRef] [PubMed]
- Ambrose, A.R.; Baxter, W.L.; Wong, C.S.; Næsborg, R.R.; Williams, C.B.; Dawson, T.E. Contrasting drought-response strategies in California redwoods. Tree Physiol. 2015, 35, 453–469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez Vilalta, J.; Garcia Forner, N. Water potential regulation, stomatal behavior and hydraulic transport under drought: Deconstructing the iso/anisohydric concept. Plant Cell Environ. 2017, 40, 962–976. [Google Scholar] [CrossRef] [Green Version]
- Pivovaroff, A.L.; Cook, V.M.; Santiago, L.S. Stomatal behavior and stem xylem traits are coordinated for woody plant species under exceptional drought conditions. Plant Cell Environ. 2018, 41, 2617–2626. [Google Scholar] [CrossRef]
- Guo, J.S.; Hultine, K.R.; Koch, G.W.; Kropp, H.; Ogle, K. Temporal shifts in iso/anisohydry revealed from daily observations of plant water potential in a dominant desert shrub. New Phytol. 2020, 225, 713–726. [Google Scholar] [CrossRef]
- Martínez-Sancho, E.; Vásconez Navas, L.K.; Seidel, H.; Dorado-Liñán, I.; Menzel, A. Responses of contrasting tree functional types to air warming and drought. Forests 2017, 8, 450. [Google Scholar] [CrossRef] [Green Version]
- Wieser, G.; Leo, M.; Oberhuber, W. Transpiration and canopy conductance in an inner alpine Scots pine (Pinus sylvestris L.) forest. Flora 2014, 209, 491–498. [Google Scholar] [CrossRef] [Green Version]
- Song, L.N.; Zhu, J.J.; Li, M.C.; Zhang, J.X.; Zheng, X.; Wang, K. Canopy transpiration of Pinus sylvestris var. mongolica in a sparse wood grassland in the semiarid sandy region of Northeast China. Agric. For. Meteorol. 2018, 250, 192–201. [Google Scholar]
- Han, H.; Bai, X.F.; Xu, G.J.; Zhang, B.X.; You, G.C.; Shang, S.C. Study on the water balance of Pinus sylvestris var. mongolica plantation in Zhanggutai, China. J. For. Sci. Technol. Liaoning Prov. China 2012, 6, 8–11, 14. (In Chinese) [Google Scholar]
- Song, L.N.; Zhu, J.J.; Zheng, X.; Wang, K.; Lu, L.Y.; Zhang, X.L.; Hao, G.Y. Transpiration and canopy conductance dynamics of Pinus sylvestris var. mongolica in its natural range and in an introduced region in the sandy plains of Northern China. Agric. For. Meteorol. 2020, 281, 107830. [Google Scholar]
- Zhu, J.J.; Zheng, X.; Yan, Q.L. Assessment of impacts of the Three-North Protective Forest Program on Ecological Environments by Remote Sensing Technology; China Science Press: Beijing, China, 2016. (In Chinese) [Google Scholar]
- Zheng, X.; Zhu, J.J.; Yan, Q.L.; Song, L.N. Effects of land use changes on the groundwater table and the decline of Pinus sylvestris var. mongolica plantations in southern Horqin Sandy Land, Northeast China. Agric. Water Manag. 2012, 109, 94–106. [Google Scholar]
- Fyllas, N.M.; Michelaki, C.; Galanidis, A.; Evangelou, E.; Zaragoza-Castells, J.; Dimitrakopoulos, P.G.; Tsadilas, C.; Arianoutsou, M.; Lloyd, J. Functional trait variation among and within species and plant functional types in mountainous Mediterranean forests. Front. Plant Sci. 2020, 11, 212. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.S.; Defrenne, C.; Mccormack, M.L.; Yang, L.; Tian, D.S.; Luo, Y.Q.; Hou, E.Q.; Yan, T.; Li, Z.L.; Bu, W.S.; et al. Fine-root functional trait responses to experimental warming: A global meta-analysis. New Phytol. 2021, 230, 1856–1867. [Google Scholar] [CrossRef]
- Martínez-Vilalta, J.; Cochard, H.; Mencuccini, M.; Sterck, F.; Herrero, A.; Korhonen, J.F.J.; Llorens, P.; Nikinmaa, E.; Nole, A.; Poyatos, R.; et al. Hydraulic adjustment of Scots pine across Europe. New Phytol. 2009, 184, 353–364. [Google Scholar]
- Song, L.N.; Zhu, J.J.; Li, M.C.; Yan, T.; Zhang, J.X. Stable carbon isotope composition and traits of needles of Pinus sylvestris var. mongolica in sparse wood grassland in the south edge of the Horqin Sandy Land under conditions with different precipitation levels. Chin. J. Appl. Ecol. 2012, 23, 1435–1440. (In Chinese) [Google Scholar]
- Wei, Y.F.; Fang, J.; Zhao, X.Y.; Li, S.G. Eco-physiological traits of needles at different ages for Pinus sylvestris var. mongolica plantations in Horqin Sandy Land of China. Chin. J. Plant Ecol. 2011, 35, 1271–1280. (In Chinese) [Google Scholar] [CrossRef]
- Palmroth, S.; Berninger, F.; Nikinmaa, E.; Lloyd, J.; Pulkkinen, P.; Hari, P. Structural adaptation rather than water conservation was observed in Scots pine over a range of wet to dry climates. Oecologia 1999, 121, 302–309. [Google Scholar] [CrossRef]
- Qiu, J.; Gu, J.C.; Jiang, H.Y.; Wang, Z.Q. Factors influencing fine root longevity of Pinus sylvestris var. mongolica plantation. Chin. J. Plant Ecol. 2010, 34, 1066–1074. (In Chinese) [Google Scholar]
- Zackrisson, O.; Nilsson, M.C.; Steijlen, I.; Hornberg, G. Regeneration pulses and climate vegetation interactions in nonpyrogenic boreal Scots pine stands. J. Ecol. 1995, 83, 469–483. [Google Scholar] [CrossRef]
- Jiang, F.Q.; Zeng, D.H.; Fan, Z.P.; Zhu, J.J. Simulation of individual tree growth of Mongolian Scots pine forest in sandy land. Chin. J. Appl. Ecol. 1996, 7, 1–5. (In Chinese) [Google Scholar]
- Zani, D.; Crowther, T.W.; Mo, L.D.; Renner, S.S.; Zohner, C.M. Increased growing-season productivity drives earlier autumn leaf senescence in temperate trees. Science 2020, 370, 1066. [Google Scholar] [CrossRef]
- Yan, W.M.; Zhong, Y.; Shangguan, Z.P. Responses of different physiological parameter thresholds to soil water availability in four plant species during prolonged drought. Agric. For. Meteorol. 2017, 247, 311–319. [Google Scholar] [CrossRef]
- Peters, M.P.; Iverson, L.R.; Matthews, S.N. Long-term droughtiness and drought tolerance of eastern US forests over five decades. For. Ecol. Manag. 2015, 345, 56–64. [Google Scholar] [CrossRef] [Green Version]
- Anderegg, W.R.; Plavcová, L.; Anderegg, L.D.; Hacke, U.G.; Berry, J.A.; Field, C.B. Drought’s legacy: Multiyear hydraulic deterioration underlies widespread aspen forest die-off and portends increased future risk. Glob. Chang. Biol. 2013, 19, 1188–1196. [Google Scholar] [CrossRef]
- Wu, X.Y.; Jiang, F.Q.; Li, X.D.; Xue, Y.; Qiu, S.F. Decline regularity and causes of Pinus sylvestris var. mongolica plantation in sandy land. Chin. J. Appl. Ecol. 2004, 15, 2225–2228. (In Chinese) [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dang, H.; Han, H.; Zhang, X.; Chen, S.; Li, M.; Liu, C. Key Strategies Underlying the Adaptation of Mongolian Scots Pine (Pinussylvestris var. mongolica) in Sandy Land under Climate Change: A Review. Forests 2022, 13, 846. https://0-doi-org.brum.beds.ac.uk/10.3390/f13060846
Dang H, Han H, Zhang X, Chen S, Li M, Liu C. Key Strategies Underlying the Adaptation of Mongolian Scots Pine (Pinussylvestris var. mongolica) in Sandy Land under Climate Change: A Review. Forests. 2022; 13(6):846. https://0-doi-org.brum.beds.ac.uk/10.3390/f13060846
Chicago/Turabian StyleDang, Hongzhong, Hui Han, Xueli Zhang, Shuai Chen, Mingyang Li, and Chunying Liu. 2022. "Key Strategies Underlying the Adaptation of Mongolian Scots Pine (Pinussylvestris var. mongolica) in Sandy Land under Climate Change: A Review" Forests 13, no. 6: 846. https://0-doi-org.brum.beds.ac.uk/10.3390/f13060846