2. Materials and Methods
The studies were conducted in the northern part of the Kamchatka Peninsula (
Figure 1a). Two sections were plotted: at the foot of volcano Shiveluch (
Figure 1b), one of the largest and most active volcanoes of the peninsula [
13], and at approximately 100 km from active volcanoes, in the middle ridge of Kamchatka on the southwestern foot of the extinct volcanoes Alnei–Chashakonda massif (
Figure 1c). In both cases, the soil profile is a fractional interstratification of buried soil horizons and volcanic ash and is identified as a soil-pyroclastic cover (SPC).
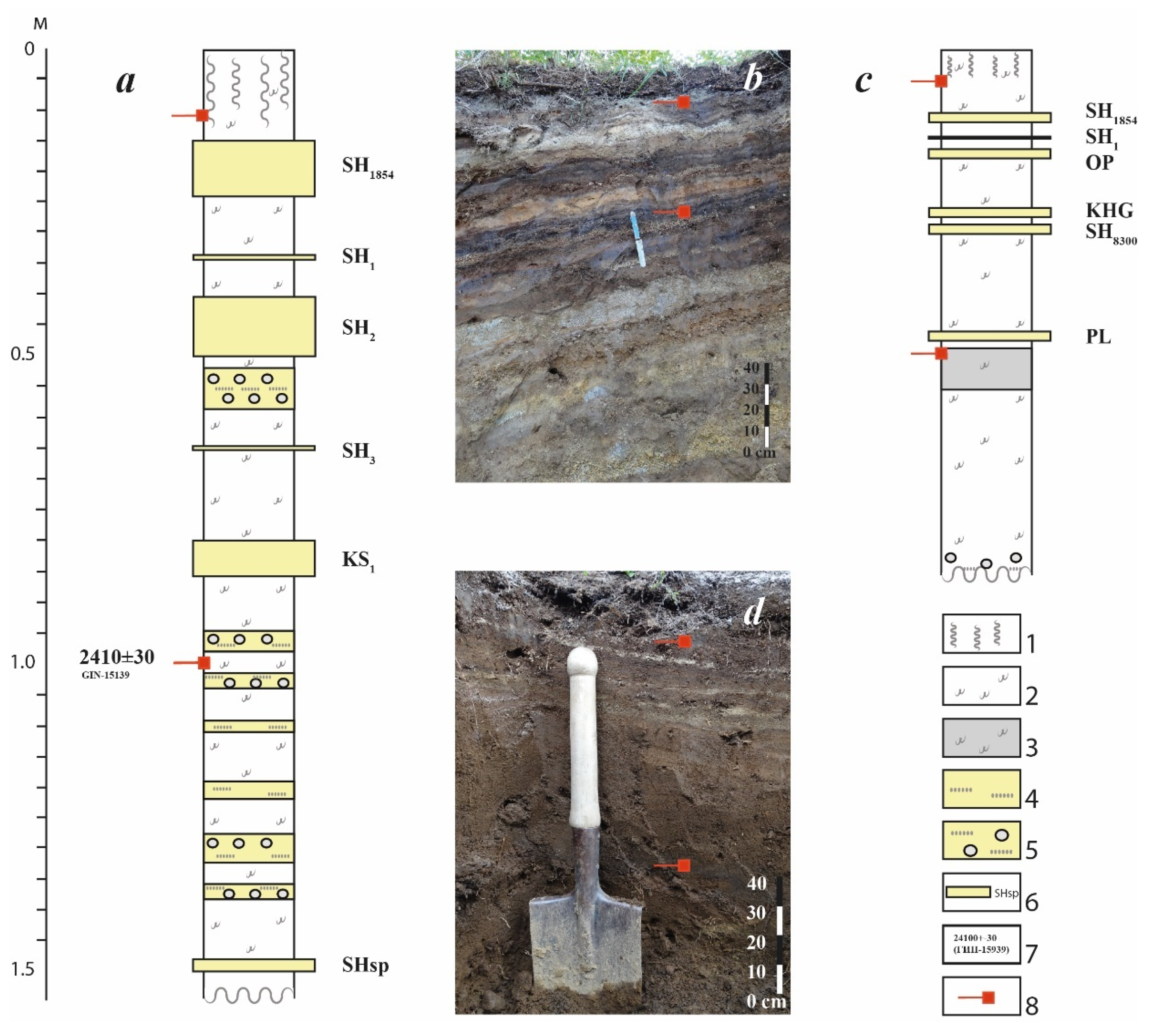
The first section is located at the northern foot of the Shiveluch volcano, 20 km from its peak (56°49′42″ N 161°19′28″ E). The SPC is formed on deposits of a large clastic avalanche dated 11.3 ka BP in a stone-birch forest [
14].
Figure 2a,b demonstrates clearly that, near the active volcano, where the maximum frequency of ashfalls is observed, the soil profile is saturated with volcanic material of different dimensions, from fine sand to gravel and lapilli pumice. A large amount of pyroclastic material also leads to an increase in the thickness of the entire section, with the soil interlayers occupying a subordinate position. Two horizons (modern and buried) of Aluandic Andosols Dystric stratified ochre volcanic soil (IUSS Working Group, 2015) were sampled to characterize the prokaryotic complex. The characteristics of the samples (Shiveluch) are presented in
Table 1.
The second section was plotted in the valley of the Tigil River, whose headwaters are located in the massif of extinct volcanoes of Alnay–Chashakonja (56°29′52″ N 159°31′03″ E). The SPC is formed on lava deposits of Quaternary period in the mountain tundra zone.
Figure 2c,d shows that the amount of volcanic ash in the section decreases considerably at a significant distance from active volcanoes. They noticeably decrease in thickness and are represented only by thin sands. The total thickness of the soil profile also decreases. At the same time, the thickness of organogenic soil horizons in the section increases considerably. The soil is characterized as poorly developed volcanic humus-ochric, Aluandic Andosols Dystric [
16]. Two soil samples (“Tigil”) were collected from the modern and buried horizons (
Table 1).
Description of the section of volcanic stratified ochre soil (Shiveluch).
A depth of 15 cm—turf and loose semi-decomposed forest litter; 10 cm—dirty-white, loose, coarse-grained sand with a single gravel (SH1854); 17 cm—dark brown soil horizon, loamy, almost structureless with light layers of unidentified ash; 12 cm—the horizon of marking volcanic ash, stratified, in the upper part light fawn fine-grained to siltstone, in the lower part dark fawn of coarser granulation (SH2); 25 cm—the soil horizon is brownish-gray, with a large number of organic residues of varying degrees of decomposition, lumpy structure, loose, intertwined with a dense network of roots. There is a marking layer of ash of light fawn color, fine-grained (SH3); up to 6 cm—marking layer of ash, ochreous siltstone (KS1); 60 cm—interlayer of ash horizons and buried soil (grayish-ochre and grayish-brown) lightly loamy, desalinated; 1–1.5 cm—bright bluish-gray fine-grained sand, hardened along the lower border.
Description of the section of volcanic humus-ochre soil (Tigil).
A depth of 3 cm—turf, 1 cm from the roof light gray siltstone with an admixture of fine-grained sand (unidentified ash); 9 cm—humus horizon, dark gray to brown, slightly structured, powdery, loose with stratified layers of black fine-grained sands of basalt composition; 0.2–1.2 cm—pinkish-light gray siltstone with an admixture of fine-grained sand (SH1); 2 cm—humus horizon, dark gray to brown, slightly structured, powdery, loose, with stones not rolled in diameter up to 0.5 cm; 0.5 cm—bright creamy white siltstone with biotite (OP); 6 cm—dark coffee, with an unclear structure, with stones and volcanic sand of different composition; 1–2 cm—fawn siltstone (KHG); 2 cm—dark coffee, with an unclear structure; up to 2 cm—pale gray fine-grained sand of the “salt and pepper” type with rusty fine grains (SH8300); 54 cm—grayish-ochreous, loamy, fragile lumpy–powdery structure. Decayed, with fragments up to 15 cm in diameter throughout the power. At a depth of approximately 50 cm, a dark coffee to black high-humus horizon.
Radiocarbon dating of buried soils was carried out at the Geological Institute of the Russian Academy of Sciences (Moscow) according to the standard soil methodology, which is discussed in detail in Pevzner, 2015 [
17]. Measurements were carried out on radiometric installations with a counting efficiency of approximately 50% in non-replaceable calibrated cuvettes with working volumes from 1.5 to 22.03 mL. E-5 and background samples were used as standards. The measurement of the age of one counting drug was carried out at least twice (one measurement lasts approximately a day), with an interval between them of 5–10 days on average. With a satisfactory score (the values of different measurements converge within the error), the average radiocarbon age of the sample was taken. The radiocarbon dates were calibrated to calendar age using Oxford University calibration software OxCalv4.4.2IntCal 20 [
18,
19]. The studied soil horizon of the volcanic stratified ochre soil at the foot of Shiveluch volcano was formed approximately 2.5 thousand years ago (calBP). The age of the volcanic hummocky-ochre soil in the Tigil River valley was estimated based on the stratigraphy of marker ashes. The age of the Sopok Plosky eruption, resulting in the deposition of marker ash PL, is estimated at 11,650 years calBP [
15]. Since the soil is deposited directly beneath the PL ash (
Figure 2c), its age is approximately 12 thousand years calBP. Soil microbial succession was initiated by humidification (soil water pressure was −3.2 kPa) and application of 0.6% biopolymer chitin suspension (0.3 g/5 g soil), as well as crude oil in an amount largely exceeding the MAC of 10% (1 mL/5 g soil). The soil weight was 5 g. Purified chitin was obtained from Sigma-Aldrich. Oil was obtained from the Azov–Kuban oil-and-gas-bearing basin (OGB, Kubanskaya area field, Well C-1,
Table 2 and
Table 3).
The use of two substrates (oil and chitin) is methodologically justified by different carbon-to-nitrogen ratios (C:N). This should lead to different degrees and speeds of substrate utilization and allows for assessment of the metabolic potential of the soil microbial community.
The following parameters were used as indicators of microbial decomposition of chitin and oil in soils: dynamics of bacterial numbers and biomass, assessment of metabolically active prokaryotic cells, presence or absence of functional genes characterizing the main metabolic reactions associated with N and C cycles in the system.
Epifluorescence microscopy using different fluorochromes—acridine orange, Sy3—was used to determine the total number and the number of metabolically active cells in the studied samples [
20]. The total number of prokaryotes was determined using acridine orange dye, which reacts with cell DNA. Soil samples were pre-treated with Bandel in Sonopuls HD 2070 (Germany) for 2 min and 50% power for better cell desorption. Assessment of diversity and abundance of the metabolically active representatives of individual phylogenetic groups of microorganisms in the soils was carried out by in situ hybridization with rRNA-specific fluorescent-labeled oligonucleotide probes (fluorescent in situ hybridization, FISH) [
21,
22,
23,
24]. In this study, we used an array of probes specific for representatives of Archaea and Bacteria domains, as well as representatives of several phylogenetic groups (
Acidobacteria,
Actinobacteria,
Bacteroidetes,
Firmicutes,
α-Proteobacteria,
β-Proteobacteria,
γ-Proteobacteria,
δ-Proteobacteria,
Planctomyces,
Verrucomicrobia,
Euryarchaeota,
Crenoarchaeota, and
Thaumarchaeota). This allowed for analysis of the bacterial and archaeal community of the soils under study. Hybridization temperature regime and hybridization and washing buffers were optimized for each bacterial phylogenetic group [
25]. Prepared samples were observed using PrimoStar microscope (Zeiss, Germany) with a light filter (AmScope, Irvine, CA, USA). Prokaryotic biomass (B) was calculated according to Formula (1):
where N is the number of prokaryotic cells in 1 g of soil sample of bacteria in 1 g of sample, and the biomass of dry matter for 1 bacterial cell with volume of 0.1 µm
3 is 2 × 10–14 g [
20]. The specific gravity of microorganisms was taken as 1 g/cm
3; the water content of the cells was 80%.
Total DNA extraction followed by sequencing was performed for all soil sample types (buried and modern). Metagenomic DNA was extracted from a 0.5 g soil sample using a specialized DNeasy Power Lyzer Power Soil Kit (MoBioLaboratories, Inc., Carlsbad, CA, USA) according to the manufacturer’s protocol. The nucleotide sequences of V3-V4 region of variable sections of 16S rRNA gene from metagenomic DNA samples were determined by high-throughput sequencing [
26]. Sequencing was performed on Illumina Miseq sequencer by Evrogen Research Company (Moscow, Russia) with 39 h read time and 8 mln pair-end reads. Then, a file with forward and reverse reads from both ends of DNA (301 cycles from each end of the fragments) was generated, which was a textual description of the primary structure of linear macromolecules in the form of a monomer sequence. Sequencing data processing (FASTQ files) was performed using the DADA2 package functions for the programming language R.
The software tools were used to perform the following: check sequencing quality and create a library of sequences; generate OTUs (de novo OTU picking) with 97% similarity threshold; remove singletons (“singletons” are OTUs containing only one sequence) and sequences belonging to plant chloroplasts; remove chimeras.
In addition, random samples were created from each table: 2500 from initial coverage, 100,000 from final coverage, 2500 step, with the number of replicates—5. For each sample, Sørensen–Dyce and Bray–Curtis pairwise distance matrices were calculated. We calculated Shannon, Simpson, Pielu, and Chao alpha diversity scores using functions from the scikit-bio package for the Python programming language for each sample. Values for the same depth grouped by indicator and sample were averaged.
Sequences were visualized using Krona [
27].
The abundance of ribosomal genes in bacteria and archaea was estimated by real-time polymerase chain reaction (RT-PCR) [
28]. The reaction was performed in a DTLite4 DNA-Technology amplifier. The reaction mixture was prepared from Super Mix Eva Green (Carlsbad, CA, USA). The instrument was calibrated using the dependence of fluorescence intensity on the logarithm of DNA concentrations of standard solutions. Solutions of cloned fragments of ribosomal operon from
Escherichia coli strain K12 were used as standards for 16S rRNA gene concentration for bacteria, and those of
Halobacterium salinarum strain FG-07 were used for archaea. For each experimental variant (sample), the reactions were performed in three replicates and with negative control (sample without DNA matrix). Data processing was performed using the Real time_PCR software package. All DNA-specific primers used in the work, as well as reaction protocols (amplification temperature profile), are described in
Table 4.
To better characterize the nitrogen cycle in the studied microbial communities, the genes responsible for the nitrification (ammonium oxidation) process were also amplified:
amoA (AOB) for bacteria [
29] and
amoA (AOA) for archaea [
30]. The reactions used a series of dilutions of a known number of copies of linearized plasmid (PCR4-TOPO, Aberdeen, Scotland) containing the
amoA gene
Nitrosospira multiformis ATCC25196. The following reaction protocol was used to amplify ammonium oxidizing bacteria: 95 °C 10 min; 94 °C 45 s, 58 °C 45 s, 72 °C 45 s—39 cycles; 95 °C 15 s, 60° C 30 s, 95 °C 15 s. The protocol for amplification of ammonium oxidizing archaea was: 94 °C 45 s, 55 °C 45 s, 72 °C 45 s—39 cycles; 95 °C 15 s, 60 °C 30 s, 95 °C 15 s.
Amplification of the genes responsible for the denitrification process of
nirK [
31] (
Table 4) was carried out according to the following scheme: 95 °C 15 min; 95 °C 15 s, 60 °C 30 s, 72 °C 30 s, 80 °C 15 s—40 cycles; 95 °C 15 s, 72 °C 30 s. The
nifH gene was amplified to determine the presence of the gene responsible for the ability to supply available nitrogen to the community (nitrogenase activity) [
32]; the amplification temperature profile is shown in
Table 4. DNA solutions extracted from cultures of
Bacillus cereus and
Azotobacter chroococcum were used as gene concentration standards for
nirK gene and
nifH gene copies, respectively.
The primer system described in Smith et al., 2013 [
33] was used to identify functional genes indicative of n-alkane degradation processes.
Streptomyces violaceus (
sec. Roseus: ser. Roseoviolaceus) culture capable of using hydrocarbons as a single food source (toluene) was used as a standard for detection of
alkB gene copies. The following protocol for the amplification was used: 1 cycle: 94.0 °C—5 min; 30 cycles: 94.0 °C—1 min; 60 °C—1 min; 72 °C—1 min; 1 cycle: 72 °C—3 min; 1 cycle—4 °C—cooling [
34] (
Table 4).
3. Results
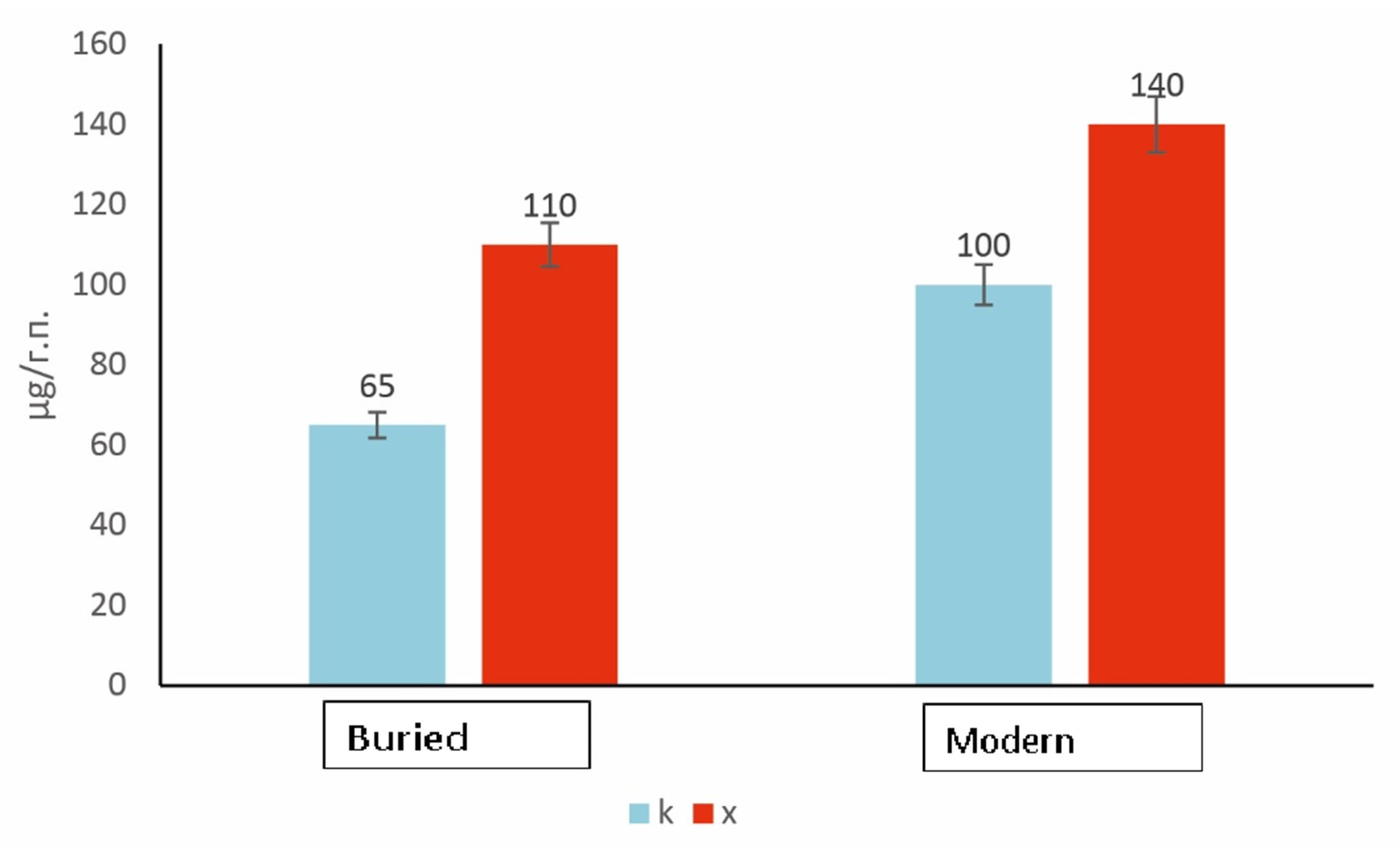
The biomass of prokaryotic communities in the modern and buried horizons of the studied soils was determined by the direct accounting method. A greater prokaryotic biomass in the modern horizons than in the buried ones was observed for the studied samples (
Figure 3). The addition of chitin leads to an increase in biomass in both buried and modern soil samples. However, the relative response of the microbial community of the buried soils to substrate application is significantly higher than that of modern soils. Similar patterns were noted by the authors earlier for the microbial hydrolytic community of buried paleochannels compared to modern chestnut soils [
20]. When polysaccharide was applied to chestnut paleosoils, the proportion of metabolically active prokaryotic cells reached 50% of the total detectable prokaryotic community, and the response to the substrate application was more intense compared to modern soils. Similar results were obtained for the structure of the prokaryotic complex of Antarctic permafrost [
35].
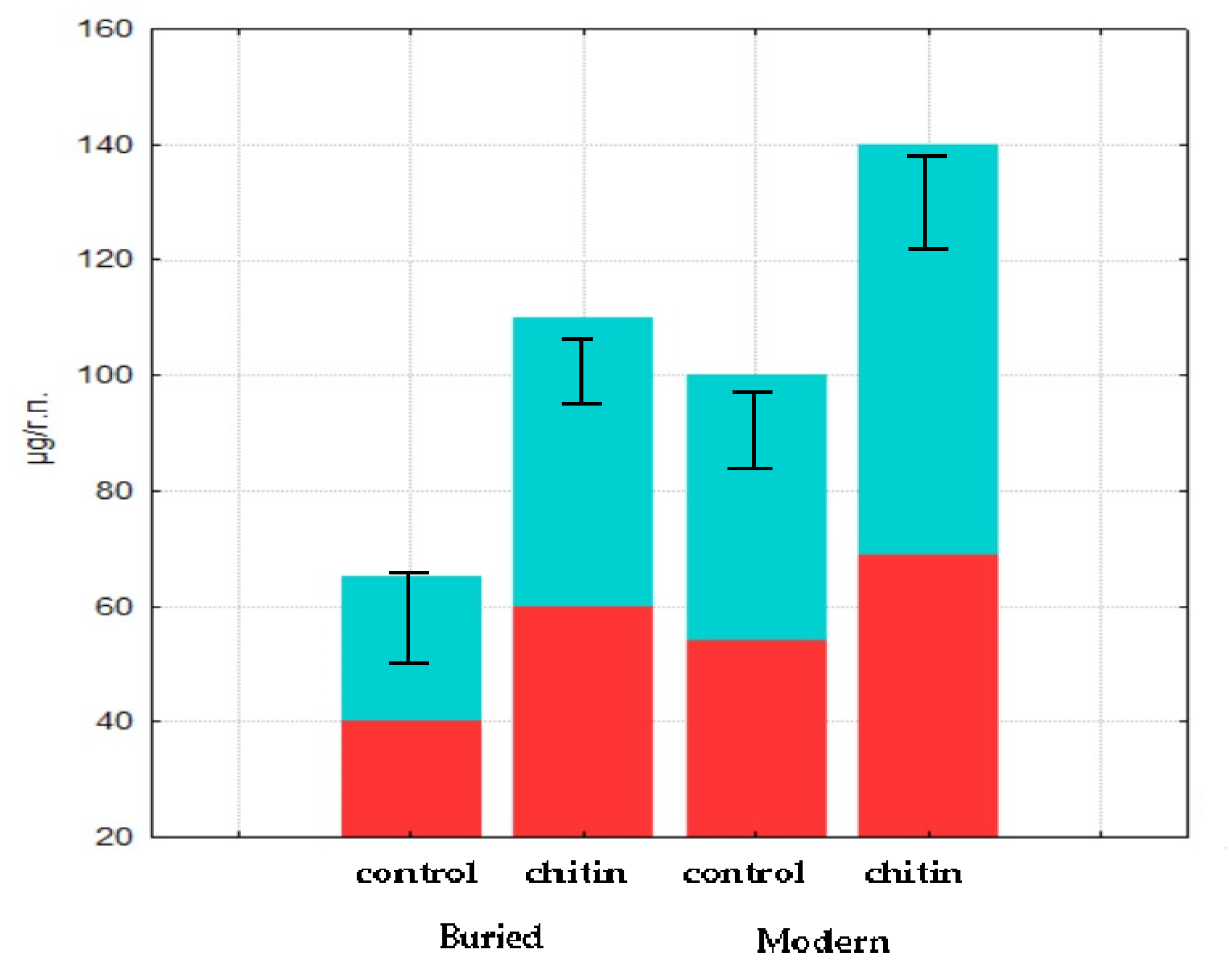
By the tenth day of the succession, the biomass of metabolically active representatives of the prokaryotic component of the sample collected from the buried horizon of volcanic stratified ochre soil moistened with water was 40 µg/g.p. (of which, 35 µg/g.p. was the bacterial component). An increase in biomass was observed in the microcosm following the addition of chitin compared to the control sample; the biomass value was 60 µg/g.p. (of which, 44 µg/g.p. was the bacterial component) (
Figure 4).
By the thirtieth day of the succession, the metabolically active biomass of the sample of the buried horizon of volcanic stratified ochre soil moistened with water was 57 µg/g.p. (of which, 48 µg/g.p. was the bacterial component). The prokaryotic community biomass was 32 µg/g.p. (of which, 29 µg/g.p. was the bacterial component) in the variant with chitin by the thirtieth day of the succession. Thus, our results confirm the data obtained earlier for samples of chestnut modern and paleo-soil soils. The proportion of metabolically active prokaryotic components in the buried soil horizons reached 50% of all of the detected biomass when the microbial community was reactivated by the resource.
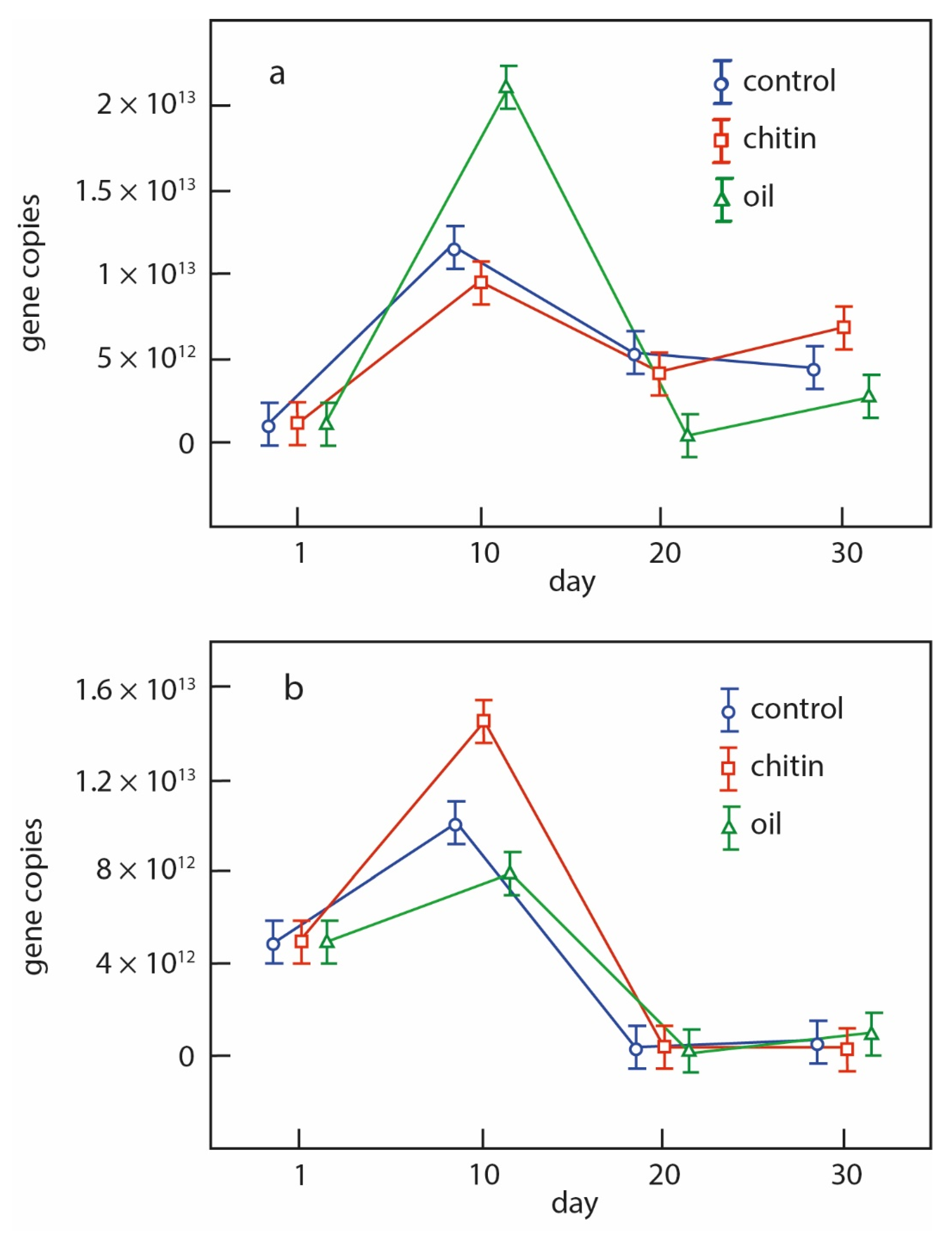
The number of copies of bacterial rRNA genes in the studied soils was determined using a real-time polymerase chain reaction. The gene copy number in the soils was measured dynamically during the succession. The variants with oil and chitin addition, as well as the control, were considered. The values of bacterial genes in the buried horizon were comparable to those of modern soils, reaching n × 10
13 genes (
Figure 5). The greatest response to the application of the resource was noted in the variants with chitin in the buried sample. This coincides with the results of total and metabolically active prokaryotic biomass in the buried soil samples with biopolymers, indicating a possible high metabolic potential of the studied soils.
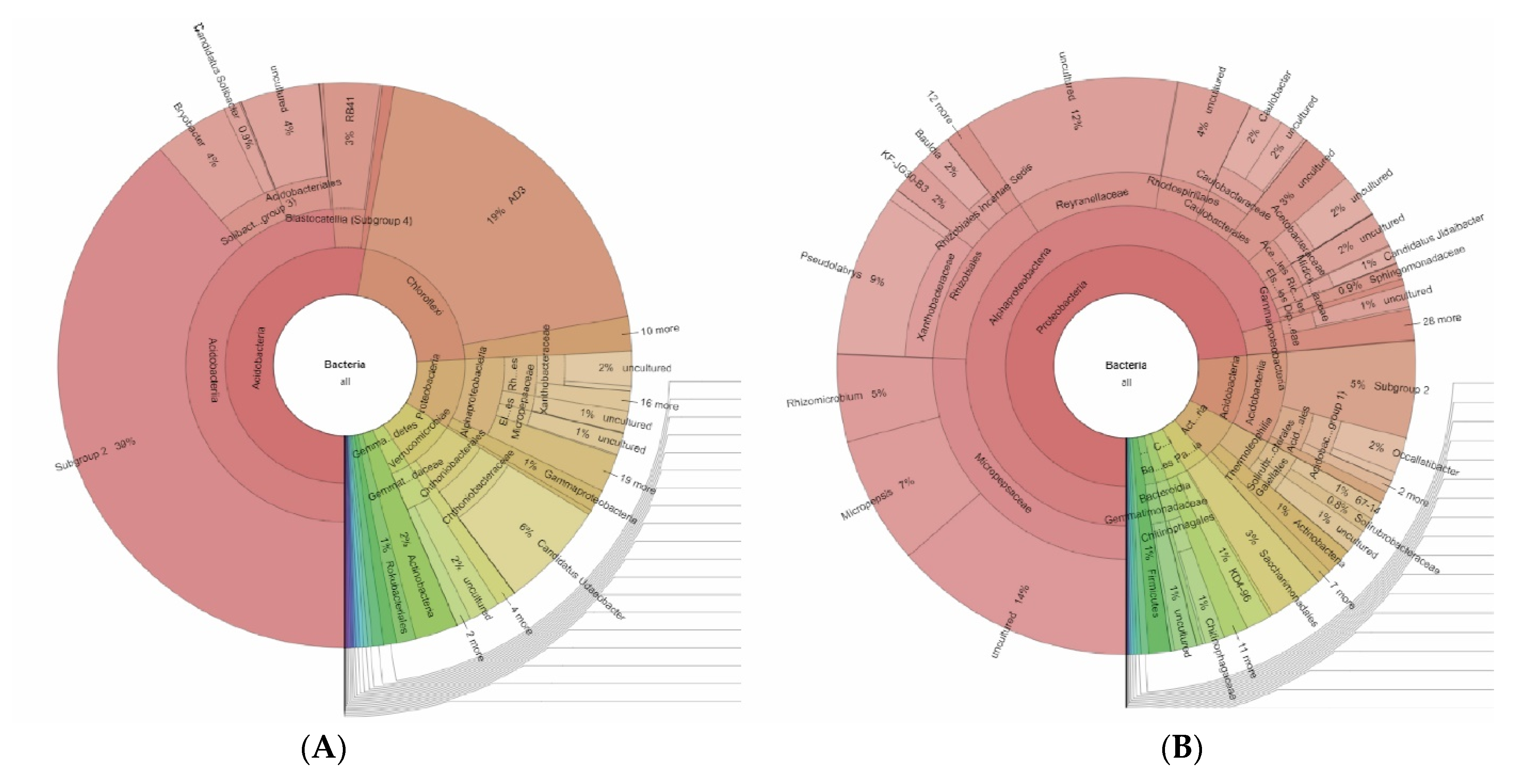
The phylogenetic structure of the prokaryotic communities of two studied soils—volcanic stratified ochre and volcanic humus-ochre—was determined by the FISH method and metagenomic analysis. Significant differences in the structure of the bacterial complex of modern and buried soils were revealed.
The microbial community structure of the studied samples was determined using Krona software (
Figure 6). For the modern horizon, it was determined that phylum
Proteobacteria was dominant (74%); representatives of phyla
Actinobacteria and
Acidobacteria (14% together) were included in the moderate abundance groups, and the minor group included
Gemmatimonadetes,
Chloroflexi, and
Verrucomicrobia (8% together) in this community (
Figure 6B). Representatives of the other domain groups are only marginally represented, totaling less than 1%.
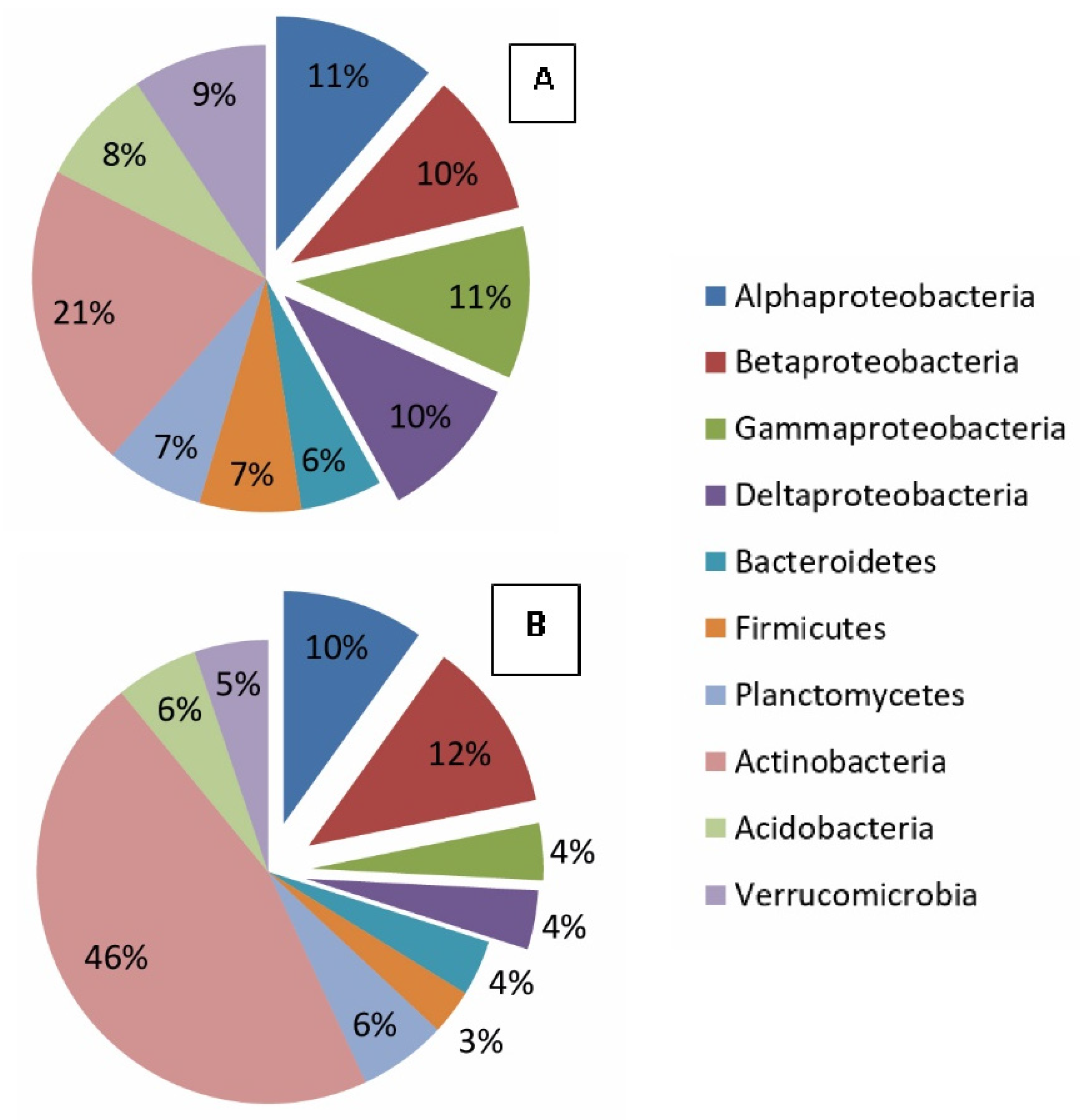
The biomass of metabolically active Proteobacteria phylum was 17 µg/g of the sample. Among Proteobacteria, representatives of class Alfaproteobacteria and orders (families) Ellin329, Rhodospirillales (Rhodospirillaceae, Acetobacteraceae), Rhizobiales (Hyphomicrobiaceae, Bradyrhizobiaceae), Caulobacterales (Caulobacteraceae), Rhodospirillales (Acetobacteraceae), and Sphingomonadales (Erythrobacteraceae) dominated. The classes Betaproteobacteria and Gammaproteobacteria are less represented.
Saccharibacteria (candidate phylum TM7) were detected in significant numbers.
For the Acidobacteria division (biomass of metabolically active representatives was 10 µg/g of the sample), the most characteristic orders were Acidobacteriales (Acidobacteriaceae, Koribacteraceae) and Solibacterales.
The Actinobacteria group having a biomass of metabolically active representatives in the amount of 8 µg/g of the sample was mainly represented by the class Thermoleophilia and its orders (Solirubrobacterales—family Patulibacteraceae, Gaiellales—family Gaiellaceae). The Acidimicrobiales (EB1017) and Actinomycetales (Nocardioidaceae) orders were also identified among Actinobacteria representatives.
Among the Chloroflexi, the classes Ellin6529 and Anaerolineae (GCA004) were identified.
Members of the family Chitinophagaceae of the order Bacteroidetes were detected in significant numbers.
Clostridiales (Clostridiaceae) was the most common among Firmicutes, and N1423WL among Gemmatimonadetes. The phylum Nitrospirae was represented by the class Nitrospira (order Nitrospirales, family Nitrospiraceae).
The archaeal community complex of the modern volcanic stratified ochre soil horizon was dominated by Crenarchaeota (NRJP) whereas Cenarchaeales was only minor. Among the Euryarchaeota, representatives of the class Thermoplasmata (order E2, family Methanomassilii coccaceae) were identified.
For the buried horizon (
Figure 6A), the phylum
Acidobacteria was dominant (51.8%), with subdominants being
Chloroflexi (21%),
Proteobacteria (9%), and
Verrucomicrobia (7%). Representatives of
Actinobacteria and
Gemmatimonadetes phyla held minor positions in the structure of the studied community (4% together). The conditions for the existence of microorganisms change greatly during the transition from modern soils to their buried analogues. First of all, this affects the content of the resource available to microbes. Paleosols are characterized by a lower content of organic matter, which entails a change in the structure of the microbial community of soils. Thus, earlier, the authors found that, in the microbial complex of buried chestnut soils, in comparison with their modern counterparts, the number of spore-forming actinobacteria increases noticeably. In addition, with the introduction of chitin, this pattern was significantly enhanced [
20].
The biomass of metabolically active Acidobacteria representatives in paleochannels reached 20 µg/g of the sample. Acidobacteria were represented by classes DA052 (order Ellin6513), Chloracidobacteria, Chloracidobacteria (o__RB41), orders Acidobacteriales (Acidobacteriaceae, Koribacteraceae), and Solibacteres (Solibacterales).
The following representatives were found in large numbers: Dormibacteraeota (formerly known as AD3) from the phylum Chloroflexi typical for deep soil horizons with a reduced organic carbon content; also active hydrolytes from division Verrucomicrobia (class Spartobacteria, order Chthoniobacterales, family Chthoniobacteraceae); and, finally, class Methylacidiphilae (S-BQ2-57) from Verrucomicrobia.
Proteobacteria with a biomass of metabolically active representatives in the amount of 8 µg/g of the sample, mainly Alphaproteobacteria (order Ellin329), as well as orders (Rhodospirillales, family Rhodospirillaceae) and Rhizobiales (Hyphomicrobiaceae), were detected. Among Betaproteobacteria, the order Ellin 6067 was detected.
Actinobacteria were represented by the classes Acidimicrobia (Acidimicrobiales, family EB1017) and Thermoleophilia (order Gaiellales, family Gaiellaceae).
Gemmatimonadetes (Gemmatimonadetes, order N1423WL, family c__Gemm-1), Nitrospirae (Nitrospira, order Nitrospirales, family 0319-6A21), and Bacteroidetes (Sphingobacteriia order Sphingobacteriales) were identified.
The archaeal community complex of the buried volcanic stratified ochre soil horizon was dominated by Euryarchaeota, mainly Methanomicrobia (Methanocellale). The representatives of pGrfC26 were typical for Crenarchaeota in the buried horizon. Thaumarchaeota were represented by the order Cenarchaeales (SAGMA-X).
Biodiversity indices (i.e., alpha-diversity) were calculated for each sample using the Python scikit-bio software package. For the modern horizon, the following indices were calculated: the Chao1 index was 669.5, Pielu index (“alignment” of the community) = 0.6, Shannon index = 5.65, Simpson index = 0.95. For the buried horizon, the Chao index was 448.3, Pielu index = 0.59, Shannon index = 5.26, Simpson index = 0.91.
The addition of chitin to the studied samples leads to a change in the structure of the metabolically active prokaryotic component of the microbial community. The number and biomass of actinobacteria increase, whereas the number of proteobacteria decreases (
Figure 7). More evidence of the ability of prokaryotes to synthesize a variety of instances of depolymerization was obtained [
36,
37]. A high chitinolytic activity of
Actinobacteria is known from the literature [
38]. Actinomycete complexes have a high chitinolytic activity in various soils [
39,
40] The intensity of prokaryotes-destroyers, as well as that of all microorganisms, strongly depends on environmental factors, including temperature, moisture, redox conditions, etc.
In parallel, the functional activity of the prokaryotic community was assessed in the studied soil samples by detecting the presence of nitrogen cycle genes in the system: nifH—nitrogenase activity, amoA (AOB)—ammonium oxidizing bacteria monooxygenase activity, amoA (AOA)—ammonium oxidizing archaea monooxygenase activity, and nirK—nitrate reductase activity.
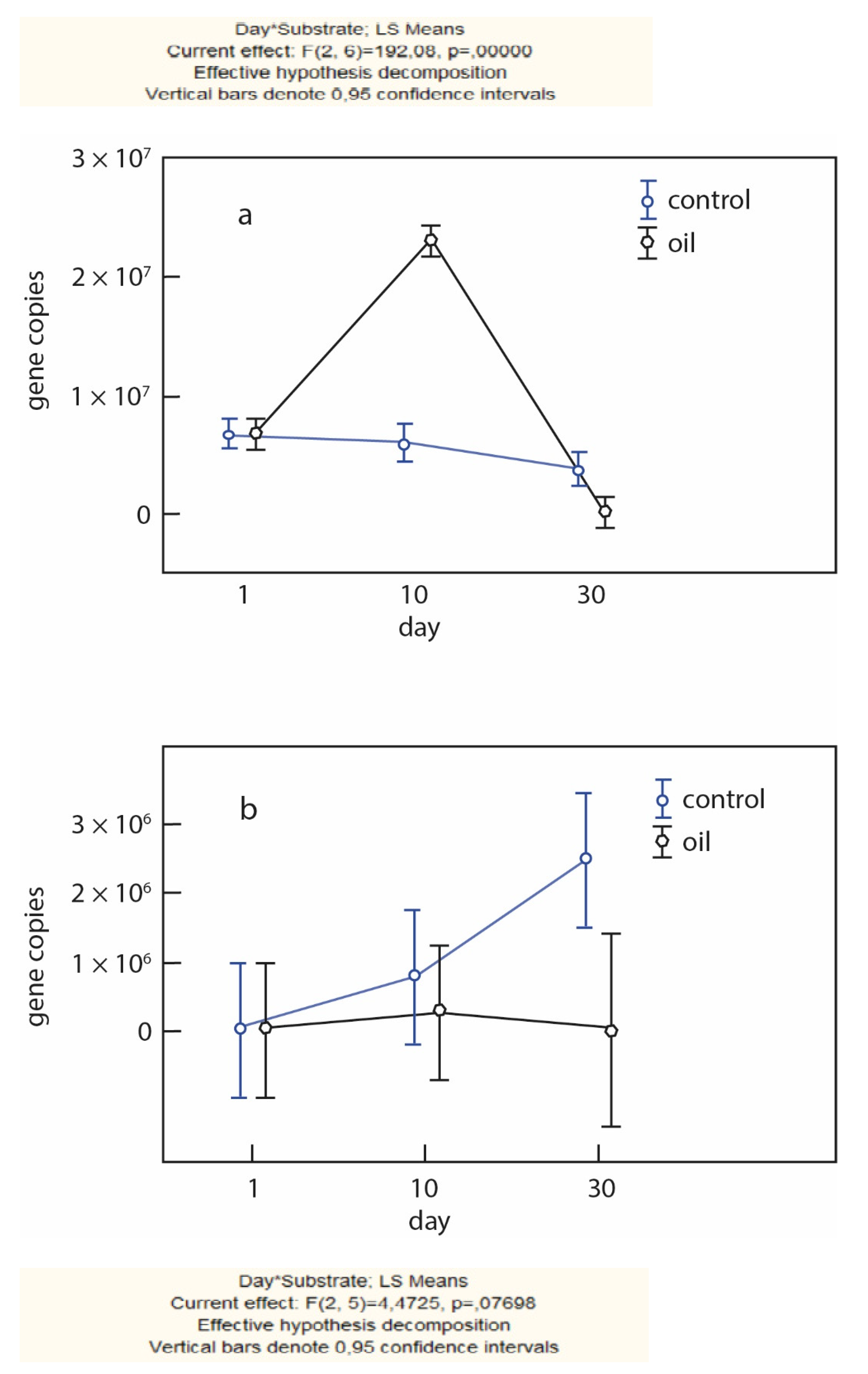
As expected, the highest values for the functional gene nifH in the studied soils were observed in the control variant without the addition of nitrogen-containing polymers. Moreover, the maximum values were observed for the modern horizon and reached 1.54 × 107 gene copies/g.p. by the 30th day of succession. For the buried horizon, despite an overall increase in gene concentration by the 30th day, its maximum values were observed at the beginning of succession (2.5 × 105 gene copies/g.p.). The values of the gene content in the modern horizon were two orders of magnitude higher than in the buried. For all soils, there was a tendency in the lower content of the nifH gene when introducing substrates into the soil. This feature can be explained by the presence of nitrogen forms available to microorganisms in the soil with polymers (especially chitin), which leads to an intensification of mineralization processes and a decrease in nitrogenase activity. It is important to note the presence, albeit insignificant, of the nitrogenase gene in the deeper layers of the soil, which indicates the possible potential of microbial communities living there.
An amplification of the key nitrification gene was also carried out. For volcanic soils (both modern and buried), the presence of ammonium oxidizing bacterial and archaeal genes was detected (
Figure 8 and
Figure 9).
An increase in the gene concentration in the buried samples after 30 days of succession in the control variants and the variant with oil was noted, indicating a reactivation of nitrifiers by the end of succession (
Figure 9a). The dynamics of the
amoA gene in the modern horizon of the volcanic humus-ochre soil shows its increased concentration in the bacterial complex in the variants with oil (2.3 × 10
7 DNA copies/g.p.) by the 10th day of succession, whereas a low content of the gene is noted in the control. In the archaeal complex, there is an increase in the gene concentration in the control variant after 30 days (2.5 × 10
6 DNA copies/g.p.), whereas the indicators remain very low in the variant with oil.
Based on the data obtained, it can be concluded that archaea play a leading role in the nitrification process in soils without oil (undisturbed community). The bacterial complex is capable of nitrification with a high oil contamination of the soil, and its role also increases in the lower layers of the soil profile. The contribution of archaea to the nitrification process decreases with the introduction of oil, and also decreases with depth.
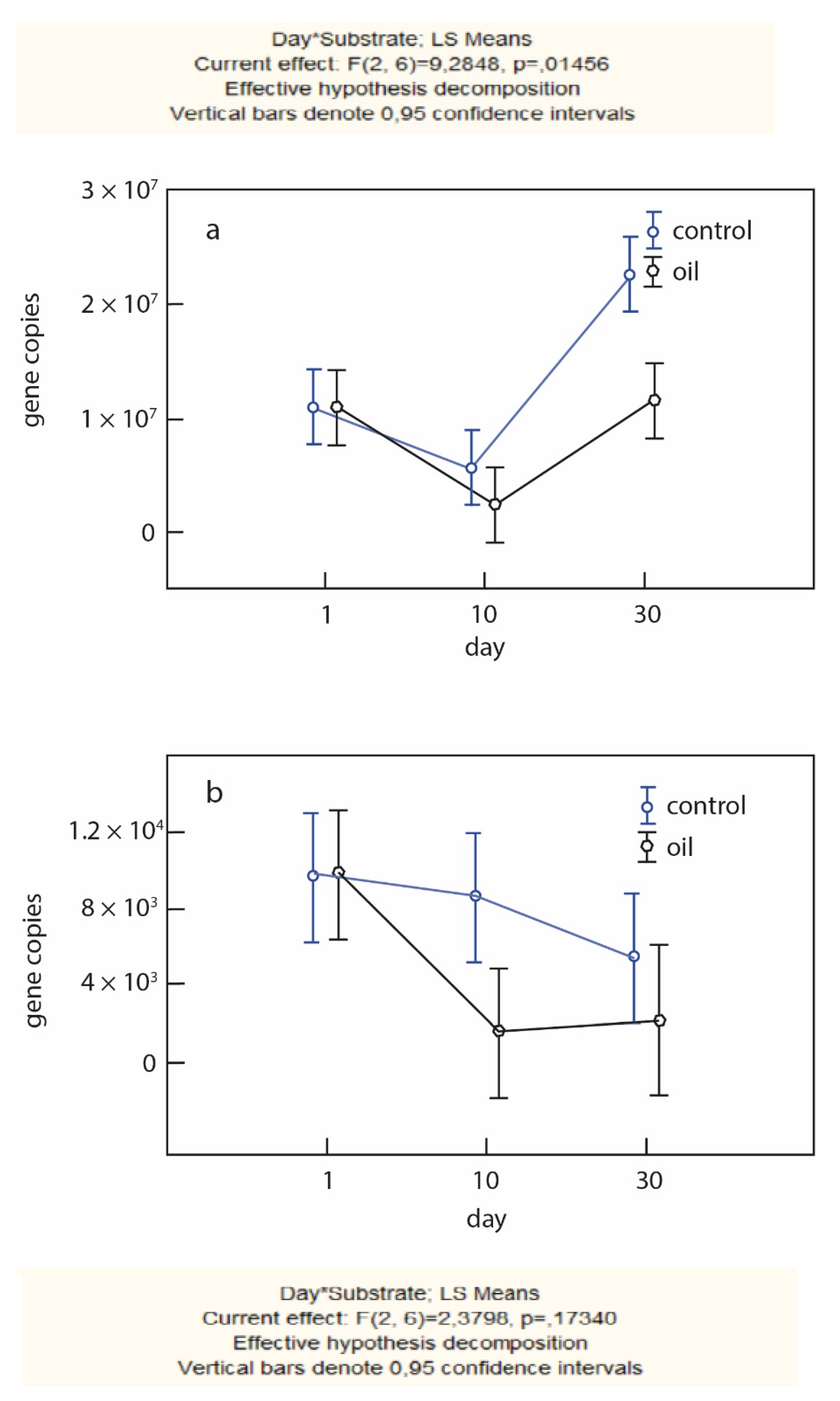
Copies of the nir K gene, a molecular marker of denitrification, were detected in volcanic humus-ochre soil samples in all variants of the experiment (control, oil) during succession. The absolute values were the highest in the modern horizon (1.25 × 107 DNA copies/g p), whereas, in the buried horizon, they were two orders of magnitude lower (1.2 × 105 DNA copies/g p).
In all horizons, there was a reduced content of the gene in the variant with the introduction of oil. The greatest negative impact of oil was noted in the buried horizon.
The presence of alkB, a gene responsible for the activity of the n-alkanoxygenase enzyme, was assessed for the volcanic humus-ochre soil system. The alkB gene was detected in all variants; its absolute concentration was maximal in the modern horizon for a variant with oil on the 10th day of succession (2.5 × 104 DNA copies/g.p.). Thereafter, it decreased in the oil and control variants, and, after 30 days of succession, was 5 × 105 DNA copies/g.p. and 1.3 × 106 DNA copies/g.p., respectively.
There is a notable trend toward an increasing gene concentration during succession for all variants in the buried horizon. Absolute values of the gene content in buried soil by 30 days are 4.8 × 104 DNA/g.p for the control variant and 1.8 × 104 DNA copies/g.p for a variant with oil. Similar dependencies were found when considering the activity of hydrolytic (chitinolytic) microbial complexes of modern and buried soils.
4. Discussion
The work complements the previously obtained data from studies of relict microbial communities, and also reveals previously unknown features of such microbial complexes. Previously, studies of ancient communities were limited due to the imperfection of the methodological base. The applied complex of methods of classical microbiology and new molecular biological methods allows for the most complete characterization of the metabolically active prokaryotic community of buried soils. Buried soils, apparently, for the first time, became the object of a study on metagenomics. The analysis of the data of the high-performance sequencing of the 16S rRNA gene and the subsequent restoration of the functional genetic complex allowed us to identify the main dominants of the hydrolytic community of ancient soils. Presumably, the dominants of ancient hydrolytic communities may have higher activity compared to their collectible counterparts. The revealed increase in the potential activity of communities as the degree of “conservation” of the community increases opens up opportunities for the biotechnological use of strains isolated from being buried. The conducted research, apparently, is one of the most comprehensive and complete works devoted to the ancient microbial complex.
To summarize, the prokaryotic heterotrophic mesophilic community has been successfully studied in volcanic soil samples from Central Kamchatka. Paleolithic soils of different ages and their modern analogues were investigated. The age of the buried horizon of volcanic stratified ochre soil was 2500 years old and the horizon of volcanic humus-ochre soil was approximately 11,300 years old. It was found that the biomass of metabolically active prokaryotes of volcanic layered-ochre soil reached 54 µg/g in the control sample and 69 µg/g of the sample in the variant with the addition of chitin, which is approximately 40% of all detected prokaryote biomass. In paleovolcanic soil, the biomass indices of metabolically active prokaryotes were 40 µg/g and 60 µg/g, respectively. The addition of the chitin biopolymer to the paleovolcanic soil caused the indicators of the metabolically active prokaryotes share to reach 50% of the detected prokaryote biomass. Similar patterns were noted by the authors earlier for the microbial hydrolytic community of buried paleochannels compared to modern chestnut soils [
20]. When polysaccharide was applied to chestnut paleosoils, the proportion of metabolically active prokaryotic cells reached 50% of the total detectable prokaryotic community, and the response to substrate application was more intense compared to modern soils.
The proportion of archaea in the metabolically active prokaryotic complex ranged from 20% to 30% and increased in variants with resource addition, which is consistent with the results of other researchers [
4].
The features and differences in the phylogenetic structure of the prokaryotic metabolically active component of modern and buried volcanic stratified ochre soil were established by molecular biological methods (metagenomic analysis, FISH method). Proteobacteria (74%), Acidobacteria, and Actinobacteria (14% combined) were dominant in modern soil phylum; Acidobacteria was dominant (51.8%), and Chloroflexi (21%) and Proteobacteria (9%) were among subdominants in buried soil phylum. Obviously, such a feature of the structure of the complex is associated with a change in the resources present in the environment and used by microorganisms. The total diversity of the studied microbiomes (alpha diversity) differs slightly in buried and modern soils. The Shannon index does not differ significantly in buried and modern soils. At the same time, the amount of OTE (species richness) in modern soils is higher than in buried ones.
The introduction of an additional substrate (chitin) into the studied soil samples caused a change in the structure of the metabolically active prokaryotic component of the microbial community. This was especially noticeable for samples of buried soils. The number and biomass of
Actinobacteria doubled in the bacterial hydrolytic complex of stratified ochre paleosoils compared to its modern counterpart. The development of actinobacteria on such an inaccessible polysaccharide, nitrogen-containing biopolymer as chitin has been repeatedly noted by the authors [
7,
20]. The functional role of mycelial actinobacteria in the metabolism of chitin consists, on the one hand, of the active decomposition of this biopolymer, and, on the other hand, of the regulation of microbial hydrolytic complex activity through the production of biologically active regulatory metabolites, which occurs in a wide range of environmental parameters (moisture, temperature, organic matter, successional time).
The introduction of the substrate into both buried and modern soil reduced the content of nifH genes. This feature can be explained by the presence of nitrogen forms available to microorganisms in the soil with polymers (especially chitin), which leads to an intensification of mineralization processes and a decrease in nitrogenase activity. It is important to note the presence, albeit insignificant, of the nifH gene in the deeper layers of the soil, which indicates the possible potential of microbial communities living there.
An interesting feature is noted when considering the functional genes responsible for the nitrification process. For uncontaminated territories, the archaeal component of the prokaryotic community of soils plays a significant role in the process of ammonium oxidation. Similar results were obtained in the works of other authors, which provide evidence that archaea play a leading role in the oxidation of ammonium in soils [
41,
42,
43]. In soils without oil, archaea play a leading role in the nitrification process. The bacterial complex is capable of nitrification with a high oil contamination of the soil, and its role also increases in the lower layers of the soil profile. The contribution of archaea to the nitrification process decreases with the introduction of oil, and also decreases with depth.
The identified patterns indicate a possible high metabolic potential of the studied soils.


 ,
, 









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}