
Spatial Distribution Pattern and Genetic Diversity of Quercus wutaishanica Mayr Population in Loess Plateau of China
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
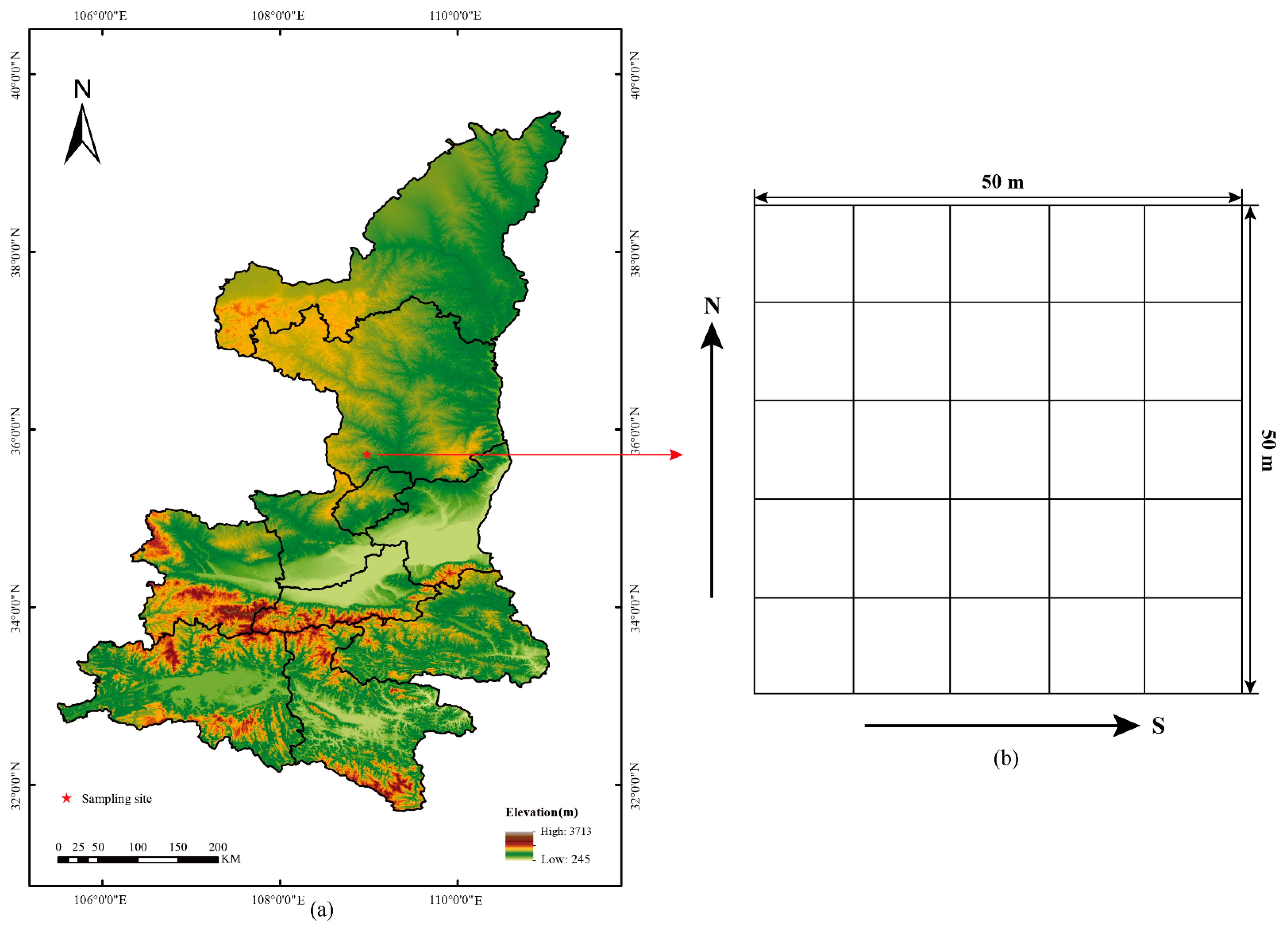
2.1. Study Area
2.2. Data Collection
2.3. DNA Extraction and SSR Analysis
2.4. Point Pattern Analysis and Null Model
3. Results
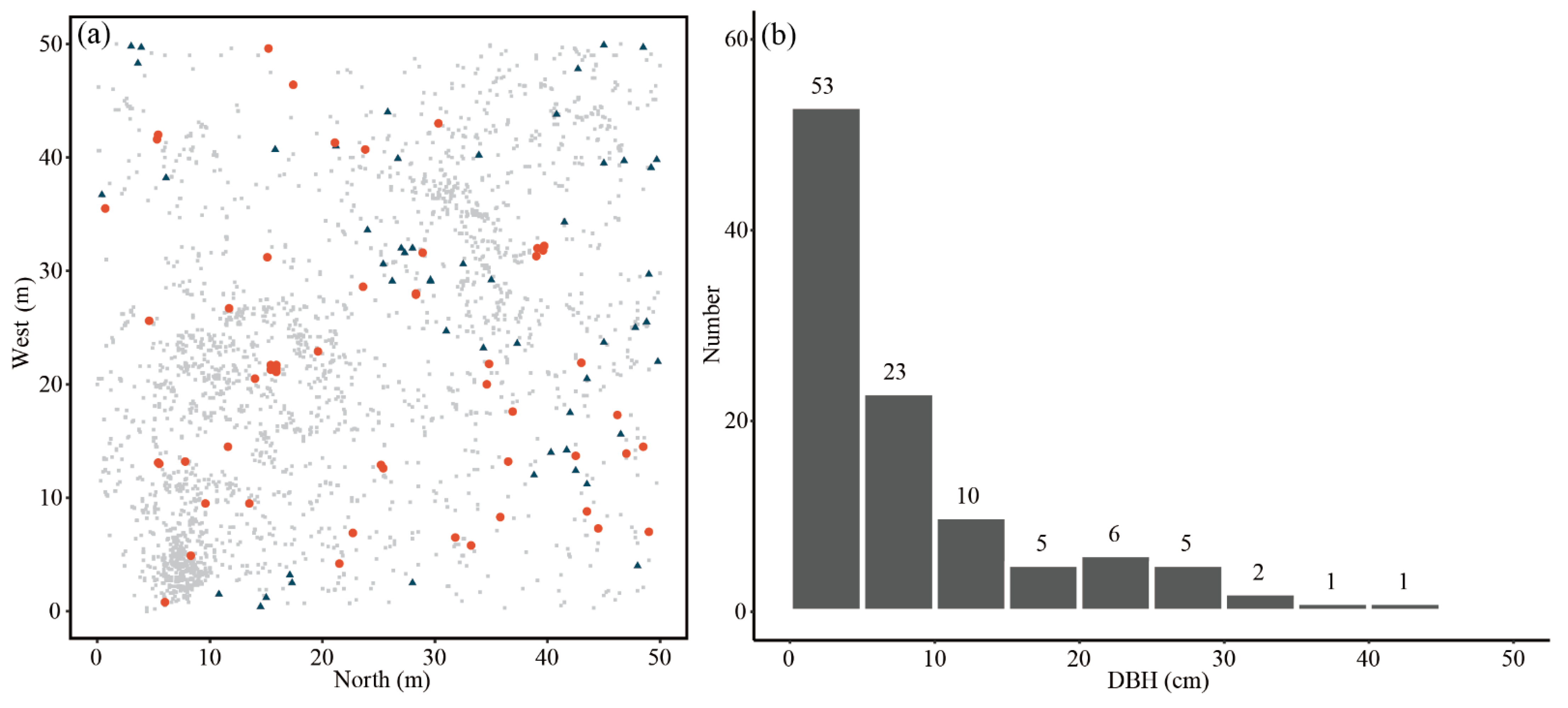
3.1. Individual Distribution and Population Structure
3.2. Genetic Diversity
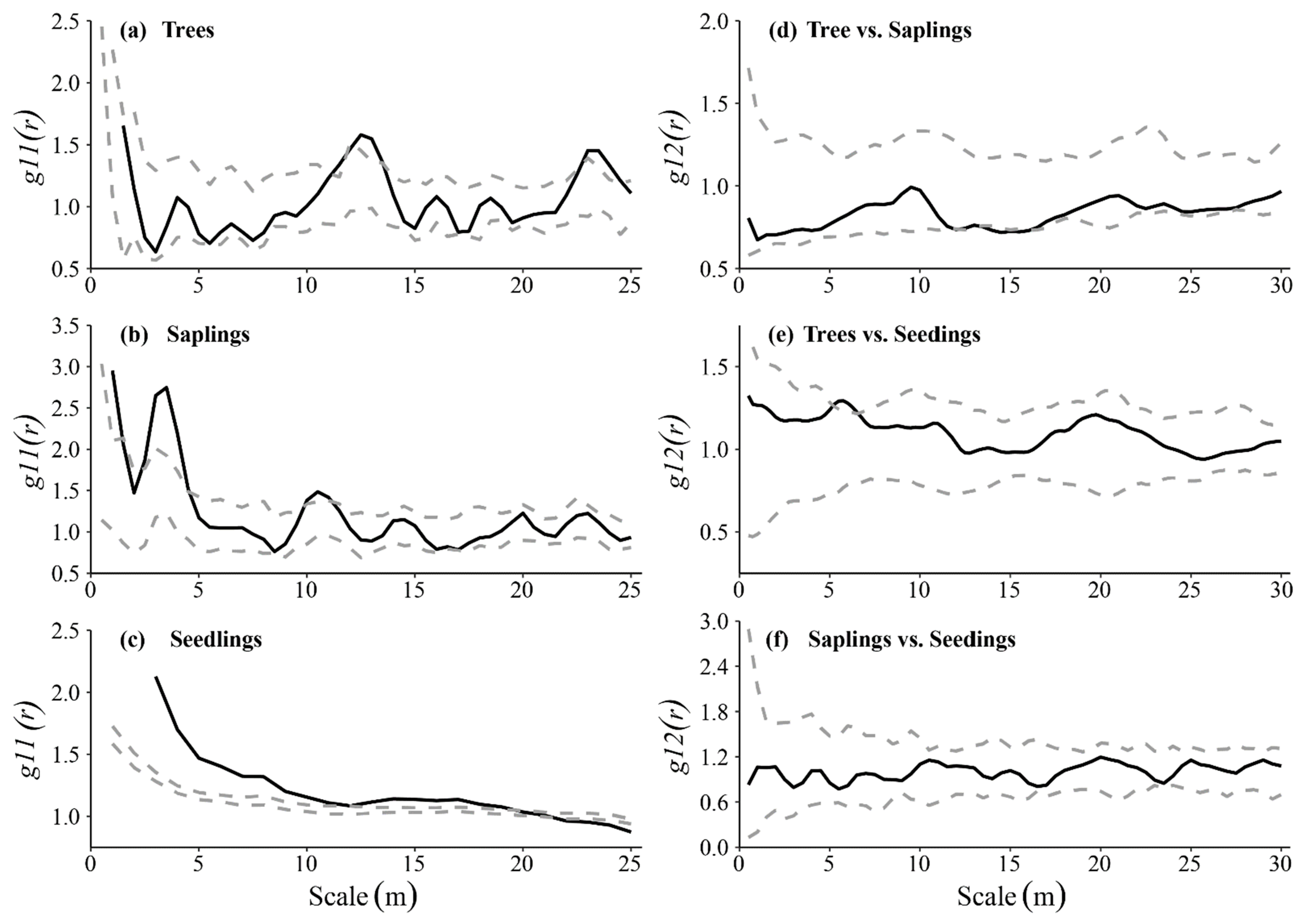
3.3. Spatial Distributions Patterns and Intraspecific Association
4. Discussion
4.1. Diameter Classes Structure of Q. wutaishanica
4.2. Gene Diversity of Q. wutaishanica
4.3. Spatial Distribution Pattern of Q. wutaishanica at Different Life Stages
4.4. Spatial Correlation of Q. wutaishanica at Different Life Stages
4.5. Suggestions for Conservation and Management
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Condit, R.; Ashton, P.S.; Baker, P.; Bunyavejchewin, S.; Gunatilleke, S.; Gunatilleke, N.; Hubbell, S.P.; Foster, R.B.; Itoh, A.; LaFrankie, J.V.; et al. Spatial Patterns in the Distribution of Tropical Tree Species. Science 2000, 288, 1414–1418. [Google Scholar] [CrossRef] [PubMed]
- Lydersen, J.M.; North, M.P.; Knapp, E.E.; Collins, B.M. Quantifying Spatial Patterns of Tree Groups and Gaps in Mixed-Conifer Forests: Reference Conditions and Long-Term Changes Following Fire Suppression and Logging. For. Ecol. Manag. 2013, 304, 370–382. [Google Scholar] [CrossRef]
- May, F.; Huth, A.; Wiegand, T. Moving beyond Abundance Distributions: Neutral Theory and Spatial Patterns in a Tropical Forest. Proc. Royal Soc. B 2015, 282, 20141657. [Google Scholar] [CrossRef]
- Lin, Y.; Chang, L.; Yang, K.; Wang, H.; Sun, I. Point Patterns of Tree Distribution Determined by Habitat Heterogeneity and Dispersal Limitation. Oecologia 2011, 165, 175–184. [Google Scholar] [CrossRef]
- Tuo, F.; Liu, X.; Liu, R.; Zhao, W.; Jing, W.; Ma, J.; Wu, X.; Zhao, J.; Ma, X. Spatial distribution patterns and association of Picea crassifolia population in Dayekou Basin of Qilian Mountains, northwestern China. Chin. J. Plant Ecol. 2020, 44, 1172–1183. [Google Scholar] [CrossRef]
- Huang, X.; Li, S.; Su, J.; Liu, W.; Lang, X. Distribution of Pinus yunnanensis Natural Population in Yunlong Tianchi National Nature Reserve. For. Res. 2018, 31, 47–52. [Google Scholar] [CrossRef]
- Liu, H.; Chen, Q.; Xu, Z.; Wu, C.; Chen, Y. Natural population structure and spatial distribution pattern of rare and endangered species Dacrydium pectinatum. Acta Ecol. Sin. 2020, 40, 2985–2995. [Google Scholar] [CrossRef]
- Martínez, I.; Wiegand, T.; González-Taboada, F.; Obeso, J.R. Spatial Associations among Tree Species in a Temperate Forest Community in North-Western Spain. For. Ecol. Manag. 2010, 260, 456–465. [Google Scholar] [CrossRef]
- Tilman, D. Resource Competition between Plankton Algae: An Experimental and Theoretical Approach. Ecology 1977, 58, 338–348. [Google Scholar] [CrossRef]
- Wright, A.; Schnitzer, S.A.; Reich, P.B. Living Close to Your Neighbors: The Importance of Both Competition and Facilitation in Plant Communities. Ecology 2014, 95, 2213–2223. [Google Scholar] [CrossRef] [Green Version]
- Peters, H.A. Neighbour-Regulated Mortality: The Influence of Positive and Negative Density Dependence on Tree Populations in Species-Rich Tropical Forests. Ecol. Lett. 2003, 6, 757–765. [Google Scholar] [CrossRef]
- Getzin, S.; Dean, C.; He, F.A.; Trofymow, J.; Wiegand, K.; Wiegand, T. Spatial Patterns and Competition of Tree Species in a Douglas-Fir Chronosequence on Vancouver Island. Ecography 2006, 29, 671–682. [Google Scholar] [CrossRef]
- Zhang, C.; Jin, W.; Gao, L.; Zhao, X. Scale Dependent Structuring of Spatial Diversity in Two Temperate Forest Communities. For. Ecol. Manag. 2014, 316, 110–116. [Google Scholar] [CrossRef]
- Tilman, D. Resource Competition and Community Structure; Princeton University Press: Princeton, NJ, USA, 1982; ISBN 978-0-691-08302-5. [Google Scholar]
- Hughes, A.R.; Inouye, B.D.; Johnson, M.T.J.; Underwood, N.; Vellend, M. Ecological Consequences of Genetic Diversity. Ecol Lett 2008, 11, 609–623. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Etienne, R.S.; Liang, M.; Wang, Y.; Yu, S. Experimental Evidence for an Intraspecific Janzen-Connell Effect Mediated by Soil Biota. Ecology 2015, 96, 662–671. [Google Scholar] [CrossRef] [PubMed]
- Campoy, J.A.; Lerigoleur-Balsemin, E.; Christmann, H.; Beauvieux, R.; Girollet, N.; Quero-García, J.; Dirlewanger, E.; Barreneche, T. Genetic Diversity, Linkage Disequilibrium, Population Structure and Construction of a Core Collection of Prunus Avium L. Landraces and Bred Cultivars. BMC Plant Biol. 2016, 16, 49. [Google Scholar] [CrossRef]
- Mas-Gómez, J.; Cantín, C.M.; Moreno, M.Á.; Prudencio, Á.S.; Gómez-Abajo, M.; Bianco, L.; Troggio, M.; Martínez-Gómez, P.; Rubio, M.; Martínez-García, P.J. Exploring Genome-Wide Diversity in the National Peach (Prunus persica) Germplasm Collection at CITA (Zaragoza, Spain). Agronomy 2021, 11, 481. [Google Scholar] [CrossRef]
- Topp, B.L.; Russell, D.M.; Neumüller, M.; Dalbó, M.A.; Liu, W. Plum. In Fruit Breeding; Badenes, M.L., Byrne, D.H., Eds.; Springer: Boston, MA, USA, 2012; pp. 571–621. ISBN 978-1-4419-0762-2. [Google Scholar]
- Mason, A.S. SSR Genotyping. Methods Mol. Biol. 2015, 1245, 77–89. [Google Scholar] [CrossRef]
- Vieira, M.L.C.; Santini, L.; Diniz, A.L.; Munhoz, C.d.F. Microsatellite Markers: What They Mean and Why They Are so Useful. Genet. Mol. Biol. 2016, 39, 312–328. [Google Scholar] [CrossRef]
- Dow, B.D.; Ashley, M.V. Factors Influencing Male Mating Success in Bur Oak, Quercus Macrocarpa. New For. 1998, 15, 161–180. [Google Scholar] [CrossRef]
- Lefort, F.; Echt, C.; Streiff, R.; Giovanni Giuseppe, V. Microsatellite Sequences: A New Generation of Molecular Markers for Forest Genetics. For. Genet. 1999, 6, 15–20. [Google Scholar]
- Lefort, F.; Lally, M.; Thompson, D.; Douglas, G. Morphological Traits, Microsatellite Fingerprinting and Genetic Relatedness of a Stand of Elite Oaks (Q. Robur L.) at Tullynally, Ireland. Silvae Genet. 1998, 47, 257–262. [Google Scholar]
- Liu, M. The Research on Genetic Evolution Relationship of Quercus. mongolia and Quercus. wutaishanica. Master’s Thesis, Northeast Forestry University, Haerbin, China, 2012. [Google Scholar]
- Qin, Y. Study on the Genetic Diversity of Quercus liaotungensis Natural Population in Shanxi Province. Master’s Thesis, Beijing Forestry University, Beijing, China, 2012. [Google Scholar]
- Zeng, Y.; Liao, W.; Petit, R.J.; Zhang, D. Exploring Species Limits in Two Closely Related Chinese Oaks. PLoS ONE 2010, 5, e15529. [Google Scholar] [CrossRef]
- Wang, M.; Xu, J.; Chai, Y.; Guo, Y.; Liu, X.; Yue, M. Differentiation of Environmental Conditions Promotes Variation of Two Quercus Wutaishanica Community Assembly Patterns. Forests 2020, 11, 43. [Google Scholar] [CrossRef]
- Chen, C.; Liu, D.; Wu, J.; Kang, M.; Zhang, J.; Liu, Q.; Liang, Y. Leaf traits of Quercus wutaishanica and their relationship with topographic factors in Mount Dongling. Chin. J. Ecol. 2015, 34, 2131–2139. [Google Scholar] [CrossRef]
- Yang, J.; Lv, J.; He, Q.; Yan, M.; Li, G.; Du, S. Time lag of stem sap flow and its relationships with transpiration characteristics in Quercus liaotungensis and Robina pseudoacacia in the loess hilly region, China. Chin. J. Appl. Ecol. 2019, 30, 2607–2613. [Google Scholar] [CrossRef]
- Ou, R.; Yan, X.; Ma, H.; Jiang, Y. Predation and removal of Quercus wutaishanica and Prunus davidiana seeds of different size by rodents. Seed 2017, 36, 76–80. [Google Scholar] [CrossRef]
- Chai, Y.; Yue, M.; Liu, X.; Guo, Y.; Wang, M.; Xu, J.; Zhang, C.; Chen, Y.; Zhang, L.; Zhang, R. Patterns of Taxonomic, Phylogenetic Diversity during a Long-Term Succession of Forest on the Loess Plateau, China: Insights into Assembly Process. Sci. Rep. 2016, 6, 27087. [Google Scholar] [CrossRef] [PubMed]
- Chai, Y.; Yue, M.; Wang, M.; Xu, J.; Liu, X.; Zhang, R.; Wan, P. Plant Functional Traits Suggest a Change in Novel Ecological Strategies for Dominant Species in the Stages of Forest Succession. Oecologia 2016, 180, 771–783. [Google Scholar] [CrossRef]
- Li, Y.; Shao, M. The change of plant diversity during natural recovery process of vegetation in Ziwuling area. Acta Ecol. Sin. 2004, 24, 252–260. [Google Scholar] [CrossRef]
- Wang, J.; Chen, Y.; Cao, Y.; Zhou, J.; Hou, L. Carbon concentration and carbon storage in different components of natural Quercus wutaishanica forest in Ziwuling of Loess Plateau, Northwest China. Chin. J. Ecol. 2012, 31, 3058–3063. [Google Scholar] [CrossRef]
- Zou, H.; Liu, G.; Wang, H. The vegetation development in North Ziwulin forest region in last fifty years. Acta Bot. Boreali Occident. Sin. 2002, 22, 1–8. [Google Scholar]
- Wang, S.; Wang, X.; Guo, H.; Fan, W.; Lv, H.; Duan, R. Distinguishing the Importance between Habitat Specialization and Dispersal Limitation on Species Turnover. Ecol. Evol. 2013, 3, 3545–3553. [Google Scholar] [CrossRef]
- Liu, P.; Wang, W.; Bai, Z.; Guo, Z.; Ren, W.; Huang, J.; Xu, Y.; Yao, J.; Ding, Y.; Zang, R. Competition and Facilitation Co-Regulate the Spatial Patterns of Boreal Tree Species in Kanas of Xinjiang, Northwest China. For. Ecol. Manag. 2020, 467, 118167. [Google Scholar] [CrossRef]
- Qiu, J.; Han, A.; He, C.; Yin, Q.; Jia, S.; Luo, Y.; Li, C.; Hao, Z. Spatial distribution pattern and intraspecific association of the dominant species Quercus aliena var. acutiserrata in Qinling Mountains, China. Chin. J. Appl. Ecol. 2022, 33, 1–9. [Google Scholar] [CrossRef]
- Hormaza, J.I. Molecular Characterization and Similarity Relationships among Apricot (Prunus armeniaca L.) Genotypes Using Simple Sequence Repeats. Theor. Appl. Genet. 2002, 104, 321–328. [Google Scholar] [CrossRef] [PubMed]
- Guerra, M.E.; Guerrero, B.I.; Casadomet, C.; Rodrigo, J. Self-(in)Compatibility, S-RNase Allele Identification, and Selection of Pollinizers in New Japanese Plum-Type Cultivars. Sci. Hortic. 2020, 261, 109022. [Google Scholar] [CrossRef]
- Guerra, M.E.; López-Corrales, M.; Wünsch, A. Improved S-Genotyping and New Incompatibility Groups in Japanese Plum. Euphytica 2012, 186, 445–452. [Google Scholar] [CrossRef]
- Guerrero, B.I.; Guerra, M.E.; Rodrigo, J. Establishing Pollination Requirements in Japanese Plum by Phenological Monitoring, Hand Pollinations, Fluorescence Microscopy and Molecular Genotyping. JoVE 2020, 165, e61897. [Google Scholar] [CrossRef]
- Kampfer, S.; Lexer, C.; Glössl, J.; Steinkellner, H. Characterization of (GA)n Microsatellite Loci from Quercus Robur. Hereditas 1998, 129, 183–186. [Google Scholar] [CrossRef]
- Steinkellner, H.; Fluch, S.; Turetschek, E.; Lexer, C.; Streiff, R.; Kremer, A.; Burg, K.; Glössl, J. Identification and Characterization of (GA/CT)n- Microsatellite Loci from Quercus petraea. Plant. Mol. Biol. 1997, 33, 1093–1096. [Google Scholar] [CrossRef] [PubMed]
- Aldrich, P.R.; Michler, C.H.; Sun, W.; Romero-Severson, J. Microsatellite Markers for Northern Red Oak (Fagaceae: Quercus rubra). Mol. Ecol. Notes 2002, 2, 472–474. [Google Scholar] [CrossRef]
- Berner, D. Allele Frequency Difference AFD–An Intuitive Alternative to FST for Quantifying Genetic Population Differentiation. Genes 2019, 10, 308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stölting, K.N.; Paris, M.; Meier, C.; Heinze, B.; Castiglione, S.; Bartha, D.; Lexer, C. Genome-Wide Patterns of Differentiation and Spatially Varying Selection between Postglacial Recolonization Lineages of Populus Alba (Salicaceae), a Widespread Forest Tree. New Phytol. 2015, 207, 723–734. [Google Scholar] [CrossRef]
- Chen, J.; Källman, T.; Ma, X.-F.; Zaina, G.; Morgante, M.; Lascoux, M. Identifying Genetic Signatures of Natural Selection Using Pooled Population Sequencing in Picea Abies. G3 Genes|Genomes|Genet. 2016, 6, 1979–1989. [Google Scholar] [CrossRef]
- Wiegand, T.A.; Moloney, K. Rings, Circles, and Null-Models for Point Pattern Analysis in Ecology. Oikos 2004, 104, 209–229. [Google Scholar] [CrossRef]
- Wiegand, T.; Moloney, K.A. Handbook of Spatial Point-Pattern Analysis in Ecology; Chapman and Hall/CRC Press: Boca Raton, FL, USA, 2013; ISBN 978-1-4200-8254-8. [Google Scholar]
- Suriguga; Zhang, J.; Cheng, J.; Zhang, B. Population structure and distribution pattern of dominant species in Tilia mandshurica forest in Dongling Mountain of Beijing. Chin. J. Ecol. 2009, 28, 1253–1258. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, X.; Guo, C.; Zhang, Q. Point pattern analysis of Pteroceltis tatarinowii population at its different development stages in limestone mountain area of north Anhui, East China. Chin. J. Ecol. 2013, 32, 542–550. [Google Scholar] [CrossRef]
- Shimada, T. Nutrient Compositions of Acorns and Horse Chestnuts in Relation to Seed-Hoarding. Ecol. Res. 2001, 16, 803–808. [Google Scholar] [CrossRef]
- Yang, X.; Anwar, E.; Zhou, J.; He, D.; Gao, Y.; Lv, G.; Cao, Y. Higher Association and Integration among Functional Traits in Small Tree than Shrub in Resisting Drought Stress in an Arid Desert. Environ. Exp. Bot. 2022, 201, 104993. [Google Scholar] [CrossRef]
- Zhu, Y.; Bai, F.; Liu, H.; Li, W.; Li, L.; Li, G.; Wang, S.; Sang, W. Population distribution patterns and interspecific spatial associations in warm temperate secondary forests, Beijing. Biodivers. Sci. 2011, 19, 252–259. [Google Scholar] [CrossRef]
- Gao, J.; Liu, Z.-L.; Zhao, W.; Tomlinson, K.W.; Xia, S.-W.; Zeng, Q.-Y.; Wang, X.-R.; Chen, J. Combined Genotype and Phenotype Analyses Reveal Patterns of Genomic Adaptation to Local Environments in the Subtropical Oak Quercus Acutissima. J. Syst. Evol. 2021, 59, 541–556. [Google Scholar] [CrossRef]
- Yang, M.; Zhang, M.; Shi, S.; Kang, Y.; Liu, J. Analysis of Genetic Structure of Magnolia sprengeri Populations Based on ISSR Markers. Sci. Silvae Sin. 2014, 50, 76–81. [Google Scholar] [CrossRef]
- Tian, H.; Kang, M.; Li, L.; Yao, X.; Huang, H. Genetic diversity in natural populations of Castanea mollissima inferred from nuclear SSR markers. Biodivers. Sci. 2009, 17, 296–302. [Google Scholar] [CrossRef]
- Tang, S.; Dai, W.; Li, M.; Zhang, Y.; Geng, Y.; Wang, L.; Zhong, Y. Genetic Diversity of Relictual and Endangered Plant Abies Ziyuanensis (Pinaceae) Revealed by AFLP and SSR Markers. Genetica 2008, 133, 21–30. [Google Scholar] [CrossRef]
- Song, Y.; Jiang, X.; Zhang, M.; Wang, Z.; Bo, W.; An, X.; Zhang, Z. Genetic differences revealed by Genomic-SSR and EST-SSR in poplar. J. Beijing For. Univ. 2010, 32, 1–7. [Google Scholar] [CrossRef]
- Wen, Y.; Han, W.; Wu, S. Plant genetic diversity and its influencing factors. J. Cent. South Univ. For. Technol. 2010, 30, 80–87. [Google Scholar] [CrossRef]
- O’Connell, L.M.; Mosseler, A.; Rajora, O.P. Extensive Long-Distance Pollen Dispersal in a Fragmented Landscape Maintains Genetic Diversity in White Spruce. J. Hered. 2007, 98, 640–645. [Google Scholar] [CrossRef]
- Rushton, B.S. Natural Hybridization within the Genus Quercus L. Ann. For. Sci. 1993, 50, 73s–90s. [Google Scholar] [CrossRef]
- Souza, I.G.B.; Souza, V.A.B.; Lima, P.S.C. Molecular Characterization of Platonia Insignis Mart. (“Bacurizeiro”) Using Inter Simple Sequence Repeat (ISSR) Markers. Mol. Biol. Rep. 2013, 40, 3835–3845. [Google Scholar] [CrossRef]
- Ma, X.; Zhao, C.; Zhang, Q.; Li, Y.; Hou, Z. Spatial pattern and spatial association of Melica przewalskyi and Artemisia frigida in degraded grassland. Chin. J. Ecol. 2013, 32, 299–304. [Google Scholar] [CrossRef]
- Zhang, J. Analysis of spatial point pattern for plant species. Acta Phytopathol. Sin. 1998, 22, 344–349. [Google Scholar]
- Jin, X.; Zhang, Q.; Xu, Q.; Ji, Y.; Bi, R. Population distribution patterns and interspecific spatial associations of Acanthopanax senticosus populations in Lingkong Mountain, Shanxi Province, China. Plant Sci. J. 2018, 36, 327–335. [Google Scholar] [CrossRef]
- Wiegand, T.; Gunatilleke, S.; Gunatilleke, N. Species Associations in a Heterogeneous Sri Lankan Dipterocarp Forest. Am. Nat. 2007, 170, e77–e95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, G.; Ding, Y.; Zang, R.; Guo, Z.; Zhang, X.; Cheng, K.; Bai, Z.; Ayoufu, B. Distribution patterns of Picea schrenkiana var. tianschanica population in Tianshan Mountains. Chin. J. Appl. Ecol. 2011, 22, 9–13. [Google Scholar] [CrossRef]
- Zhang, Z.; Hu, G.; Zhu, J.; Luo, D.; Ni, J. Spatial Patterns and Interspecific Associations of Dominant Tree Species in Two Old-Growth Karst Forests, SW China. Ecol. Res. 2010, 25, 1151–1160. [Google Scholar] [CrossRef]
- Carrer, M.; Castagneri, D.; Popa, I.; Pividori, M.; Lingua, E. Tree Spatial Patterns and Stand Attributes in Temperate Forests: The Importance of Plot Size, Sampling Design, and Null Model. For. Ecol. Manag. 2018, 407, 125–134. [Google Scholar] [CrossRef]
- Wang, X.; Ye, J.; Li, B.; Zhang, J.; Lin, F.; Hao, Z. Spatial Distributions of Species in an Old-Growth Temperate Forest, Northeastern China. Can. J. For. Res. 2010, 40, 1011–1019. [Google Scholar] [CrossRef]
- Jiang, D.; Tang, Y.; Busso, C.A. Effects of Vegetation Cover on Recruitment of Ulmus Pumila L. in Horqin Sandy Land, Northeastern China. J. Arid Land 2014, 6, 343–351. [Google Scholar] [CrossRef]
- Murrell, L.D.J. The Community-Level Consequence of Seed Dispersal Patterns. Annu. Rev. Ecol. Evol. Syst. 2003, 34, 549–574. [Google Scholar] [CrossRef]
- Ismail, S.A.; Ghazoul, J.; Ravikanth, G.; Kushalappa, C.G.; Uma Shaanker, R.; Kettle, C.J. Evaluating Realized Seed Dispersal across Fragmented Tropical Landscapes: A Two-Fold Approach Using Parentage Analysis and the Neighbourhood Model. New Phytol. 2017, 214, 1307–1316. [Google Scholar] [CrossRef] [PubMed]
- Bohrer, G.; Katul, G.G.; Nathan, R.; Walko, R.L.; Avissar, R. Effects of Canopy Heterogeneity, Seed Abscission and Inertia on Wind-Driven Dispersal Kernels of Tree Seeds. J. Ecol. 2008, 96, 569–580. [Google Scholar] [CrossRef]
- Liu, S.; Jiao, J.; Hu, S.; Wu, D.; Deng, N. Effect of flood runoff on seed dispersal and population regeneration—A case study of Salix matsudana in the Loess Hill and Gully Region. Res. Soil Water Conserv. 2018, 25, 99–103. [Google Scholar] [CrossRef]
- Fraver, S.; D’Amato, A.W.; Bradford, J.B.; Jonsson, B.G.; Jönsson, M.; Esseen, P.-A. Tree Growth and Competition in an Old-Growth Picea Abies Forest of Boreal Sweden: Influence of Tree Spatial Patterning. J. Veg. Sci. 2014, 25, 374–385. [Google Scholar] [CrossRef]
- Zhang, Y. Seed Rain Dynamics and Seedling Spatial Pattern of Deciduous Broad-Leaved Forest in Malan Forest Region of Loess Plateau. Master’s Thesis, Shaanxi Normal University, Xi’an, China, 2015. [Google Scholar]
- Wang, X.; Liang, C.; Wang, W. Balance between Facilitation and Competition Determines Spatial Patterns in a Plant Population. Chin. Sci. Bull. 2014, 59, 1405–1415. [Google Scholar] [CrossRef]
- Yang, X.; Miao, Y.; Zhang, Q.; Zhang, L.; Bi, R. Spatial Pattern Analysis of Individuals in Different Age-classes of Pinus bungeana in Wulu Mountain Reserve, Shanxi, China. Bull. Bot. Res. 2013, 33, 24–30. [Google Scholar]
- Shen, Z.; Hua, M.; Dan, Q.; Lu, J.; Fang, J. Spatial pattern analysis and associations of Quercus aquifolioides population at different growth stages in Southeast Tibet, China. Chin. J. Appl. Ecol. 2016, 27, 387–394. [Google Scholar] [CrossRef]
- Omelko, A.; Ukhvatkina, O.; Zhmerenetsky, A.; Sibirina, L.; Petrenko, T.; Bobrovsky, M. From Young to Adult Trees: How Spatial Patterns of Plants with Different Life Strategies Change during Age Development in an Old-Growth Korean Pine-Broadleaved Forest. For. Ecol. Manag. 2018, 411, 46–66. [Google Scholar] [CrossRef]
- Cai, F. A stuidy on the structure and dynamics of Cyclobalanopsis glauca population at hills around West Lake in Hangzhou. Sci. Silvae Sin. 2000, 36, 67–72. [Google Scholar] [CrossRef]
- Hubbell, S.P. Neutral Theory and the Evolution of Ecological Equivalence. Ecology 2006, 87, 1387–1398. [Google Scholar] [CrossRef]
- Brooker, R.W.; Maestre, F.T.; Callaway, R.M.; Lortie, C.L.; Cavieres, L.A.; Kunstler, G.; Liancourt, P.; Tielbörger, K.; Travis, J.M.J.; Anthelme, F.; et al. Facilitation in Plant Communities: The Past, the Present, and the Future. J. Ecol. 2008, 96, 18–34. [Google Scholar] [CrossRef]
- Metz, J.; Annighöfer, P.; Schall, P.; Zimmermann, J.; Kahl, T.; Schulze, E.-D.; Ammer, C. Site-Adapted Admixed Tree Species Reduce Drought Susceptibility of Mature European Beech. Glob. Chang. Biol. 2016, 22, 903–920. [Google Scholar] [CrossRef] [PubMed]
- Muhamed, H.; Maalouf, J.-P.; Michalet, R. Summer Drought and Canopy Opening Increase the Strength of the Oak Seedlings–Shrub Spatial Association. Ann. For. Sci. 2013, 70, 345–355. [Google Scholar] [CrossRef]
- Zanini, L.; Ganade, G.; Hübel, I. Facilitation and Competition Influence Succession in a Subtropical Old Field. Plant Ecol. 2006, 185, 179–190. [Google Scholar] [CrossRef]
- Kubota, Y. Spatial Pattern and Regeneration Dynamics in a Temperate Abies–Tsuga Forest in Southwestern Japan. J. For. Res. 2006, 11, 191–201. [Google Scholar] [CrossRef]
- Gray, L.; He, F. Spatial Point-Pattern Analysis for Detecting Density-Dependent Competition in a Boreal Chronosequence of Alberta. For. Ecol. Manag. 2009, 259, 98–106. [Google Scholar] [CrossRef]
- Muhamed, H.; Touzard, B.; Le Bagousse-Pinguet, Y.; Michalet, R. The Role of Biotic Interactions for the Early Establishment of Oak Seedlings in Coastal Dune Forest Communities. For. Ecol. Manag. 2013, 297, 67–74. [Google Scholar] [CrossRef]
- Comita, L.S.; Hubbell, S.P. Local Neighborhood and Species’ Shade Tolerance Influence Survival in a Diverse Seedling Bank. Ecology 2009, 90, 328–334. [Google Scholar] [CrossRef]
- Han, A.; Qiu, J.; He, C.; Yin, Q.; Jia, S.; Luo, Y.; Li, C.; Hao, Z. Spatial distribution patterns and intraspecific and interspecific associations of the dominant shrub species Lonicera fragrantissima var. lancifolia in Huangguan of Qinling Mountains, China. Chin. J. Appl. Ecol. 2022, 33, 1–9. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Locus | Primer Sequence (5′–3′) | Annealing Temperature (°C) |
|---|---|---|
| PL123-124 | (F) GCTTGAGAGTTGAGATTTGT | 55 |
| (R) GCAACACCCTTTAACTACCA | ||
| PL127-128 | (F) GCAATTACAGGCTAGGCTGG | 55 |
| (R) GTCTGGACCTAGCCCTCATG | ||
| PL125-126 | (F) CTTCACTGGCTTTTCCTCCT | 58 |
| (R) TGAAGCCCTTGTCAACATGC | ||
| E79 | (F) CCATTAAAAGAAGCAGTATTTTGT | 52 |
| (R) GCAACACTCAGCCTATATCTAGAA | ||
| E71 | (F) CGTCTATAAGTTCTTGGGTGA | 46 |
| (R) GTAACTATGATGTGATTCTTACTTCA | ||
| Qden 05011 | (F) CCCACTCCCTGTCCATTGT | 59 |
| (R) CACTGTGTGCTGCGACTTG | ||
| ssrQrZAG 96 | (F) CCCAGTCACATCCACTACTGTCC | 59 |
| (R) GGTTGGGAAAAGGAGATCAGA | ||
| ssrQrZAG 7 | (F) CAACTTGGTGTTCGGATCAA | 55 |
| (R) GTGCATTTCTTTTATAGCATTCAC | ||
| 01b | (F) GTTCAACAATTTTATTAGGGTGC | 56 |
| (R) GCCTATTACACACAACAAGCC | ||
| 02b | (F) ATGTCAATATGGTCACCTACCG | 53 |
| (R) TTTTTGTAGATTTTTAAGCACGC | ||
| 04b | (F) TTCCTTTTCCTCAGTTTGGG | 52 |
| (R) CCCGCATCAAAGAACTATTG | ||
| 10b | (F) GAATGGATCTTCATTTATCGTTG | 55 |
| (R) TCTGCATATTTTCAACATACATTTAG |
| Life Stage | Locus | N | Na | Ne | PIC | I | He | Ho | Fis |
|---|---|---|---|---|---|---|---|---|---|
| Trees and saplings | 02b | 105 | 7 | 3.8677 | 0.5525 | 1.5015 | 0.7450 | 0.8000 | −0.0790 |
| 04b | 106 | 22 | 9.3555 | 0.4253 | 2.5257 | 0.8973 | 0.8208 | 0.0810 | |
| 10b | 99 | 13 | 7.3060 | 0.5260 | 2.1323 | 0.8675 | 0.7980 | 0.0755 | |
| E71 | 101 | 11 | 5.6719 | 0.5156 | 1.9138 | 0.8278 | 0.8713 | −0.0578 | |
| E79 | 105 | 13 | 3.8215 | 0.2940 | 1.6315 | 0.7419 | 0.8952 | −0.2125 | |
| Mean | — | 13.2 | 6.0045 | 0.4627 | 1.9410 | 0.8159 | 0.8371 | — | |
| Trees | 02b | 53 | 7 | 3.6985 | 0.5284 | 1.4758 | 0.7736 | 0.7366 | 0.0370 |
| 04b | 53 | 18 | 9.3478 | 0.5193 | 2.4809 | 0.8302 | 0.9015 | −0.0713 | |
| 10b | 49 | 10 | 6.7067 | 0.6707 | 2.0250 | 0.7143 | 0.8597 | −0.1454 | |
| E71 | 50 | 10 | 5.7670 | 0.5767 | 1.9331 | 0.8800 | 0.8349 | 0.0451 | |
| E79 | 53 | 11 | 4.3652 | 0.3968 | 1.7378 | 0.8302 | 0.7783 | 0.0519 | |
| Mean | — | 11.2 | 5.9770 | 0.5384 | 1.9305 | 0.8057 | 0.8222 | — | |
| Saplings | 02b | 52 | 6 | 3.9132 | 0.6522 | 1.4981 | 0.7517 | 0.8269 | −0.0752 |
| 04b | 53 | 16 | 7.9575 | 0.4973 | 2.2968 | 0.8827 | 0.8113 | −0.0714 | |
| 10b | 50 | 13 | 7.8125 | 0.6010 | 2.1930 | 0.8808 | 0.8800 | 0.0008 | |
| E71 | 51 | 9 | 5.2229 | 0.5803 | 1.8111 | 0.8165 | 0.8627 | −0.0462 | |
| E79 | 52 | 8 | 3.1333 | 0.3917 | 1.3315 | 0.6875 | 0.9615 | −0.2740 | |
| Mean | — | 10.4 | 5.6079 | 0.5445 | 1.8261 | 0.8038 | 0.8685 | — |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, D.; Xu, Y.; Chai, Y.; Tian, T.; Wang, K.; Liu, P.; Wang, M.; Zhu, J.; Hou, D.; Yue, M. Spatial Distribution Pattern and Genetic Diversity of Quercus wutaishanica Mayr Population in Loess Plateau of China. Forests 2022, 13, 1375. https://0-doi-org.brum.beds.ac.uk/10.3390/f13091375
Hu D, Xu Y, Chai Y, Tian T, Wang K, Liu P, Wang M, Zhu J, Hou D, Yue M. Spatial Distribution Pattern and Genetic Diversity of Quercus wutaishanica Mayr Population in Loess Plateau of China. Forests. 2022; 13(9):1375. https://0-doi-org.brum.beds.ac.uk/10.3390/f13091375
Chicago/Turabian StyleHu, Dong, Yao Xu, Yongfu Chai, Tingting Tian, Kefeng Wang, Peiliang Liu, Mingjie Wang, Jiangang Zhu, Dafu Hou, and Ming Yue. 2022. "Spatial Distribution Pattern and Genetic Diversity of Quercus wutaishanica Mayr Population in Loess Plateau of China" Forests 13, no. 9: 1375. https://0-doi-org.brum.beds.ac.uk/10.3390/f13091375