
Carbon Allocation of Quercus mongolica Fisch. ex Ledeb. across Different Life Stages Differed by Tree and Shrub Growth Forms at the Driest Site of Its Distribution

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area and Site Features
2.2. Field Survey
2.3. Sample Collection
2.4. Measurements of NSC Concentration
2.5. Statistical Analysis
3. Results
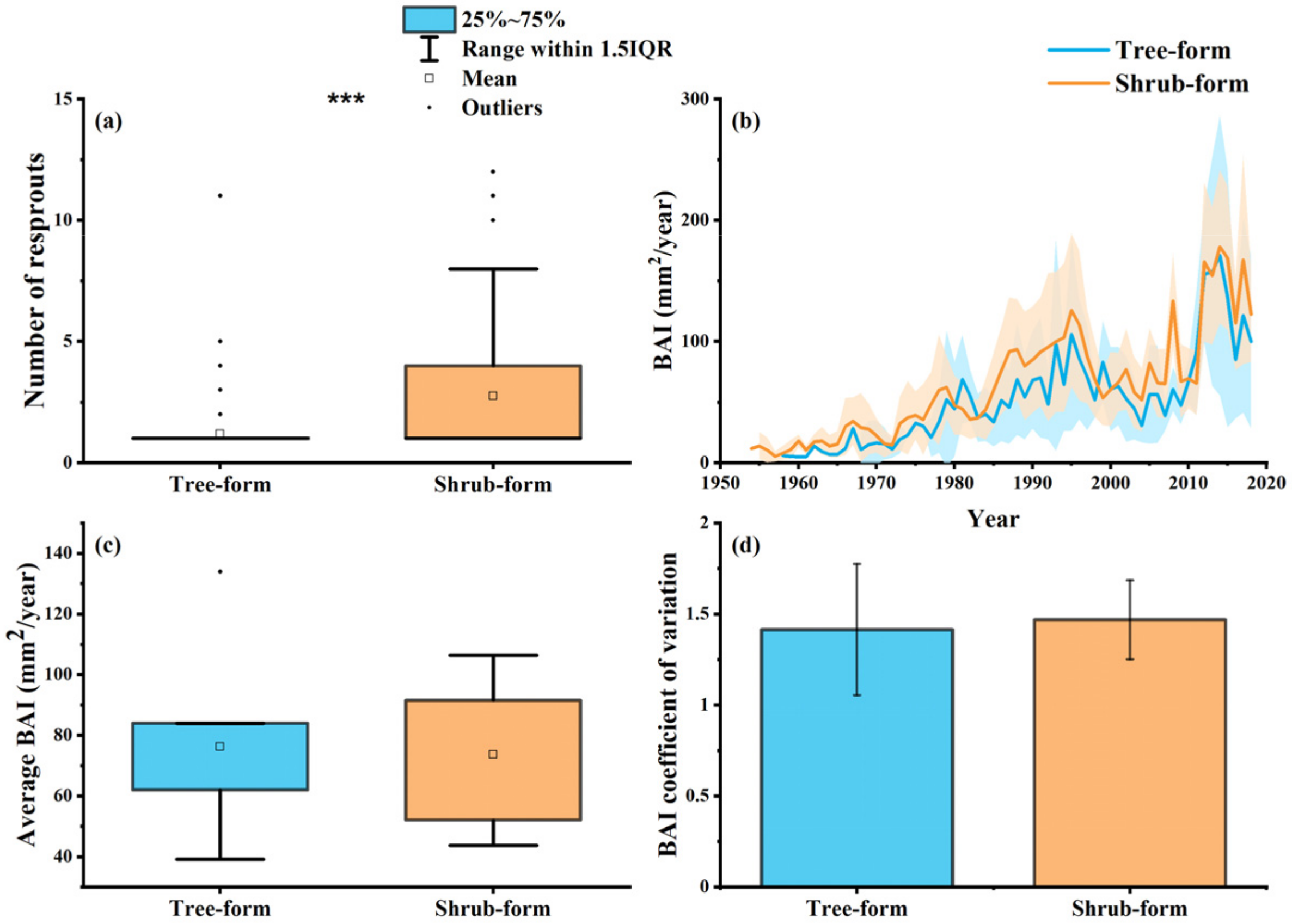
3.1. Growth Features of Tree and Shrub Form Quercus mongolica
3.2. Carbon Allocation for Reproduction and Growth
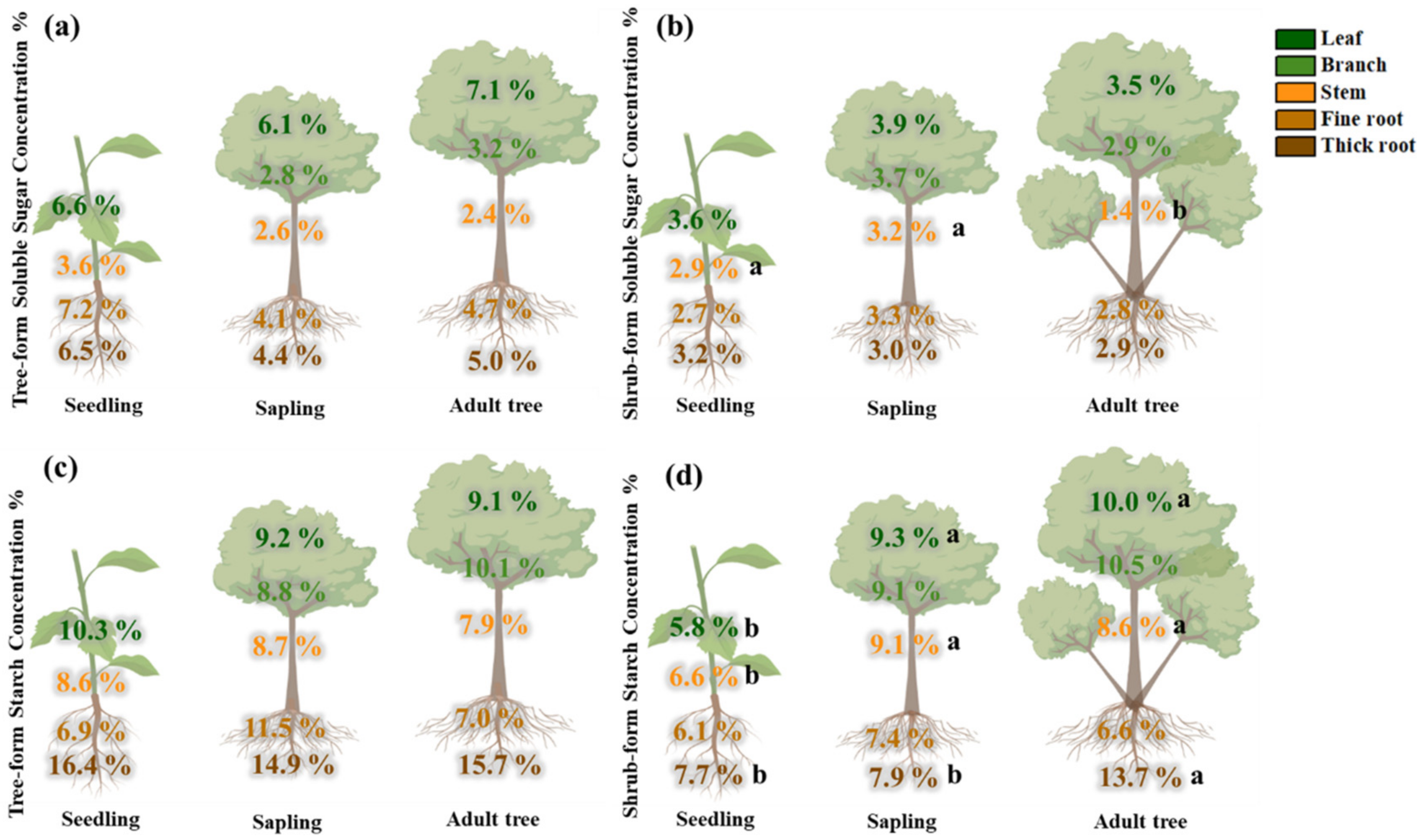
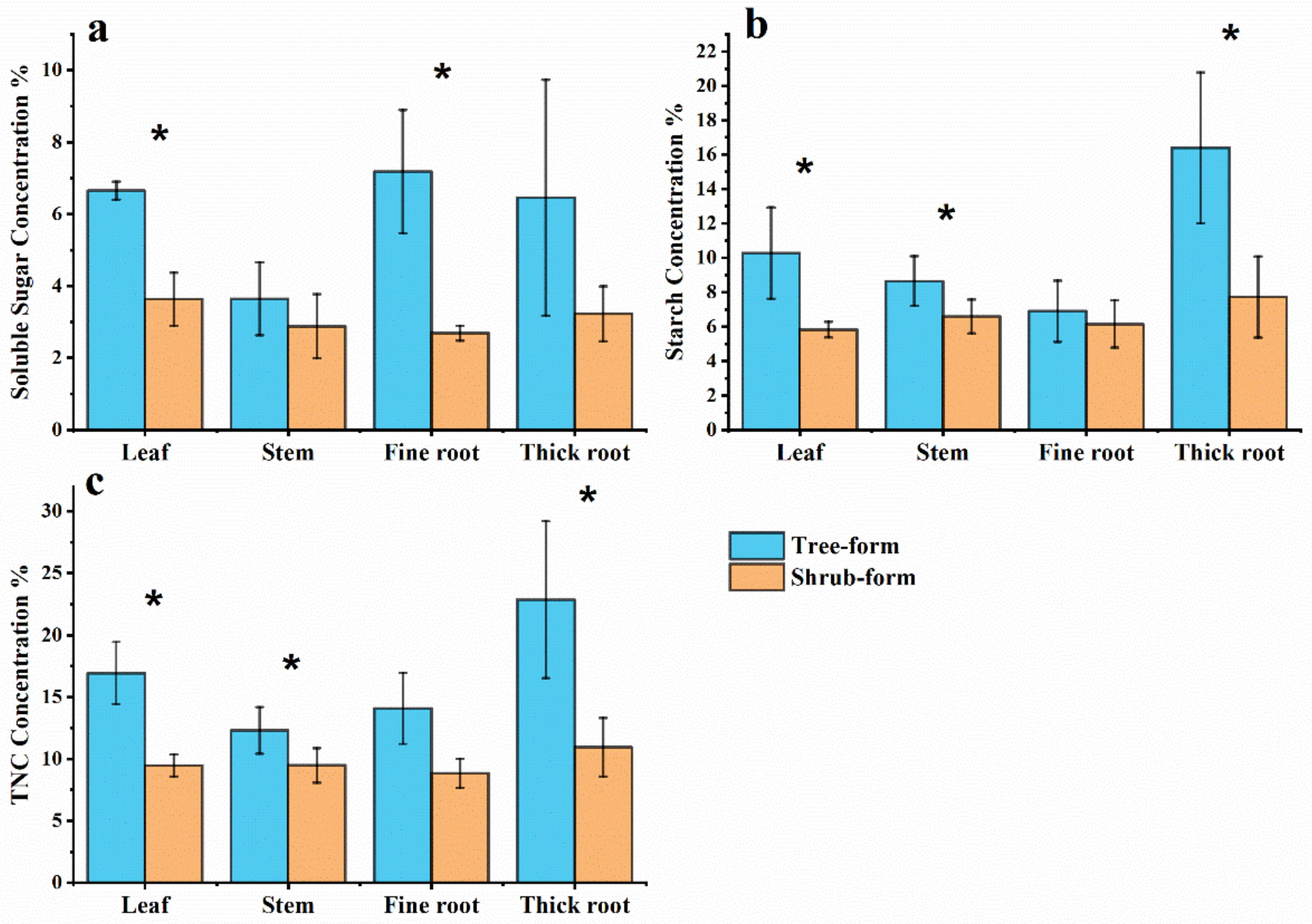
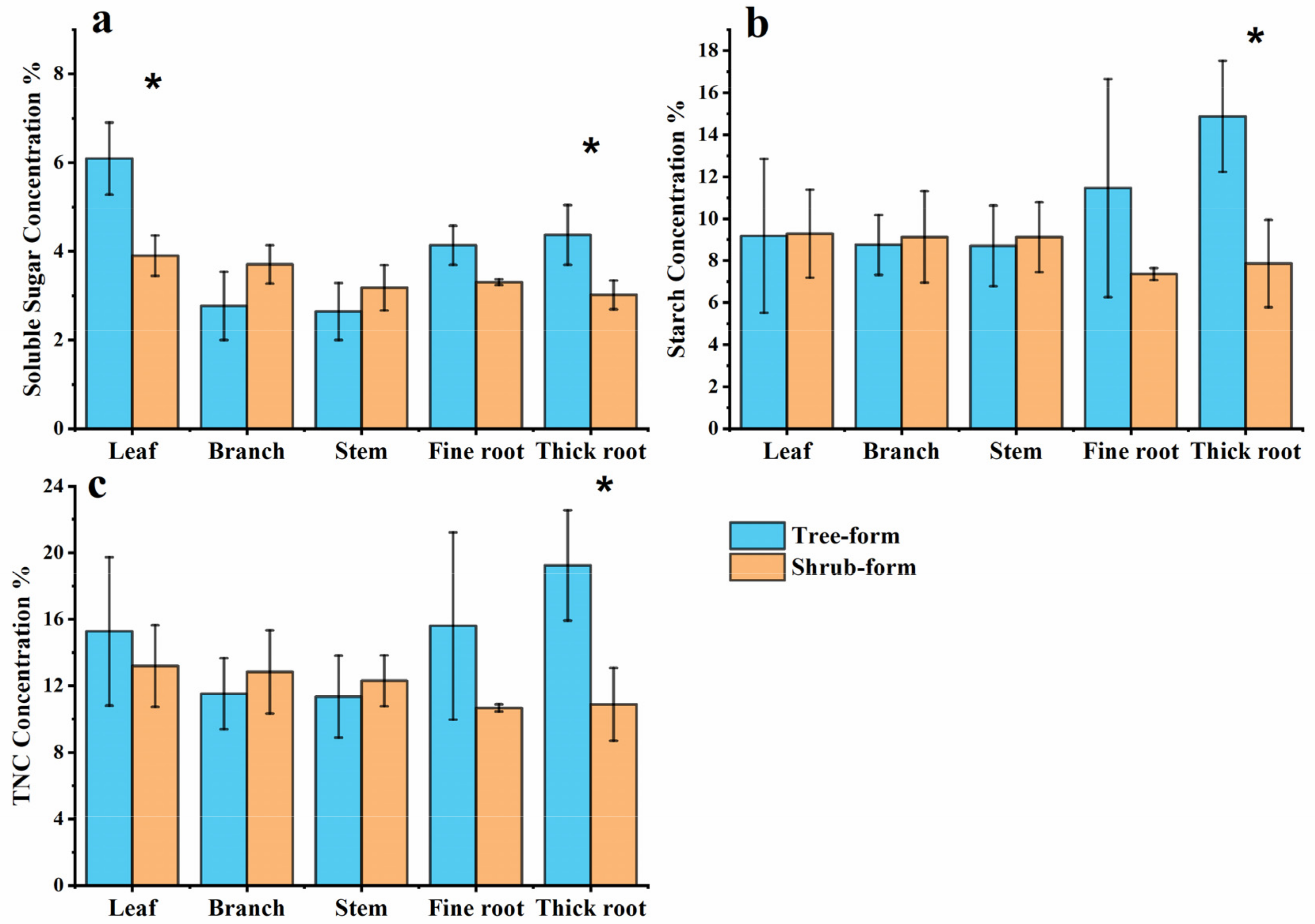
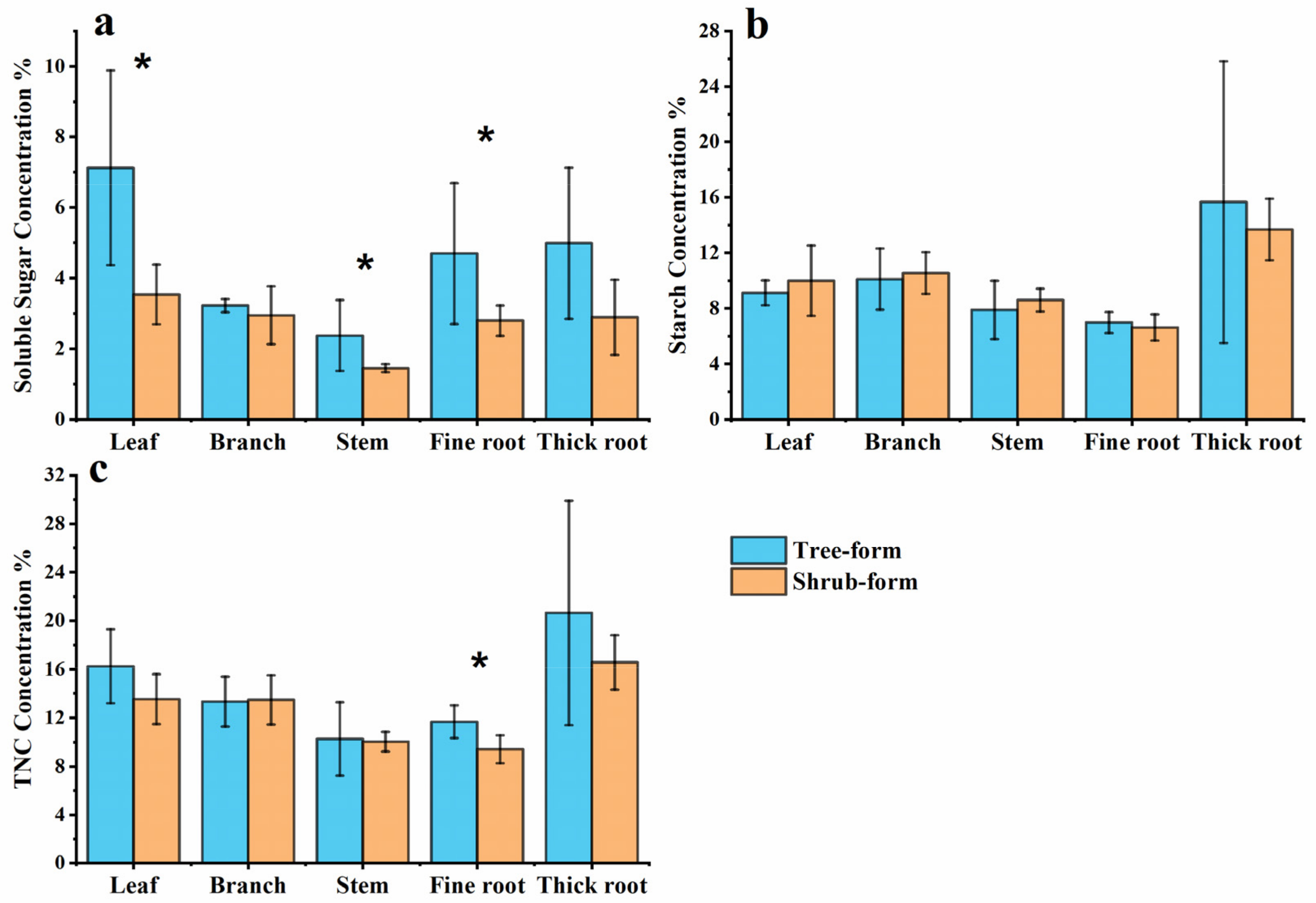
3.3. Carbon Allocation for NSC Storage
3.4. Effects of Different Life Stages on NSC Storage
4. Discussion
4.1. Growth Form Affecting Carbon Allocation for Both Tree Growth and Reproduction
4.2. Divergent Carbon Allocation at Different Life Stages
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Jucker, T.; Fischer, F.J.; Chave, J.; Coomes, D.A.; Caspersen, J.; Ali, A.; Loubota Panzou, G.J.; Feldpausch, T.R.; Falster, D.; Usoltsev, V.A.; et al. Tallo: A global tree allometry and crown architecture database. Global. Change Biol. 2022, 28, 5254–5268. [Google Scholar] [CrossRef] [PubMed]
- Scheffer, M.; Vergnon, R.; Cornelissen, J.H.C.; Hantson, S.; Holmgren, M.; van Nes, E.H.; Xu, C. Why trees and shrubs but rarely trubs? Trends Ecol. Evol. 2014, 29, 433–434. [Google Scholar] [CrossRef] [PubMed]
- Vincent, G.; Harja, D. Exploring Ecological Significance of Tree Crown Plasticity through Three-dimensional Modelling. Ann. Bot. 2007, 101, 1221–1231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stovall, A.E.L.; Shugart, H.; Yang, X. Tree height explains mortality risk during an intense drought. Nat. Commun. 2019, 10, 4385. [Google Scholar] [CrossRef] [Green Version]
- Dai, J.; Liu, H.; Wang, Y.; Guo, Q.; Hu, T.; Quine, T.; Green, S.; Hartmann, H.; Xu, C.; Liu, X.; et al. Drought-modulated allometric patterns of trees in semi-arid forests. Commun. Biol. 2020, 3, 405. [Google Scholar] [CrossRef]
- Barrett, S.C.H. Influences of clonality on plant sexual reproduction. Proc. Natl. Acad. Sci. USA 2015, 112, 8859–8866. [Google Scholar] [CrossRef] [Green Version]
- Qiu, T.; Andrus, R.; Aravena, M.; Ascoli, D.; Bergeron, Y.; Berretti, R.; Berveiller, D.; Bogdziewicz, M.; Boivin, T.; Bonal, R.; et al. Limits to reproduction and seed size-number trade-offs that shape forest dominance and future recovery. Nat. Commun. 2022, 13, 2381. [Google Scholar] [CrossRef]
- Wenk, E.H.; Falster, D.S. Quantifying and understanding reproductive allocation schedules in plants. Ecol. Evol. 2015, 5, 5521–5538. [Google Scholar] [CrossRef] [Green Version]
- Xu, C.; Liu, H.; Zhou, M.; Xue, J.; Zhao, P.; Shi, L.; Shangguan, H. Enhanced sprout-regeneration offsets warming-induced forest mortality through shortening the generation time in semiarid birch forest. Forest Ecol. Manag. 2018, 409, 298–306. [Google Scholar] [CrossRef]
- Zeppel, M.J.B.; Harrison, S.P.; Adams, H.D.; Kelley, D.I.; Li, G.; Tissue, D.T.; Dawson, T.E.; Fensham, R.; Medlyn, B.E.; Palmer, A.; et al. Drought and resprouting plants. New Phytol. 2015, 206, 583–589. [Google Scholar] [CrossRef]
- Malanson, G.P.; Trabaud, L. Vigour of post-fire resprouting by Quercus coccifera L. J. Ecol. 1988, 76, 351–365. [Google Scholar] [CrossRef]
- Bellingham, P.J.; Sparrow, A.D. Resprouting as a life history strategy in woody plant communities. Oikos 2000, 89, 409–416. [Google Scholar] [CrossRef]
- Moles, A.T.; Ackerly, D.D.; Webb, C.O.; Tweddle, J.C.; Dickie, J.B.; Pitman, A.J.; Westoby, M. Factors that shape seed mass evolution. Proc. Natl. Acad. Sci. USA 2005, 102, 10540–10544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frank, D.; Reichstein, M.; Bahn, M.; Thonicke, K.; Frank, D.; Mahecha, M.D.; Smith, P.; Velde, M.; Vicca, S.; Babst, F.; et al. Effects of climate extremes on the terrestrial carbon cycle: Concepts, processes and potential future impacts. Global. Chang. Biol. 2015, 21, 2861–2880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lauder, J.D.; Moran, E.V.; Hart, S.C. Fight or flight? Potential tradeoffs between drought defense and reproduction in conifers. Tree Physiol. 2019, 39, 1071–1085. [Google Scholar] [CrossRef] [PubMed]
- Genet, H.; Breda, N.; Dufrene, E. Age-related variation in carbon allocation at tree and stand scales in beech (Fagus sylvatica L.) and sessile oak (Quercus petraea (Matt.) Liebl.) using a chronosequence approach. Tree Physiol. 2010, 30, 177–192. [Google Scholar] [CrossRef]
- Green, J.K.; Keenan, T.F. The limits of forest carbon sequestration. Science 2022, 376, 692–693. [Google Scholar] [CrossRef]
- Long, R.W.; Dudley, T.L.; D’Antonio, C.M.; Grady, K.C.; Bush, S.E.; Hultine, K.R. Spenders versus savers: Climate-induced carbon allocation trade-offs in a recently introduced woody plant. Funct. Ecol. 2021, 35, 1640–1654. [Google Scholar] [CrossRef]
- Yin, X.H.; Hao, G.Y.; Sterck, F. A trade-off between growth and hydraulic resilience against freezing leads to divergent adaptations among temperate tree species. Funct. Ecol. 2022, 36, 739–750. [Google Scholar] [CrossRef]
- Wiley, E.; Casper, B.B.; Helliker, B.R. Recovery following defoliation involves shifts in allocation that favour storage and reproduction over radial growth in black oak. J. Ecol. 2017, 105, 412–424. [Google Scholar] [CrossRef]
- Jacquet, J.S.; Bosc, A.; O’Grady, A.; Jactel, H. Combined effects of defoliation and water stress on pine growth and non-structural carbohydrates. Tree Physiol. 2014, 34, 367–376. [Google Scholar] [CrossRef] [PubMed]
- Piper, F.I. Patterns of carbon storage in relation to shade tolerance in southern South American species. Am. J. Bot. 2015, 102, 1442–1452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maguire, A.J.; Kobe, R.K. Drought and shade deplete nonstructural carbohydrate reserves in seedlings of five temperate tree species. Ecol. Evol. 2015, 5, 5711–5721. [Google Scholar] [CrossRef] [PubMed]
- Ferrenberg, S.; Kane, J.M.; Langenhan, J.M. To grow or defend? Pine seedlings grow less but induce more defences when a key resource is limited. Tree Physiol. 2015, 35, 107–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morgan, J.M. Osmoregulation and water-stress in higher-plants. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1984, 35, 299–319. [Google Scholar] [CrossRef]
- Guo, J.S.; Gear, L.; Hultine, K.R.; Koch, G.W.; Ogle, K. Non-structural carbohydrate dynamics associated with antecedent stem water potential and air temperature in a dominant desert shrub. Plant Cell Environ. 2020, 43, 1467–1483. [Google Scholar] [CrossRef]
- Hartmann, H.; Trumbore, S. Understanding the roles of nonstructural carbohydrates in forest trees—From what we can measure to what we want to know. New Phytol. 2016, 211, 386–403. [Google Scholar] [CrossRef] [Green Version]
- Drake, P.L.; Mendham, D.S.; White, D.A.; Ogden, G.N. A comparison of growth, photosynthetic capacity and water stress in Eucalyptus globulus coppice regrowth and seedlings during early development. Tree Physiol. 2009, 29, 663–674. [Google Scholar] [CrossRef] [Green Version]
- He, W.; Liu, H.; Qi, Y.; Liu, F.; Zhu, X. Patterns in nonstructural carbohydrate contents at the tree organ level in response to drought duration. Global. Chang. Biol. 2020, 26, 3627–3638. [Google Scholar] [CrossRef]
- Hartmann, H.; Adams, H.D.; Hammond, W.M.; Hoch, G.; Landhäusser, S.M.; Wiley, E.; Zaehle, S. Identifying differences in carbohydrate dynamics of seedlings and mature trees to improve carbon allocation in models for trees and forests. Environ. Exp. Bot. 2018, 152, 7–18. [Google Scholar] [CrossRef]
- Xu, X.; Wang, Z.; Rahbek, C.; Sanders, N.J.; Fang, J. Geographical variation in the importance of water and energy for oak diversity. J. Biogeogr. 2016, 43, 279–288. [Google Scholar] [CrossRef]
- Dai, J.; Lu, S.; Qi, Y.; Liu, H. Tree-to-Shrub Shift Benefits the Survival of Quercus mongolica Fisch. ex Ledeb. at the Xeric Timberline. Forests 2022, 13, 244. [Google Scholar] [CrossRef]
- Zhu, Y.; Comita, L.S.; Hubbell, S.P.; Ma, K. Conspecific and phylogenetic density-dependent survival differs across life stages in a tropical forest. J. Ecol. 2015, 103, 957–966. [Google Scholar] [CrossRef]
- Schweingruber, F.H. Tree Rings: Basics and Applications of Dendrochronology; Springer Science & Business Media: Dordrecht, The Netherlands, 2012. [Google Scholar]
- Holmes, R.L. Computer-assisted quality control in tree-ring dating and measurement. Tree-Ring Bull. 1983, 43, 69–78. [Google Scholar]
- Monserud, R.A.; Sterba, H. A basal area increment model for individual trees growing in even-and uneven-aged forest stands in Austria. For. Ecol. Manag. 1996, 80, 57–80. [Google Scholar] [CrossRef]
- Li, M.H.; Hoch, G.; Körner, C. Spatial variability of mobile carbohydrates within Pinus cembra trees at the alpine treeline. Phyton 2001, 41, 203–213. [Google Scholar]
- Trevelyan, W.E.; Harrison, J.S. Studies on yeast metabolism. I. Fractionation and microdetermination of cell carbohydrates. Biochem. J. 1952, 50, 298–303. [Google Scholar] [CrossRef]
- Osaki, M.H.U.S.; Shinano, T.; Tadano, T. Redistribution of carbon and nitrogen compounds from the shoot to the harvesting organs during maturation in field crops. Soil Sci. Plant Nutr. 1991, 37, 117–128. [Google Scholar] [CrossRef]
- Quentin, A.G.; Pinkard, E.A.; Ryan, M.G.; Tissue, D.T.; Baggett, L.S.; Adams, H.D.; Maillard, P.; Marchand, J.; Landhäusser, S.M.; Lacointe, A.; et al. Non-structural carbohydrates in woody plants compared among laboratories. Tree Physiol. 2015, 35, 1146–1165. [Google Scholar] [CrossRef] [Green Version]
- Ashwell, G. Colorimetric analysis of sugars. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 1957; Volume 3, pp. 73–105. [Google Scholar]
- Niinemets, Ü. Responses of forest trees to single and multiple environmental stresses from seedlings to mature plants: Past stress history, stress interactions, tolerance and acclimation. Forest Ecol. Manag. 2010, 260, 1623–1639. [Google Scholar] [CrossRef]
- Wiley, E.; Helliker, B. A re-evaluation of carbon storage in trees lends greater support for carbon limitation to growth. N. Phytol. 2012, 195, 285–289. [Google Scholar] [CrossRef] [PubMed]
- Vesk, P.A. Plant size and resprouting ability: Trading tolerance and avoidance of damage? J. Ecol. 2006, 94, 1027–1034. [Google Scholar] [CrossRef]
- Weber, R.; Schwendener, A.; Schmid, S.; Lambert, S.; Wiley, E.; Landhausser, S.M.; Hartmann, H.; Hoch, G. Living on next to nothing: Tree seedlings can survive weeks with very low carbohydrate concentrations. New Phytol. 2018, 218, 107–118. [Google Scholar] [CrossRef] [Green Version]
- Hoch, G.; Richter, A.; Körner, C. Non-structural carbon compounds in temperate forest trees. Plant Cell Environ. 2003, 26, 1067–1081. [Google Scholar] [CrossRef]
- Sala, A.; Hoch, G. Height-related growth declines in ponderosa pine are not due to carbon limitation. Plant Cell Environ. 2009, 32, 22–30. [Google Scholar] [CrossRef] [PubMed]
- Martin, A.R.; Thomas, S.C. Size-dependent changes in leaf and wood chemical traits in two Caribbean rainforest trees. Tree Physiol. 2013, 33, 1338–1353. [Google Scholar] [CrossRef] [Green Version]
- Qiu, T.; Sharma, S.; Woodall, C.W.; Clark, J.S. Niche Shifts From Trees to Fecundity to Recruitment That Determine Species Response to Climate Change. Front. Ecol. Evol. 2021, 9, 719141. [Google Scholar] [CrossRef]
- Bowen, B.J.; Pate, J.S. The Significance of Root Starch in Post-fire Shoot Recovery of the Resprouter Stirlingia latifolia R. Br. (Proteaceae). Ann. Bot. 1993, 72, 7–16. [Google Scholar] [CrossRef]
- Tixier, A.; Guzmán-Delgado, P.; Sperling, O.; Amico Roxas, A.; Laca, E.; Zwieniecki, M.A. Comparison of phenological traits, growth patterns, and seasonal dynamics of non-structural carbohydrate in Mediterranean tree crop species. Sci. Rep. 2020, 10, 347. [Google Scholar] [CrossRef] [Green Version]
- Gilson, A.; Barthes, L.; Delpierre, N.; Dufrene, E.; Fresneau, C.; Bazot, S. Seasonal changes in carbon and nitrogen compound concentrations in a Quercus petraea chronosequence. Tree Physiol. 2014, 34, 716–729. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Growth Form | Seedling | Sapling | Adult Tree/Shrub | ||||
|---|---|---|---|---|---|---|---|
| Height (m) | Seedlings from Seed Germination | Height (m) | DBH (cm) | Height (m) | DBH (cm) | Age (Year) | |
| Tree | 0.14 ± 0.06 a | 33.3% | 3.10 ± 2.25 a | 5.60 ± 2.49 a | 7.21 ± 1.81 a | 11.65 ± 1.32 a | 33.8 ± 16.6 a |
| Shrub | 0.16 ± 0.08 a | 0 | 2.41 ± 1.55 b | 5.13 ± 2.90 a | 4.26 ± 1.23 b | 11.33 ± 2.13 a | 48.1 ± 12.7 a |
| Fixed Factors | Soluble Sugar | Starch | TNC |
|---|---|---|---|
| Growth form | 37.7754 *** | 7.9883 ** | 27.0347 *** |
| Life stage | 5.1772 * | 1.0088 | 0.0049 |
| Organ | 12.2682 *** | 13.7526 *** | 14.6334 *** |
| Growth form × Life stage | 2.3192 | 3.4001 * | 3.9453 * |
| Growth form × Organ | 5.9409 ** | 4.9803 ** | 6.7068 *** |
| Fixed Factors | Tree Form | Shrub Form | ||||
|---|---|---|---|---|---|---|
| Soluble Sugar | Starch | TNC | Soluble Sugar | Starch | TNC | |
| Life stage | 5.00 * | 1.07 | 4.00 | 3.17 | 19.01 *** | 12.65 ** |
| Organ | 12.83 *** | 10.85 *** | 13.17 *** | 10.67 *** | 7.55 *** | 8.21 *** |
| Fixed Factors | Seedling | Sapling | Adult Tree | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Soluble Sugar | Starch | TNC | Soluble Sugar | Starch | TNC | Soluble Sugar | Starch | TNC | |
| Growth form | 16.53 ** | 32.10 ** | 42.17 ** | 5.30 | 1.75 | 2.60 | 12.38 ** | 0.11 | 2.62 |
| Organ | 3.93 * | 9.14 ** | 7.04 ** | 9.00 *** | 1.81 | 2.83 * | 6.07 *** | 7.81 *** | 9.48 *** |
| Growth form × Organ | 2.30 | 3.63 * | 3.17 * | 6.22 ** | 7.05 *** | 6.55 *** | 1.84 | 0.41 | 0.71 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qi, Y.; Liu, H.; He, W.; Dai, J.; Shi, L.; Song, Z. Carbon Allocation of Quercus mongolica Fisch. ex Ledeb. across Different Life Stages Differed by Tree and Shrub Growth Forms at the Driest Site of Its Distribution. Forests 2022, 13, 1745. https://0-doi-org.brum.beds.ac.uk/10.3390/f13111745
Qi Y, Liu H, He W, Dai J, Shi L, Song Z. Carbon Allocation of Quercus mongolica Fisch. ex Ledeb. across Different Life Stages Differed by Tree and Shrub Growth Forms at the Driest Site of Its Distribution. Forests. 2022; 13(11):1745. https://0-doi-org.brum.beds.ac.uk/10.3390/f13111745
Chicago/Turabian StyleQi, Yang, Hongyan Liu, Wenqi He, Jingyu Dai, Liang Shi, and Zhaopeng Song. 2022. "Carbon Allocation of Quercus mongolica Fisch. ex Ledeb. across Different Life Stages Differed by Tree and Shrub Growth Forms at the Driest Site of Its Distribution" Forests 13, no. 11: 1745. https://0-doi-org.brum.beds.ac.uk/10.3390/f13111745