
Soil Chemical Properties Strongly Influence Distributions of Six Kalidium Species in Northwest China

1
State Key Laboratory of Herbage Improvement and Grassland Agro-Ecosystems, College of Ecology, Lanzhou University, Lanzhou 730000, China
2
Northwest Institute of Eco-Environment and Resources, Chinese Academy of Sciences, Lanzhou 730000, China
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Forests 2022, 13(12), 2178; https://0-doi-org.brum.beds.ac.uk/10.3390/f13122178
Submission received: 21 November 2022
/
Revised: 12 December 2022
/
Accepted: 16 December 2022
/
Published: 19 December 2022
(This article belongs to the Special Issue Plant Adaptation to Extreme Environments in Drylands)
Abstract
:The degrees of adaptive responses of different halophytes to saline–alkali soil vary substantially. Kalidium (Amaranthaceae), a genus comprised of six species of succulent euhalophytes with significantly differing distributions in China, provides ideal material for exploring the ecophysiological relationships involved in these variations. Thus, in a large-scale field survey in 2014–2018, samples of soil (at 20 cm depth intervals spanning 0 to 100 cm) and seeds were collected from areas where these six species are naturally distributed. Chemical properties of soils in the areas and germinability of the species’ seeds in media with 0–500 mM NaCl and 0–250 mM Na2SO4 were then analyzed to test effects of salinity-related factors on the species’ distributions. The pH of the soil samples mainly ranged between 8.5 and 10.5 and positively correlated with their mean total salt contents. Germination rates of all six species’ seeds were negatively correlated with concentrations of NaCl and Na2SO4 in the media, and their recovery germination rates in distilled water were high (>74%). The results show that the species’ distributions and chemical properties of their saline soils are strongly correlated, notably the dominant cation at all sites is Na+, but the dominant anions at K. cuspidatum and K. caspicum sites are Cl− and SO42−, respectively. Species-associated variations in concentrations of Ca2+ were also detected. Thus, our results provide clear indications of major pedological determinants of the species’ geographic ranges and strong genotype-environment interactions among Kalidium species.
Keywords:
germination percentage; Kalidium; halophytes; pH; ion content; total salt contents; saline soil1. Introduction
One of the diverse environmental factors that strongly affect terrestrial plants’ natural distributions is soil salinity, which has growth-impairing and lethal effects on all plants when it exceeds species-dependent thresholds [1,2,3]. Further, salinity reportedly reduces crop yields on about a fifth of all irrigated land and, in combination with increasing global scarcity of water resources, salinization of soil and water is seriously threatening crop yields and future food production [4,5]. However, halophytes, comprising about 1% of the world’s flora, can grow in saline environments with relatively high concentrations of electrolytes [6]. For example, the growth and development of glycophytes is severely inhibited by exposure to 100–200 mM of NaCl, while halophytes can tolerate and complete their life cycles at substantially higher concentrations [2,4,7]. This is due to anatomical adaptations in halophytes, such as salt bladders, salt hairs, and/or salt glands in the leaves [7,8] and various physiological tolerance mechanisms. For example, excess salt may be excreted through trichomes of halophytic grasses [8], or diluted by increases in the water content and thickness of succulent halophytes’ leaves [9,10]. Moreover, different halophytes growing together on the same saline–alkali soil often have substantially differing elemental concentrations, indicating that their physiological selectivity varies [11]. Generally, the dominant ions in salty habitats are Na+ and Cl−, but other ions (including Ca2+, Mg2+, K+, SO42−, and CO32−) may also be abundant [9,12]. Moreover, both their absolute and relative concentrations may vary, and influence the composition of the associated plant communities [13,14,15].
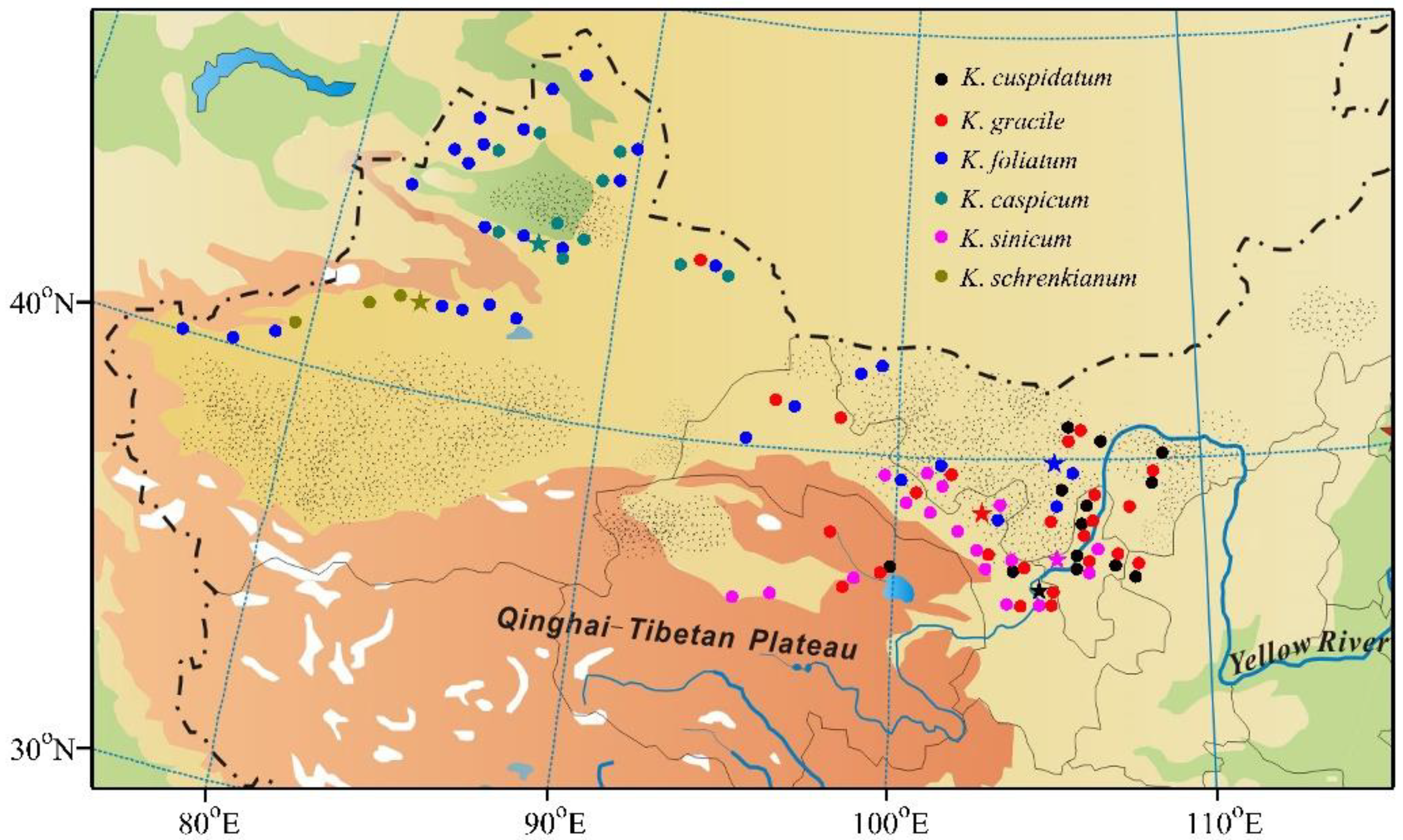
Kalidium (Amaranthaceae) is a genus of succulent halophytes with five species (Kalidium caspicum (L.) Ung.-Sternb., Kalidium cuspidatum (Ung.-Sternb.) Grubov., Kalidium foliatum (Pall.) Moq., Kalidium gracile Fenzl, and Kalidium schrenkianum Bunge ex Ung.-Sternb.), mainly distributed as shrubs in Northwest Asia and Southeast Europe. Some authorities have also recognized K. wagenitzii as an endemic species in Turkey, but others include it in K. foliatum [16]. In addition, two varieties of K. cuspidatum (var. cuspidatum and sinicum A. J. Li) have been recognized as separate species [17]. Here these varieties are called species, mainly due to genetic differences in DNA barcodes [17], but partly because our results indicate that they have significant adaptive differences. All six Kalidium species (including the two varieties of K. cuspidatum as species) naturally grow in deserts in northwest China. These succulent halophytes provide important fodder for livestock in winter, and have high ecological value for soil and water conservation in their semi-arid and arid areas. An extensive field survey showed that their distributions in China significantly differ (Figure 1). K. foliatum is the most widespread. Most K. cuspidatum sites are in Ningxia, K. gracile is mostly located in Gansu, Qinghai and Inner Mongolia. K. sinicum is naturally distributed in East Gansu and West Ningxia, while K. caspicum and K. schrenkianum are mainly restricted to regions north and south of the Tianshan Mountains in Xinjiang, respectively.
The adaptive evolution and origin of key halotolerance mechanisms have been intensively studied, as reviewed for example [2,18]. However, the NaCl concentration is not the sole stressor in saline environments. Thermal and water stresses are also often important [19,20]. Furthermore, salinity may be associated with extreme pH and/or variations in relative proportions of both cations and anions [21]. Effects of these variations have been less extensively studied, so this study focused on their impacts on distributions of the six Kalidium species in China. For this purpose, soil samples were collected from sites of the six species, across their ranges in China, then the pH, total salt contents, and ion contents of the soil were assayed at 20 cm depth intervals spanning 0 to 100 cm. In addition, the germinability of seeds of the six species was determined under different concentrations of NaCl and Na2SO4. Correlations between these abiotic factors and distributions of the Kalidium species were then examined, to explore mechanisms affecting the relationship between biodiversity and ecosystem functions.
2. Materials and Methods
2.1. Field Investigation and Sampling
In a comprehensive field investigation of the areas where Kalidium Moq is distributed in northwest China during 2014–2018, we identified 103 representative sites, in total, of the six Kalidium species. These sites are mainly located in gravel deserts and/or gravel dunes, according to the FAO soil classification system (FAO 2016). Vegetation at the sites is dominated by Kalidium Moq and Halocnemum strobilaceum of the Amaranthaceae, Halogeton arachnoideus (Amaranthaceae), Stipa glareosa (Gramineae) and various other halophytes. Distances between neighboring sites mainly ranged from 150 to 200 km. Soil samples were collected from centers of these areas using a soil auger, with three replications (three-point sampling), at 20 cm depth intervals from 0 to 100 cm. Furthermore, mature seeds were collected from the six Kalidium species and stored in a refrigerator at −20 °C before the start of experiments. The altitude and geographic coordinates of each site were measured using an Etrex GIS unit (Garmin, Taiwan). Locations of the collection sites are shown in Figure 1.
2.2. Determination of Soil Chemical Properties
The soil samples were air-dried, passed through a 1 mm sieve, then their pH was measured at a 1:5 soil: water ratio (w/v) using a PHS-25 pH meter (Shanghai Biocotek Inc., Shanghai, China), and their electrical conductivity (mS/cm) using a DDSJ-318 conductivity meter (Shanghai Biocotek Inc., Shanghai, China). Their contents of eight ions were also analyzed: Na+, K+, Ca2+, and Mg2+ using a 180-80 Polarized Zeeman atomic absorption spectrophotometer (Hitachi Inc., Tokyo, Japan); CO32− and HCO3− by the double indicator titration method; Cl− by silver nitrate titration; and SO42− by the turbidimetric method [22].
2.3. Determination of Seeds’ Germinability
Mean seed mass was calculated by weighing 1000 seeds of each Kalidium species with three replications (Table S1). Then germination and recovery experiments were conducted in an LRH 550-G programmed controlled-environment chamber (Shaoguan taihong, China) providing 16 h light/8 h dark cycles with cool white fluorescent lamps 100 μmol m−2 s−1 (Philips), and 25/19 °C day/night temperatures.
Seeds of the six Kalidium species were subjected to treatment with NaCl at seven concentrations by placing them on filter paper in Petri dishes (9 cm diameter) moistened with 10 mL of 0, 50, 100, 200, 300, 400, and 500 mM NaCl solution. Other batches were exposed to Na2SO4 with corresponding Na concentrations (0, 25, 50, 100, 150, 200 and 250 mM). Germination (regarded as emergence of the radicle from the seed by about 1 mm) was scored every day for 7 days. The germination rate at that point was calculated for each species, then ungerminated seeds were transferred to distilled water, and incubated under otherwise identical conditions. Germination of these seeds was scored for a further 7 days, after which the recovery germination percentage was calculated for each species.
2.4. Data Analysis
The soils’ total salt contents were estimated from electrical conductivity measurements, using the following empirically determined linear relationship between NaCl concentration (y) and conductivity (x): y = 0.0159x (R2 = 0.9811) [23]. Relationships between total salt contents and pH in 20 cm layers of the top 100 cm of soil at sites of the six species were examined by linear regression. The significance of differences in germination rates and recovery germination rates of seeds at different salt concentrations was tested by one-way ANOVA. Values of 18 bioclimatic factors covering most of the distributions of the six species were downloaded from the Global Climate Database (http://www.worldclim.org/bioclim, accessed on 9 November 2021). After excluding redundant bioclimatic factors, by applying a cumulative contribution ratio threshold of 80% [24], eight remained (Table S2). These were used in combination with altitude and two soil factors (pH and total salt concentration of the soil) in the Principal Component Analysis (PCA) of abiotic factors affecting distributions of each of the six studied Kalidium species. For this, SPSS (Version 19.0., Chicago, IL, USA) was used (with the significance threshold set at p < 0.05).
3. Results
3.1. pH and Total Salt Contents
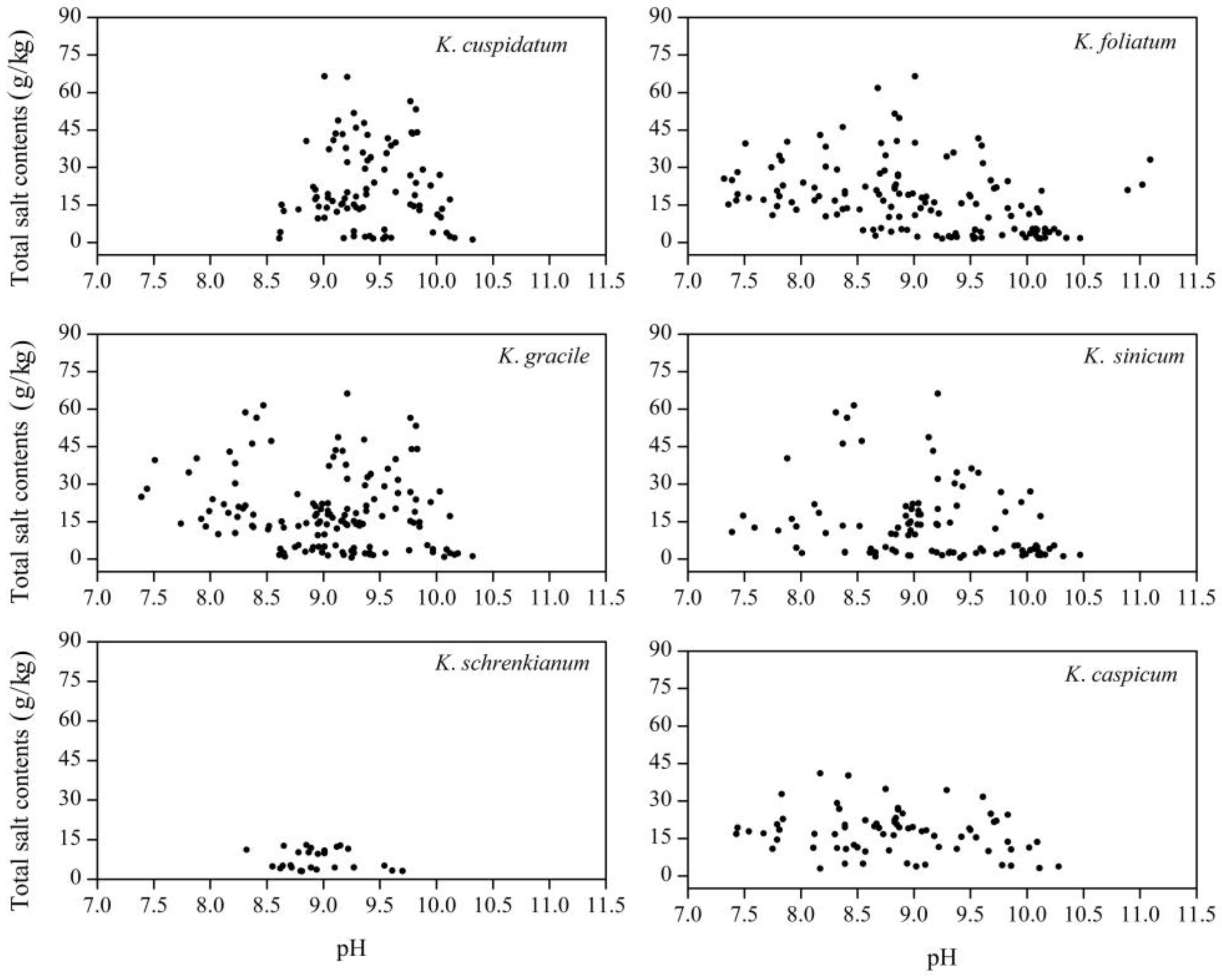
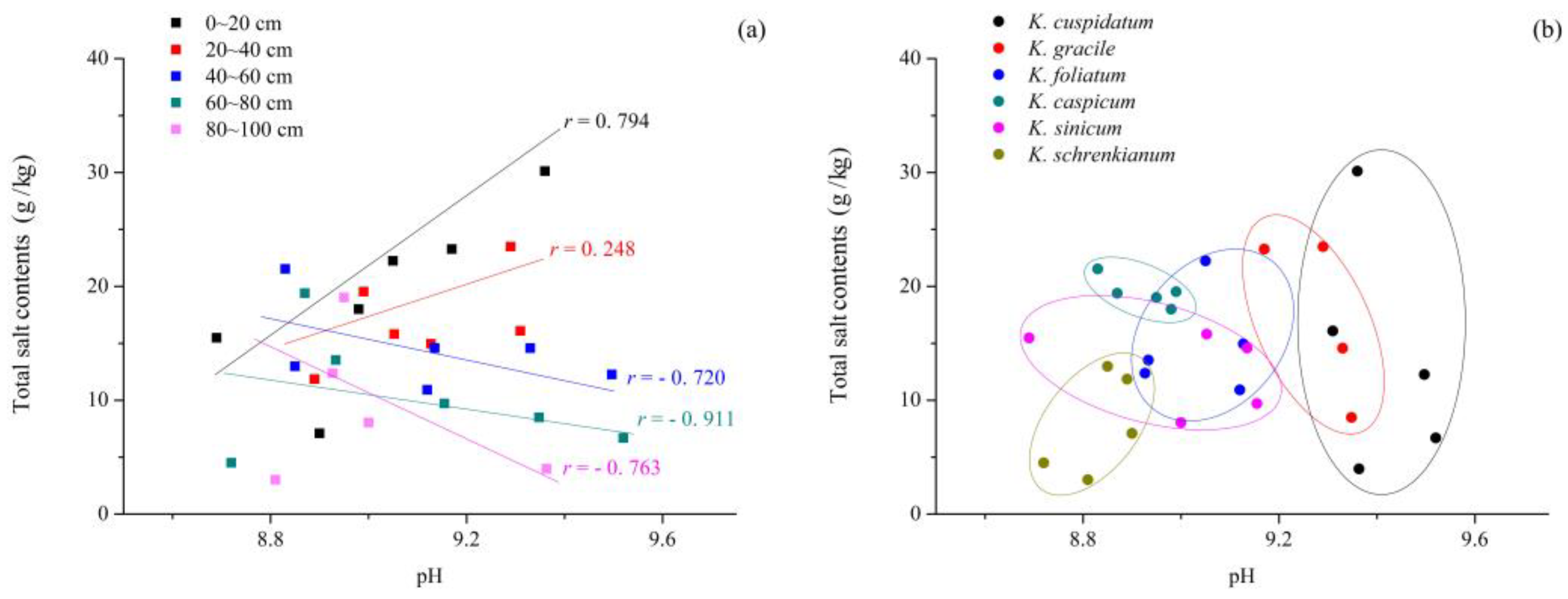
The pH of soils in the areas occupied by the Kalidium species mainly ranged from 8.5 to 10.5, although the K. foliatum sites had a wider range (7.3–11). The mean pH of soil samples from areas occupied by K. cuspidatum, K. gracile, K. sinicum, K. foliatum, K. caspicum, and K. schrenkianum was 9.41, 9.05, 9.13, 9.03, 8.94, and 8.90, respectively (Figure 2). Mean total salt contents in areas occupied by K. cuspidatum, K. gracile, K. foliatum, K. caspicum and K. sinicum were 22.4, 19.5, 17.6, 16.5 and 14.3 g/kg, respectively (Figure 2), while the highest mean value for any layer in K. schrenkianum areas was just 13.0 g/kg (in the 0–20 cm layer) and the overall mean was just 7.08 g/kg (Figure 2 and Figure 3a). Generally, there was a positive correlation between pH values and mean salt contents in the topsoil (0–20 cm), but at >40 cm depths the relationship turned negative in areas occupied by all six species (Figure 3a). However, salt contents of samples from K. cuspidatum areas (with the highest mean pH and salt contents) significantly varied with depth while they were much more constant in K. caspicum areas (Figure 3). In addition, pH and total salt contents varied much more widely in soil samples from K. foliatum, K. gracile, and K. sinicum areas than in samples from K. schrenkianum areas (Figure 2 and Figure 3).
3.2. Na+, Ca2+, K+, and Mg2+ Concentrations
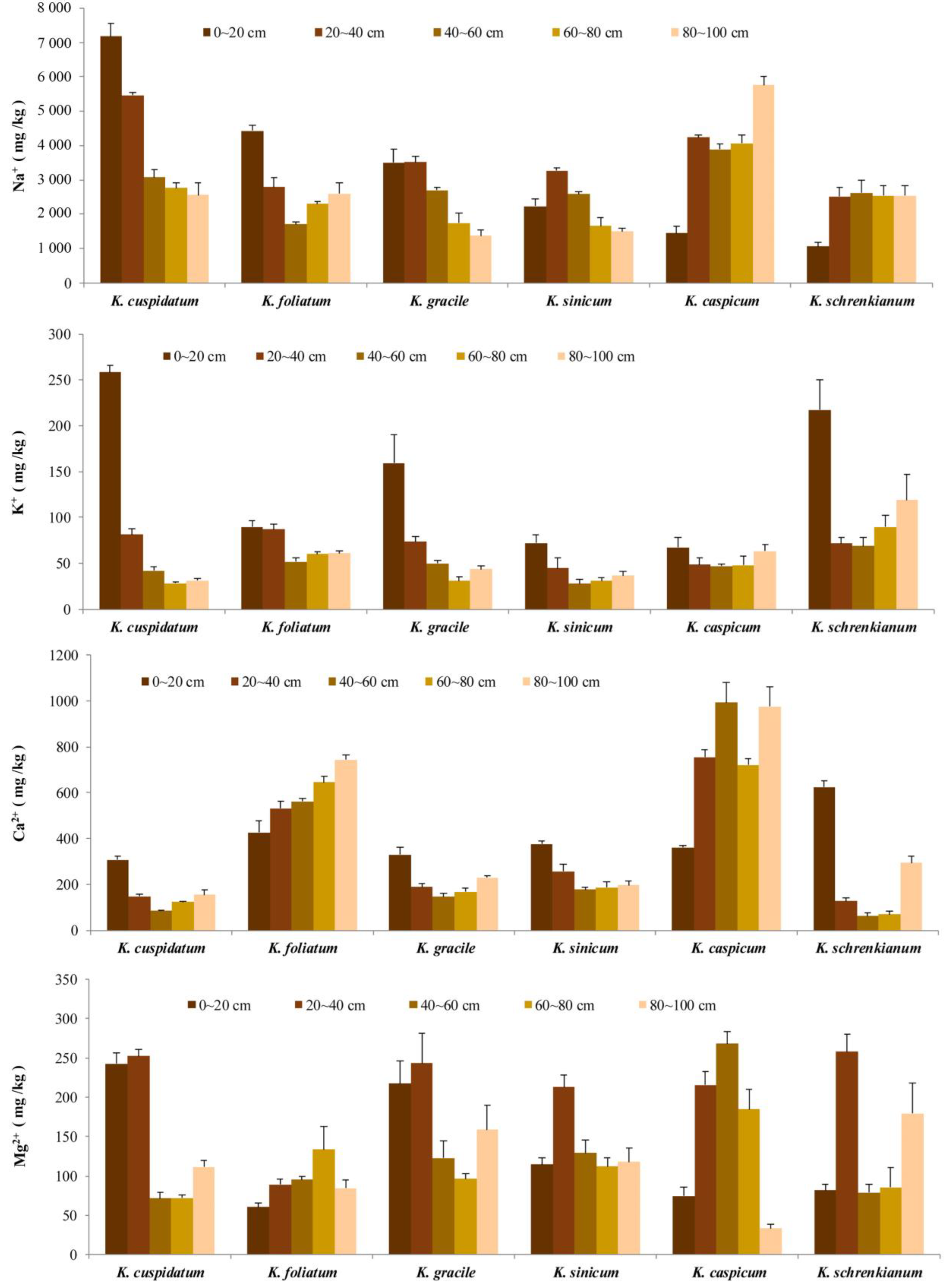
Na+ concentrations in areas occupied by the six species were all high (Figure 4), especially in K. cuspidatum areas, where they declined with increases in depth but even at 80–100 cm exceeded 2.5 g/kg, the upper limit for sensitive crops according to the FAO. In K. gracile and K. schrenkianum areas, Na+ concentrations peaked at 20–40 cm depth. Mean Na+ concentrations in samples from all soil layers in areas occupied by the six species ranged from 0.90 to 4.22 g/kg. As shown in Table 1, Na+ contents were also significantly positively correlated (r > 0.89) with total salt contents in soil from the six species’ areas, except for 20–40 cm samples (r = 0.632), strongly suggesting that Na+ made the largest contributions to the total salt contents.
Ca2+ was the second most abundant cation in all tested soil samples (Figure 4). In K. foliatum and K. caspicum areas, mean concentrations increased as soil depth increased, reaching 0.74 and 0.98 g/kg at 80–100 cm, respectively. In contrast, in K. cuspidatum and K. sinicum areas, mean Ca2+ concentrations declined as depth increased and then increased (Figure 4), to <86 mg/kg at 40–60 cm in K. cuspidatum areas (Figure 4). Overall, as shown in Table 1, there was a negative correlation between mean Ca2+ concentrations and total salt contents in the topsoil (r = −0.841, p < 0.05), but a positive correlation between them at 60–80 cm (r = 0.934, p < 0.05) and 80–100 cm (r = 0.965, p < 0.01).
K+ concentrations did not exceed 260 mg/kg in any tested soil samples. In K. gracile and K. sinicum areas, as depth increased they first declined and then increased, and were almost twice as high in the former as the latter at each soil depth (0–60 cm) (Figure 4). K+ concentrations significantly declined with depth (from 216, and 257 mg/kg, respectively, in topsoil) at all sites occupied by K. schrenkianum and K. cuspidatum (Figure 4).
Mg2+ concentrations were also substantially lower than Na+ and Ca2+ concentrations (consistently < 300 mg/kg). In areas occupied by K. caspicum, K. gracile, and K. sinicum they first increased and then declined with increases in soil depth (Figure 4), peaking at 268 mg/kg at 40–60 cm in samples from K. caspicum areas (Figure 4). Overall, as shown in Table 1, at sites of all six species, Mg2+ concentrations were positively correlated with total salt contents at 40–60 cm (r = 0.980, p < 0.01) and 80–100 cm (r = 0.991, p < 0.01) depths.
3.3. Cl−, SO42− and HCO3− Concentrations
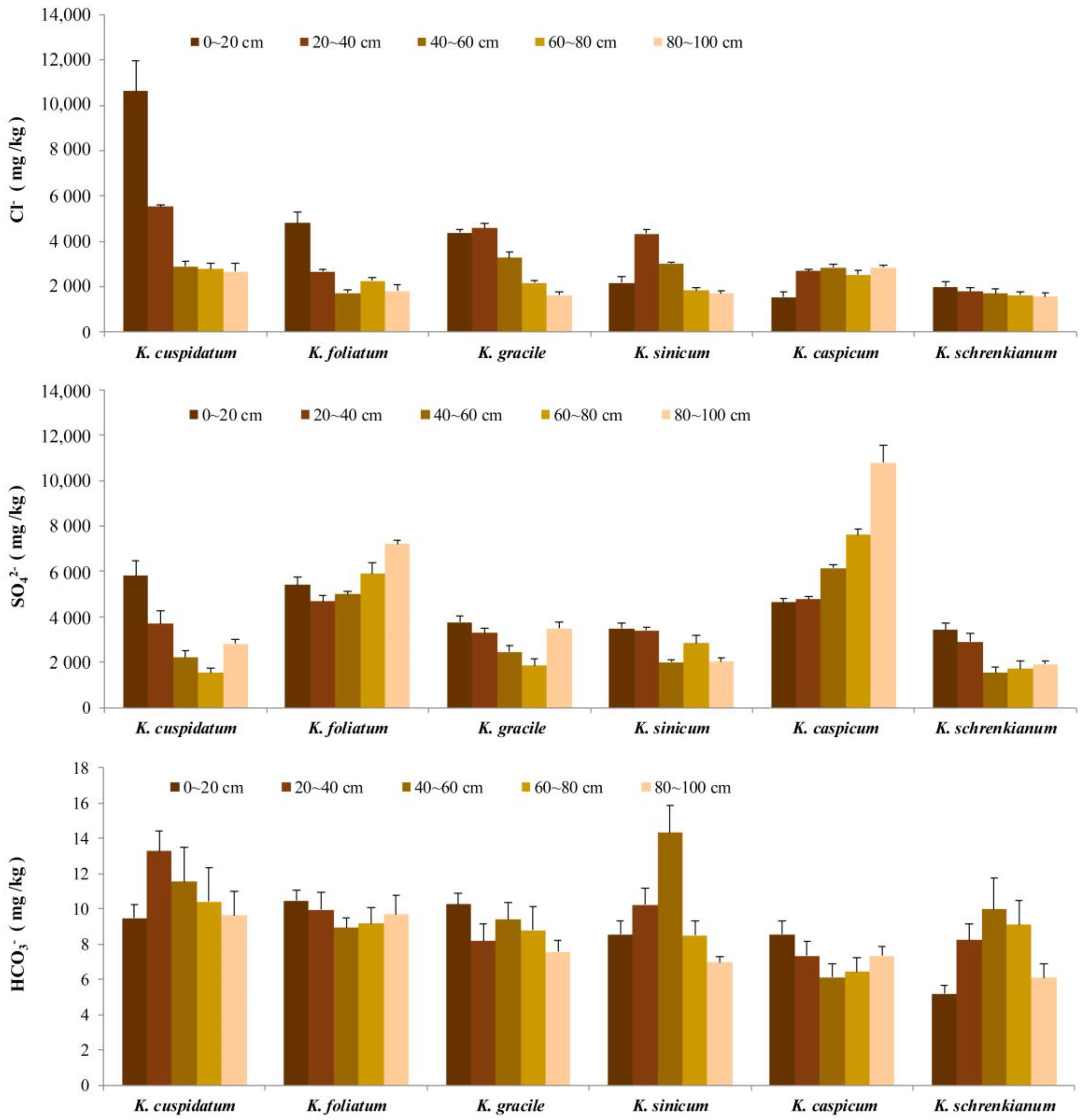
Mean Cl− concentrations were highest in K. cuspidatum areas, where they declined from 10.62 g/kg in topsoil but still exceeded 2.5 g/kg at 80–100 cm depth, and lowest in K. caspicum areas, where the mean topsoil concentration was only 1.53 g/kg (Figure 5). It was also a major anion in K. sinicum, K. foliatum and K. gracile areas, where mean concentrations were 2.6, 2.7 and 3.2 g/kg, respectively (Figure 5). Overall, as shown in Table 1, Cl− concentrations were positively correlated with total salt contents at 0–20 cm (r = 0.856, p < 0.05), 40–60 cm (r = 0.880, p < 0.05) and 80–100 cm (r = 0.933, p < 0.05) depths.
In K. caspicum areas, SO42− was the main anion, and its mean concentration increased as soil depth increased, reaching 11.78 g/kg at 80–100 cm, while concentrations were much lower in K. cuspidatum areas (just 1.52 g/kg at 60–80 cm depth) (Figure 5). In areas occupied by the other four species—K. gracile, K. foliatum, K. sinicum, and K. schrenkianum—the mean concentration first declined (from 3.7, 5.4, 3.5 and 3.43 g/kg, respectively, in topsoil) and then increased as depth increased. Overall, it was only correlated with total salt contents at 60–80 cm depth (r = 0.966, p < 0.01) in areas occupied by the six Kalidium species (Figure 5, Table 1).
HCO3− concentrations in soil from areas occupied by all six species were very low. Its mean concentrations first increased then declined as depth increased in K. schrenkianum, K. cuspidatum and K. sinicum areas (Figure 5), and were highest in the 40–60 cm layer of soil in K. sinicum areas, at just 14 mg/kg. Moreover, HCO3− concentrations were negatively correlated with total salt contents at depths below 20 cm in areas occupied by all species (Figure 5, Table 1). CO32− was undetectable with the applied equipment in most samples.
3.4. Germination Rates
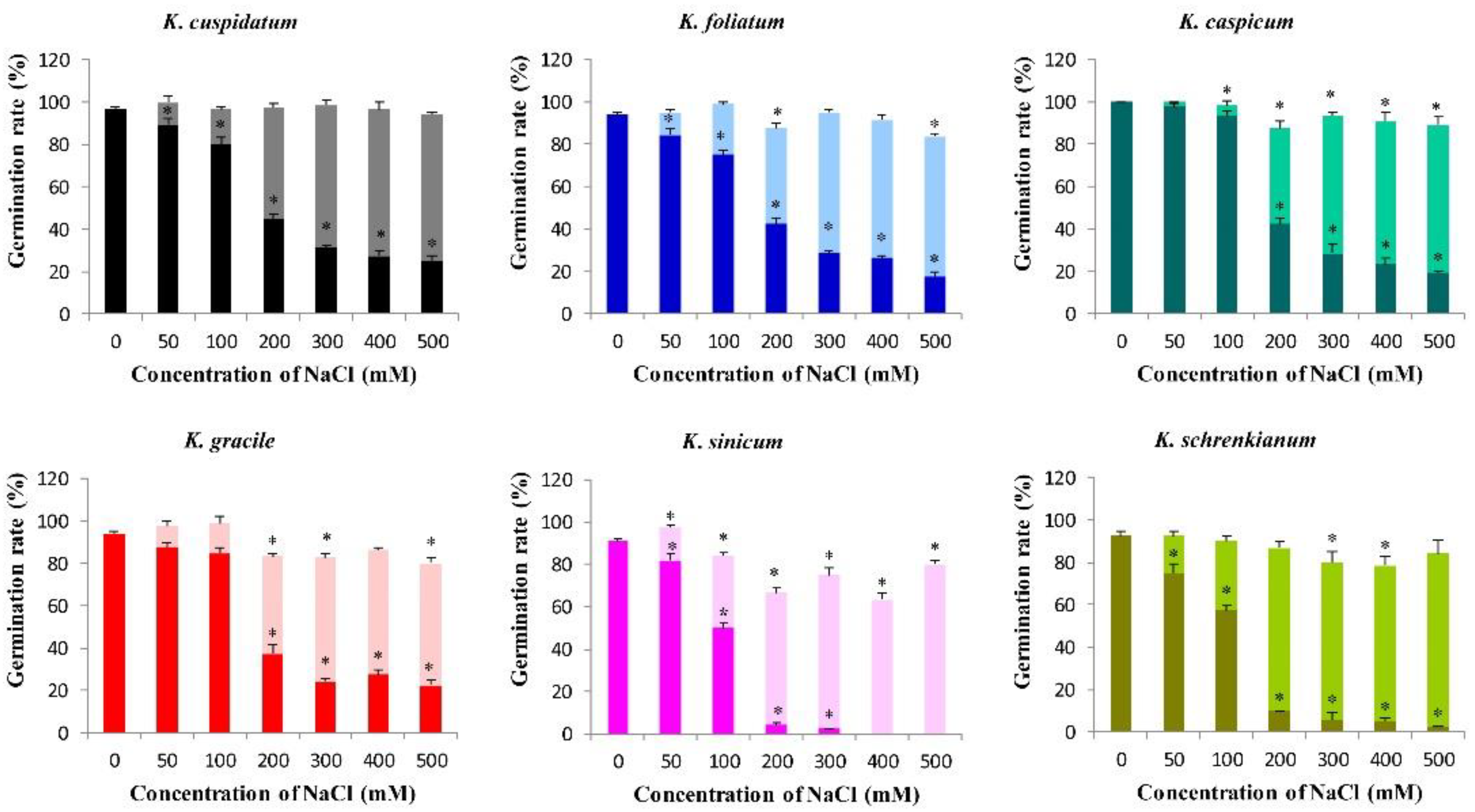
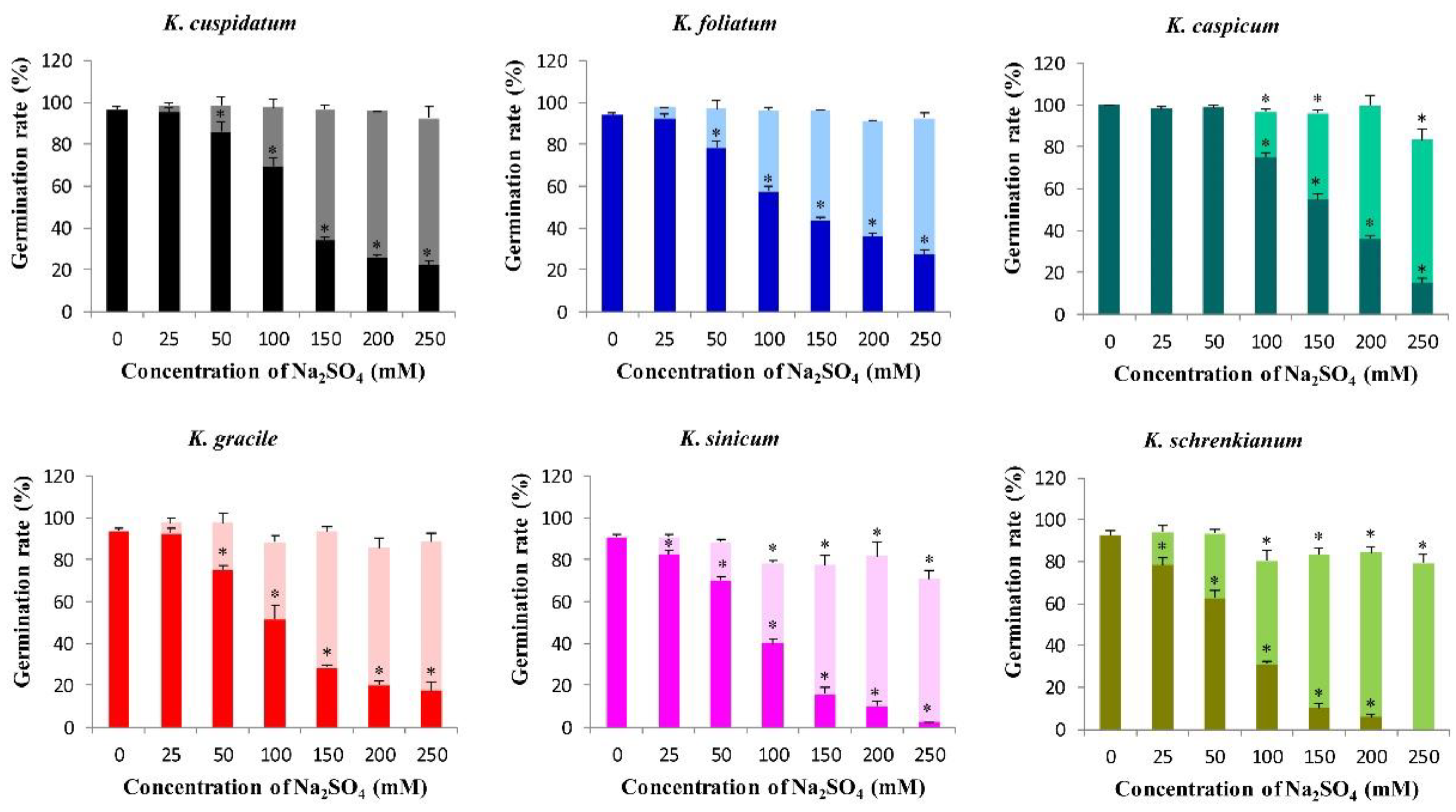
Germination rates of seeds of the six Kalidium species were negatively correlated with concentrations of both NaCl and Na2SO4 in the media (p < 0.05, Figure 6 and Figure 7). In distilled water their germination rates ranged from 90.8% for K. sinicum to 100% for K. caspicum. However, less than half of all species’ seeds germinated when the concentration exceeded 200 mM NaCl, and no K. sinicum seeds germinated at higher concentrations (400 or 500 mM NaCl) (Figure 6). Germination rates also declined with increases in Na2SO4 concentrations in the medium, and at 150 mM exceeded 50% (56.3%) for seeds of only one species (K. caspicum) (Figure 7).
The recovery germination rates of seeds of all six Kalidium species after treatment with both salts were high. The recovery germination rates of K. gracile seeds following the NaCl treatment were negatively correlated with the NaCl concentration during the treatment, declining from 92.2 to 74.2% following exposure to 500 and 200 mM NaCl, respectively (Figure 6). However, the recovery germination rate of K. sinicum seeds exposed to 500 mM NaCl was high (>80%). Moreover, there were no significant differences in recovery germination rates of K. cuspidatum seeds exposed to different NaCl concentrations (p >0.05) and those of the other three species were all above 78% at 500 mM NaCl (Figure 6). Following treatment with Na2SO4, the recovery germination percentages of K. sinicum and K. schrenkianum seeds were lower than those of the other four species at the same concentrations, but were still high (76.6 and 83.2%, respectively), following exposure to the highest Na2SO4 concentration, 250 mM (Figure 7).
All six species produce small seeds (<0.5 g/1000 seeds, Table S1), and there was no linear relationship between the mean mass and germination rate of their seeds.
3.5. Principal Component Analyses
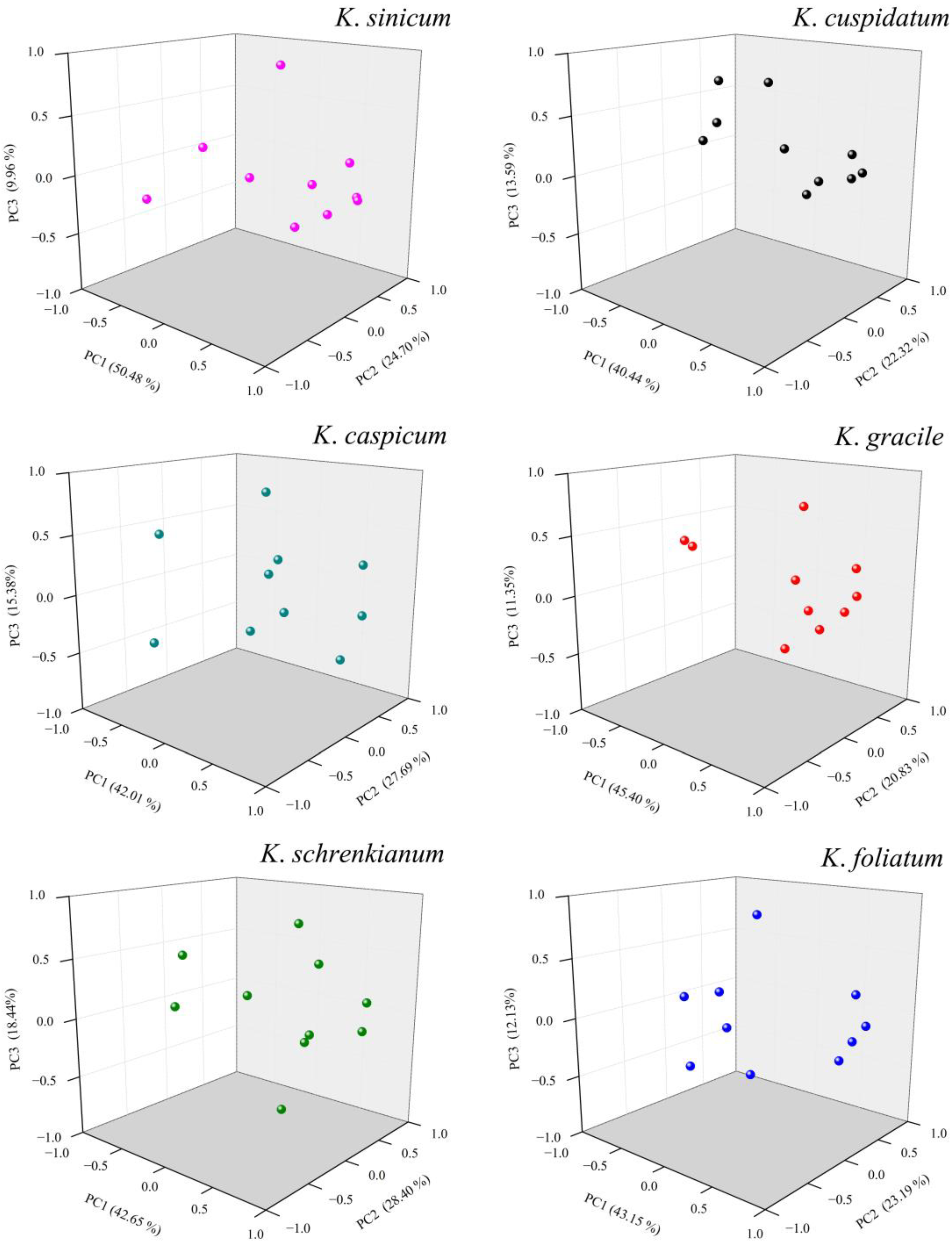
Two soil factors (pH and total salt contents), altitude, four temperature factors and two precipitation factors were used in PCA to explore relationships between abiotic factors and spatial distributions of each of the six Kalidium species (Figure 8 and Table S2). Principal Component (PC1) explained 40.4–50.5% of the variance and was most strongly influenced by temperature factors (maximum temperature of the warmest month, annual mean temperature, and mean temperature of the warmest quarter; Table 2 and Table S2). PC2 explained 20.8–28.4% of the variance and largely reflected effects of precipitation factors on four of the six species. In addition, the two soil factors (total salt contents and pH) strongly contributed to PC3, explaining 9.96–18.4% of the variation in distribution of the six species. In total, PCs 1–3 explained >85% of the variation in the six species’ distributions (Figure 8, Table 2 and Table S2). The results strongly indicate that the most important ecological variables for adaptation to the species’ saline environments are temperature during the driest month and precipitation. They also indicate that the selected ecological variables are strongly associated with the six Kalidium species’ spatial distributions through their effects on the soil environment (Figure 1).
4. Discussion
The mean pH was high (8.9–9.4) in soil samples from areas occupied by all six of the Kalidium species. Mean total salt contents were also high, but covered a substantial range (22.4, 19.5, 17.6, 16.5, 14.3, and 7.08 g/kg in soils from K. cuspidatum, K. gracile, K. foliatum, K. caspicum, K. sinicum, and K. schrenkianum areas, respectively). The indication that K. schrenkianum has relatively low salt tolerance, according to the low mean salt content of samples from areas it occupies, may at least partly explain why this species is restricted to a narrow range to the south of the Tianshan Mountains [18]. In areas occupied by all six species, total salt concentrations were positively related to pH at 0–20 cm and 20–40 cm soil depths (r = 0.794 and r = 0.248, respectively), indicating that salt contents are particularly strongly correlated to pH in topsoil in the study region.
In areas occupied by all the species, Na+ was the main cation, and its concentrations were significantly correlated with total salt contents at all soil depths except 20–40 cm (r = 0.632), showing that they either require high Na+ concentrations for optimal growth and development or at least tolerate them [6]. Ca2+ was the next most abundant cation at 0–40 cm soil depths, indicating that high concentrations of Ca2+ and Na+ likely accumulate in all the Kalidium species, as previously found in roots and leaves of K. foliatum, K. cuspidatum, and various other halophytes [25,26]. As a kind of antagonist, absorption of large amounts of Ca2+ by roots could potentially alleviate damage to plants by other ions [27]. Concentrations of the major nutrient K+, which is required by all living cells and often deficient in barren soil [28,29], ranged from 28.2 to 256.9 mg/kg in soil from K. sinicum and K. cuspidatum areas, respectively. Generally, in soils from areas of all six species the K+ concentration was much lower than the Na+ concentration and (hence) the Na+/K+ ratio was high (>30:1). In addition, there was no significant correlation between the K+ concentration and total salt contents, indicating that the species’ requirements for Na+ and K+ ions significantly differ. Mean Mg2+ concentrations were highest and lowest in soil from K. gracile and K. foliatum areas (ca. 217 mg/kg and 2.3-fold lower, respectively), and substantially lower than concentrations of the other measured cations in all surveyed areas.
The most abundant anion was Cl− in K. cuspidatum areas (where concentrations of both Na+ and Cl− were highest) and SO42− in K. caspicum areas (where Cl− concentrations were lowest). Thus, anions in soils in these areas were strongly dominated by Cl− and SO42−, respectively (mainly balanced in both cases by Na+). These were also the major ions in habitats of the other species, but there was less dominance by Cl− or SO42−, e.g., mean SO42−, Cl− and Na+ concentrations in K. foliatum areas were 5.5, 2.6, and 2.8 g/kg, respectively. There were also wide variations in pH and total salt contents in soil samples from K. foliatum areas, indicating that the species has strong adaptive ability and, thus, can thrive in relatively diverse habits. Concentrations of CO32− were nondetectable and HCO3− concentrations were very low (with no significant differences) in areas of the Kalidium species.
Plants must be sufficiently adapted to the salinity of their environments to germinate [19,30] and establish [31,32,33,34]. In our assays, the germination rates of seeds of the six Kalidium species were all negatively correlated with NaCl and Na2SO4 contents of the medium. Soil salinity fluctuates with precipitation, and can be alleviated in periods with high precipitation, so high proportions of seeds of many halophytes stored in highly saline soil may germinate during such periods [30,35,36]. Moreover, their recovery germination parameters may be major determinants of their distributions. The recovery germination rates of all six Kalidium species exceeded 74% after the NaCl and Na2SO4 treatments, corroborating the conclusions that the quality of topsoil is the first selective barrier affecting plants’ distributions [14,35]. In addition, PCA showed that maximum temperature, summer rainfall and total salt contents of the soil strongly affect geographic distributions of the six Kalidium species. Similarly, distributions and yields of wild barley are clearly related to climatic factors, especially precipitation [37], and distributions of Arabidopsis halleri and A. lyrate are apparently linked to differences in their tolerance of the heavy metals Zn and Cd [38]. Clearly, therefore, ecological factors (and genetically-based adaptations to them) are key determinants of plants’ distributions [18,37,38].
5. Conclusions
Adaptation to topsoil salinity in early stages is a major determinant of the six Kalidium species’ geographic distributions in the study region. The dominant cation at all sites is Na+, but the dominant anions at K. caspicum and K. cuspidatum sites are SO42− and Cl−, respectively. Both salinity and their distributions are affected by numerous interacting factors. Inter alia, temperatures during the driest month and precipitation directly and/or indirectly affect soils’ salt contents and pH, which also strongly influence the six Kalidium species’ distributions. Clearly, high tolerance of salinity stress is a key adaptive trait of halophytes, which has multiple evolutionary origins. Moreover, major changes have occurred in plants’ distributions and population sizes during desertification, following which halophytes may occupy extensive semi-arid and arid regions.
Supplementary Materials
The following supporting information can be downloaded at: https://0-www-mdpi-com.brum.beds.ac.uk/article/10.3390/f13122178/s1:, Table S1: Masses of 1000 seeds of the six Kalidium species; Table S2: Loading matrix of the Principle Components (PCs).
Author Contributions
Y.W. and Z.C. conceived and managed the project; Y.W., D.L. and X.L. performed the experiments; Y.W., Z.C., D.L. and X.L. analyzed the data; Z.C. and D.L. performed statistical analyses; Y.W. and D.L. wrote the original manuscript; and Y.W. and Z.C. reviewed and edited the final manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Central Government Guides Local Scientific and Technological Development Programs of Gansu Province (Grant Number 22ZY2QG001), National Key Research and Development Program of China (Grant Number 2022YFF1303301), the National Natural Science Foundation of China (NSFC, Grant Number 41871092), and Science and Technology Project of Forestry and Grassland Bureau of Gansu Province (Grant Number 2022kj063).
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
We are grateful to Kuibing Meng and Fengzhu Zhang for collecting samples in the field.
Conflicts of Interest
The authors declare that they have no conflict of interest.
References
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheeseman, J.M. The integration of activity in saline environments: Problems and perspectives. Unct. Plant Biol. 2013, 40, 759–774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanin, M.; Ebel, C.; Ngom, M.; Laplaze, L.; Masmoudi, K. New insights on plant salt tolerance mechanisms and their potential use for breeding. Front. Plant Sci. 2016, 7, 1787. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horie, T.; Karahara, I.; Katsuhara, M. Salinity tolerance mechanisms in glycophytes: An overview with the central focus on rice plants. Rice 2012, 5, 11. [Google Scholar] [CrossRef] [Green Version]
- FAO. FAO Soils Portal. 2016. Available online: http://www.fao.org/soils-portal/soil-management/management-of-some-problem-soils/salt-affected-soils/more-information-on-salt-affected-soils/en (accessed on 9 November 2021).
- Flowers, T.J.; Colmer, T.D. Plant salt tolerance: Adaptations in halophytes. Ann. Bot. 2015, 115, 327–331. [Google Scholar] [CrossRef] [Green Version]
- Kachout, S.S.; Mansoura, A.B.; Jaffel, K.; Leclerc, J.C.; Rejeb, M.N. The effect of salinity on the growth of the halophyte Atriplex hortensis (Chenopodiaceae). Appl. Ecol. Env. Res. 2009, 7, 319–332. [Google Scholar] [CrossRef]
- Shabala, S. Learning from halophytes: Physiological basis and strategies to improve stress tolerance in crops. Ann. Bot. 2013, 112, 1209–1221. [Google Scholar] [CrossRef]
- Waisel, Y. Biology of Halophytes; Academic Press: New York, NY, USA, 1972. [Google Scholar]
- Parida, A.K.; Jha, B. Salt tolerance mechanisms in mangroves: A review. Trees 2010, 24, 199–217. [Google Scholar] [CrossRef]
- Zhao, K.F.; Fan, H.; Jiang, X.Y.; Song, J. Improvement and utilization of saline soil by planting halophytes. Chin. J. Appl. Env. Biol. 2002, 8, 31–35. [Google Scholar]
- Cram, W.J. Negative feedback regulation of transport in cells. The maintenance of turgor, volume and nutrient supply. In Encyclopaedia of Plant Physiology; Luttge, U., Pitman, M.G., Eds.; Springer: Berlin, Germany, 1976; pp. 284–316. [Google Scholar]
- Richau, K.H.; Schat, H. Intraspecific variation of nickel and zinc accumulation and tolerance in the hyperaccumulator Thlaspi caerulescens. Plant and Soil 2009, 314, 253–262. [Google Scholar] [CrossRef] [Green Version]
- Turner, T.L.; Bourne, E.C.; Von Wettberg, E.J.; Hu, T.T.; Nuzhdin, S.V. Population resequencing reveals local adaptation of Arabidopsis lyrata to serpentine soils. Nat. Genet 2010, 42, 260–263. [Google Scholar] [CrossRef] [PubMed]
- Lazarus, B.E.; Richards, J.H.; Claassen, V.P.; O’Dell, R.E.; Ferrell, M.A. Species-specific plant-soil interactions influence plant distribution on serpentine soils. Plant and Soil 2011, 342, 327–344. [Google Scholar] [CrossRef] [Green Version]
- Piirainen, M. Kalidium (Chenopodiaceae) the information resource for Euro-Mediterranean plant diversity (Uotila, P., Eds.). Eur. Med. Plantbase 2009, 13, 146. [Google Scholar]
- Liang, X.H.; Wu, Y.X. Identification of Kalidium species (Chenopodiaceae) by DNA barcoding. Sci. Cold. Ari. Regi. 2017, 9, 89–96. [Google Scholar]
- Flowers, T.J.; Galal, H.K.; Bromham, L. Evolution of halophytes: Multiple origins of salt tolerance in land plants. Funct. Plant Biol. 2010, 37, 604–612. [Google Scholar] [CrossRef]
- Tobe, K.; Li, X.M.; Omasa, K. Seed germination and radicle growth of a halophyte, Kalidium caspicum (Chenopodiaceae). Ann. Bot. 2000, 85, 391–396. [Google Scholar] [CrossRef] [Green Version]
- Song, J.; Feng, G.; Zhang, F.S. Salinity and temperature effects on germination for three salt resistant euhalophytes, Halostachys caspica, Kalidium foliatum and Halocnemum Strobilaceu. Plant and Soil 2006, 279, 201–207. [Google Scholar] [CrossRef]
- Haris, S.; Xia, H.; Elisabeth, B.; Camile, M.; Lindell, B. Predicting species’ tolerance to salinity and alkalinity using distribution data and geochemical modelling: A case study using Australian grasses. Ann. Bot. 2015, 115, 343–351. [Google Scholar]
- Lu, R.K. Analytical Methods for Soil and Agro-Chemistry; China Agricultural Science and Technology Press: Beijing, China, 1999. [Google Scholar]
- Li, R.A.; Wang, F.; Qin, F.J.; Lou, F.; Wu, D.Y. Studies on the best curve equation between the total salts and the electrical conductivity of the coastal saline soil. J. Agric. 2015, 5, 59–62. [Google Scholar]
- Wang, W.X.; Li, Z.Z.; Chang, Z.Q. Principal component analysis of geological factors related to landforms-hydrological system of Qilian mountain region. Acta Bot. Boreali-Occident. Sin. 2004, 24, 533–537. (In Chinese) [Google Scholar]
- Tobe, K.; Li, X.M.; Omasa, K. Effects of sodium, magnesium and calcium salts on seed germination and radicle survival of a halophyte, Kalidium caspicum (Chenopodiaceae). Aust. J. Bot. 2002, 50, 163–169. [Google Scholar] [CrossRef] [Green Version]
- Gao, R.R.; Zhao, R.H.; Du, X.M.; Huang, Z.Y.; Yang, X.J.; Wei, X.Z.; Huang, P.Y. Characteristics of root systems of two halophytes for adaptability to salinity. Sci. Silvae Sin. 2010, 46, 176–182. [Google Scholar]
- Gul, B.; Khan, M.A. Role of calcium in alleviating salinity effects in coastal halophytes. In Ecophysiology of High Salinity Tolerant Plants; Khan, M.A., Weber, D.J., Eds.; Springer: Netherlands, Switzerland, 2006; pp. 107–114. [Google Scholar]
- Very, A.A.; Sentenac, H. Molecular mechanisms and regulation of K+ transport in higher plants. Annu. Rev. Plant Biol. 2003, 54, 575–603. [Google Scholar] [CrossRef]
- Shabala, S.; Cuin, T.A. Potassium transport and plant salt tolerance. Physiol. Plant. 2007, 133, 651–669. [Google Scholar] [CrossRef] [PubMed]
- El-Keblawy, A.; Elnaggar, A.; Tammam, A.; Mosa, K.A. Seed provenance affects salt tolerance and germination response of the habitat-indifferent Salsola drummondii halophyte in the arid Arabian deserts. Flora 2020, 266, 151592. [Google Scholar] [CrossRef]
- Attia, H.; Al-Yasi, H.; Alamer, K.; Esmat, F.; Hassan, F.; Elshazly, S.; Hessini, K. Induced anti-oxidation efficiency and others by salt stress in Rosa damascena Miller. Sci. Hortic. 2020, 274, 109681. [Google Scholar] [CrossRef]
- Yasir, T.A.; Khan, A.; Skalicky, M.; Wasaya, A.; Rehmani, M.; Sarwar, N.; Mubeen, K.; Aziz, M.; Hassan, M.M.; Hassan, F.A.S. Exogenous sodium nitroprusside mitigates salt stress in lentil (Lens culinaris medik.) by affecting the growth, yield, and biochemical properties. Molecules 2021, 26, 2576. [Google Scholar] [CrossRef]
- Hassan, F.; Al-Yasi, H.; Ali, E.F.; Alamer, K.; Hessini, K.; Attia, H.; El-Shazly, S. Mitigation of salt stress effects by moringa leaf extract or salicylic acid through motivating antioxidant machinery in damask rose. Can. J. Plant Sci. 2021, 101, 157–165. [Google Scholar] [CrossRef]
- Hassan, F.; Ali, E.; Gaber, A.; Fetouh, M.I.; Mazrou, R. Chitosan nanoparticles effectively combat salinity stress by enhancing antioxidant activity and alkaloid biosynthesis in Catharanthus roseus (L.) G. Don. Plant Physiol. Biochem. 2021, 162, 291–300. [Google Scholar] [CrossRef]
- Bhatt, A.; Gallacher, D.J.; Jarma-Orozco, A.; Fernandes, D.; Pompelli, M.F. Seed provenance selection of wild halophyte seeds improves coastal rehabilitation efficiency. Estuar. Coast. Shelf Sci. 2021, 265, 107657. [Google Scholar] [CrossRef]
- Vicente, O.; Boscaiu, M.; Naranjo, M.O.; Estrelles, E.; Bellés, J.M.; Soriano, P. Responses to salt stress in the halophyte Plantago crassifolia (Plantaginaceae). J. Arid Environ. 2004, 58, 463–481. [Google Scholar] [CrossRef]
- Cronin, J.K.; Bundock, P.C.; Henry, R.J.; Nevo, E. Adaptive climatic molecular evolution in wild barley at the Isa defense locus. PNAS 2007, 104, 2773–2778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alcazar, R.; Pecinka, A.; Aarts, M.G.; Fransz, P.F.; Koornneef, M. Signals of speciation within Arabidopsis thaliana in comparison with its relatives. Curr. Opin. Plant Biol. 2012, 15, 205–211. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Map showing soil sampling sites in areas occupied by the six Kalidium species. Stars indicate seed sampling sites.
Figure 1.
Map showing soil sampling sites in areas occupied by the six Kalidium species. Stars indicate seed sampling sites.

Figure 2.
Scatter plots of total salt contents and pH of soil samples from areas occupied by indicated Kalidium species.
Figure 2.
Scatter plots of total salt contents and pH of soil samples from areas occupied by indicated Kalidium species.

Figure 3.
Linear regression plots showing the relationship between total salt contents and pH at indicated depths, and overall, in soil from areas occupied by the six Kalidium species. (a) total salt contents at different soil depth; (b) total salt contents at different species.
Figure 3.
Linear regression plots showing the relationship between total salt contents and pH at indicated depths, and overall, in soil from areas occupied by the six Kalidium species. (a) total salt contents at different soil depth; (b) total salt contents at different species.

Figure 4.
Concentrations of Na+, Ca2+, K+, and Mg2+ in soil from indicated depths in areas occupied by the six Kalidium species. The values shown are means with SE (n = 3).
Figure 4.
Concentrations of Na+, Ca2+, K+, and Mg2+ in soil from indicated depths in areas occupied by the six Kalidium species. The values shown are means with SE (n = 3).

Figure 5.
Concentrations of Cl−, SO42−, and HCO3− in soil from indicated depths in areas occupied by the six Kalidium species. The values shown are means with SE (n = 3).
Figure 5.
Concentrations of Cl−, SO42−, and HCO3− in soil from indicated depths in areas occupied by the six Kalidium species. The values shown are means with SE (n = 3).

Figure 6.
Total germination percentages after 14 days at the indicated NaCl concentrations in the six studied Kalidium species. Germination rates (%, means and standard deviations) of the six Kalidium species after treatments with indicated concentrations of NaCl (0, 50, 100, 200, 300, 400, and 500 mM NaCl) after 7 days. The dark bars indicate germination rates after the treatments and light bars the total percentages that germinated during the recovery treatment in distilled water with additional 7 days. Asterisks indicate significant differences in each condition respect to the corresponding control (according to Dunnet test, p < 0.05).
Figure 6.
Total germination percentages after 14 days at the indicated NaCl concentrations in the six studied Kalidium species. Germination rates (%, means and standard deviations) of the six Kalidium species after treatments with indicated concentrations of NaCl (0, 50, 100, 200, 300, 400, and 500 mM NaCl) after 7 days. The dark bars indicate germination rates after the treatments and light bars the total percentages that germinated during the recovery treatment in distilled water with additional 7 days. Asterisks indicate significant differences in each condition respect to the corresponding control (according to Dunnet test, p < 0.05).

Figure 7.
Total germination percentages after 14 days at the indicated Na2SO4 concentrations in the six studied Kalidium species. Germination rates (%, means and standard deviations) of the six Kalidium species after treatments with indicated concentrations of Na2SO4 (0, 25, 50, 100, 150, 200, and 250 mM) after 7 days. The dark bars indicate germination rates after the treatments and light bars the total percentages that germinated during the recovery treatment in distilled water with additional 7 days. Asterisks indicate significant differences in each condition respect to the corresponding control (according to Dunnet test, p < 0.05).
Figure 7.
Total germination percentages after 14 days at the indicated Na2SO4 concentrations in the six studied Kalidium species. Germination rates (%, means and standard deviations) of the six Kalidium species after treatments with indicated concentrations of Na2SO4 (0, 25, 50, 100, 150, 200, and 250 mM) after 7 days. The dark bars indicate germination rates after the treatments and light bars the total percentages that germinated during the recovery treatment in distilled water with additional 7 days. Asterisks indicate significant differences in each condition respect to the corresponding control (according to Dunnet test, p < 0.05).

Figure 8.
Score plots obtained from Principal Component Analysis (PCA) of effects of abiotic factors on distributions of the six Kalidium species. x axis, y axis, and z axis indicate the first, the second and the third Principle Component, respectively.
Figure 8.
Score plots obtained from Principal Component Analysis (PCA) of effects of abiotic factors on distributions of the six Kalidium species. x axis, y axis, and z axis indicate the first, the second and the third Principle Component, respectively.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Correlation coefficient (r) between total salt contents and concentrations of indicated ions in soil from indicated depths in areas occupied by the six Kalidium species.
Table 1.
Correlation coefficient (r) between total salt contents and concentrations of indicated ions in soil from indicated depths in areas occupied by the six Kalidium species.
| Soil Depth | Ca2+ | Mg2+ | K+ | Na+ | HCO3− | Cl− | SO42− |
|---|---|---|---|---|---|---|---|
| 0~20 cm | −0.841 * | 0.725 | 0.018 | 0.896 * | 0.992 ** | 0.856 * | 0.793 |
| 20~40cm | −0.093 | −0.156 | −0.680 | 0.632 | −0.969 ** | 0.689 | 0.031 |
| 40~60 cm | 0.677 | 0.980 ** | −0.140 | 0.986 ** | −0.809 | 0.880 * | 0.784 |
| 60~80 cm | 0.934 * | 0.991 ** | 0.567 | 0.972 ** | −0.873 | 0.933 * | 0.966 ** |
| 80~100 cm | 0.965 ** | −0.958 * | 0.660 | 0.973 ** | −0.502 | 0.167 | 0.344 |
*, p < 0.05; **, p < 0.01.
Table 2.
Loadings of the main factors influencing the first three Principle Components obtained from Principle Component Analysis of the relationships between abiotic factors and distributions of the six Kalidium species.
Table 2.
Loadings of the main factors influencing the first three Principle Components obtained from Principle Component Analysis of the relationships between abiotic factors and distributions of the six Kalidium species.
| Species | Main Factors and Loadings (Correlation Coefficients) | ||
|---|---|---|---|
| PC1 | PC2 | PC3 | |
| K. schrenkianum | bio5 (0.971) | pH (0.797) | TS (0.719) |
| K. sinicum | bio9 (0.975) | bio14 (0.812) | TS (0.906) |
| K.cuspidatum | bio5 (0.892) | bio14 (0.707) | TS (0.766) |
| K. capsicum | bio1 (0.949) | bio14 (0.870) | pH (0.833) |
| K. gracile | bio1 (0.993) | bio12 (0.832) | bio14 (0.658) |
| K. foliatum | bio9 (0.924) | bio5 (0.713) | TS (0.840) |
bio1, annual mean temperature; bio5, max temperature of the warmest month; bio9, mean temperature of the driest quarter; bio12, annual average precipitation; bio14, precipitation of the driest month; TS, total salt contents.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Liu, D.; Chang, Z.; Liang, X.; Wu, Y. Soil Chemical Properties Strongly Influence Distributions of Six Kalidium Species in Northwest China. Forests 2022, 13, 2178. https://0-doi-org.brum.beds.ac.uk/10.3390/f13122178
AMA Style
Liu D, Chang Z, Liang X, Wu Y. Soil Chemical Properties Strongly Influence Distributions of Six Kalidium Species in Northwest China. Forests. 2022; 13(12):2178. https://0-doi-org.brum.beds.ac.uk/10.3390/f13122178
Chicago/Turabian StyleLiu, Decheng, Zongqiang Chang, Xiaohui Liang, and Yuxia Wu. 2022. "Soil Chemical Properties Strongly Influence Distributions of Six Kalidium Species in Northwest China" Forests 13, no. 12: 2178. https://0-doi-org.brum.beds.ac.uk/10.3390/f13122178
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.