

How to Capture Thousands of Genotypes—Initiation of Somatic Embryogenesis in Norway Spruce

Natural Resources Institute Finland (Luke), Vipusenkuja 5, FI-57200 Savonlinna, Finland
*
Author to whom correspondence should be addressed.
Forests 2023, 14(4), 810; https://0-doi-org.brum.beds.ac.uk/10.3390/f14040810
Submission received: 14 March 2023
/
Revised: 11 April 2023
/
Accepted: 13 April 2023
/
Published: 14 April 2023
(This article belongs to the Special Issue Application of Plant Biotechnology in Forestry)
Abstract
:Somatic embryogenesis (SE) is considered the most effective method for vegetative propagation of Norway spruce (Picea abies L. Karst). When the aim is commercial production, the process needs scaling up. This includes many initiations to increase the number of available genotypes in the cryo-bank. Numerous genotypes are needed to maintain genetic diversity in reforestation and, at the same time, are a prerequisite for the efficient improvement of breeding traits. Norway spruce is also highly susceptible to Heterobasidion root rot. We analysed the data from the SE initiations of Norway spruce from six different years, including a total of 126 families and almost 13,000 initiations, and used several genetic (including allele PaLAR3B improving Heterobasidion resistance), environmental, and operational variables to explain the initiation success and the number of cryopreserved embryogenic tissue (ET). Overall, the cone collection date was the best and most comprehensive single variable for predicting the initiation success and the number of cryopreserved ET in the logistic regression models. PaLAR3B allele did not interfere with SE initiation or the cryopreservation. In the optimal scenario, according to the current data, Norway spruce cones would be collected in southern Finland during the first two weeks of July (in approximately 800 d.d. accumulation) from the seed orchard or greenhouse and delivered quickly to the laboratory, and the cones would be cold-stored for five days or less before initiations on mLM media. Lower initiation frequencies in some families can be compensated by increasing the number of explants—however, taking operational limitations into account.
1. Introduction
To increase forest growth and, thus, increase the possibility of their sustainable use, the best possible regeneration material, i.e., superior tree genotypes, should be used [1]. To ensure the availability of good-quality forest regeneration material, effective vegetative propagation methods such as somatic embryogenesis (SE) were introduced [2]. SE must provide improved genetic gain to be utilised in forestry and breeding, i.e., SE plants must be proven to be superior to unimproved seedlings (at least) to support the costs of production [3]. Genetic gain can be proven by field testing [4].
From the practical perspective, establishing field trials with SE plants requires numerous genotypes per family to ensure the possibility of genotype selection and to avoid reduced genetic gain [4,5,6,7]. To obtain the required number of plants for field testing, well-performing cell lines should be initiated, i.e., lines that have vigorous proliferation, survive cryopreservation, and produce a large number of somatic embryos after cryopreservation [5,6]. Moreover, embryos must germinate and perform well in the nursery. For this, many initiations, 60 to 80 explants per family, from several families must be undertaken [7]. These preceding requirements regarding field testing combined with irregular flowering of Norway spruce in Finland lay out the framework for the magnitude of annual initiations for thousands rather than hundreds [8]. Labour and financial resources are often limited and to optimise successful SE propagation, variables affecting initiation should be carefully analysed.
Genetic variation in the outcome of SE initiations was addressed in white spruce (Picea glauca) [9,10], loblolly pine (Pinus taeda) [11], and black spruce (Picea mariana) [12,13]. Park et al. [9] found that the initiation phase of SE was under strong genetic control, but genetic influence declined steadily through the proliferation, maturation, and germination phases [2]. In Norway spruce, differences in the initiation rates and establishment of embryogenic cultures were reported among families [14]. However, to estimate genetic parameters for the initiation capacity of a population, much larger samples are needed than those reported above [13].
SE propagation enables the efficient propagation of selected genotypes with desired traits. Norway spruce is highly susceptible to Heterobasidion root rot, and the pressure of disease is predicted to increase with ongoing climate change [15]. Therefore, an effort was made to identify more root-rot-tolerant genotypes through genome-based tools as marker-assisted selection [16,17]. It was shown that in the presence of leucoanthocyanidin reductase, PaLAR3B allele root rot spread slowed down by 27% [16]. In a previous study, we showed that the resistance allele Palar3B is successfully delivered through the SE process [18]. The scale of the present study enables us to study whether this allele interferes with the SE initiation and cryopreservation process.
The developmental stage of explants had a significant effect on the initiation of Norway spruce [19], black spruce [20], and white spruce [9]. The optimal stage depends on the species. For example, for maritime pine (Pinus pinaster), the precotyledonary stage of embryo development was the most responsive [21], and for jack pine (Pinus banksiana), only embryos at the polyembryonic cleavage stage will respond [22]. The initiation rate gradually diminishes as the zygotic embryo matures, although at the earliest stage, immature ZEs may not be capable of forming embryogenic tissue (ET) [2,9,19]. Various environmental factors and genetic variability affect ZE development, making it difficult to predict the right moment for cone collection.
Due to limited labour resources, immature cones are usually cold-stored before preparations. However, cold storage may reduce initiation potential remarkably, as observed in Park’s study [9]. Another storage option is to keep sterilised immature seeds in sterile water in the cold prior to initiation, but, to our knowledge, there are no previous studies about the effect of this.
Many studies were conducted on optimising SE initiation media for different species [23,24]. In spruce, the media for SE are usually based either on those described by von Arnold and Eriksson (LP) [25], Litvay et al. (LM) [26], or Murashige-Skoog medium (MS) [27]. Modifications were made, for example, by reducing the amount of macroelements [14,28]. Media are supplemented with organic nitrogen (L-glutamine and casein hydrolysate), and typically contain 2,4-dichlorophenoxyacetic acid (2,4-D) at 9.0–10 M, benzyl adenine (BA) at 4.5–5.0 M, sucrose at 1 or 2%, and are solidified with agar (0.8%) or gellan gum (0.4%) [28]. In some cases, proliferation media may need changes especially in the concentrations of plant growth regulators [23,29], but media of the same composition are usually used [28].
Proliferating ET should be cryopreserved when enough culture is grown to avoid losing ET’s embryogenic potential and save resources [14]. The criteria for cryopreservation should be the growth rate to avoid the delay caused by waiting for the results of embryo productivity testing [14,30].
In this study, we utilise the extensive data from SE initiations of Norway spruce made in several years to identify the genetic, environmental, and operational variables affecting initiation and proliferation up to cryostorage. Several studies were previously conducted on factors affecting the SE success of Norway spruce and other coniferous species, but this study covers families and initiations among families more than previous ones. Compared to other studies, this unique dataset is also extended over a decade. To our knowledge, this is the first study in which such extensive SE initiation and cryopreservation data are analysed for root rot tolerance related to the genetic component for resistance breeding. Our findings will help optimise the successful initiation and cryopreservation of Norway spruce using the SE technique.
2. Materials and Methods
2.1. Plant Material
Embryogenic lines of Norway spruce were initiated from immature and mature seed embryos originating from the controlled crossings of a Finnish tree breeding programme (Supplementary Table S1). Crossings were made in 2011, 2012, 2014, 2015, 2019, and 2022 using seed orchard grafts in southern Finland (60°55′ N, 26°13′ E, 80 m, 60°41′ N, 24°02′ E, 130 m) or grafted trees in a greenhouse. Grafted trees and pollen donors originated from different locations in southern and central Finland. From 2011 to 2015, the plus trees used in the SE were chosen based on the flowering of the grafts and the sufficiency of cones. In 2019, the measured progeny testing results (unpublished data) were available for selection [31]. In 2022, the plus trees were genotyped and chosen based on the root rot resistance marker PaLAR3 alleles [16,18]. Over the years, a total of 126 crossing combinations (families) were made, using 69 mother and 85 father genotypes. Three combinations were repeated in two different years, and 14 mothers and 13 fathers were used in more than one year.
Controlled crossings were conducted on 7 May at the earliest in 2019, and on 17 May at the latest in 2011 and 2014. Immature cones were collected at the earliest on 24 June in the summer (2019) and at the latest on 9 August (2012). Mature cones were collected in November or December (2011 and 2012).
Days between the first pollination day and cone collection were used to describe the development of ZE. In addition, the temperature sum accumulation in degree days (d.d.) was used to describe the timing of cone collection. The d.d. was calculated by summing the daily mean temperatures exceeding the threshold value of 5 °C from 1 January of that year. Temperature sums were available from 2011, 2012, 2014 (seed orchards), and 2019 (greenhouse).
2.2. Initiation, Proliferation, and Cryopreservation
Embryogenic lines were initiated and proliferated by applying the methods developed by Klimaszewska et al. [32] and Lelu-Walter et al. [33] and described in Varis et al. [34]. In short, immature cones were wiped with 70% ethanol, seeds were dissected in sterile water, and washing-up liquid (15%–30% anionic surfactants, 5%–15% non-ionic surfactants, benzisothiazolinone, methylisothiazolinone, phenoxyethanol, perfumes) was added to clean the seeds. After one rinse in sterile water, the seeds were surface-sterilised in 70% ethanol for 5 min and rinsed three times in sterilised water. The ZEs were dissected from megagametophytes and placed on modified Litvay’s medium (mLM) [26,32] containing 10 µM 2,4-dicholophenoxyacetic acid (2,4-D) and 5 µM 6-benzyladenine (BA) as plant growth regulators (Supplementary Table S2). The sucrose concentration of the medium was 0.03 M. The pH of the medium was adjusted to 5.8 prior to adding gelling agent (Phytagel 4 g/L) and sterilisation in the autoclave. After autoclaving at 120 degrees for 20 min, the medium was cooled to 60 °C, and 500 mg/L of L-glutamine was added using filter sterilising.
In 2011, half the initiations were placed on 1/2LP medium [25], modified according to [14], containing 9 µM 2.4-D and 4.4 µM BA (Supplementary Table S2). In both cases, Petri dishes (9 cm in diameter) were filled with 20 mL of the medium. Ten ZEs were placed on the same Petri dish, and they were kept in the dark and at 24 °C. The ZEs were kept on the same media until ET started to grow and then transferred to a fresh medium, each genotype on a separate dish. The established ETs were subcultured bi-weekly. The proliferation media was the same as that used for culture initiation.
Embryogenic lines were chosen for cryopreservation based on their embryo productivity (2011 and 2012) or good growth in proliferation (2014 and since). In 2011, the two-day pre-treatment method in liquid mLM media described by Kartha [35] and modified by Find et al. [36] was applied to cryopreserve six samples from every 260 cell lines. From 2012 to 2019, a two-day pre-treatment method in semi-solid media was used as described in Varis et al. [34], and in 2022, the pre-treatment method was simplified to a one-day dehydration in semi-solid media [37]. After pre-treatment, 1.5 mL suspended ET was placed in a 2 mL sterile cryovial, or 200 mg of ET from semi-solid media was placed in sterile cryovials containing 400 mL of liquid mLM medium with 0.4 M of sucrose without plant growth regulators (PGR) or glutamine, after which 400 mL of preservative mixture containing polyethylene glycol 6000, glucose, and DMSO 10% w/v each were added. The cryovials were cooled at the rate of 0.17 °C/min to −38 °C in a programmable cooling device (Planer, Kryo 10 Series III, Planer Products, Middlesex, UK), followed by immersion in LN.
2.3. Cone and Seed Cold Storage
Because of the limited human resources or for experimental reasons, some of the cones were stored in a cold room at +2 °C. The cones were in paper bags and stored from 1 to 38 days. For the same reason, sterilised seeds were left in the last sterile rinsing water and stored in sterile water in a refrigerator at +4 °C from 1 to 22 days.
When the effect of storing cones in the cold room was analysed, initiations in which the seed was also stored in cold water were not used. Because there was an unbalanced number of families between cone storing days, the data were merged into groups of five days. When the effect of seed storage in cold water was described, only data in which cones were cold-stored for one to seven days were used.
2.4. Statistical Analyses
The initiation was defined as successful when the ET was growing and could be separated from the ZE and moved to its own medium. Each initiation was counted as successful (1) or unsuccessful (0). Cryopreserved ET is the number of initiations which ended up in cryopreservation. The variables which were investigated reflecting initiation and cryopreservation success were categorised in three groups of variables: genetic; environmental; and operational. The genetic variables investigated were: full-sib family, father tree, mother tree, PaLAR3 cross type PaLAR3 father, PaLAR3 mother. Because mother and father PaLAR3 genotypes AA, AB, and BB were available in different years (Supplementary Table S1), the PaLAR3 cross types of the mother × father were AA × AA, AA × AB, AB × AA, AB × AB, AB × BB, i.e., BB × BB was missing.
The environmental variables were year of initiation, pollination date, cone collection d.d. and date, and time period from pollination to cone collection and to initiation. The operational variables tested were location of the mother graft (in a seed orchard or in a greenhouse), mature or immature ZEs, duration of cone or seed cold storage before initiation, initiation date, and initiation media lot.
The data in which ½LP was used as an initiation medium were included only when the initiation rate between different media was analysed but excluded from further analysis. Cryopreservation data from 2011 and 2012 was excluded from the analysis because ETs for cryopreservation were selected based on embryo production capacity, i.e., following maturation experiments, and the material was, therefore, not comparable with the ETs selected for their vigorous proliferation as in later years.
Differences in initiation and cryopreservation success between seed orchard and greenhouse grafts, initiation success between mature and immature seeds, and the effect of the PaLAR3 genotype were analysed by the Pearson chi-square test. The Pearson correlation test was used to analyse the correlation between the number of initiations in the full-sib family and initiation success %.
The cryopreservation percentages were calculated as cryopreserved ET/initiated explants × 100 if not mentioned otherwise. Binary logistic regression (log(p/1 − p)) modelling was used to estimate the effects of genetic, environmental, and operational factors on the initiation success and cryopreserved ET, because they are 0/1 data. Variables were added to the model if values were available throughout the data (although several were included with missing values, which was considered in the selection of final models and in comparison of variables).
In the final models, only cone collection date was used as an experimental variable, as it was the best (and most comprehensive) single variable predicting initiation and cryopreservation success in the overall data. When more than one variable was accepted for the model, the best model response was achieved including full-sib family as a genetic variable and the initiation date as an operational variable for the models (Supplementary Table S3).
3. Results
3.1. Genetic Variables in Initiation and Cryopreserved ET
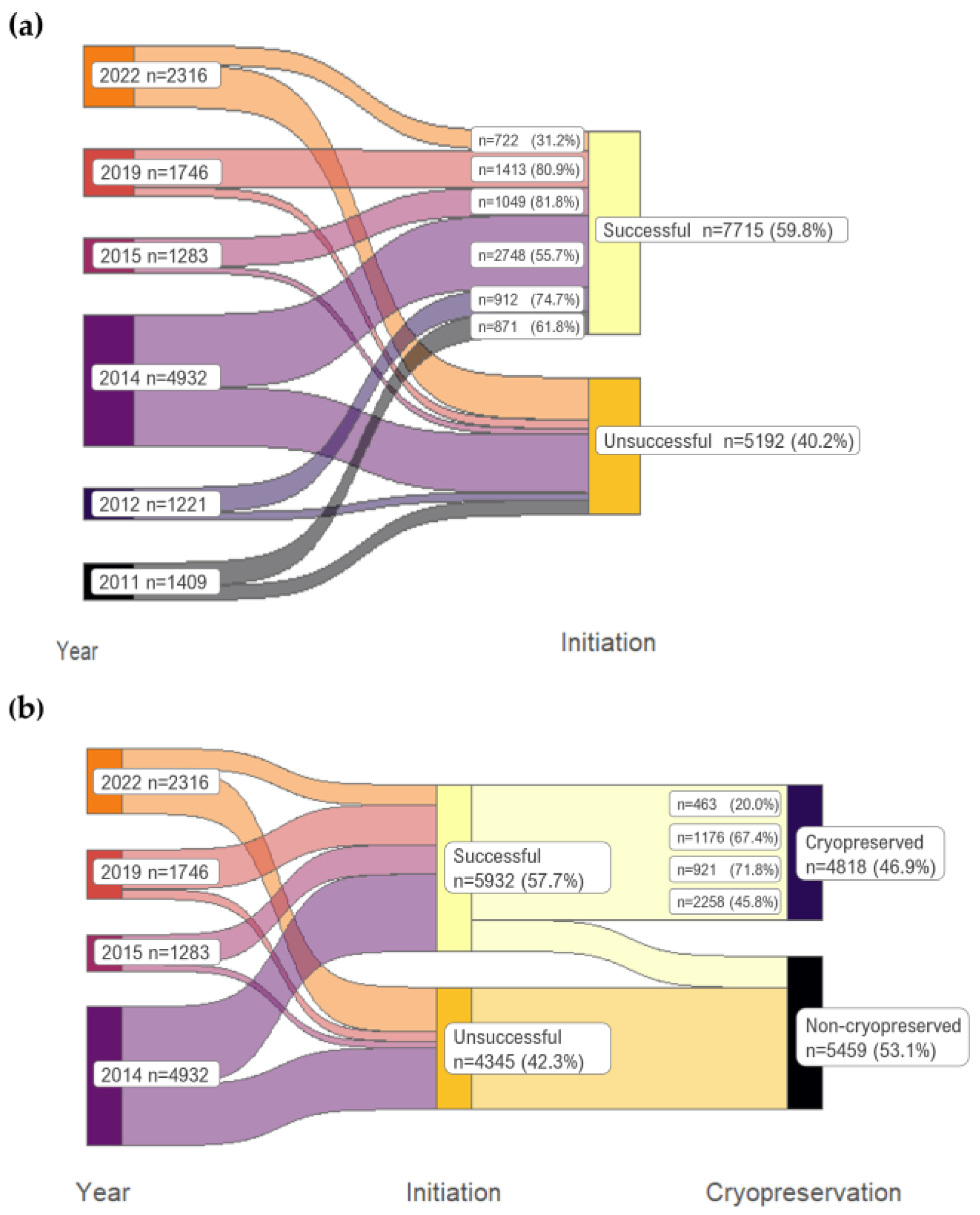
Embryogenic cell lines were obtained from 118 full-sib families out of the 126 (Supplementary Table S1). ET initiation % varied from 0 to 100 among full-sib families (Supplementary Table S1), and the mean ET initiation % of all 12,907 initiations was 59.8% (Figure 1a). The ET initiation % of the full-sib family was positively correlated with the number of initiations made (Pearson correlation 0.233, p < 0.01). Not all ET continued growing vigorously, and in 2014–2022, 46.9% of the original initiations ended up in cryopreservation (=81.2% of the succeeded initiations, i.e., growing cell lines) (Figure 1b).
In the logistic regression model, the full-sib family, father tree, and mother tree were significant (p < 0.01, in all). The full-sib family improved the model most by increasing the overall % of cases predicted correctly to 69.3% (improvement in the model 9.5 units of %). The father tree and mother tree resulted in 68% (improvement in the model 8.2 units of %) and 65.2% (improvement in the model 5.4 units of %) of cases predicted correctly, respectively.
The effect of the PaLAR3 cross type, PaLAR3 father and PaLAR3 mother, was significant (p < 0.01, in all both in initiation and cryopreservation). However, the improvements in the initiation model were only 1.5, 0.0, and 0.2 units of %, respectively. The effect of PaLAR3 variables was similar in the model of cryopreserved ET. The PaLAR3 cross type, PaLAR3 father and PaLAR3 mother, improved the model by 8.4, 4.8, and 6.1 units of %. Additionally, when PaLAR3 variables were included in the model with the full-sib family, the PaLAR3 variables did not improve the model at all.
The effect of PaLAR3 genotype was further studied using the chi-squared test of independence. The PaLAR3 cross type and allele frequency of root rot resistance-related B-allele had a significant effect on the initiation and cryopreserved ET when the data from all years were pooled. However, when these variables were tested for each year separately, the PaLAR3 genotype had no effect on the initiation and cryopreserved ET result.
3.2. Environmental Variables in Initiation and Cryopreserved ET
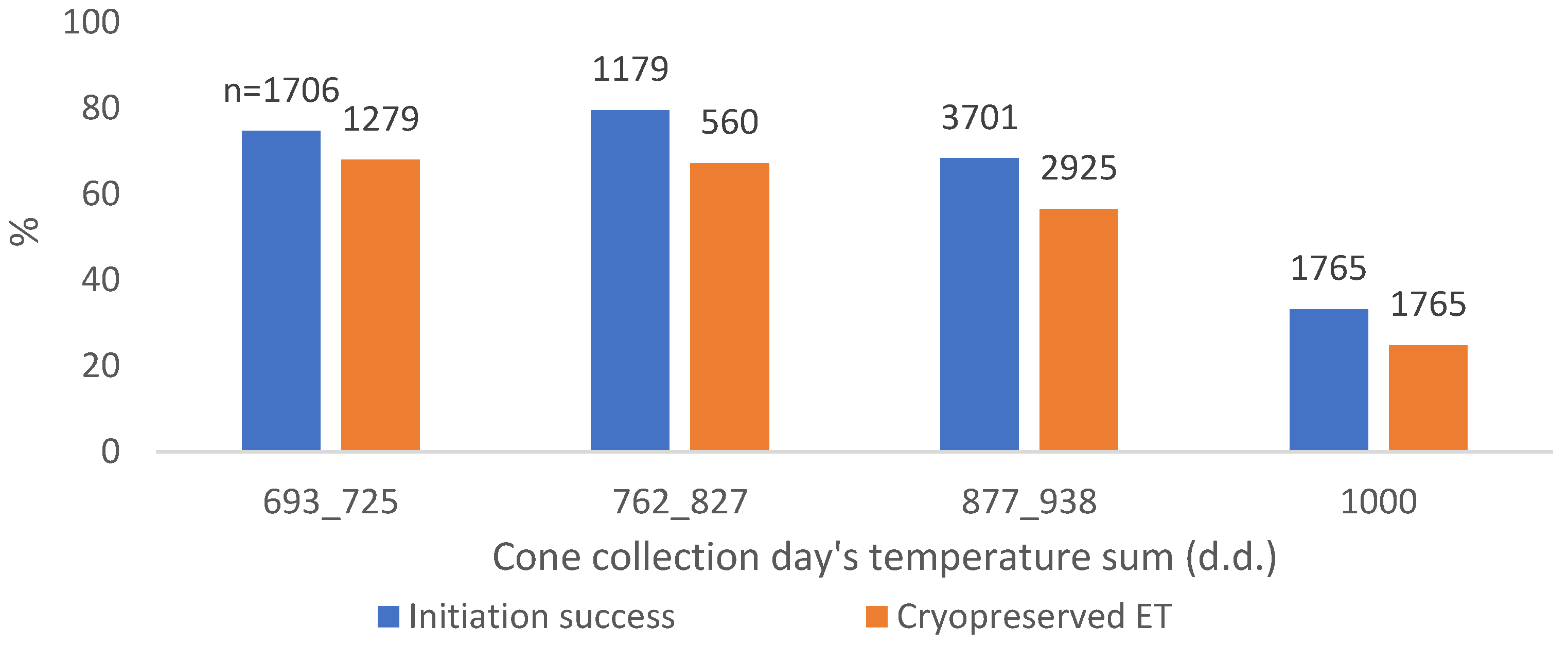
The highest percentage of cases predicted correctly in initiation was found with cone collection d.d. (71.6%, improving the cases predicted correctly by 7.8 units of %) (Figure 2). However, there were missing data in cone collection d.d. (4556 cases of the overall 12,907), making the variable not comprehensive and not fully comparable with other variables. In cryopreserved ET, the situation was similar in the cone collection d.d. (64.7% of cases predicted correctly, improving the cases predicted correctly by 13.6 units of %).
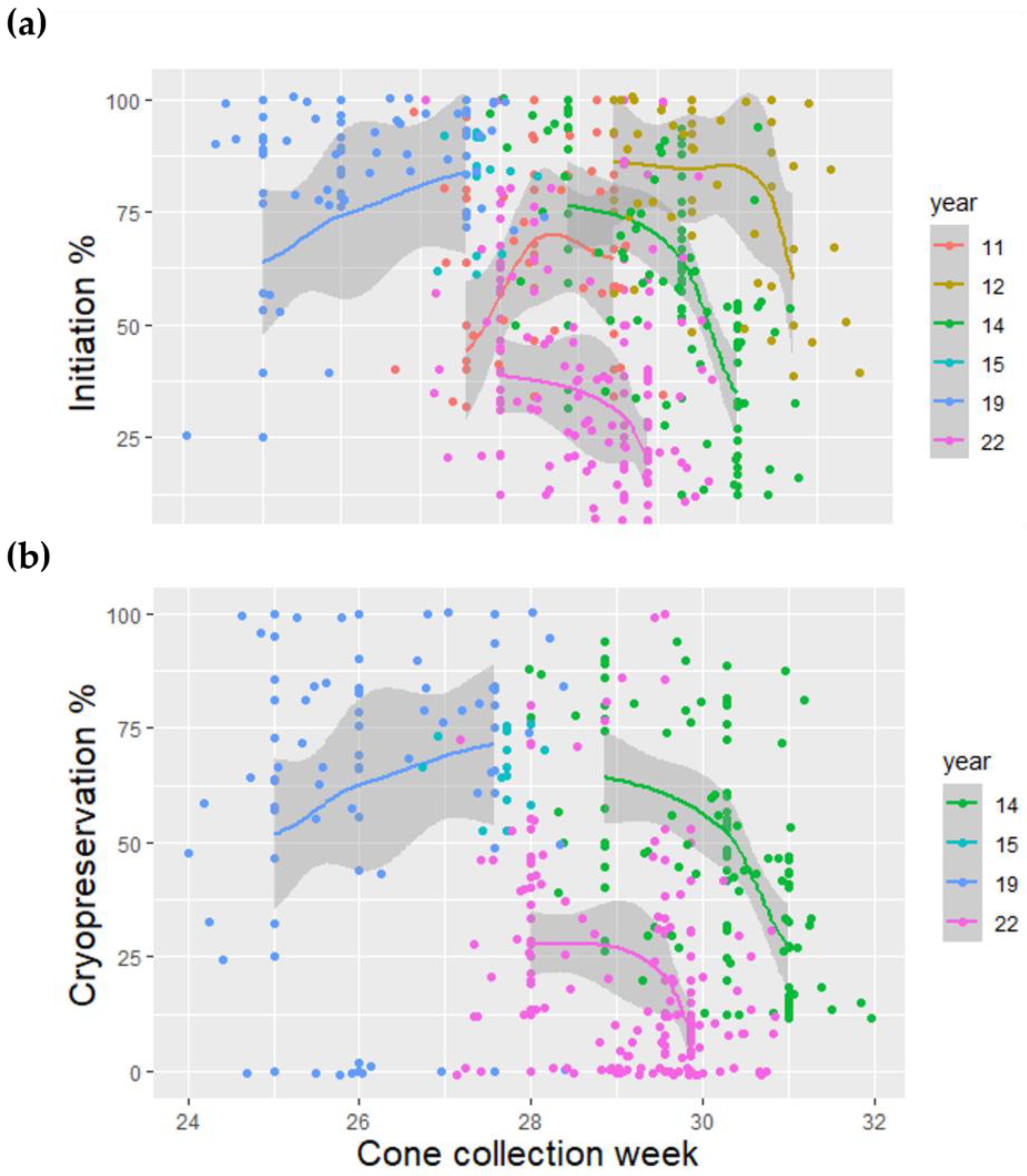
Cone collection date was the best and most comprehensive single variable for predicting the initiation and cryopreserved ET in the logistic regression models (71.0% and 69.1%, respectively) and predicted correctly by 11.2 and 16.0 units of %, respectively (Table 1, Figure 3).
The pollination date was found to have a significant effect on the initiation and cryopreserved ET (p < 0.01 for both in the logistic regression model). Including pollination date in the model resulted in the logistic regression model correctly predicting 63.9 and 64.9% percent of cases in initiation and cryopreserved ET. The improvement in the model was 5.7 and 11.8 units of % in predicting the initiation and cryopreservation success, respectively.
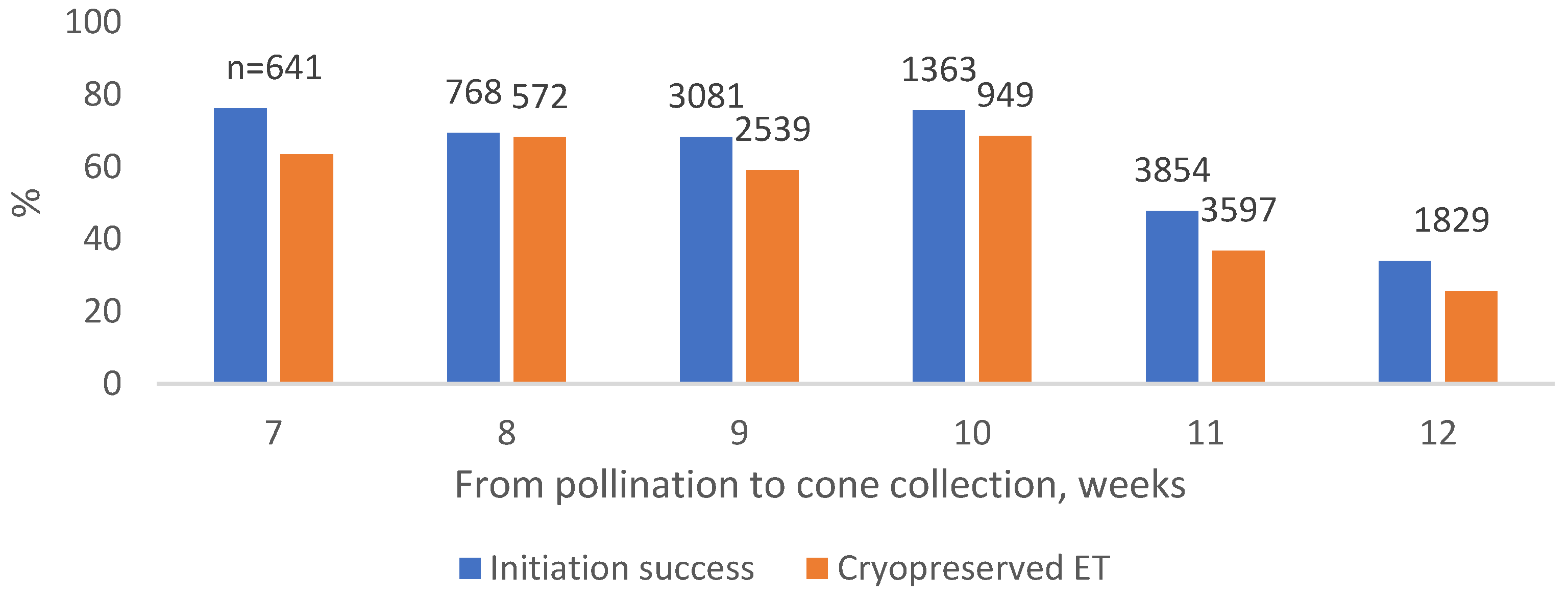
The duration between pollination and cone collection, i.e., cone development duration, had a significant effect on initiation and cryopreserved ET (p < 0.01 for both in the logistic regression model) (Figure 4). It improved the cases predicted correctly by the model by 8.9 and 12.7 units of % to 66.9 and 66.8% in initiation and cryopreserved ET, respectively.
The period from the pollination of flowers to initiation had a significant effect on the initiation and cryopreserved ET (p < 0.01 for both in the logistic regression model). It improved the cases predicted correctly by the model by 10.3 and 12.7 units of % to 68.4 and 66.8% in initiation and cryopreserved ET, respectively.
In 2022, the mean initiation success % was lowest, 31.2%, and in 2015, it was highest, 81.8% (Figure 1a). In 2022, the cryopreserved ET was also low, when only 20.2% of initiations led to a cryopreserved cell line (Figure 1b), which was 64.1% of the initiations that started to grow in the first place. In the best year, 2015, 71.8% of the initiations led to cryopreservation, which was 87.8% of successful initiations. The initiation year had a significant effect on initiation and cryopreserved ET in the logistic regression model (p < 0.01 for both). In the model explaining initiation success, initiation year improved the cases predicted correctly by 6.7 units of % to 66.5, and in the cryopreservation model, 11.4 units to 64.5%.
3.3. Operational Variables in Initiation and Cryopreserved ET
When mother trees were grafts in the seed orchard, 59.2% of the initiations started to grow ET (n = 6605 initiations), which is the same as when grafts were potted in the greenhouse (n = 6302 initiations). From seed orchards, 45.2% of the original explants, i.e., 81.9% of the initiated ET lines were cryopreserved (in 2014, n = 4783 initiations), and from the greenhouse grafts, the corresponding cryopreserved ET % was 65.8 of the original explants, i.e., 87.5% of the initiated ETs (n = 149 initiations). The difference in cryopreserved ET % was significant: χ2 = 10.641, p = 0.001.
Mature (468 initiations) and immature (1891 initiations) seeds were used as a source of ZEs from 17 families in 2011 and 2012. When mature ZEs were used, 47.4% started to grow ET, and when immature ZEs were used, the ET initiation % was 69.1 (χ2 = 76.909, p < 0.01).
The initiation date had a significant effect on initiation and cryopreserved ET in the logistic regression model (p < 0.01 in both). When the initiation date was included in the model, the cases predicted correctly rose to 68.0 (8.2 units of % increment) and 68.5% (15.4 units of % increment).
In 2011, the ET initiation % from 1/2LP media was 16.2%, and from mLM media 61.8%. After the first year, only mLM media was used in the SE initiations of Norway spruce. The initiation media (mLM) lot also had a significant effect on the initiation and cryopreserved ET (p < 0.01 in both). When the initiation media lot was added to the model, the cases predicted correctly rose to 69.4 (6 units of % increment) and 66.0 (14.7 units of % increment).
When cones were cold-stored for one to seven days, and initiations were made immediately afterwards, the ET initiation % was 73.8, and 70.4% of the original initiations led to cryopreservation between 2014 and 2022 (Figure 5a). When cones were stored for more than one week, the ET initiation % was 50.7, and 39.1% of the original initiations led to cryopreservation (2014 to 2022). When both cones and seeds were stored for one to seven days, the ET initiation % was 73.6, and 65.5% of the original initiations led to cryopreservation (2014 to 2022), and if seeds were stored for more than one week, 55.3% of the initiations started to grow ET (Figure 5b).
When the cold storage of cones or seeds was added to the logistic regression model, it was found to have a significant effect on initiation and cryopreserved ET (p < 0.01 for all in the logistic regression model). However, the cold storage of cones (65.0 and 64.3% cases predicted correctly with an increment of 4.8 and 11.9 units of % in initiation and cryopreserved ET, respectively) improved the model much more than the cold storage of seeds (54.7 and 57.4% cases predicted correctly with an increment of 0.4 and 1 units of % in initiation and cryopreserved ET, respectively).
4. Discussion
4.1. Relations of the Different Variables and Limitations of the Study
When more than one variable was included in the model, the highest percentage of cases predicted correctly was found with full-sib family and initiation date in initiation (73.9%, improving the cases predicted correctly by 14.1 units of %) and cryopreserved ET (72.0% success, improving the cases predicted correctly by 18.9 units of %) (Supplementary Table S3).
In the current data, several of the studied variables were partially overlapping among different variable categories. For example, the genetic full-sib family (and other genetic variables) also reflects the variation between the year of initiation and the initiation date (as full-sib families were not replicated every year), and the variables were also derived from the aforementioned ones.
The cone collection date variable, which was used in the final models, reflects the variation caused partly by the different full-sib families and other genetic variables, cone collection d.d., and the time related variables resulting from biological development. The cone collection day also had a significant effect on the initiation day. The cone collection day variable worked well in the model because it was the single variable which most comprehensively predicted the effects caused by other variables. The cone collection day will probably work well as an operational indicator when carrying out the collections. However, it must be noted that unexpected change in the other variables may have a drastic effect on the outcome if the whole process from flowering to initiation is not well known and monitored.
Although the data were very extensive, covering dozens of full-sib families and thousands of initiations, initiated, and cryopreserved cell lines, there were weaknesses that complicated the interpretation of results. First, there were a fair amount of missing data in several variables, which weakened their potential in the statistical modelling. Second, although the data were vast, there were very few replicates with the same genetic background in different experimental variables. Third, in the large dataset, very small differences became significant in the models, which needs to be considered when making conclusions.
4.2. The Effects of Genetic Variables on Initiation Success and the Number of Cryopreserved ET
The overall initiation percentage, 59.8%, in the current study was higher than the 15% in Egertsdotter et al. [13], who used mature ZEs as explants. In Högberg et al.’s [14] study with initiations using immature ZEs, 53.5% of the initiations were growing vigorously after four months. Small steps were taken since Hakman et al. started in 1985 and reported that about 50% of the cultures obtained from ZEs were embryogenic [40].
In the present study, ETs were obtained from 94% of families, which was similar to the previous report by Egertsdotter et al. [13] in which 1 to 3% of Norway spruce families did not generate any SE initiations at all. In our study, the unsuccessful 6% consisted of eight families, of which seven were initiated in 2022. When they were further investigated, it was obvious that there were difficulties in seed and ZE development, because only one to six ZEs were found from two dissected cones per family. It must also be noted that the cones from the aforementioned seven families were cold-stored for 14 days on average (varying from 8 to 18). Judging those families as recalcitrant to SE may, therefore, be misleading. Additionally, five of the mother trees from those eight families were also crossed with different fathers (the number of ZEs was higher in these cases). From these families, successful initiations and cryopreserved ETs were achieved, which can be related to an earlier cone collection date and the greater number of ZEs or shorter cone cold storage (mean 12 days, varying from three to 38 days).
Of the genetic variables related to the parent trees, the full-sib family predicted the outcome best in the logistic regression model. This is understandable, as it covers the variation related to both the mother and father trees. Of the parent trees, the mother tree was slightly more effective in the model, probably because mother tree as a variable also covers some environmental variation due to its growing location and the physiological effects of the mother tree on the development of ZE.
Another tested genetic variable was the effect of PaLAR3B allele on root rot tolerance. The fact that all parents in the present study were genotyped for the B allele enabled it to be confirmed that the allele did not interfere with SE initiation or ET growth that led to cryopreservation, based on this very large dataset extending over different years. In the present study, only the parent genotype was known, whereas the progeny genotype was not analysed. However, we assume that the allele frequency among progeny of different PaLAR3 cross types follows the Mendelian inheritance rules as shown in a previous study [18]. In that study, the progeny from seven different PaLAR3 crosses from 2014 initiations was shown to follow the inheritance rule as expected.
In 2022, the initiation and cryopreserved ET percentages were lower than in previous years. One difference from previous years is that the proportion of heterozygous crosses for the PaLAR3 allele was more numerous than in other years. However, there were no differences in initiation or cryopreserved ET percentages between different PaLAR3 cross types or the amount of B-allele frequency in 2022. The same result was obtained for all previous years in the present study. The reason for the lower initiation percentages is, therefore, probably related to factors other than the PaLAR3 genotype, such as the long storage of cones.
4.3. The Effects of Environmental Variables on Initiation Success and the Number of Cryopreserved ET
The time period variables, duration from pollination to cone collection, and initiation were included to explain the developmental stage of the cone and ZE better than just the date variables (date of collection, cone collection date d.d., and initiation date). Surprisingly, the date variables were more effective in the model, probably because they also reflected variation caused by other factors. It was shown that maritime pine explant’s ability to initiate embryogenic masses was significantly influenced by the collection date of the cones [21,41], and Kvaalen et al. [42] state that the most important factor that determined the initiation success in subalpine fir (Abies lasiocarpa) was the time of cone collection.
The cone collection d.d. and cone collection date are both variables that reflect the developmental stage of the cones and ZEs within the cones. In addition to this, cone collection d.d. reflects the annual variation in temperature and the relationship of the growing season with the calendar, which the cone collection date does not [43]. This probably explains why cone collection d.d. worked better in the part of the data where it was available. On the other hand, the cone collection date affects and partly determines the following operational variables.
Pollination date is highly dependent on flower development, which is affected by the timing of the spring, i.e., temperature sum accumulation. In addition to the d.d. of the year, Norway spruce flower development, especially the number of flowers, is affected by the environmental conditions a year before [44]. Lindgren et al. [44] states that the weather conditions after flowering certainly influence the production of good seeds. If the summer and autumn are cool, the seeds may not mature properly, and this will be reflected by a high proportion of embryos that have not fully developed [44]. In our model, pollination date only covers the running years’ environmental variation before actual pollination, so it does not explain the initiation success and the number of cryopreserved ET very well.
Overall initiation success varied remarkably in the years of our study, although the initiation year as a variable did not work well in the models. This is probably related to genetic material and operational factors varying from year to year, with different goals set for initiations.
4.4. The Effects of Operational Variables on Initiation Success and the Number of Cryopreserved ET
Labour resources are limited in the laboratory, which affects the scheduling of the initiations, as well as the operational workflow later when the ETs are picked and placed in subculture. Moreover, the cryopreservation of hundreds of cell lines with more than one sample is itself a massive process. Previously, e.g., Häggman et al. [45] proposed cold storage of the collected cones to even the laboratory workload, and reported that cold storage could be applied for up to two months without an effect on SE initiation in Scots pine (Pinus sylvestris). However, in the present study, the initiation success decreased when cones were stored in the cold for more than one week, either reflecting a difference between Pinus and Picea or suggesting that the overall low initiation frequency in Scots pine in Häggman’s study masked the cold storage effect. The present result is confirmed in the study of Park [9], in which a significantly higher percentage of the white spruce explants excised from fresh material immediately after cone collection resulted as ETs than of explants excised from cold-stored cones. It seems that sterilised seeds could be stored longer than cones, but the data from long storage are limited in the present study, and the good outcome may be due to a genetic effect.
Due to the limited resources or varying objectives, different approaches to initiation may be applied. For example, in 2019, the aim was to initiate and cryopreserve enough cell lines to serve as future forest regeneration material in Finland, with enough genotypes per family to achieve improved genetic gain after field testing. When the first forest regeneration material was compiled from a collection of families initiated in 2014, we were able to estimate an initiation threshold for 2019 families. The initiation threshold was achieved when the number of initiations that was estimated to produce enough cell lines was achieved from a family [7]. When this threshold was applied, the initiations were carried out in a short period (no excess cold storage), and cryopreservation was carried out in a larger number per cell line than was possible in 2014. Applying this strict threshold probably affected the higher initiation and cryopreserved ET rates in 2019 than in previous years.
Another approach to the conduct of initiations was applied in 2022, when families with a different PaLAR3 background were initiated. The aim was to obtain as many cell lines as possible from the 54 genotyped families. This resulted in a prolonged initiation period, which, in turn, resulted in a lower initiation and cryopreservation rate than in previous years. However, the sheer number of the obtained and cryopreserved cell lines was vast, and the aim was achieved.
Being able to collect material for SE initiations from small potted grafts in the greenhouse near you also saves resources compared to collections from seed orchards or outdoor clonal archives with bigger trees and potentially located at a distance. Based on our results, both options are equally good from the SE initiation perspective, with factors other than the location of donor trees having a greater effect. However, when planning initiations, one should remember that the temperature in the greenhouse is higher than the temperature outdoors, and this is related to the accumulation of the d.d. sum and development of explants.
In 2011, the first year of our Norway spruce SE studies, we tested two different media for initiations, 1/2LP and mLM (Supplementary Table S2), with a drastic superiority of mLM. There were remarkable differences in the quantities of inorganic elements, vitamins, and amino acids, and in addition, ½LP contains more different amino acids and sugars. Interestingly, the quantities of plant growth regulators were similar, suggesting that Norway spruce SE benefits from a richer medium, and that additional amino acids and sugars present in ½LP were not threshold components for SE initiation or proliferation in this species. mLM is used in conifer SE, being preferable for initiation not only in spruce species [28,46] but also, e.g., in Pinus banksiana, P. strobus, P. pinaster, and P. sylvestris [22,47]. However, it should be noted that there are also interactions among basal media and plant growth regulators and trace elements, as described by Park et al. [22].
As previously elaborated with other variables, the effect of the media lot probably describes variation caused by the combination of other factors, i.e., variation among years, cone collection, and initiation dates, as well as genetic variables. Explants prepared previously normally form ETs earlier and are cryopreserved earlier than those managed later in the initiation season, when different media lots were used. In the models, the media lot did not cover any variation which could not be covered with other variables.
4.5. Conclusions and Future Remarks
When trying to capture thousands of genotypes, it is critical to make a strategic plan keeping in mind what the purpose of the material will be. Operational decisions (such as the use of either immature or mature ZE as explants, cold-storing explants awaiting preparation versus collecting them later, etc.) will also affect the outcome, but knowing these effects helps in the optimisation of resources.
According to the current data, the best overall crossings subjected to SE were E799 × E1366 (carried out in 2014), E9990 × 9902, and K1181 × E318 (both carried out in 2019). In the two first mentioned crossings, few ZEs (45 and 5, respectively) were found, and the rates in initiation (93.3 and 100%, respectively) and cryopreserved ET (80 and 100% of ZEs) were high. Thus, the focus was shifted to the K1181 × E318 full-sib family, where 184 ZEs were found, and the initiation (92.4%) and cryopreserved ET (84.8% of ZE’s) rates were excellent. In this crossing, the mother tree was a potted greenhouse graft, and there were two collection dates (24 June in 718 d.d. and 8 July in 883 d.d.), which were both rather early. From the first collection, initiations were made on 1 July (30 ZEs with 96.7% initiation and 93.3% cryopreservation from ZEs) and 5 July (40 ZEs with 87.5% initiation and 80.0% cryopreservation from ZEs) as from the latter collection initiations were made on 11 July (114 ZEs 93.0% initiation and 84.8% cryopreservation from ZEs).
In the optimal scenario, according to the current data, the Norway spruce cones would be collected in southern Finland during the first two weeks of July (in approximately 800 d.d. accumulation) from the seed orchard or greenhouse and delivered quickly to the laboratory, and the cones would be cold-stored for five days or less before initiations on mLM. In the current data, this happened in practice in two years (2015 and 2019) for four full-sib families, with an initiation rate of 94%, and ET from 87% of the original ZEs were cryopreserved. Our results show that with the right timing of explant collection and an efficient SE initiation procedure, the genotype capture from amenable full-sib families of Norway spruce can be very high. Furthermore, almost all families are responsive, and lower initiation frequencies in some families can be compensated by increasing the number of explants—however, considering operational limitations. In practice, biological and societal constraints may have a conflict here because the optimal initiation time with the highest labour intensity coincides with the well-established holiday season in our latitudes.
Supplementary Materials
The following are available online at https://0-www-mdpi-com.brum.beds.ac.uk/article/10.3390/f14040810/s1, Table S1: year of the crossing, mother tree and its location, pollen donor tree, number of initiations, cell lines picked up, and the percentage of picked up cell lines; Table S2: media used in the initiations and proliferation of Norway spruce somatic embryogenesis; Table S3: logistic regression models used for analysing a binary response (living or dead) in the initiation and cryopreservation success. In the models, a1 to a125 are design variables for full-sib families, and b1 to b57 are design variables for the cone collection date.
Author Contributions
Conceptualisation, M.T. and S.V.; methodology, T.A., M.T., S.V. and J.E.; validation, T.A., M.T., S.V. and J.E.; formal analysis, M.T., J.E. and S.V.; investigation, M.T., J.E. and S.V.; resources, T.A.; writing—original draft preparation, S.V., M.T. and J.E.; writing—review and editing, T.A.; visualisation, J.E. and S.V.; supervision, T.A.; project administration, T.A., M.T. and S.V.; funding acquisition, T.A., M.T. and S.V. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by European Regional Development Fund (A31652, A70606, A72814), and Savonlinna municipality, the ForestValue Research Program, which is a transnational research, development, and innovation program jointly funded by national funding organizations within the framework of the ERA-NET Cofund ‘ForestValue—Innovating forest-based bioeconomy’, grant agreement No. 773324, Public Private Partnership—project in Norway spruce vegetative propagation, and The Royal Swedish Academy of Agriculture and Forestry (KSLA) under Tandem Forest Values research program, grant number TFV 2018-0027.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
The authors wish to thank the technical personnel of Natural Resources Institute Finland for their skilful contributions.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; the collection, analyses, or interpretation of data; the writing of the manuscript; or in the decision to publish the results.
References
- Bonga, J. Conifer Clonal Propagation in Tree Improvement Programs. In Vegetative Propagation of Forest Trees; Springer: Berlin/Heidelberg, Germany, 2016; pp. 3–31. [Google Scholar]
- Park, Y.-S. Implementation of Conifer Somatic Embryogenesis in Clonal Forestry: Technical Requirements and Deployment Considerations. Ann. For. Sci. 2002, 59, 651–656. [Google Scholar] [CrossRef] [Green Version]
- Sutton, B. Commercial Delivery of Genetic Improvement to Conifer Plantations Using Somatic Embryogenesis. Ann. For. Sci. 2002, 59, 657–661. [Google Scholar] [CrossRef]
- Haapanen, M.; Mikola, J. Metsänjalostus 2050—Pitkän Aikavälin Metsänjalostusohjelma; Working Papers of the Finnish Forest Research Institute; METLA: Helsinki, Finland, 2008; 50p. Available online: http://jukuri.luke.fi/handle/10024/535985 (accessed on 8 July 2016).
- Haines, R.J.; Woolaston, R.R. The Influence of Reproductive Traits on the Capture of Genetic Gain. Can. J. For. Res. 1991, 21, 272–275. [Google Scholar] [CrossRef]
- Högberg, K.-A. Possibilities and Limitations of Vegetative Propagation in Breeding and Mass Propagation of Norway Spruce; Dept. of Plant Biology and Forest Genetics, Swedish Univ. of Agricultural Sciences: Uppsala, Sweden, 2003; Volume 294, ISBN 91-576-6528-1. [Google Scholar]
- Tikkinen, M. Improved Propagation Efficiency in a Laboratory–Nursery Interface for Somatic Embryogenesis in Norway Spruce. Diss. For. 2018, 265, 35. [Google Scholar] [CrossRef]
- Haapanen, M.; Leinonen, H.; Leinonen, K. Männyn ja kuusen siemenviljelyssiemenen taimitarhakäytön kehitys 2006–2016: Alueellinen tarkastelu. Metsätieteen Aikakauskirja 2017, 2017, 7716. [Google Scholar] [CrossRef] [Green Version]
- Park, Y.S.; Pond, S.E.; Bonga, J.M. Initiation of Somatic Embryogenesis in White Spruce (Picea glauca): Genetic Control, Culture Treatment Effects, and Implications for Tree Breeding. Theor. Appl. Genet. 1993, 86, 427–436. [Google Scholar] [CrossRef]
- Park, Y.S.; Pond, S.E.; Bonga, J.M. Somatic Embryogenesis in White Spruce (Picea glauca): Genetic Control in Somatic Embryos Exposed to Storage, Maturation Treatments, Germination, and Cryopreservation. Theor. Appl. Genet. 1994, 89, 742–750. [Google Scholar] [CrossRef]
- MacKay, J.J.; Becwar, M.R.; Park, Y.-S.; Corderro, J.P.; Pullman, G.S. Genetic Control of Somatic Embryogenesis Initiation in Loblolly Pine and Implications for Breeding. Tree Genet. Genomes 2006, 2, 1–9. [Google Scholar] [CrossRef]
- Cheliak, W.M.; Klimaszewska, K. Genetic Variation in Somatic Embryogenic Response in Open-Pollinated Families of Black Spruce. Theor. Appl. Genet. 1991, 82, 185–190. [Google Scholar] [CrossRef]
- Egertsdotter, U. Plant Physiological and Genetical Aspects of the Somatic Embryogenesis Process in Conifers. Scand. J. For. Res. 2019, 34, 360–369. [Google Scholar] [CrossRef] [Green Version]
- Högberg, K.-A.; Ekberg, I.; Norell, L.; Von Arnold, S. Integration of Somatic Embryogenesis in a Tree Breeding Programme: A Case Study with Picea abies. Can. J. For. Res. 1998, 28, 1536–1545. [Google Scholar] [CrossRef]
- Thor, M.; Moykkynen, T.; Pratt, J.E.; Pukkala, T.; Ronnberg, J.; Shaw, C.G.; Stenlid, J.; Stahl, G.; Woodward, S. Modeling Infection and Spread of Heterobasidion annosum in Coniferous Forests in Europe. U. S. Dep. Agric. For. Serv. Gen. Tech. Rep. PNW 2005, 656, 105. [Google Scholar]
- Nemesio-Gorriz, M.; Hammerbacher, A.; Ihrmark, K.; Källman, T.; Olson, Å.; Lascoux, M.; Stenlid, J.; Gershenzon, J.; Elfstrand, M. Different Alleles of a Gene Encoding Leucoanthocyanidin Reductase (PaLAR3) Influence Resistance against the Fungus Heterobasidion parviporum in Picea abies. Plant Physiol. 2016, 171, 2671–2681. [Google Scholar] [CrossRef] [PubMed]
- Lind, M.; Källman, T.; Chen, J.; Ma, X.-F.; Bousquet, J.; Morgante, M.; Zaina, G.; Karlsson, B.; Elfstrand, M.; Lascoux, M. A Picea abies Linkage Map Based on SNP Markers Identifies QTLs for Four Aspects of Resistance to Heterobasidion parviporum Infection. PLoS ONE 2014, 9, e101049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edesi, J.; Tikkinen, M.; Elfstrand, M.; Olson, Å.; Varis, S.; Egertsdotter, U.; Aronen, T. Root Rot Resistance Locus PaLAR3 Is Delivered by Somatic Embryogenesis (SE) Pipeline in Norway Spruce (Picea abies (L.) Karst.). Forests 2021, 12, 193. [Google Scholar] [CrossRef]
- Hakman, I.; Fowke, L.C.; Von Arnold, S.; Eriksson, T. The Development of Somatic Embryos in Tissue Cultures Initiated from Immature Embryos of Picea abies (Norway spruce). Plant Sci. 1985, 38, 53–59. [Google Scholar] [CrossRef]
- Tautorus, T.E.; Attree, S.M.; Fowke, L.C.; Dunstan, D.I. Somatic Embryogenesis from Immature and Mature Zygotic Embryos, and Embryo Regeneration from Protoplasts in Black Spruce (Picea mariana Mill.). Plant Sci. 1990, 67, 115–124. [Google Scholar] [CrossRef]
- Bercetche, J.; Pâques, M. Somatic Embryogenesis in Maritime Pine (Pinus pinaster). In Somat. Embryogenesis Woody Plants Vol. 3 Gymnosperms; Springer: Berlin/Heidelberg, Germany, 1995; Volume 3, p. 221. [Google Scholar]
- Park, Y.S.; Lelu-Walter, M.A.; Harvengt, L.; Trontin, J.F.; MacEacheron, I.; Klimaszewska, K.; Bonga, J.M. Initiation of Somatic Embryogenesis in Pinus banksiana, P. strobus, P. pinaster, and P. sylvestris at Three Laboratoriesin Canada and France. Plant Cell Tissue Organ Cult. 2006, 86, 87–101. [Google Scholar] [CrossRef]
- Gupta, P.K.; Durzan, D.J. Plantlet Regeneration via Somatic Embryogenesis from Subcultured Callus of Mature Embryos of Picea abies (Norway spruce). Vitro Cell Dev. Biol. 1986, 22, 685–688. [Google Scholar] [CrossRef]
- Phillips, G.C.; Garda, M. Plant Tissue Culture Media and Practices: An Overview. Vitro Cell Dev. Biol.-Plant 2019, 55, 242–257. [Google Scholar] [CrossRef]
- Arnold, S.V.; Eriksson, T. In Vitro Studies of Adventitious Shoot Formation in Pinus contorta. Can. J. Bot. 1981, 59, 870–874. [Google Scholar] [CrossRef]
- Litvay, J.D.; Verma, D.C.; Johnson, M.A. Influence of a Loblolly Pine (Pinus taeda L.). Culture Medium and Its Components on Growth and Somatic Embryogenesis of the Wild Carrot (Daucus carota L.). Plant Cell Rep. 1985, 4, 325–328. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A Revised Medium for Rapid Growth and Bio Assays with Tobacco Tissue Cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Klimaszewska, K.; Cyr, D.R. Conifer Somatic Embryogenesis: I. Development. Dendrobiology 2002, 74, 69–76. [Google Scholar]
- Salaj, T.; Matusova, R.; Salaj, J. Conifer Somatic Embryogenesis-an Efficient Plant Regeneration System for Theoretical Studies and Mass Propagation. Dendrobiology 2015, 35, 209–224. [Google Scholar] [CrossRef] [Green Version]
- Nørgaard, J.V.; Duran, V.; Johnsen, Ø.; Krogstrup, P.; Baldursson, S.; Arnold, S.V. Variations in Cryotolerance of Embryogenic Picea abies Cell Lines and the Association to Genetic, Morphological, and Physiological Factors. Can. J. For. Res. 1993, 23, 2560–2567. [Google Scholar] [CrossRef]
- Haapanen, M. Performance of Genetically Improved Norway Spruce in One-Third Rotation-Aged Progeny Trials in Southern Finland. Scand. J. For. Res. 2020, 35, 221–226. [Google Scholar] [CrossRef]
- Klimaszewska, K.; Lachance, D.; Pelletier, G.; Lelu, M.-A.; Séguin, A. Regeneration of Transgenic Picea glauca, P. mariana, and P. abies after Cocultivation of Embryogenic Tissue with Agrobacterium Tumefaciens. Vitro Cell Dev. Biol.-Plant 2001, 37, 748–755. [Google Scholar] [CrossRef]
- Lelu-Walter, M.-A.; Bernier-Cardou, M.; Klimaszewska, K. Clonal Plant Production from Self- and Cross-Pollinated Seed Families of Pinus sylvestris (L.) through Somatic Embryogenesis. Plant Cell Tissue Organ Cult. 2008, 92, 31–45. [Google Scholar] [CrossRef]
- Varis, S.; Ahola, S.; Jaakola, L.; Aronen, T. Reliable and Practical Methods for Cryopreservation of Embryogenic Cultures and Cold Storage of Somatic Embryos of Norway Spruce. Cryobiology 2017, 76, 8–17. [Google Scholar] [CrossRef]
- Kartha, K.K.; Fowke, L.C.; Leung, N.L.; Caswell, K.L.; Hakman, I. Induction of Somatic Embryos and Plantlets from Cryopreserved Cell Cultures of White Spruce (Picea glauca). J. Plant Physiol. 1988, 132, 529–539. [Google Scholar] [CrossRef]
- Find, J.I.; Floto, F.; Krogstrup, P.; Møller, J.D.; Nørgaard, J.V.; Kristensen, M.M.H. Cryopreservation of an Embryogenic Suspension Culture of Picea sitchensis and Subsequent Plant Regeneration. Scand. J. For. Res. 1993, 8, 156–162. [Google Scholar] [CrossRef]
- Varis, S.A.; Virta, S.; Montalbán, I.A.; Aronen, T. Reducing Pre-and Post-Treatments in Cryopreservation Protocol and Testing Storage at −80 °C for Norway Spruce Embryogenic Cultures. Int. J. Mol. Sci. 2022, 23, 15516. [Google Scholar] [CrossRef] [PubMed]
- Wickham, H. Data Analysis. In Ggplot2: Elegant Graphics for Data Analysis; Wickham, H., Ed.; Springer International Publishing: Cham, Switzerland, 2016; pp. 189–201. ISBN 978-3-319-24277-4. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2008; Available online: https://www.R-project.org/ (accessed on 15 March 2023).
- Hakman, I.; Arnold, S.V. Plantlet Regeneration through Somatic Embryogenesis in Picea abies (Norway spruce). J. Plant Physiol. 1985, 121, 149–158. [Google Scholar] [CrossRef]
- Miguel, C.; Gonçalves, S.; Tereso, S.; Marum, L.; Maroco, J.; Margarida Oliveira, M. Somatic Embryogenesis from 20 Open-Pollinated Families of Portuguese plus Trees of Maritime Pine. Plant Cell Tissue Organ Cult. 2004, 76, 121–130. [Google Scholar] [CrossRef]
- Kvaalen, H.; Daehlen, O.G.; Rognstad, A.T.; Grønstad, B.; Egertsdotter, U. Somatic Embryogenesis for Plant Production of Abies Lasiocarpa. Can. J. For. Res. 2005, 35, 1053–1060. [Google Scholar] [CrossRef]
- Almqvist, C.; Bergsten, U.; Bondesson, L.; Eriksson, U. Predicting Germination Capacity of Pinus sylvestris and Picea abies Seeds Using Temperature Data from Weather Stations. Can. J. For. Res. 1998, 28, 1530–1535. [Google Scholar] [CrossRef]
- Lindgren, K.; Ekberg, I.; Eriksson, G. External Factors Influencing Female Flowering in Picea abies (L.) Karst.; Community Organized Relief Effort: Los Angeles, CA, USA, 1977; ISBN 91-38-03762-9. [Google Scholar]
- Häggman, H.; Jokela, A.; Krajnakova, J.; Kauppi, A.; Niemi, K.; Aronen, T. Somatic Embryogenesis of Scots Pine: Cold Treatment and Characteristics of Explants Affecting Induction. J. Exp. Bot. 1999, 50, 1769–1778. [Google Scholar] [CrossRef]
- Hazubska-Przybyl, T.; Bojarczuk, K. Somatic Embryogenesis of Selected Spruce Species [Picea abies, P. omorika, P. pungens ‘Glauca’ and P. breweriana]. Acta Soc. Bot. Pol. 2008, 77, 189–199. [Google Scholar] [CrossRef]
- Harju, A.; Heiska, S.; Julkunen-Tiitto, R.; Venäläinen, M.; Aronen, T. Somatic Embryogenesis of Pinus sylvestris L. from Parent Genotypes with High-and Low Stilbene Content in Their Heartwood. Forests 2022, 13, 557. [Google Scholar] [CrossRef]
Figure 1.
Sankey diagram of Norway spruce initiation success (a) and cryopreserved ET (b) percentages in different years. n = the number of initiations.
Figure 1.
Sankey diagram of Norway spruce initiation success (a) and cryopreserved ET (b) percentages in different years. n = the number of initiations.

Figure 2.
Norway spruce SE initiation success percentages from 2011 to 2022, and the percentage of initiations which led to cryopreservation from 2014 to 2022 in different cone collection days’ temperature sums (d.d.).
Figure 2.
Norway spruce SE initiation success percentages from 2011 to 2022, and the percentage of initiations which led to cryopreservation from 2014 to 2022 in different cone collection days’ temperature sums (d.d.).

Figure 3.
The effect of cone collection date on initiation (a) and cryopreserved ET (b) of Norway spruce SE cultures. Each dot represents the mean initiation or cryopreserved ET percentage of each family according to the cone collection date.
Figure 3.
The effect of cone collection date on initiation (a) and cryopreserved ET (b) of Norway spruce SE cultures. Each dot represents the mean initiation or cryopreserved ET percentage of each family according to the cone collection date.

Figure 4.
Norway spruce SE initiation success percentages from 2011 to 2022, and the percentage of initiations which led to cryopreservation from 2014 to 2022 in different cone development duration.
Figure 4.
Norway spruce SE initiation success percentages from 2011 to 2022, and the percentage of initiations which led to cryopreservation from 2014 to 2022 in different cone development duration.

Figure 5.
Norway spruce initiation success percentages from 2011 to 2022 and the percentage of initiations which led to cryopreservation from 2014 to 2022 when (a) cones were stored at +2 °C, and (b) sterilised seeds were stored in cold water at +4 °C. n = number of initiations.
Figure 5.
Norway spruce initiation success percentages from 2011 to 2022 and the percentage of initiations which led to cryopreservation from 2014 to 2022 when (a) cones were stored at +2 °C, and (b) sterilised seeds were stored in cold water at +4 °C. n = number of initiations.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
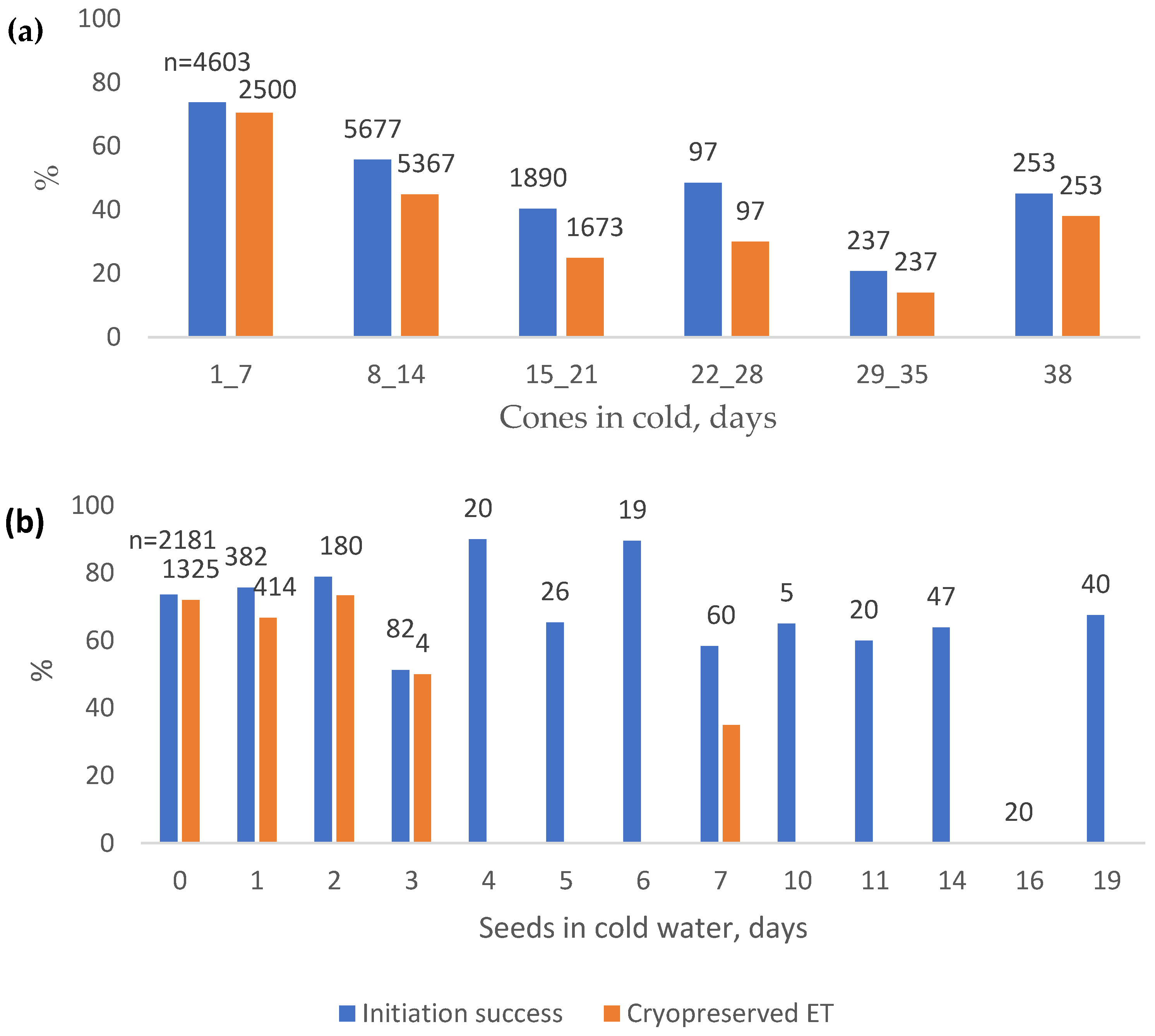{kind=link}
Table 1.
Logistic regression models used to analyse binary response in the initiation (proliferating ET or not) and cryopreservation (cryopreserved or not). In the models, a1 to a15 are design variables for cone collection date.
Table 1.
Logistic regression models used to analyse binary response in the initiation (proliferating ET or not) and cryopreservation (cryopreserved or not). In the models, a1 to a15 are design variables for cone collection date.
| Dependent Variable | Model, log(p/1 − p) | Variable | p-Value | Odds Ratio (95% CI) | Date | % of Cases Predicted Correctly by Model |
|---|---|---|---|---|---|---|
| Initiation | log(p/1 − p) = 1.160 + 0.321a1 − 0.045a2 − 1.038a3 + 0.340a4 − 1.349a5 − 0.349a6 + 159a7 + 0.287a8 − 0.566a9 − 2.183a10 − 2.474a11 − 0.426a12 − 1.816a13 − 0.049a14 | 71.0 | ||||
| Collection date | <0.001 | 1 | 24 June | |||
| 1.378 (1.043–1.821) | 1 July | |||||
| 0.956 (0.760–1.204) | 8 July | |||||
| 0.354 (0.275–0.457) | 12 July | |||||
| 1.406 (1.116–1.77) | 13 July | |||||
| 0.259 (0.207–0.325) | 15 July | |||||
| 0.674 (0.519–0.876) | 18 July | |||||
| 1.172 (0.915–1.501) | 21 July | |||||
| 1.332 (0.916–1.939) | 24 July | |||||
| 0.568 (0.433–0.744) | 25 July | |||||
| 0.113 (0.085–0.15) | 26 July | |||||
| 0.084 (0.067–0.106) | 28 July | |||||
| 0.653 (0.535–0.797) | 31 July | |||||
| 0.163 (0.132–0.200) | 5 August | |||||
| 0.952 (0.594–1.525) | 9 August | |||||
| Cryopreservation | log(p/1 − p) = 0.553 + 0.209a1 + 0.347a2 + 0.380a4 − 1.240a5 + 0.250a7 − 2.125a10 − 2.743a11 − 0.408a12 − 1.621a13 | 69.1 | ||||
| Collection date | <0.001 | 1 | 24 June | |||
| 1.232 (0.971–1.564) | 1 July | |||||
| 1.415 (1.105–1.811) | 8 July | |||||
| 1.463 (1.196–1.789) | 13 July | |||||
| 0.289 (0.234–0.359) | 15 July | |||||
| 1.284 (1.032–1.599) | 21 July | |||||
| 0.119 (0.088–0.162) | 26 July | |||||
| 0.064 (0.050–0.083) | 28 July | |||||
| 0.665 (0.555–0.796) | 31 July | |||||
| 0.198 (0.163–0.240) | 5 August | |||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Varis, S.; Tikkinen, M.; Edesi, J.; Aronen, T. How to Capture Thousands of Genotypes—Initiation of Somatic Embryogenesis in Norway Spruce. Forests 2023, 14, 810. https://0-doi-org.brum.beds.ac.uk/10.3390/f14040810
AMA Style
Varis S, Tikkinen M, Edesi J, Aronen T. How to Capture Thousands of Genotypes—Initiation of Somatic Embryogenesis in Norway Spruce. Forests. 2023; 14(4):810. https://0-doi-org.brum.beds.ac.uk/10.3390/f14040810
Chicago/Turabian StyleVaris, Saila, Mikko Tikkinen, Jaanika Edesi, and Tuija Aronen. 2023. "How to Capture Thousands of Genotypes—Initiation of Somatic Embryogenesis in Norway Spruce" Forests 14, no. 4: 810. https://0-doi-org.brum.beds.ac.uk/10.3390/f14040810
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.