
Adaptation of Forest Management Regimes in Southern Sweden to Increased Risks Associated with Climate Change

Abstract
:1. Introduction
2. Material and Methods
2.1. Growth Simulation Using Heureka-Standwise Model

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tree Species | Site Index, m | Initial Stocking | No. of Thinnings | Rotation Length, Years | MAI, m3·ha−1 |
|---|---|---|---|---|---|
| Norway spruce | G34 | 2000 | 2 | 65 | 12.7 |
| Norway spruce | G34 | 1500 | 0 | 45 | 11.5 |
| Birch | B22 | 1500 | 1 | 50 | 5.0 |
| European beech | F28 | 4000 | 5 | 105 | 7.9 |
| hybrid larch | G34 | 2000 | 3 | 35 | 12.9 |
| hybrid aspen | G34 | 1100 | 2 | 25 | 20.6/23.3 |
2.2. Implementation of Management in Heureka-Standwise Model for Norway Spruce Stand
2.3. Simulation of Risk Factors Associated with Climate Change for Norway Spruce Stand
2.3.1. Simulation of the Risk of Root Rot Infection Under Current Climate and Changing Climate
2.3.2. Simulation of the Risk of Spruce Bark Beetle Damage Under Current Climate and Changing Climate
2.3.3. Simulation of the Risk of Storm Damage Under Current Climate and Changing Climate
2.4. Sensitivity Analysis of Individual Risk Factors Based on Extent of Damage and Profitability of Norway Spruce Stand
| G34-2 | G34-0 | Hybrid Larch | Hybrid Aspen | European Beech | Birch | |
|---|---|---|---|---|---|---|
| Rotation length, years | 57 | 47 | 35 | 100 (25 × 4) | 105 | 50 |
| Regeneration cost, €·ha−1 | 1816 | 1282 | 1816 | 7358 | 3204 | 1282 |
| Total production, m3·ha−1 | 683 | 524 | 453 | 2125 | 716 | 306 |
| MAI, m3·ha−1 | 13.5 | 11.2 | 12.9 | 21.2 | 6.8 | 5.1 |
| Cash flow, €·ha−1·year−1 | 310 | 299 | 278 | 352 | 278 | 132 |
| LEV, €·ha−1 | 4518 | 4528 | 6013 | 6376 | 70 | 1314 |
2.5. Simulation of Combined Effect of Various Risk Factors on Norway Spruce Stand Under Current Climate and Changing Climate
2.6. Management Options in Adaptive Strategy to Reduce the Risk of Damages
2.6.1. Changing Thinning Regimes and Reduction in Rotation Length of Norway Spruce Stand
2.6.2. Simulation of Alternative Tree Species
3. Results
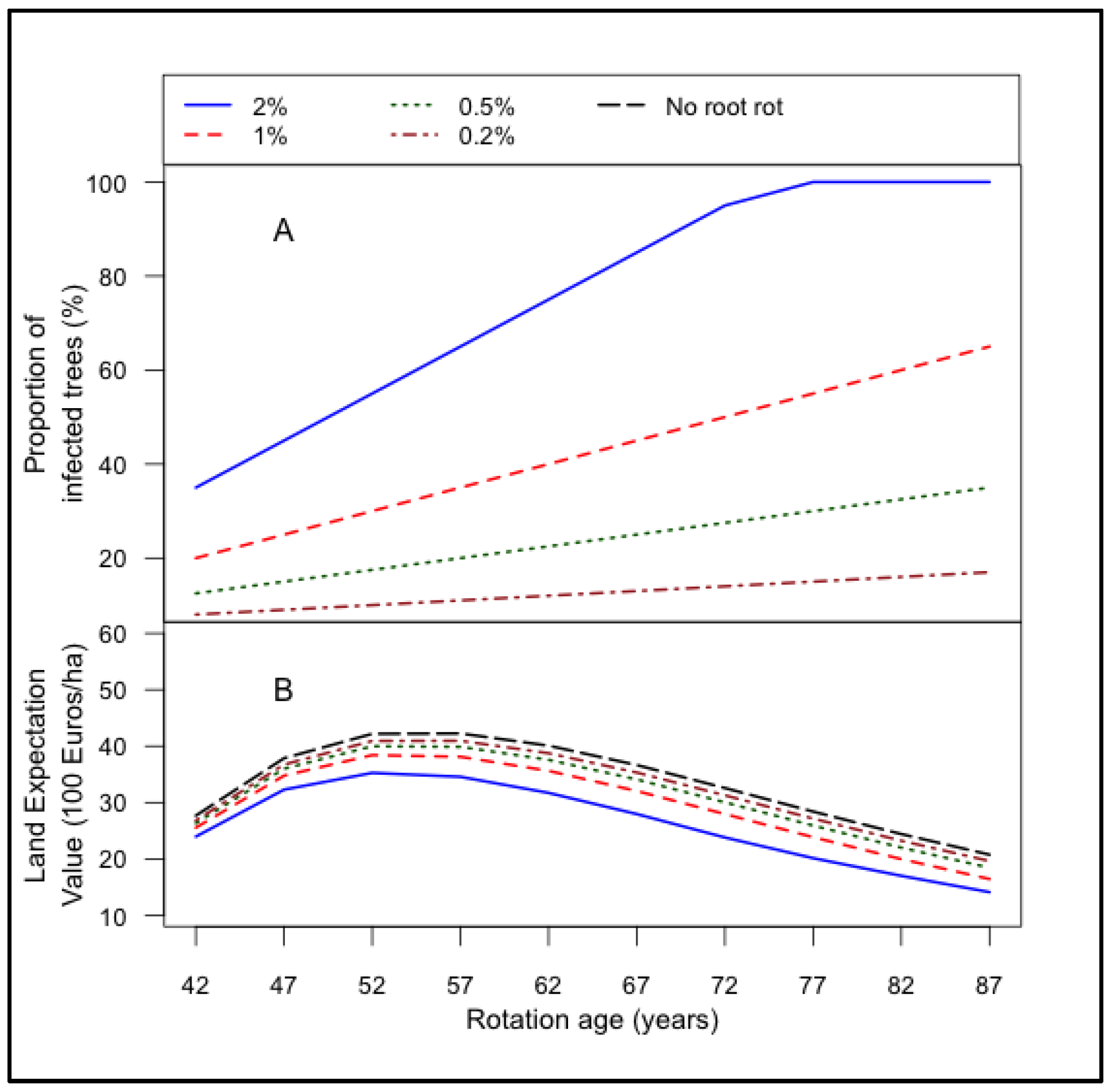
3.1. Root Rot Infection and Its Effect on Land Expectation Values

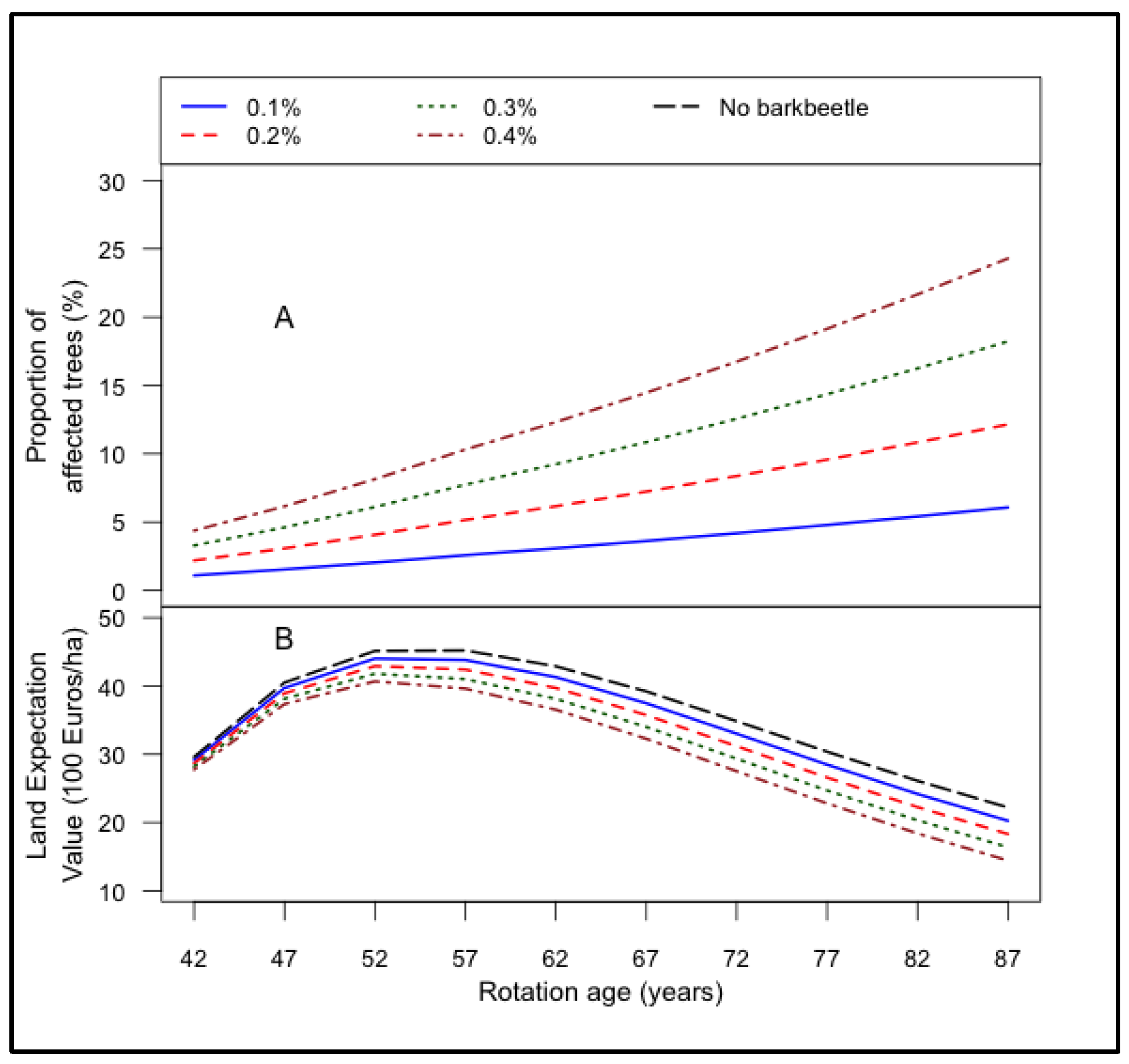
3.2. Bark Beetle Attacks and Its Effect on Land Expectation Values

3.3. Major Storms and Their Effects on Land Expectation Value
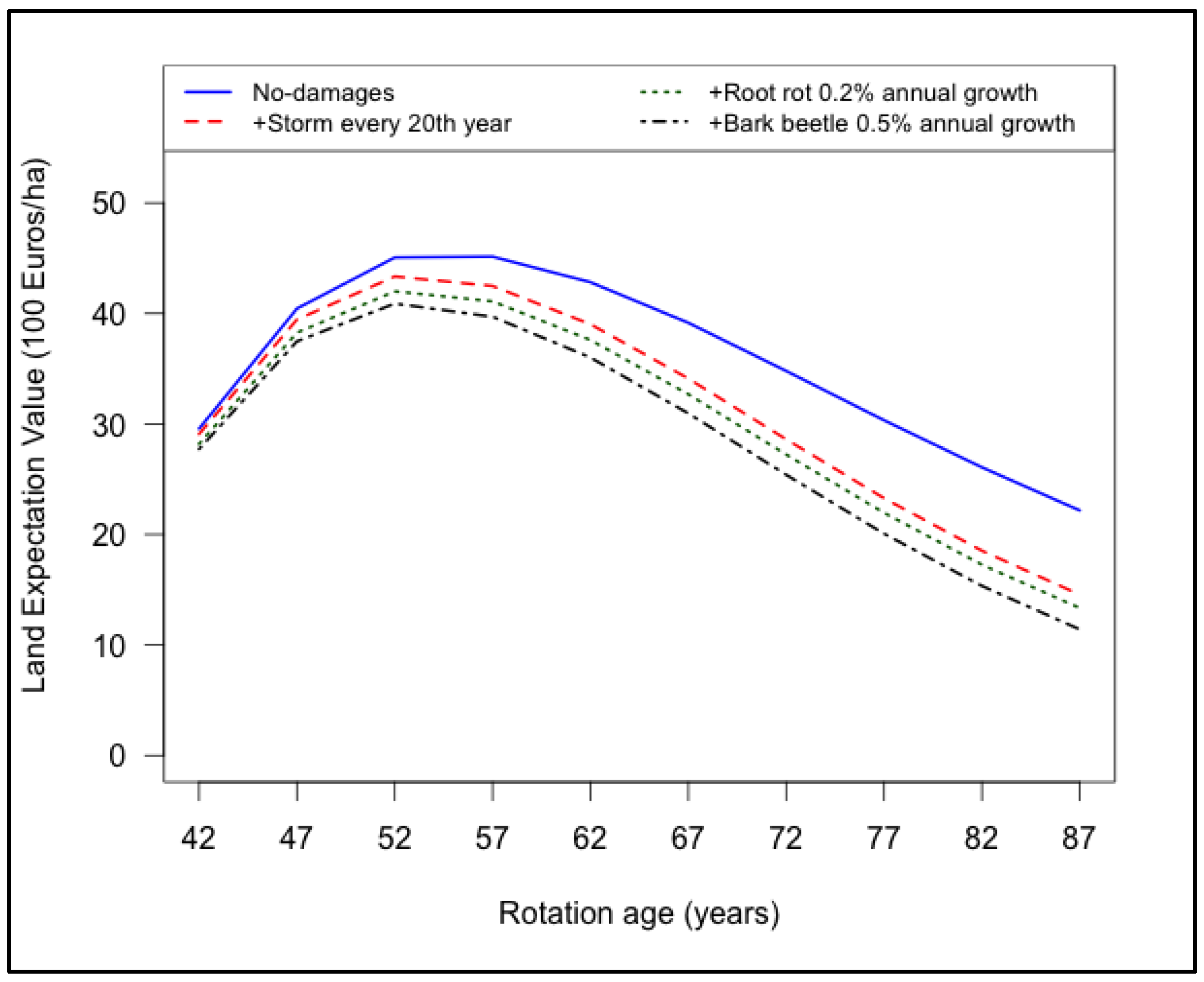
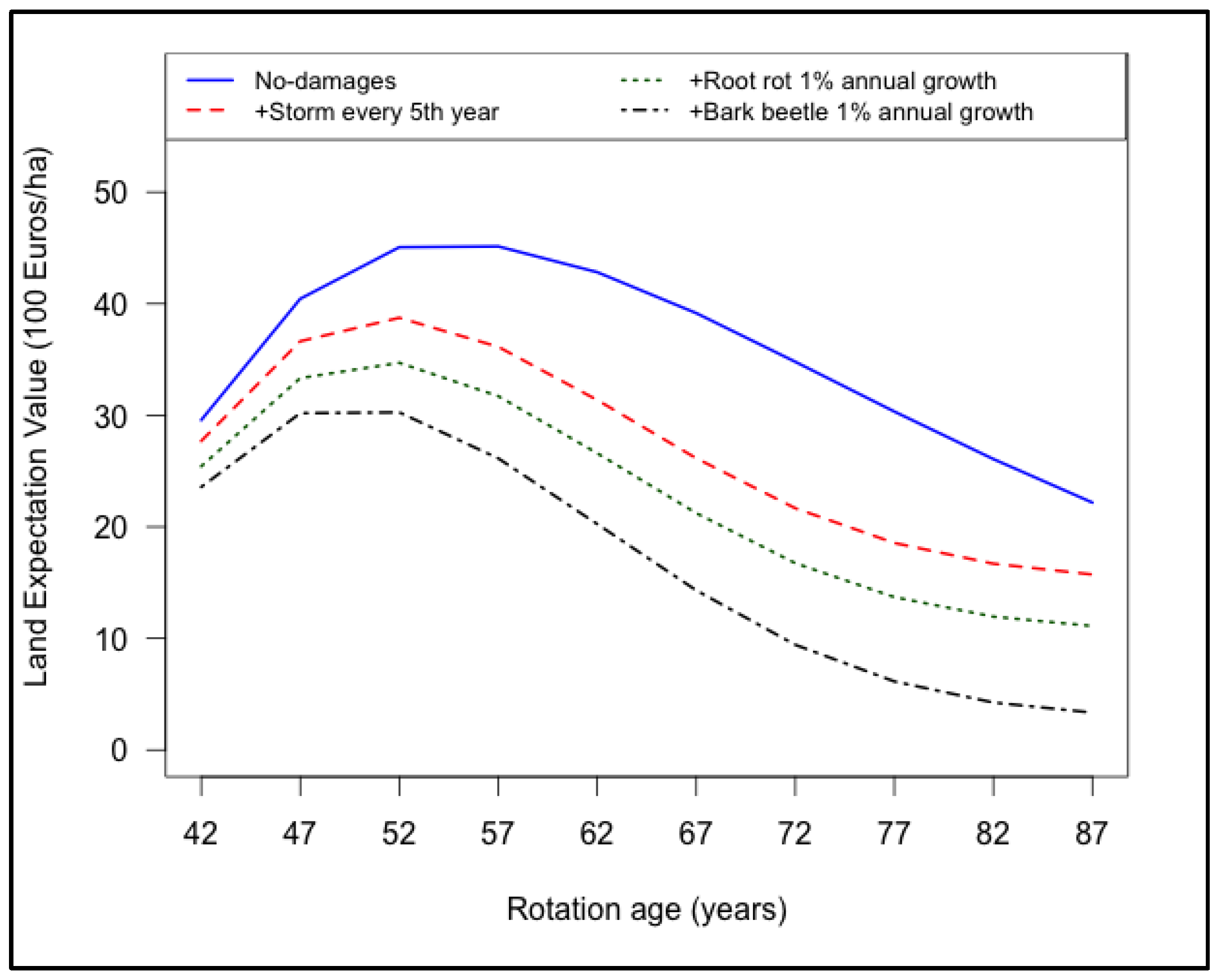
3.4. Combined Effects of Root Rot, Bark Beetle and Major Storms on Land Expectation Values



3.5. Alternative Management Programs and Land Expectation Values
4. Discussion
| Pros | Cons |
|---|---|
| Avoid thinning and reducing rotation length | |
|
|
| Switching to other native species | |
|
|
| Switching to exotic tree species | |
|
|
5. Conclusions
- Damages caused by storms, root rot and bark beetle resulted in reduction of LEV.
- The LEV peaked earlier at high damage levels of root rot (2% growth in annual infection rate), bark beetle (0.3% growth in annual damage rate) and storms.
- Even though the impact of root rot infection and bark beetle damage on LEV of Norway spruce stand is minimal under current infection rate, it is important to implement counter measures now, because in future the infection rate of these risk factors would increase and then more intense counter measures might require which would be costly.
- Avoiding thinning and reducing the rotation length in Norway spruce stands in southern Sweden could reduce the risk of damages due to root rot, storm and bark beetle as the intensity of risk factors increased with age of the stand and soon after thinning operation.
- Switching to other fast growing tree species plantations such as hybrid-larch and hybrid-aspen in southern Sweden were at least as profitable as Norway spruce whereas switching to slow growing species such as birch and European beech gave considerably lower economic compared to Norway spruce.
Acknowledgments
Author Contributions
Conflicts of Interest
References
- SLU. Historical Data from National Forest Inventory. Swedish National Forest Inventory. Swedish University of Agricultural Sciences. Available online: http://www.slu.se/en/collaborative-centres-and-projects/swedish-national-forest-inventory/historical-data/ (accessed on 1 October 2013).
- Claesson, S. Skogliga Konsekvensanalyser 2008—SKA-VB 08 (Impact Assessment in Forestry 2008); Skogsstyrelsen (Swedish Forest Agency) Rapport 25; Swedish Forest Agency: Jönköping, Sweden, 2008. (In Swedish) [Google Scholar]
- Bradshaw, R.H.; Holmqvist, B.H.; Cowling, S.A.; Sykes, M.T. The effects of climate change on the distribution and management of Picea abies in southern Scandinavia. Can. J. For. Res. 2000, 30, 1992–1998. [Google Scholar] [CrossRef]
- Stocker, T.; Qin, D.; Plattner, G.; Tignor, M.; Allen, S.; Boschung, J.; Nauels, A.; Xia, Y.; Bex, B.; Midgley, B. IPCC, 2013: Climate Change 2013: The Physical Science Basis; Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2013. [Google Scholar]
- Intergovernmental Panel on Climate Change (IPCC). Summary for Policymakers. In Climate Change 2013: The Physical Science Basis; Stocker, T.F., Qin, D., Plattner, G.K., Tignor, M., Allen, S.K., Boschung, A., Nauels, Y., Xia, Y., Bex, V., Midgley, P.M., Eds.; Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2013. [Google Scholar]
- Blennow, K.; Olofsson, E. The probability of wind damage in forestry under a changed wind climate. Clim. Chang. 2008, 87, 347–360. [Google Scholar] [CrossRef]
- Jungqvist, G.; Oni, S.K.; Teutschbein, C.; Futter, M.N. Effect of climate change on soil temperature in Swedish boreal forests. PLoS ONE 2014, 9, e93957. [Google Scholar] [CrossRef] [PubMed]
- Smith, B.; Knorr, W.; Widlowski, J.L.; Pinty, B.; Gobron, N. Combining remote sensing data with process modelling to monitor boreal conifer forest carbon balances. For. Ecol. Manag. 2008, 255, 3985–3994. [Google Scholar] [CrossRef]
- Kellomäki, S.; Peltola, H.; Nuutinen, T.; Korhonen, K.T.; Strandman, H. Sensitivity of managed boreal forests in Finland to climate change, with implications for adaptive management. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2008, 363, 2339–2349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bergh, J.; Freeman, M.; Sigurdsson, B.; Kellomaki, S.; Laitinen, K.; Niinisto, S.; Peltola, H.; Linder, S. Modelling the short-term effects of climate change on the productivity of selected tree species in Nordic countries. For. Ecol. Manag. 2003, 183, 327–340. [Google Scholar] [CrossRef]
- Koca, D.; Smith, B.; Sykes, M.T. Modelling regional climate change effects on potential natural ecosystems in Sweden. Clim. Chang. 2006, 78, 381–406. [Google Scholar] [CrossRef]
- Pussinen, A.; Nabuurs, G.; Wieggers, H.; Reinds, G.; Wamelink, G.; Kros, J.; Mol-Dijkstra, J.; de Vries, W. Modelling long-term impacts of environmental change on mid-and high-latitude European forests and options for adaptive forest management. For. Ecol. Manag. 2009, 258, 1806–1813. [Google Scholar] [CrossRef]
- Bergh, J.; Linder, S. Skogsbruket måste påbörja omställningen (Conversion should Begin in Forest Management). In Formas Fokuserar “Sverige i ett nytt Klimat-Våtvarm Utmaning” (Formas Focuses—“Sweden in a New Climate—Wet-Warm Challange”); Swedish National Forest Inventory (SLU): Alnarp, Sweden, 2010. [Google Scholar]
- Subramanian, N. Simulation of Net Primary Production (NPP) of Picea Abies in Southern Sweden: An Analysis Based on Three Forest Growth Models; Swedish University of Agricultural Science: Alnarp, Sweden, 2010; p. 43. [Google Scholar]
- Bolte, A.; Ammer, C.; Löf, M.; Madsen, P.; Nabuurs, G.-J.; Schall, P.; Spathelf, P.; Rock, J. Adaptive forest management in central Europe: Climate change impacts, strategies and integrative concept. Scand. J. For. Res. 2009, 24, 473–482. [Google Scholar] [CrossRef]
- Mason, W.; Petr, M.; Bathgate, S. Silvicultural strategies for adapting planted forests to climate change: From theory to practice. J. For. Sci. 2012, 6, 265–277. [Google Scholar]
- Meason, D.F.; Mason, W.L. Evaluating the deployment of alternative species in planted conifer forests as a means of adaptation to climate change-case studies in New Zealand and Scotland. Ann. For. Sci. 2014, 71, 239–253. [Google Scholar] [CrossRef]
- Blennow, K.; Eriksson, H.M.; Skogsstyrelsen, S. Riskhantering i Skogsbruket. (Risk Handling in Forestry); Rapport 14; Skogsstyrelsen: Jönköping, Sweden, 2006; p. 51. [Google Scholar]
- Blennow, K.; Andersson, M.; Bergh, J.; Sallnas, O.; Olofsson, E. Potential climate change impacts on the probability of wind damage in a south Swedish forest. Clim. Chang. 2010, 99, 261–278. [Google Scholar] [CrossRef]
- Clark, A.J. Climate. The changing climatic environment for New Zealand’s land-based sectors. In Impacts of Climate Change on Land-Based Sectors and Adaptation Options; Clark, A., Nottage, R., Wilcocks, L., Lee, J., Burke, C., Kalaugher, E., Roche, J., Beukes, P., Lieffering, M., Newton, P., Eds.; Stakeholder Report to the Sustainable Land Management and Climate Change; Ministry for Primary Industries: Wellington, New Zealand, 2012. [Google Scholar]
- Dukes, J.S.; Pontius, J.; Orwig, D.; Garnas, J.R.; Rodgers, V.L.; Brazee, N.; Cooke, B.; Theoharides, K.A.; Stange, E.E.; Harrington, R. Responses of insect pests, pathogens, and invasive plant species to climate change in the forests of northeastern North America: What can we predict? Can. J. For. Res. 2009, 39, 231–248. [Google Scholar] [CrossRef]
- Björkman, C.; Bylund, H.; Klapwijk, M.J.; Kollberg, I.; Schroeder, M. Insect pests in future forests: More severe problems? Forests 2011, 2, 474–485. [Google Scholar] [CrossRef]
- Stenlid, J.; Oliva, J.; Boberg, J.B.; Hopkins, A.J. Emerging diseases in European forest ecosystems and responses in society. Forests 2011, 2, 486–504. [Google Scholar] [CrossRef]
- Klapwijk, M.J.; Ayres, M.P.; Battisti, A.; Larsson, S. Assessing the impact of climate change on outbreak potential. In Insect Outbreaks Revisited; Wiley-Blackwell: Hoboken, NJ, USA, 2012; pp. 429–450. [Google Scholar]
- Keenan, R.J. Adaptation of forests and forest management to climate change: An editorial. Forests 2012, 3, 75–82. [Google Scholar] [CrossRef]
- Wallentin, C.; Nilsson, U. Storm and snow damage in a Norway spruce thinning experiment in southern Sweden. Forestry 2014, 87, 229–238. [Google Scholar] [CrossRef]
- Möykkynen, T.; Miina, J.; Pukkala, T. Optimizing the management of a Picea abies stand under risk of butt rot. For. Pathol. 2000, 30, 65–76. [Google Scholar] [CrossRef]
- Martikainen, P.; Siitonen, J.; Kaila, L.; Punttila, P.; Rauh, J. Bark beetles (Coleoptera, Scolytidae) and associated beetle species in mature managed and old-growth boreal forests in southern Finland. For. Ecol. Manag. 1999, 116, 233–245. [Google Scholar] [CrossRef]
- Lindner, M.; Maroschek, M.; Netherer, S.; Kremer, A.; Barbati, A.; Garcia-Gonzalo, J.; Seidl, R.; Delzon, S.; Corona, P.; Kolström, M.; et al. Climate change impacts, adaptive capacity, and vulnerability of European forest ecosystems. For. Ecol. Manag. 2010, 259, 698–709. [Google Scholar] [CrossRef]
- Bodin, P.; Wiman, B.L. The usefulness of stability concepts in forest management when coping with increasing climate uncertainties. For. Ecol. Manag. 2007, 242, 541–552. [Google Scholar] [CrossRef]
- Spittlehouse, D.L. Climate Change, Impacts, and Adaptation Scenarios: Climate Change and Forest and Range Management in British Columbia; Ministry of Forests and Range: Victoria, BC, Canada, 2008. [Google Scholar]
- Elfving, B.; Kiviste, A. Construction of site index equations for Pinus sylvestris L. using permanent plot data in Sweden. For. Ecol. Manag. 1997, 98, 125–134. [Google Scholar] [CrossRef]
- Carbonnier, C. Yield of Beech in Southern Sweden; Institution för Skogsproduktion: Stockholm, Sweden, 1971; pp. 1–89. (In Swedish) [Google Scholar]
- Ekö, P.M.; Stern, M.L.; Albrektson, A. Growth and yield of hybrid larch (Larix× eurolepis A. Henry) in Southern Sweden. Scand. J. For. Res. 2004, 19, 320–328. [Google Scholar] [CrossRef]
- Oikarinen, M. Etelä-Suomen Viljeltyjen Rauduskoivikoiden Kasavatusmallit (Growth and Yield Models for Silver Birch (Betula Pendula) Plantations in Southern Finland); The Finnish Forest Research Institute: Helsinki, Finland, 1983; Volume 113, p. 75. [Google Scholar]
- Fahlvik, N.; Wikström, P.; Elfving, B. Evaluation of growth models used in the Swedish Forest Planning System Heureka. Silva Fenn. 2014, 48, 1–17. [Google Scholar] [CrossRef]
- Wikström, P.; Edenius, L.; Elfving, B.; Eriksson, L.O.; Lämås, T.; Sonesson, J.; Öhman, K.; Wallerman, J.; Waller, C.; Klintebäck, F. The Heureka Forestry Decision Support System: An Overview. Math. Comput. For. Nat. Resour. Sci. 2011, 3, 87–95. [Google Scholar]
- Pettersson, N. The Effect on Stand Development of Different Spacing after Planting and Precommercial Thinning in Norway Spruce (Picea abies (L.) Karst.) and Scots Pine (Pinus sylvestris L.) Stands; Rapport-Sveriges Lantbruksuniversitet; Swedish University of Agricultural Sciences: Uppsala, Sweden, 1992. [Google Scholar]
- Elfving, B. Övre Höjdens Utveckling i Granplanteringar (Top Height Growth in Norway Spruce Plantations); Department of Silviculture, Swedish University of Agricultural Sciences: Uppsala, Sweden, 2003; p. 8. (In Swedish) [Google Scholar]
- Falvik, N.; Nilsson, U. Granskogsbruksmodell (DT) (Model for Norway Spruce Stand (DT)). Available online: http://www-gran.slu.se/Webbok/PDFdokument/GranmodellenMoM.pdf (accessed on 10 September 2015).
- Söderberg, U. Funktioner för Skogsindelning: Höjd, Formhöjd Och Barktjocklek för Enskilda Träd (Functions for Forest Management: Height, form Height and Bark Thickness of Individual Trees); Institutionen for Skogstaxering: Umea, Sweden, 1992. [Google Scholar]
- Elfving, B.; Nyström, K. Growth Modelling in the Heureka System; Department of Forest Ecology and Management, Swedish University of Agricultural Sciences: Umeå, Sweden, 2010; p. 97. [Google Scholar]
- Brandel, G. Volume Functions for Individual Trees; Scots Pine (Pinus Sylvestris), Norway Spruce (Picea Abies) and Birch (Betula Pendula & Betula Pubescens); Swedish University of Agricultural Sciences: Uppsala, Sweden, 1990. [Google Scholar]
- Fridman, J.; Ståhl, G. A three-step approach for modelling tree mortality in Swedish forests. Scand. J. For. Res. 2001, 16, 455–466. [Google Scholar] [CrossRef]
- Skogsstyrelsen. Gallringsmallar (Thinning Guides); Skogsstyrelsen: Jönköping, Sweden, 1984; p. 35. [Google Scholar]
- Thor, M.; Arlinger, J.D.; Stenlid, J. Heterobasidion annosum root rot in Picea abies: Modelling economic outcomes of stump treatment in Scandinavian coniferous forests. Scand. J. For. Res. 2006, 21, 414–423. [Google Scholar] [CrossRef]
- Windmuller-Campione, M.A.; Long, J.N. If Long-Term Resistance to a Spruce Beetle Epidemic is Futile, Can Silvicultural Treatments Increase Resilience in Spruce-Fir Forests in the Central Rocky Mountains? Forests 2015, 6, 1157–1178. [Google Scholar] [CrossRef]
- Persson, P. Stormskador på Skog: Uppkomstbetingelser och Inverkan av Skogliga Aatgaerder (Windthrow in Forests: It’s Causes and the Effect of Forestry Measures); Rapporter Och Uppsatser/Institutionen Foer Skogsproduktion; Royal College of Forestry: Stockholm, Sweden, 1975; p. 294. [Google Scholar]
- Schlyter, P.; Stjernquist, I.; Bärring, L.; Jönsson, A.M.; Nilsson, C. Assessment of the impacts of climate change and weather extremes on boreal forests in northern Europe, focusing on Norway spruce. Clim. Res. 2006, 31, 75–84. [Google Scholar] [CrossRef]
- Nilsson, C.; Stjernquist, I.; Bärring, L.; Schlyter, P.; Jönsson, A.M.; Samuelsson, H. Recorded storm damage in Swedish forests 1901–2000. For. Ecol. Manag. 2004, 199, 165–173. [Google Scholar] [CrossRef]
- Faustmann, M. Berechnung des Wertes welchen Waldboden sowie noch nicht haubare Holzbestände für die Waldwirtschaft besitzen (Calculation of the Value Which Forest Land and Immature Stands Possess for Forestry). Allg. Forst Jagdztg. 1849, 15, 7–44. [Google Scholar]
- Faustmann, M. Calculation of the value which forest land and immature stands possess for forestry. J. For. Econ. (Sweden) 1995, 1, 7–44. [Google Scholar]
- Castanheira, P.; Oliveira, J. Land Expectation Value Calculation in Pinus pinaster forestry. In Proceedings of the 21st Annual European Real Estate Society Conference, Bucharest, Romania, 25–28 June 2014.
- Swedish Forest Agency—Prices-Tables and Figures. Available online: http://www.skogsstyrelsen.se/en/AUTHORITY/Statistics/Subject-Areas/Prices/Tables-and-figures/ (accessed on 7 May 2015).
- Nilsson, U.; Southern Swedish Forest Research Centre, Rörsjövägen 1, Alnarp 230 53, Sweden. Personal Communication, 2015.
- Rytter, L. Räkna Med hybridasp (Count with hybrid Aspen). Available online: http://www.kunskapdirekt.se/sv/KunskapDirekt/Lov/Verktyg/hybridasp/ (accessed on 7 May 2015).
- Carbonnier, C.; Hägglund, B. A Comparison between the Volume and Value of the Yield from Beech and Norway Spruce; Royal College of Forestry: Stockholm, Sweden, 1969. [Google Scholar]
- Rytter, L.; Stener, L.-G. Growth and thinning effects during a rotation period of hybrid aspen in southern Sweden. Scand. J. For. Res. 2014, 29, 747–756. [Google Scholar] [CrossRef]
- Stenlid, J. Controlling and predicting the spread of Heterobasidion annosum from infected stumps and trees of Picea abies. Scand. J. For. Res. 1987, 2, 187–198. [Google Scholar] [CrossRef]
- Vollbrecht, G.; Agestam, E. Modelling incidence of root rot in Picea abies plantations in southern Sweden. Scand. J. For. Res. 1995, 10, 74–81. [Google Scholar] [CrossRef]
- Yde-Andersen, A. Stump protection with urea against Fomes annosus in Norway spruce. Det Forstl. Forsøgsvæs. Dan. 1982, 38, 209–217. [Google Scholar]
- Wermelinger, B. Ecology and management of the spruce bark beetle Ips typographus—A review of recent research. For. Ecol. Manag. 2004, 202, 67–82. [Google Scholar] [CrossRef]
- Thor, M.; Ståhl, G.; Stenlid, J. Modelling root rot incidence in Sweden using tree, site and stand variables. Scand. J. For. Res. 2005, 20, 165–176. [Google Scholar] [CrossRef]
- Långström, B.; Lindelöw, Å.; Schroeder, M.; Björklund, N.; Öhrn, P. The spruce bark beetle outbreak in Sweden following the January-storms in 2005 and 2007. In Proceedings of the Workshop on “Insect and Fungi in Strom Areas”, Štrbské Pleso, Slovakia, 15–19 September 2008; IUFRO: Štrbské Pleso, Slovakia.
- Lagergren, F.; Jönsson, A.M.; Blennow, K.; Smith, B. Implementing storm damage in a dynamic vegetation model for regional applications in Sweden. Ecol. Model. 2012, 247, 71–82. [Google Scholar] [CrossRef]
- Štefančík, I. Development of spruce (Picea abies (L.) Karst.) target (crop) trees in pole-stage stand with different initial spacing and tending regime. J. For. Sci. 2012, 58, 456–464. [Google Scholar]
- Rössler, G. The European stem number experiment in Norway spruce (Picea abies (L.) Karst.), 3rd report: Austria (A 18). Ber. Freib. Forstliche Forsch. 2006, 66, 74–86. [Google Scholar]
- Pfister, O.; Wallentin, C.; Nilsson, U.; Ekö, P.-M. Effects of wide spacing and thinning strategies on wood quality in Norway spruce (Picea abies) stands in southern Sweden. Scand. J. For. Res. 2007, 22, 333–343. [Google Scholar] [CrossRef]
- Rönnberg, J.; Vollbrecht, G. Early infection by Heterobasidion annosum in Larix × eurolepis seedlings planted on infested sites. Eur. J. For. Pathol. 1999, 29, 81–86. [Google Scholar]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Subramanian, N.; Bergh, J.; Johansson, U.; Nilsson, U.; Sallnäs, O. Adaptation of Forest Management Regimes in Southern Sweden to Increased Risks Associated with Climate Change. Forests 2016, 7, 8. https://0-doi-org.brum.beds.ac.uk/10.3390/f7010008
Subramanian N, Bergh J, Johansson U, Nilsson U, Sallnäs O. Adaptation of Forest Management Regimes in Southern Sweden to Increased Risks Associated with Climate Change. Forests. 2016; 7(1):8. https://0-doi-org.brum.beds.ac.uk/10.3390/f7010008
Chicago/Turabian StyleSubramanian, Narayanan, Johan Bergh, Ulf Johansson, Urban Nilsson, and Ola Sallnäs. 2016. "Adaptation of Forest Management Regimes in Southern Sweden to Increased Risks Associated with Climate Change" Forests 7, no. 1: 8. https://0-doi-org.brum.beds.ac.uk/10.3390/f7010008