Effects of Stand Structure, Browsing, and Biophysical Conditions on Regeneration Following Mountain Pine Beetle in Mixed Lodgepole Pine and Aspen Forests of the Southern Rockies

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Site Selections
2.3. Field Data Collection
2.4. Animal Use Information
2.5. Topographic and Landform Data
2.6. Statistical Analysis
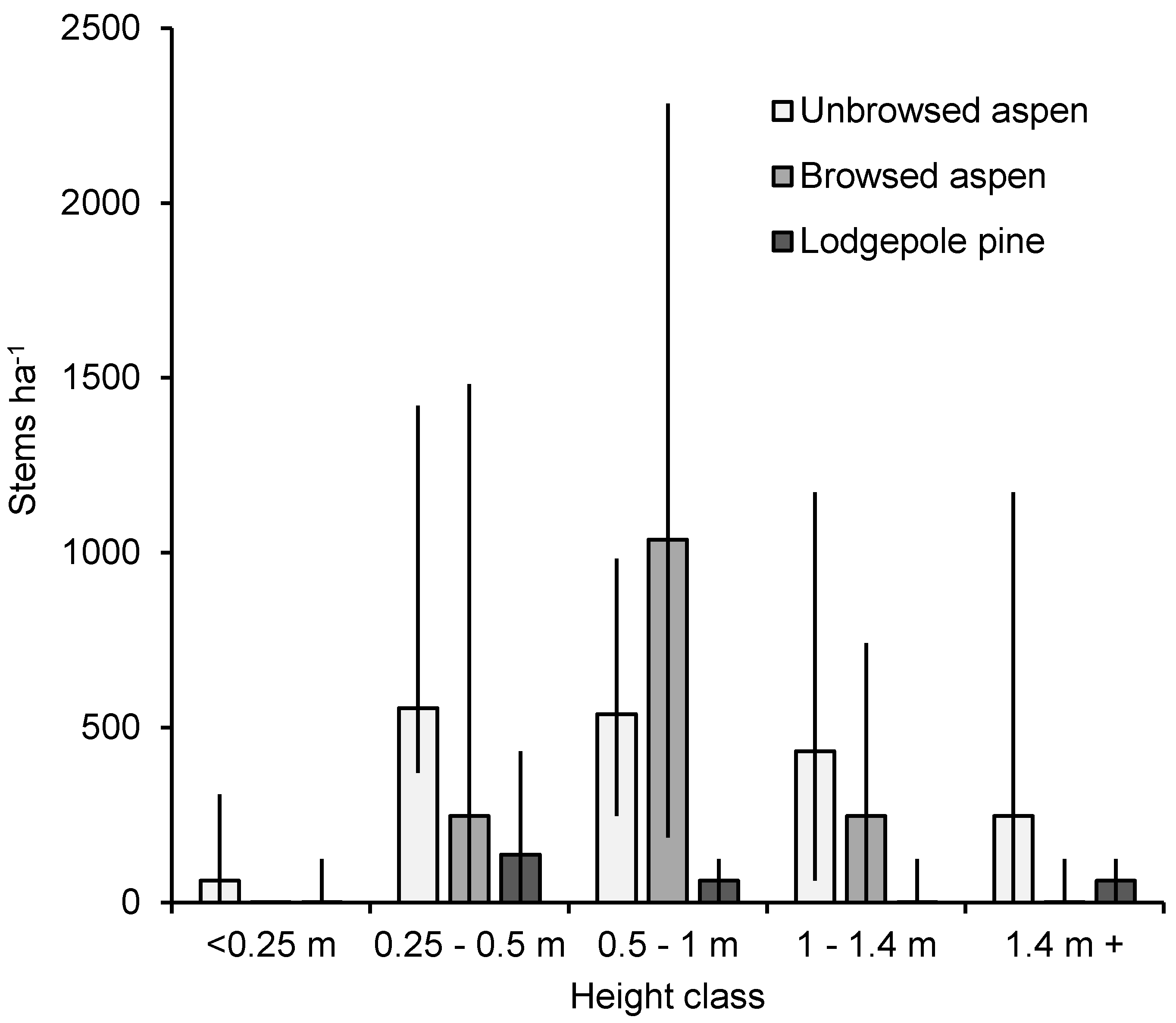
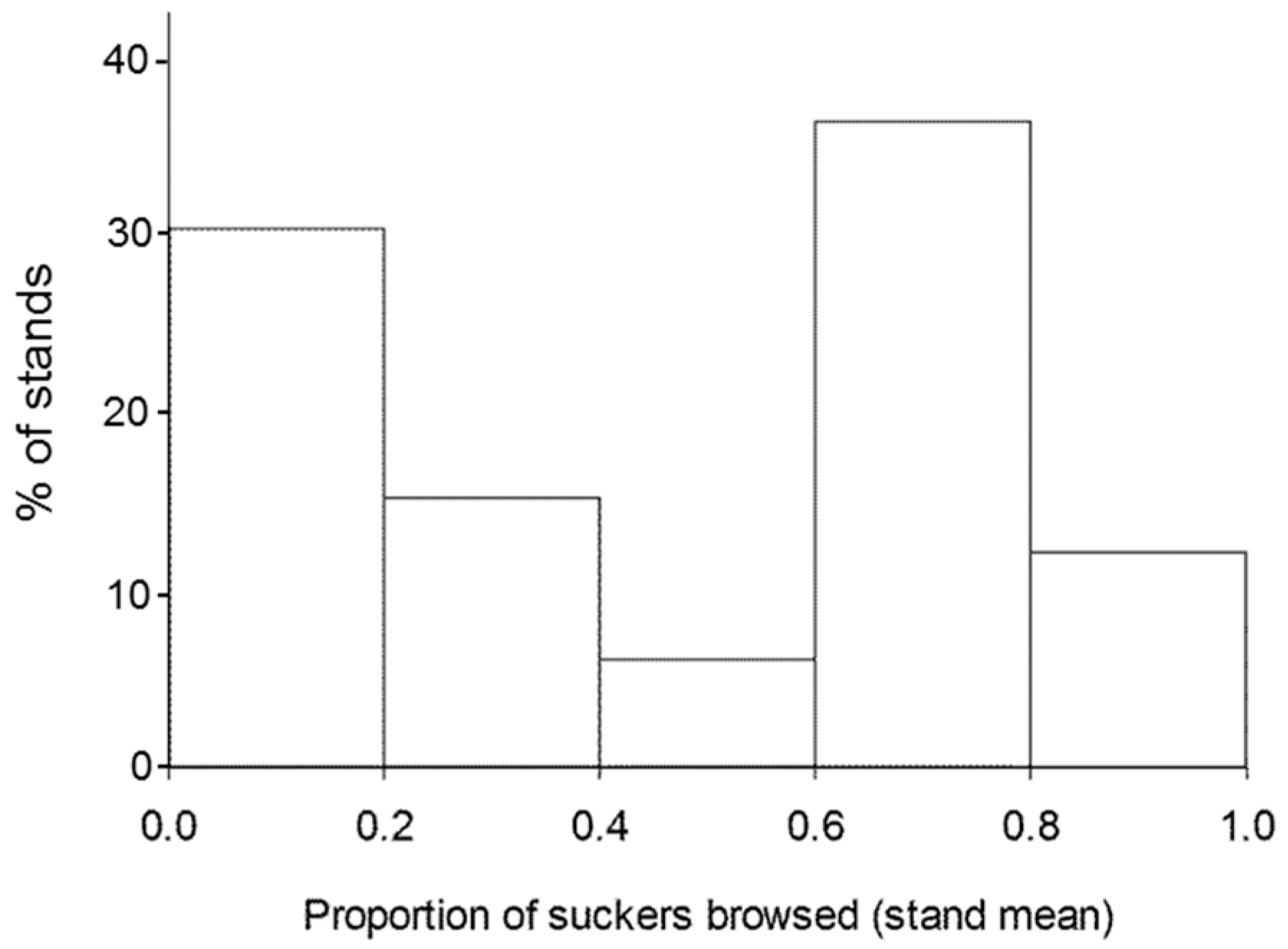
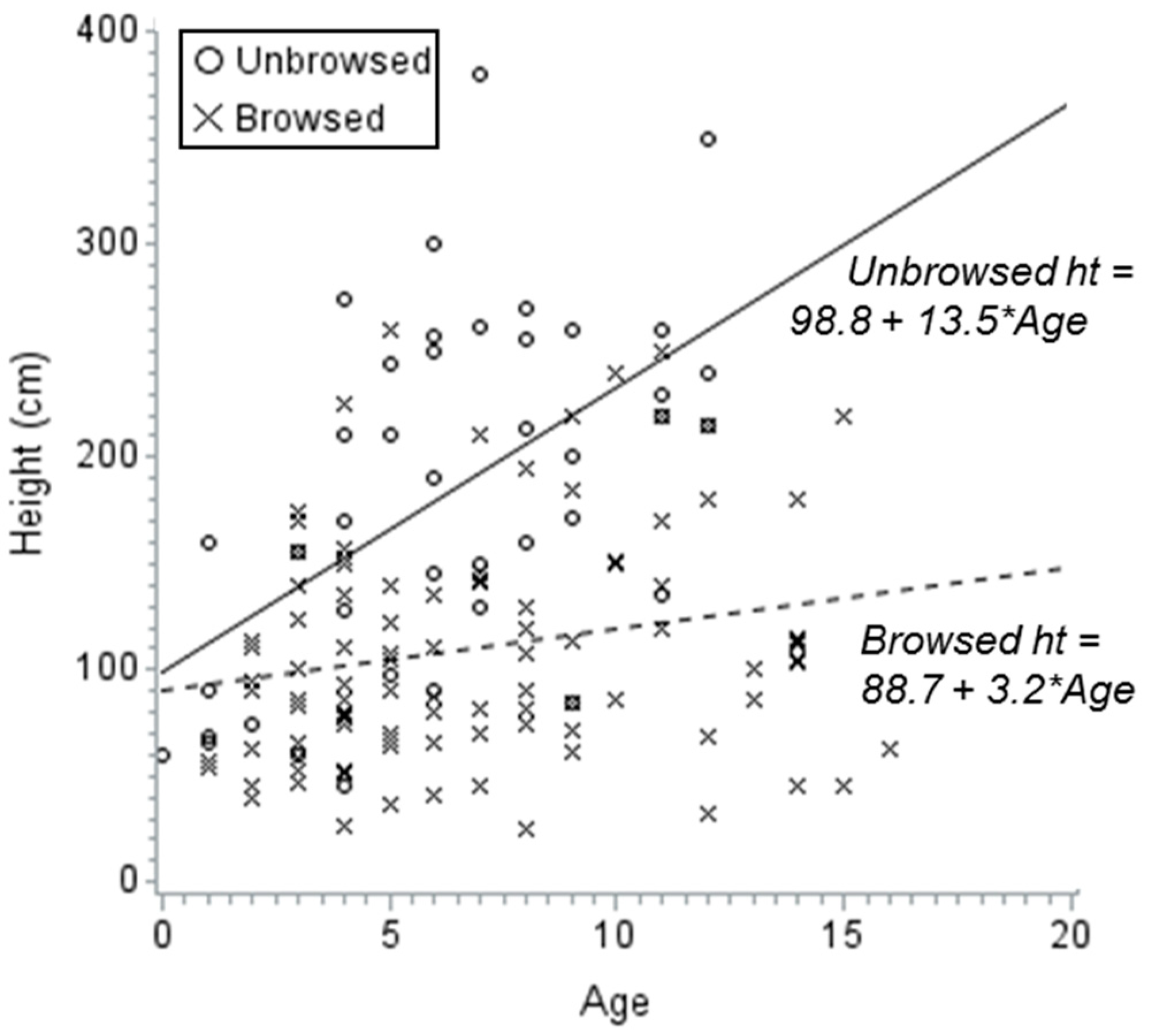
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Lotan, J.E.; Critchfield, W.B. Pinus contorta Doug. ex. Loud.—Lodgepole Pine. In Silvics of North America: Conifers; Burns, R.M., Honkala, B.H., Eds.; USDA Forest Service: Washington, DC, USA, 1990; Volume 1, pp. 302–315. [Google Scholar]
- Crouch, G.L. Aspen regeneration after commercial clearcutting in southwestern Colorado. J. For. 1983, 83, 316–319. [Google Scholar]
- Perala, D.A. Populus tremuloicies Michx. - Quaking Aspen. In Silvics of North America: Hardwoods; Burns, R.M., Honkala, B.H., Eds.; USDA Forest Service: Washington, DC, USA, 1990; Volume 2, pp. 555–569. [Google Scholar]
- Frey, B.; Lieffers, V.; Landhausser, S.; Comeau, P.; Greenway, K. An analysis of sucker regeneration of trembling aspen. Can. J. For. Res. 2003, 33, 1169–1179. [Google Scholar] [CrossRef]
- Miles, P.D. Forest Inventory EVALIDator Web-Application Version 1.6.0.03. Available online: http://apps.fs.fed.us/Evalidator/evalidator.jsp (accessed on 22 January 2015).
- Astrup, R.; Coates, K.D.; Hall, E. Recruitment limitation in forests: Lessons from an unprecedented mountain pine beetle epidemic. For. Ecol. Manag. 2008, 256, 1743–1750. [Google Scholar] [CrossRef]
- Teste, F.P.; Lieffers, V.J.; Landhäusser, S.M. Seed release in serotinous lodgepole pine forests after mountain pine beetle outbreak. Ecol. Appl. 2011, 21, 150–162. [Google Scholar] [CrossRef] [PubMed]
- Collins, B.J.; Rhoades, C.C.; Hubbard, R.M.; Battaglia, M.A. Tree regeneration and future stand development after bark beetle infestation and harvesting in Colorado lodgepole pine forests. For. Ecol. Manag. 2011, 261, 2168–2745. [Google Scholar] [CrossRef]
- Tinker, D.B.; Romme, W.H.; Hargrove, W.W.; Gardner, R.H.; Turner, M.G. Landscape-scale heterogeneity in lodgepole pine serotiny. Can. J. For. Res. 1994, 24, 897–903. [Google Scholar] [CrossRef]
- Shepperd, W.D. Initial Growth, Development, and Clonal Dynamics of Regenerated Aspen in the Rocky Mountains; USDA Forest Service: Fort Collins, CO, USA, 1993.
- Smith, E.A.; O’Loughlin, D.; Buck, J.R.; St. Clair, S.B. The influences of conifer succession, physiographic conditions and herbivory on quaking aspen regeneration after fire. For. Ecol. Manag. 2011, 262, 325–330. [Google Scholar] [CrossRef]
- Farmer, R.E. Effect of light intensity on growth of Populus tremuloides. For. Sci. 1962, 8, 403–410. [Google Scholar]
- Bartos, D.L.; Campbell, R.B. Decline of quaking aspen in the interior west-examples from Utah. Rangelands 1998, 20, 17–24. [Google Scholar]
- Worrall, J.J.; Marchetti, S.B.; Egeland, L.; Mask, R.A.; Eager, T.; Howell, B. Effects and etiology of sudden aspen decline in southwestern Colorado, USA. For. Ecol. Manag. 2010, 260, 638–648. [Google Scholar] [CrossRef]
- Rehfeldt, G.E.; Ferguson, D.E.; Crookston, N.L. Aspen, climate, and sudden decline in western USA. For. Ecol. Manag. 2009, 258, 2353–2364. [Google Scholar] [CrossRef]
- Huffman, R.D.; Fajvan, M.A.; Wood, P.B. Effects of residual overstory on aspen development in Minnesota. Can. J. For. Res. 1999, 29, 284–289. [Google Scholar] [CrossRef]
- Groot, A.; Man, R.; Wood, J. Spatial and temporal patterns of Populus tremuloides regeneration in small forest openings in northern Ontario. For. Chron. 2009, 85, 548–557. [Google Scholar] [CrossRef]
- Calder, W.J.; Horn, K.J.; St. Clair, S.B. Conifer expansion reduces the competitive ability and herbivore defense of aspen by modifying light environment and soil chemistry. Tree Phys. 2011, 31, 582–591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ripple, W.J.; Beschta, R.L. Restoring Yellowstone’s aspen with wolves. Biol. Cons. 2007, 138, 514–519. [Google Scholar] [CrossRef]
- Zeigenfuss, L.C.; Binkley, D.; Tuskan, G.A.; Romme, W.H.; Yin, T.; DiFazio, S.; Singer, F.J. Aspen Ecology in Rocky Mountain National Park: Age Distribution, Genetics, and the Effects of Elk Herbivory; U.S. Geological Survey: Reston, VA, USA, 2008.
- Nelson, K.N. The effect of mountain pine beetle caused mortality on subalpine forest stand and landscape structure in Rocky Mountain National Park, CO. Master’s Thesis, Colorado State University, Fort Collins, CO, USA, December 2009. [Google Scholar]
- Renwick, K.M. The interactive effects of climate and disturbance on tree species distributions. Ph.D. Thesis, Colorado State University, Fort Collins, CO, USA, May 2009. [Google Scholar]
- DeRose, R.J.; Long, J.N. Regeneration response and seedling bank dynamics on a Dendroctonus rufipennis-killed Picea engelmannii landscape. J. Veg. Sci. 2010, 21, 377–387. [Google Scholar] [CrossRef]
- Doesken, N.J.; Pielke Sr, R.A.; Bliss, O.A.P. Climate of Colorado, Climatography of the United States No. 60. Available online: http://climate.colostate.edu/pdfs/climateofcoloradoNo.60.pdf (accessed on 29 August 2018).
- USDA Forest Service. Forest insect and disease aerial survey data, Rocky Mountain Region. Available online: https://www.fs.usda.gov/detail/r2/forest-grasslandhealth/?cid=fsbdev3_041629 (accessed on 29 August 2018).
- USDA Natural Resources Conservation Service. National Range and Pasture Handbook; USDA Natural Resources Conservation Service: Washington, DC, USA, 2003.
- U.S. Geological Survey. National Elevation Dataset (NED), 2nd ed.; U.S. Geological Survey: Sioux Falls, SD, USA, 2009.
- Bevin, K.J.; Kirkby, M.J. A physically based, variable contributing area model of basin hydrology. Hydrol. Sci. Bull. 1979, 24, 43–69. [Google Scholar] [CrossRef]
- Franklin, J. Enhancing a regional vegetation map with predictive models of dominant plant species in chaparral. Appl. Veg. Sci. 2002, 5, 135–146. [Google Scholar] [CrossRef]
- Parolo, G.; Rossi, G.; Ferrarini, A. Toward improved species niche modelling: Arnica montana in the Alps as a case study. J. Appl. Ecol. 2008, 45, 1410–1418. [Google Scholar] [CrossRef]
- Evans, J.S.; Cushman, S.A. Gradient modeling of conifer species using random forests. Land. Ecol. 2009, 24, 673–683. [Google Scholar] [CrossRef]
- Dirnbock, T.; Hobbs, R.J.; Lambeck, R.J.; Caccetta, P.A. Vegetation distribution in relation to topographically driven processes in southwestern Australia. Appl. Veg. Sci. 2002, 5, 147–158. [Google Scholar] [CrossRef]
- Bader, M.Y.; Ruijten, J.J.A. A topography based model of forest cover at the alpine tree line in the tropical Andes. J. Biogeogr. 2008, 35, 711–723. [Google Scholar] [CrossRef]
- Burnham, K.P.; Anderson, D.R. Model Selection And Multimodel Inference: A Practical Information-Theoretic Approach; Springer: New York, NY, USA, 1998. [Google Scholar]
- Amoroso, M.M.; Coates, K.D.; Astrup, R. Stand recovery and self-organization following large-scale mountain pine beetle induced canopy mortality in northern forests. For. Ecol. Manag. 2013, 310, 300–311. [Google Scholar] [CrossRef]
- Binkley, D. Age distribution of aspen in Rocky Mountain National Park, USA. For. Ecol. Manag. 2008, 255, 797–802. [Google Scholar] [CrossRef]
- Romme, W.H.; Turner, M.G.; Wallace, L.L.; Walker, J.S. Aspen, elk, and fire in northern Yellowstone National Park. Ecology 1995, 76, 2097–2106. [Google Scholar] [CrossRef]
- Relva, M.A.; Westerholm, C.L.; Kitzberger, T. Effects of introduced ungulates on forest understory communities in northern Patagonia are modified by timing and severity of stand mortality. Plant Ecol. 2009, 201, 11–22. [Google Scholar] [CrossRef]
- Griffin, J.M.; Turner, M.G. Changes to the N cycle following bark beetle outbreaks in two contrasting conifer types. Oecologia 2012, 170, 551–565. [Google Scholar] [CrossRef] [PubMed]
- Stone, W.E.; Wolfe, M.L. Response of understory vegetation to variable tree mortality following a mountain pine beetle epidemic in lodgepole pine stands in northern Utah. Vegetatio 1996, 122, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Ripple, W.J.; Larsen, E.J. The role of postfire coarse woody debris in aspen regeneration. West. J. Appl. For. 2001, 16, 61–64. [Google Scholar]
- De Chantal, M.; Granstrom, A. Aggregations of dead wood after wildfire act as browsing refugia for seedlings of Populus tremula and Salix caprea. For. Ecol. Manag. 2007, 250, 3–8. [Google Scholar] [CrossRef]
- Forester, J.D.; Anderson, D.P.; Turner, M.G. Do high-density patches of coarse wood and regenerating saplings create browsing refugia fore aspen (Populus tremuloides Michx.) in Yellowstone National Park (USA)? For. Ecol. Manag. 2007, 253, 211–219. [Google Scholar] [CrossRef]
- Bates, P.C.; Blinn, C.R.; Alm, A.A. A survey of the harvesting histories of some poorly regenerated aspen stands in northern Minnesota. In Proceedings of the Aspen Symposium ’89; Adams, R.D., Ed.; USDA Forest Service: St. Paul, MN, USA, 1990; pp. 221–230. [Google Scholar]
- Dudley, M.M.; Burns, K.S.; Jacobi, W.R. Aspen mortality in the Colorado and southern Wyoming Rocky Mountains: Extent, severity, and causal factors. For. Ecol. Manag. 2015, 353, 240–259. [Google Scholar] [CrossRef] [Green Version]
- Stahelin, R. Factors influencing natural restocking of high altitude burns by coniferous trees in the central Rocky Mountains. Ecology 1943, 24, 19–30. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Median | Minimum | Maximum | |||
|---|---|---|---|---|---|
| Basal area m2 ha−1 | |||||
| LIVE | 7.6 | 1.1 | 37.0 | ||
| Pine | 3.3 | 0.0 | 22.9 | ||
| Aspen | 2.2 | 1.1 | 14.2 | ||
| Fir+Spruce | 0.0 | 0.0 | 2.2 | ||
| DEAD | 21.8 | 4.4 | 34.8 | ||
| Pine | Standing | 20.7 | 3.3 | 34.8 | |
| Down | 3.3 | 0.0 | 24.0 | ||
| Aspen | Standing | 2.2 | 0.0 | 10.5 | |
| Down | 2.2 | 0.0 | 13.1 | ||
| Fir + Spruce | Standing | 0.0 | 0.0 | 0.9 | |
| Down | 0.0 | 0.0 | 0.0 | ||
| Density ha−1 | |||||
| UNDERSTORY (Stems <2.5 cm diameter at breast height) | |||||
| Pine | 556 | 0 | 7040 | ||
| Aspen | 6175 | 1167 | 26861 | ||
| Fir + Spruce | 62 | 0 | 3520 | ||
| Response Variable | Independent Variable | +/− | Adjusted R2 | P |
|---|---|---|---|---|
| Mean aspen density | Down lodgepole pine | − | 0.22 | 0.0038 |
| Mean annual maximum temperature | − | 0.20 | 0.0051 | |
| Live aspen | − | 0.11 | 0.0323 | |
| Dead and down aspen | − | 0.11 | 0.0351 | |
| Mean annual precipitation | + | 0.07 | 0.0791 | |
| Mean lodgepole density | Live lodgepole pine basal area | + | 0.23 | 0.0027 |
| Topographic Wetness Index | + | 0.11 | 0.0349 | |
| Time since onset of MPB epidemic | − | 0.02 | 0.2206 | |
| Down lodgepole pine | − | −0.02 | 0.5497 | |
| Mean aspen sucker height | Down lodgepole pine | + | 0.49 | <0.0001 |
| Maximum temperature | + | 0.38 | <0.0001 | |
| Wild animal density | − | 0.27 | 0.0012 | |
| Mean annual precipitation | − | 0.23 | 0.0027 | |
| Time since onset of MPB epidemic | + | 0.06 | 0.0970 | |
| Maximum aspen sucker height | Down lodgepole pine | + | 0.39 | <0.0001 |
| Mean annual precipitation | − | 0.28 | 0.0012 | |
| Mean annual maximum temperature | + | 0.27 | 0.0010 | |
| Wild animal density | − | 0.17 | 0.0109 | |
| Topographic Wetness Index | − | 0.12 | 0.0287 | |
| Live aspen basal area | − | 0.09 | 0.0483 | |
| MPB-killed basal area | − | 0.06 | 0.0882 |
| Response Variable | Independent Variable | +/− | Adjusted R2 | P |
|---|---|---|---|---|
| Proportion of suckers browsed | Down lodgepole pine basal area | − | 0.46 | <0.0001 |
| MPB-killed basal area | + | 0.25 | 0.0027 | |
| Wild animal density | + | 0.16 | 0.0111 | |
| Live aspen basal area | − | −0.03 | 0.9437 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pelz, K.A.; Smith, F.W. Effects of Stand Structure, Browsing, and Biophysical Conditions on Regeneration Following Mountain Pine Beetle in Mixed Lodgepole Pine and Aspen Forests of the Southern Rockies. Forests 2018, 9, 525. https://0-doi-org.brum.beds.ac.uk/10.3390/f9090525
Pelz KA, Smith FW. Effects of Stand Structure, Browsing, and Biophysical Conditions on Regeneration Following Mountain Pine Beetle in Mixed Lodgepole Pine and Aspen Forests of the Southern Rockies. Forests. 2018; 9(9):525. https://0-doi-org.brum.beds.ac.uk/10.3390/f9090525
Chicago/Turabian StylePelz, Kristen A., and Frederick W. Smith. 2018. "Effects of Stand Structure, Browsing, and Biophysical Conditions on Regeneration Following Mountain Pine Beetle in Mixed Lodgepole Pine and Aspen Forests of the Southern Rockies" Forests 9, no. 9: 525. https://0-doi-org.brum.beds.ac.uk/10.3390/f9090525