Anatomical and Chemical Responses of Eastern White Pine (Pinus strobus L.) to Blue-Stain (Ophiostoma minus) Inoculation

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Field Study
2.2. Sample Collection
2.3. Fungal Identification by DNA Sequence Analysis
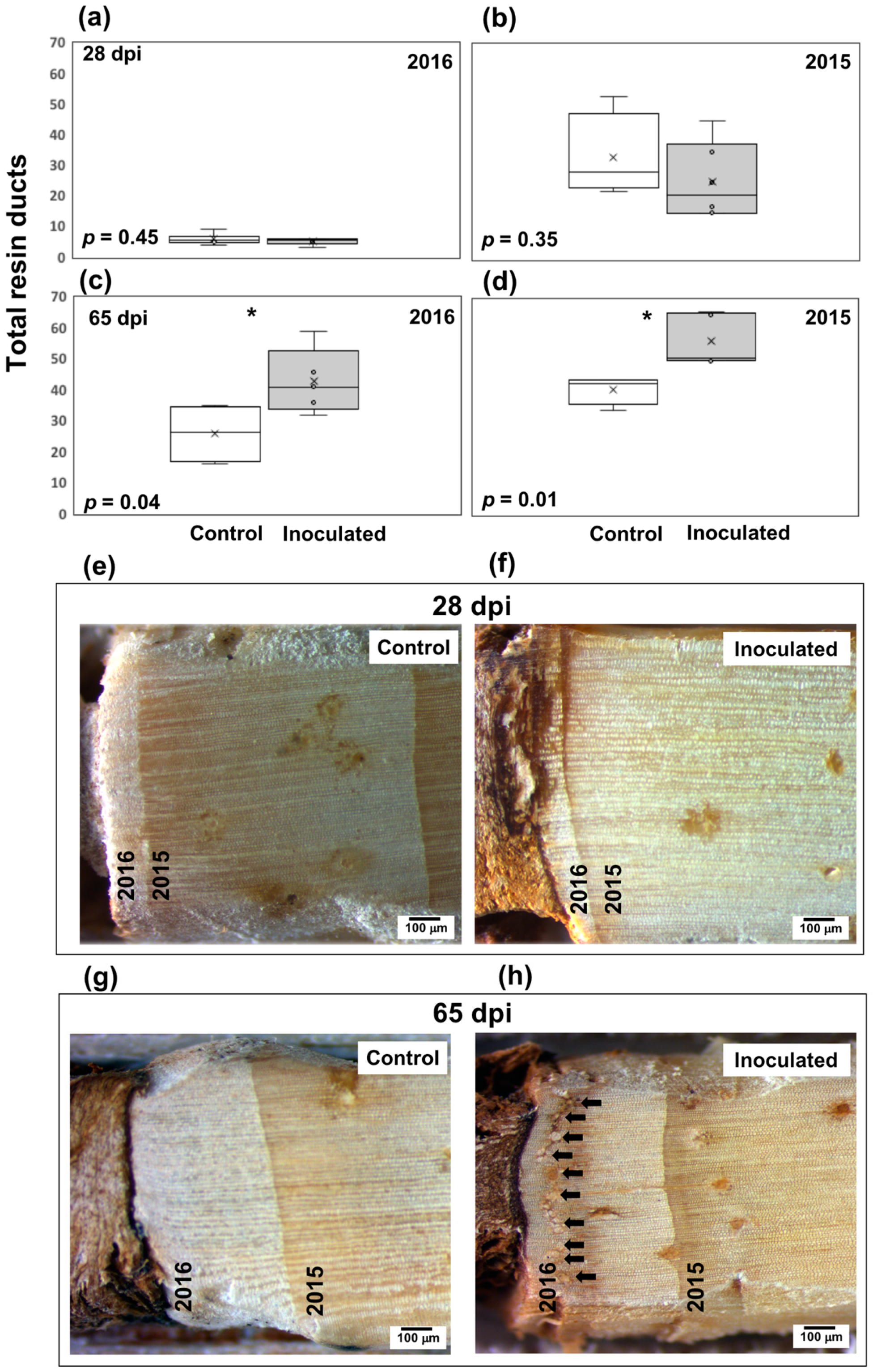
2.4. Resin Canal Traits
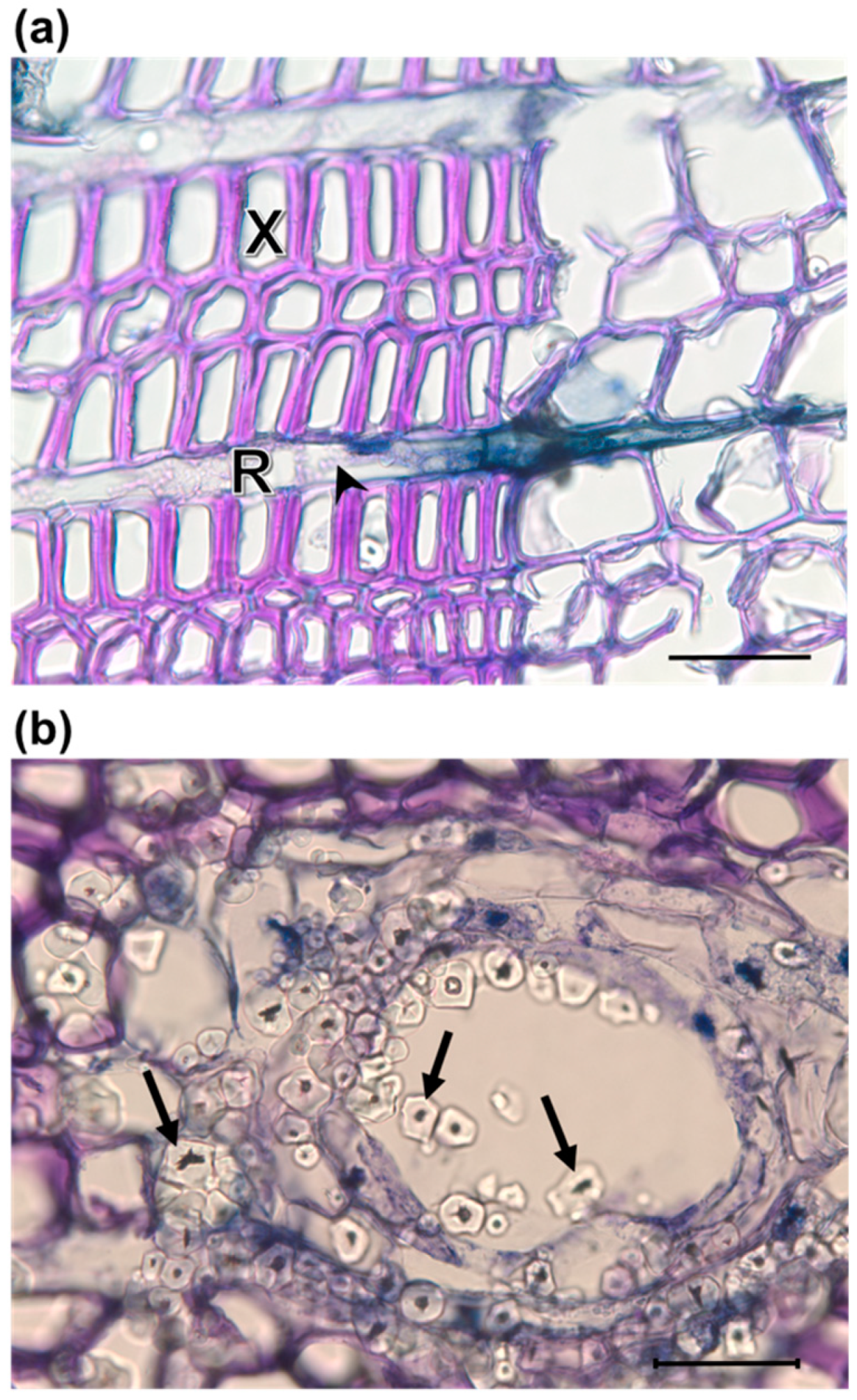
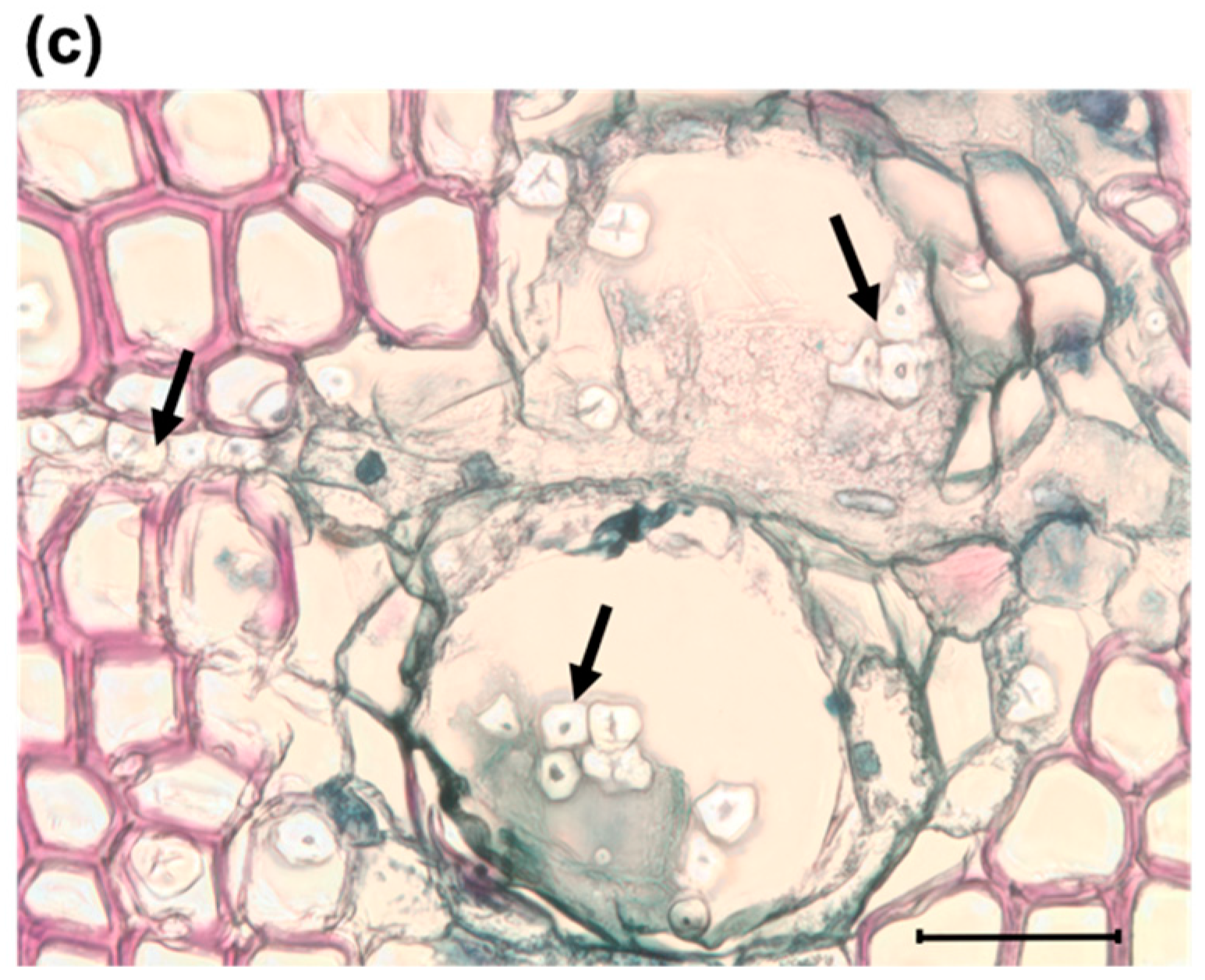
2.5. Detection of Fungus and Anatomical Traits for Plant Defense
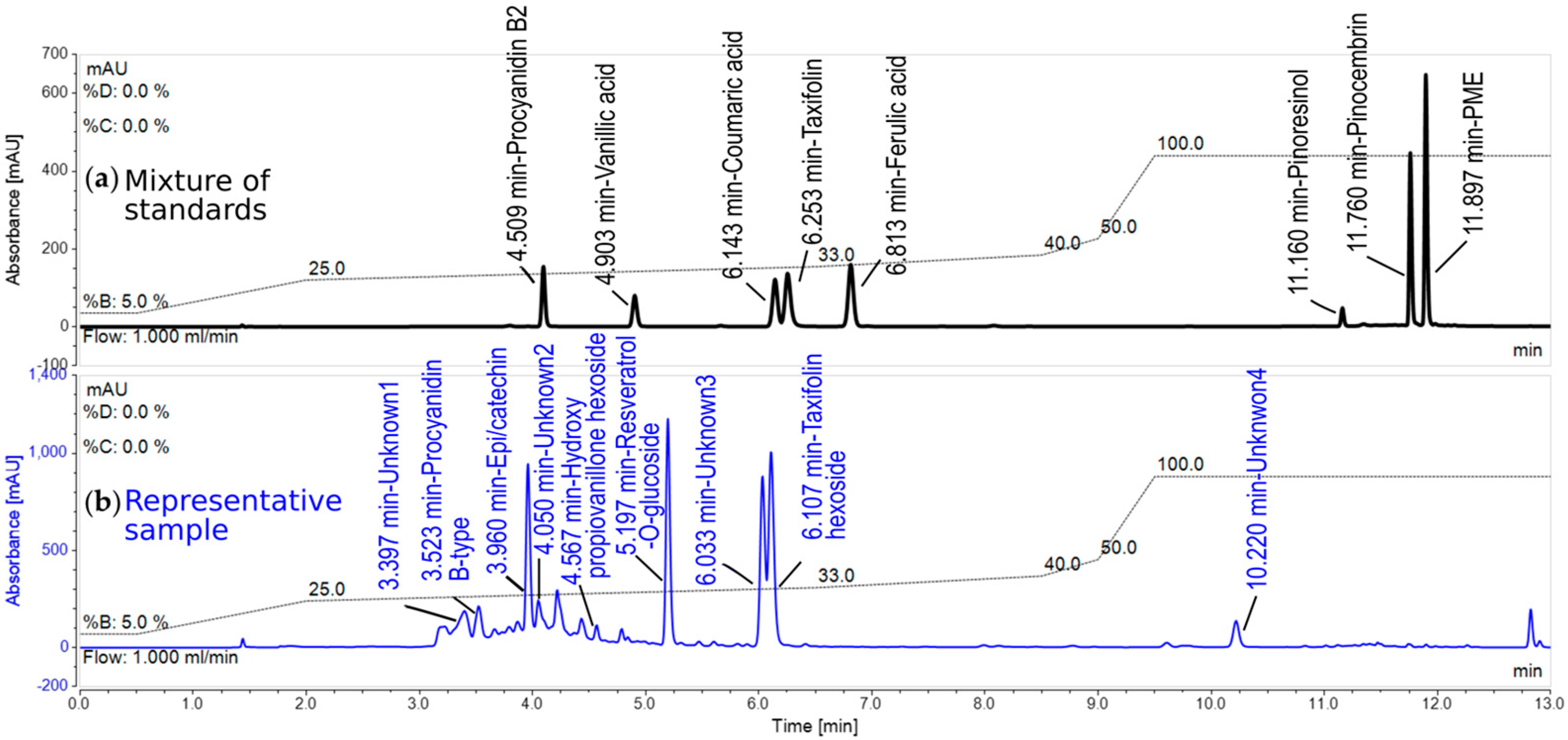
2.6. Analysis of Phenolic Compounds
2.7. Analysis of Terpenoids
2.8. Statistical Analyses
3. Results
3.1. Anatomical Responses to Fungal Inoculation
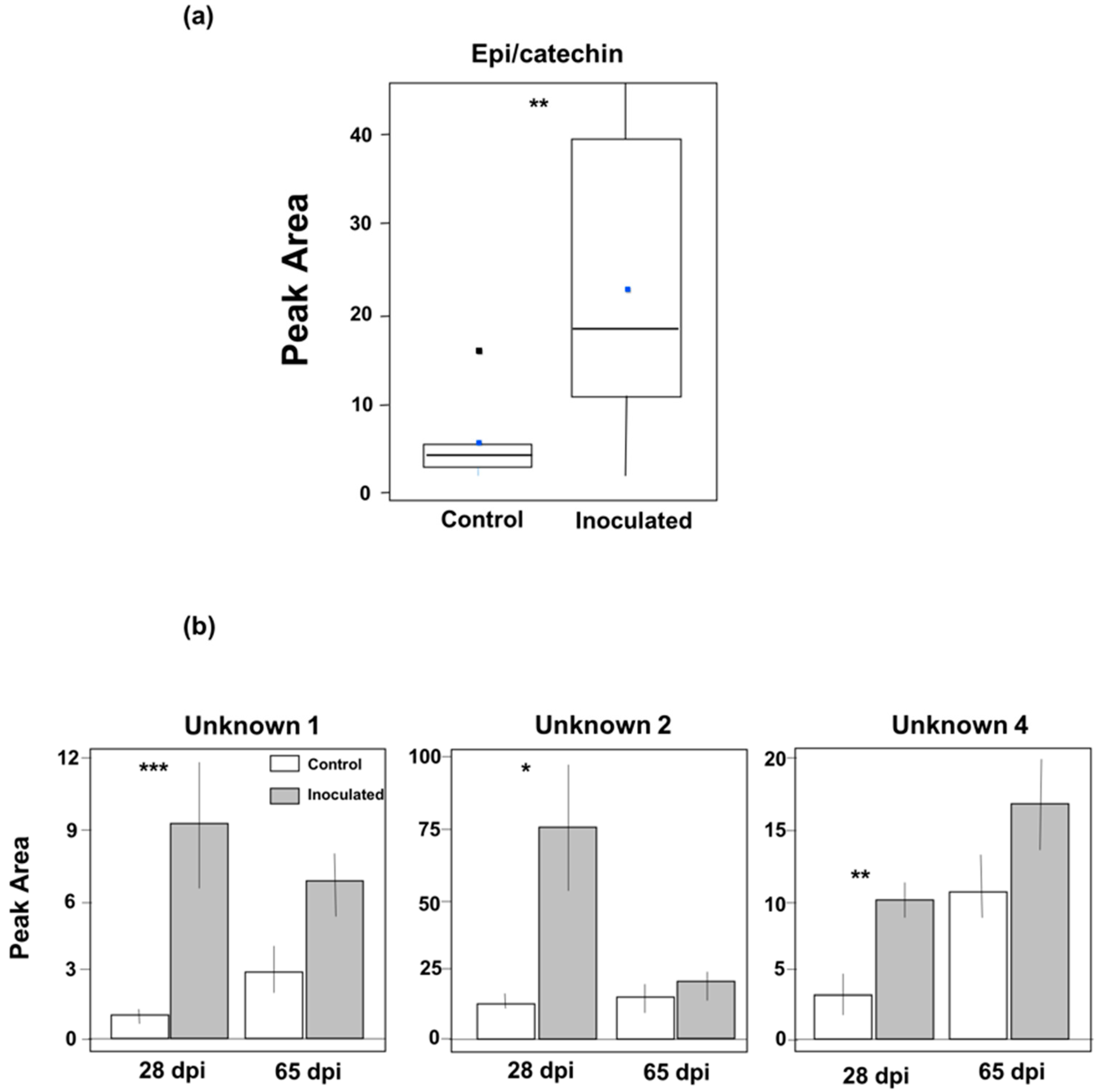
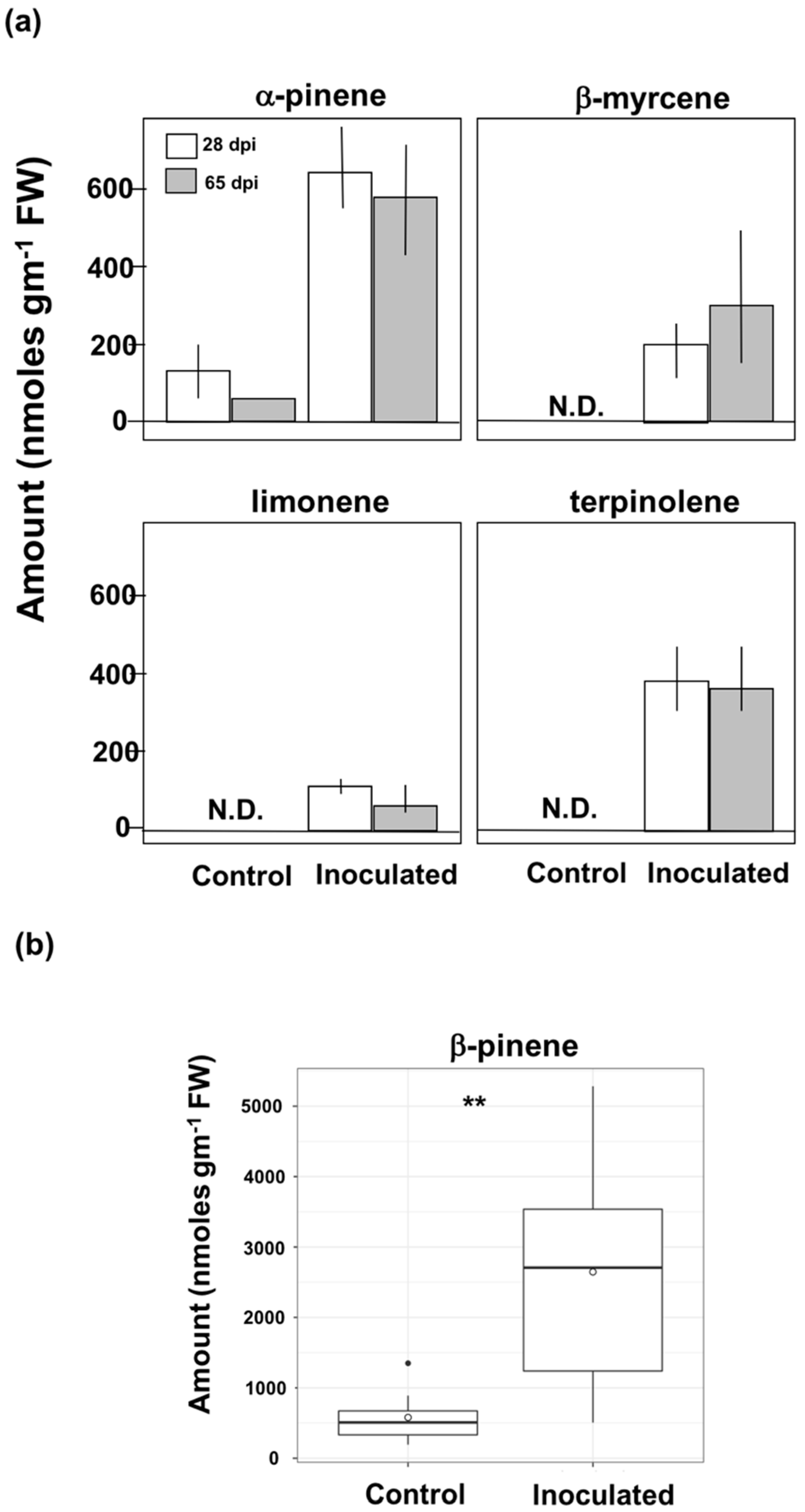
3.2. Induction of Phenolic Metabolites and Terpenoids
4. Discussion
4.1. Anatomical Responses to O. minus
4.2. Phenolics and Terpenoids Involved in Eastern White Pine Induced Response
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Nowak, J.T.; Klepzig, K.D.; Coyle, D.R.; Carothers, W.A.; Gandhi, K.J.K. Southern pine beetles in central hardwood forests: Frequency, spatial extent, and changes to forest structure. In Natural Disturbances and Historic Range of Variation: Type, Frequency, Severity, and Post-Disturbance Structure in Central Hardwood Forests; Greenberg, C.H., Collins, B.S., Eds.; Springer: Cham, Switzerland, 2016; pp. 73–88. [Google Scholar]
- Dodds, K.J.; Aoki, C.F.; Arango-Velez, A.; Cancelliere, J.; D’Amato, A.W.; DiGirolomo, M.F.; Rabaglia, R.J. Expansion of southern pine beetle into northeastern forests: Management and impact of a primary bark beetle in a new region. J. For. 2018, 116, 178–191. [Google Scholar] [CrossRef]
- Raffa, K.F.; Aukema, B.H.; Bentz, B.J.; Carroll, A.L.; Hicke, J.A.; Kolb, T.E. Responses of tree-killing bark beetles to a changing climate. Clim. Chang. Insect Pests 2015, 173–201. [Google Scholar] [CrossRef]
- Asaro, C.; Nowak, J.T.; Elledge, A. Why have southern pine beetle outbreaks declined in the southeastern U.S. with the expansion of intensive pine silviculture? A brief review of hypotheses. For. Ecol. Manag. 2017, 391, 338–348. [Google Scholar] [CrossRef]
- Bridges, J.R.; Moser, J.C. Role of two phoretic mites in transmission of bluestain fungus, Ceratocystis minor. Ecol. Entomol. 1983, 8, 9–12. [Google Scholar] [CrossRef]
- Bridges, J.R.; Moser, J.C. Relationship of phoretic mites (Acari: Tarsonemidae) to the bluestaining fungus, Ceratocystis minor, in trees infested by southern pine beetle (Coleoptera: Scolytidae). Environ. Entomol. 1986, 15, 951–953. [Google Scholar] [CrossRef]
- Barras, S.J. Antagonism between Dendroctonus frontalis and the fungus Ceratocystis minor. Ann. Entomol. Soc. Am. 1970, 63, 1187–1190. [Google Scholar] [CrossRef]
- Klepzig, K.D.; Robison, D.J.; Fowler, G.; Minchin, P.R.; Hain, F.P.; Allen, H.L. Effects of mass inoculation on induced oleoresin response in intensively managed loblolly pine. Tree Physiol. 2005, 25, 681–688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lieutier, F.; Yart, A.; Salle, A. Stimulation of tree defenses by Ophiostomatoid fungi can explain attack success of bark beetles on conifers. Ann. For. Sci. 2009, 66, 801. [Google Scholar] [CrossRef]
- Klepzig, K.D.; Wilkens, R.T. Competitive interactions among symbiotic fungi of the mountain pine beetle. Appl. Environ. Microbiol. 1997, 63, 621–627. [Google Scholar] [PubMed]
- Bridges, J.R.; Nettleton, W.A.; Connor, M.D. Southern pine beetle (Coleoptera: Scolytidae) infestations without the bluestain fungus, Ceratocystis minor. J. Econ. Entomol. 1985, 78, 325–327. [Google Scholar] [CrossRef]
- Basham, H.G. Wilt loblolly pine inoculated with blue-stain fungi of the genus Ceratocystis. Phytopathology 1970, 60, 750–754. [Google Scholar] [CrossRef]
- Arango-Velez, A.; El Kayal, W.; Copeland, C.C.J.; Zaharia, L.I.; Lusebrink, I.; Cooke, J.E.K. Differences in defence responses of Pinus contorta and Pinus banksiana to the mountain pine beetle fungal associate Grosmannia clavigera are affected by water deficit. Plant Cell Environ. 2016, 39, 726–744. [Google Scholar] [CrossRef] [PubMed]
- Bohlmann, J. Pine terpenoid defences in the mountain pine beetle epidemic and in other conifer pest interactions: Specialized enemies are eating holes into a diverse, dynamic and durable defence system. Tree Physiol. 2012, 32, 943–945. [Google Scholar] [CrossRef] [PubMed]
- Franceschi, V.R.; Krokene, P.; Krekling, T.; Christiansen, E. Phloem parenchyma cells are involved in local and distant defense responses to fungal inoculation or bark-beetle attack in Norway spruce (Pinaceae). Am. J. Bot. 2000, 87, 314–326. [Google Scholar] [CrossRef] [PubMed]
- Lippert, D.; Chowrira, S.; Ralph, S.G.; Zhuang, J.; Aeschliman, D.; Ritland, C.; Ritland, K.; Bohlmann, J. Conifer defense against insects: Proteome analysis of Sitka spruce (Picea sitchensis) bark induced by mechanical wounding or feeding by white pine weevils (Pissodes strobi). Proteomics 2007, 7, 248–270. [Google Scholar] [CrossRef] [PubMed]
- Raffa, K.F.; Aukema, B.; Erbilgin, N.; Klepzig, K.D.; Wallin, K.F. Interactions among conifer terpenoids and bark beetles across multiple levels of scale: An attempt to understand links between population patterns and physiological processes. Recent Adv. Phytochem. 2005, 39, 79–118. [Google Scholar]
- Aukema, B.H.; Powell, J.S.; Clayton, M.K.; Raffa, K.F. Variation in complex semiochemical signals arising from insects and host plants. Environ. Entomol. 2010, 39, 874–882. [Google Scholar] [CrossRef] [PubMed]
- Boone, C.K.; Aukema, B.H.; Bohlmann, J.; Carroll, A.L.; Raffa, K.F. Efficacy of tree defense physiology varies with bark beetle population density: A basis for positive feedback in eruptive species. Can. J. For. Res. 2011, 1188, 1174–1188. [Google Scholar] [CrossRef]
- Mason, C.J.; Klepzig, K.D.; Kopper, B.J.; Kersten, P.J.; Illman, B.L.; Raffa, K.F. Contrasting patterns of diterpene acid induction by red pine and white spruce to simulated bark beetle attack, and interspecific differences in sensitivity among fungal associates. J. Chem. Ecol. 2015, 41, 524–532. [Google Scholar] [CrossRef] [PubMed]
- Rosenberger, D.W.; Venette, R.C.; Maddox, M.P.; Aukema, B.H. Colonization behaviors of mountain pine beetle on novel hosts: Implications for range expansion into Northeastern North America. PLoS ONE 2017, 12, e0176269. [Google Scholar] [CrossRef] [PubMed]
- Lieutier, F.; Brignolas, F.; Sauvard, D.; Yart, A.; Galet, C.; Brunet, M.; Van De Sype, H. Intra- and inter-provenance variability in phloem phenols of Picea abies and relationship to a bark beetle-associated fungus. Tree Physiol. 2003, 23, 247–256. [Google Scholar] [CrossRef] [PubMed]
- Nerg, A.A.; Kainulainen, P.; Vuorinen, M.; Hanso, M.; Holopainen, J.K.; Kurkela, T. Seasonal and geographical variation of terpenes, resin acids and total phenolics in nursery grown seedlings of scots pine (Pinus sylvestris L.). New Phytol. 2008, 128, 703–713. [Google Scholar] [CrossRef]
- Mason, C.J.; Keefover-Ring, K.; Villari, C.; Klutsch, J.G.; Cook, S.; Bonello, P.; Erbilgin, N.; Raffa, K.F.; Townsend, P.A. Anatomical defenses against bark beetles relate to degree of historical exposure between species and are allocated independently of chemical defenses within trees. Plant Cell Environ. 2018. [Google Scholar] [CrossRef]
- Cook, S.P.; Hain, F.P. Qualitative examination of the hypersensitive response of loblolly pine, Pinus taeda L., inoculated with 2 fungal associates of the southern pine-beetle, Dendroctonus frontalis Zimmermann (Coleoptera, Scolytidae). Environ. Entomol. 1985, 14, 396–400. [Google Scholar] [CrossRef]
- Cook, S.P.; Hain, F.P. Defensive mechanisms of Loblolly pine and Shortleaf pine against attack by southern pine beetle, Dendroctonus frontalis Zimmermann, and its fungal associate, Ceratocystis minor. Chem. Ecol. 1986, 12, 1397–1406. [Google Scholar] [CrossRef] [PubMed]
- Arango-Velez, A.; González, L.M.G.; Meents, M.J.; El Kayal, W.; Cooke, B.J.; Linsky, J.; Lusebrink, I.; Cooke, J.E.K. Influence of water deficit on the molecular responses of Pinus contorta × Pinus banksiana mature trees to infection by the mountain pine beetle fungal associate, Grosmannia clavigera. Tree Physiol. 2013, 34, 1220–1239. [Google Scholar] [CrossRef] [PubMed]
- Raffa, K.F.; Smalley, E.B. Interaction of pre-attack and induced monoterpene concentrations in host conifer defense against bark beetle-fungal complexes. Oecologia 1995, 102, 285–295. [Google Scholar] [CrossRef] [PubMed]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J.W. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press, Inc.: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Madan, A. CAP3: A DNA sequence assembly program. Genome Res. 1999, 868–877. [Google Scholar] [CrossRef]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; Mcgettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, M.; Zaretskaya, I.; Raytselis, Y.; Merezhuk, Y.; Mcginnis, S.; Madden, T.L. NCBI BLAST: A better web interface. Nucleic Acid Res. 2008, 36, 5–9. [Google Scholar] [CrossRef] [PubMed]
- Ferrenberg, S.; Kane, J.M.; Mitton, J.B. Resin duct characteristics associated with tree resistance to bark beetles across lodgepole and limber pines. Oecologia 2014, 174, 1283–1292. [Google Scholar] [CrossRef] [PubMed]
- Stokes, M.A.; Smiley, T.L. An Introduction to Tree Ring Dating; The University of Chicago Press: Chicago, IL, USA, 1968. [Google Scholar]
- Pearce, R.B. Staining fungal hyphae in wood. Trans. Br. Mycol. Soc. 1984, 82, 564–567. [Google Scholar] [CrossRef]
- Villari, C.; Battisti, A.; Chakraborty, S.; Michelozzi, M.; Bonello, P.; Faccoli, M. Nutritional and pathogenic fungi associated with the pine engraver beetle trigger comparable defenses in Scots pine. Tree Physiol. 2012, 32, 867–879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- R Development Core Team. R Foundation for Statistical Computing. Available online: https://www.r-project.org (accessed on 30 September 2018).
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2009. [Google Scholar]
- De Mendiburu, F. Statistical Procedures for Agricultural Research. Available online: http://CRAN.R-project.org/package=agricolae (accessed on 30 September 2018).
- Wei, J.; Carrol, R.J.; Harden, K.K.; Wu, G. Comparisons of treatment means when factors do not interact in two-factorial studies. Amino Acids 2012, 42, 2031–2035. [Google Scholar] [CrossRef] [PubMed]
- Hudgins, J.W.; Christiansen, E.; Franceschi, V.R. Induction of anatomically based defense responses in stems of diverse conifers by methyl jasmonate: A phylogenetic perspective. Tree Physiol. 2004, 24, 251–264. [Google Scholar] [CrossRef] [PubMed]
- Pearce, B.R.B. Tansley Review No. 87 Antimicrobial defences in the wood of living trees. New Phytol. 1996, 132, 203–233. [Google Scholar] [CrossRef]
- Stephen, F.M.; Paine, T.D. Seasonal patterns of host tree resistance to fungal associates of the southern pine beetle. J. Appl. Entomol. 1985, 99, 113–122. [Google Scholar]
- Franceschi, V.R.; Krokene, P.; Christiansen, E.; Krekling, T. Anatomical and chemical defenses of conifer bark against bark beetles and other pests. New Phytol. 2005, 167, 353–375. [Google Scholar] [CrossRef] [PubMed]
- Krokene, P.; Solheim, H.; Krekling, T.; Christiansen, E. Inducible anatomical defense responses in Norway spruce stems and their possible role in induced resistance. Tree Physiol. 2003, 23, 191–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trapp, S.; Croteau, R. Defensive resin biosynthesis in conifers. Annu. Rev. Plant Physiol. Plant Mol. Biol. 2001, 52, 689–724. [Google Scholar] [CrossRef] [PubMed]
- Goodsman, D.W.; Lusebrink, I.; Landhäusser, S.M.; Erbilgin, N.; Lieffers, V.J. Variation in carbon availability, defense chemistry and susceptibility to fungal invasion along the stems of mature trees. New Phytol. 2013, 197, 586–594. [Google Scholar] [CrossRef] [PubMed]
- Bonello, P.; Blodgett, J.T. Pinus nigra–Sphaeropsis sapinea as a model pathosystem to investigate local and systemic effects of fungal infection of pines. Physiol. Mol. Plant Pathol. 2003, 63, 249–261. [Google Scholar] [CrossRef]
- Bonello, P.; Heller, W.; Sandermann, H. Ozone effects on root-disease susceptibility and defence responses in mycorrhizal and non-mycorrhizal seedlings of Scots pine (Pinus sylvestris L.). New Phytol. 1993, 12, 653–663. [Google Scholar] [CrossRef]
- Keefover-Ring, K.; Trowbridge, A.; Mason, C.J.; Raffa, K.F. Rapid induction of multiple terpenoid groups by ponderosa pine in response to bark beetle-associated fungi. J. Chem. Ecol. 2016, 42, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Raffa, K.F.; Berryman, A.A. Interacting selective pressures in conifer-bark beetle systems: A basis for reciprocal adaptations? Am. Nat. 1987, 129, 234–236. [Google Scholar] [CrossRef]
- Cale, J.A.; Muskens, M.; Najar, A.; Ishangulyyeva, G.; Hussain, A.; Kanekar, S.S.; Klutsch, J.G.; Taft, S.; Erbilgin, N. Rapid monoterpene induction promotes the susceptibility of a novel host pine to mountain pine beetle colonization but not to beetlevectored fungi. Tree Physiol. 2017, 37, 1597–1610. [Google Scholar] [CrossRef] [PubMed]
- Clark, E.L.; Huber, D.P.W.; Carroll, A.L. The legacy of attack: Implications of high phloem resin monoterpene levels in lodgepole pines following mass attack by mountain pine beetle, Dendroctonus ponderosae; Hopkins. Environ. Entomol. 2012, 41, 392–398. [Google Scholar] [CrossRef] [PubMed]








© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arango-Velez, A.; Chakraborty, S.; Blascyk, K.; Phan, M.T.; Barsky, J.; El Kayal, W. Anatomical and Chemical Responses of Eastern White Pine (Pinus strobus L.) to Blue-Stain (Ophiostoma minus) Inoculation. Forests 2018, 9, 690. https://0-doi-org.brum.beds.ac.uk/10.3390/f9110690
Arango-Velez A, Chakraborty S, Blascyk K, Phan MT, Barsky J, El Kayal W. Anatomical and Chemical Responses of Eastern White Pine (Pinus strobus L.) to Blue-Stain (Ophiostoma minus) Inoculation. Forests. 2018; 9(11):690. https://0-doi-org.brum.beds.ac.uk/10.3390/f9110690
Chicago/Turabian StyleArango-Velez, Adriana, Sourav Chakraborty, Kevin Blascyk, Mi T. Phan, Joseph Barsky, and Walid El Kayal. 2018. "Anatomical and Chemical Responses of Eastern White Pine (Pinus strobus L.) to Blue-Stain (Ophiostoma minus) Inoculation" Forests 9, no. 11: 690. https://0-doi-org.brum.beds.ac.uk/10.3390/f9110690