Allelopathic Effects of Aqueous Leaf Extracts from Four Shrub Species on Seed Germination and Initial Growth of Amygdalus pedunculata Pall.

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Seed and Plant Material
2.2. The Aqueous Leaf Extracts Preparation
2.3. Seed Germination Experiment
2.4. Seedling Growth Experiment
2.5. Statistical Analysis
3. Results
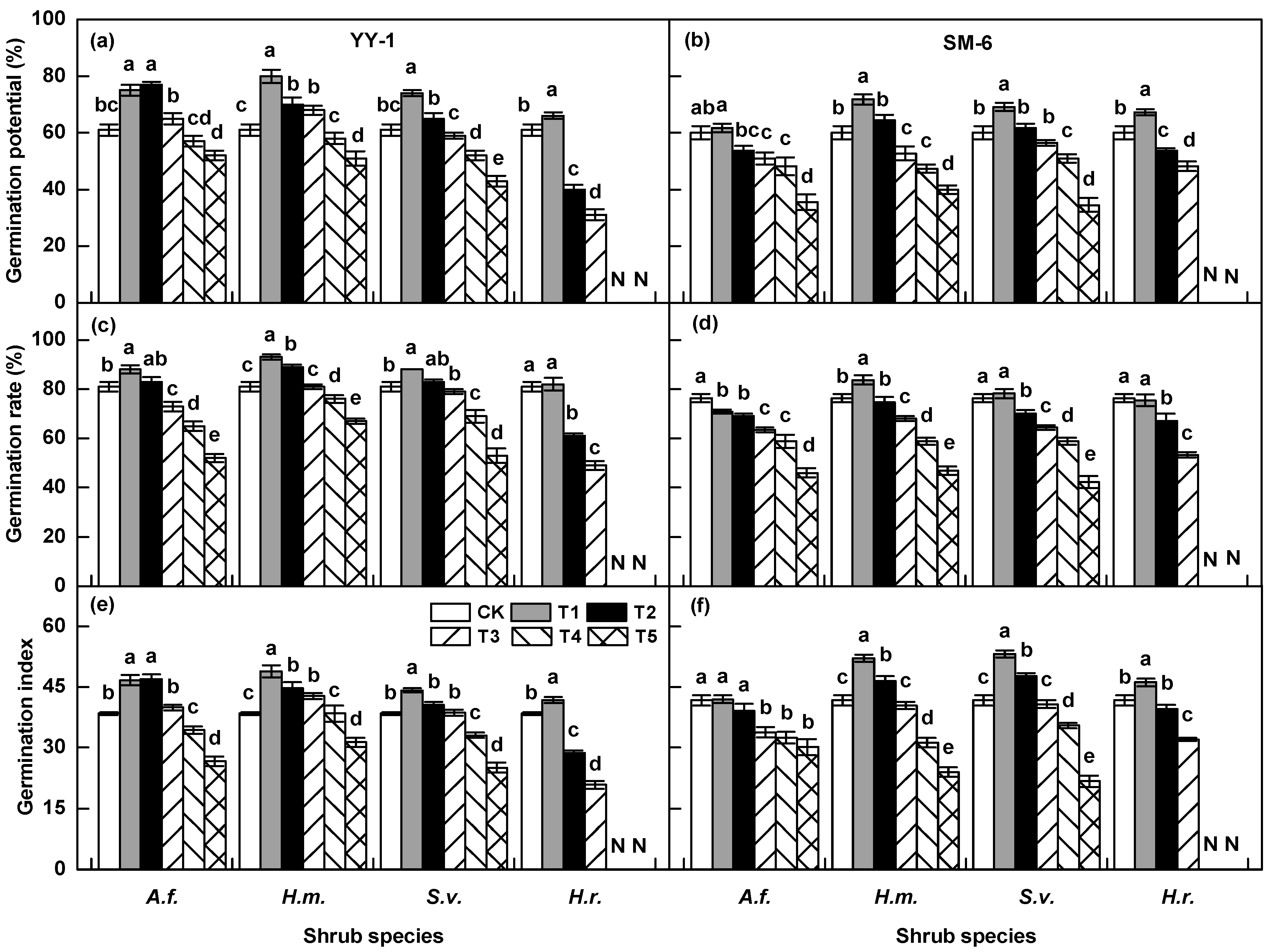
3.1. Effects of Shrub Aqueous Leaf Extracts on A. pedunculata Seed Germination
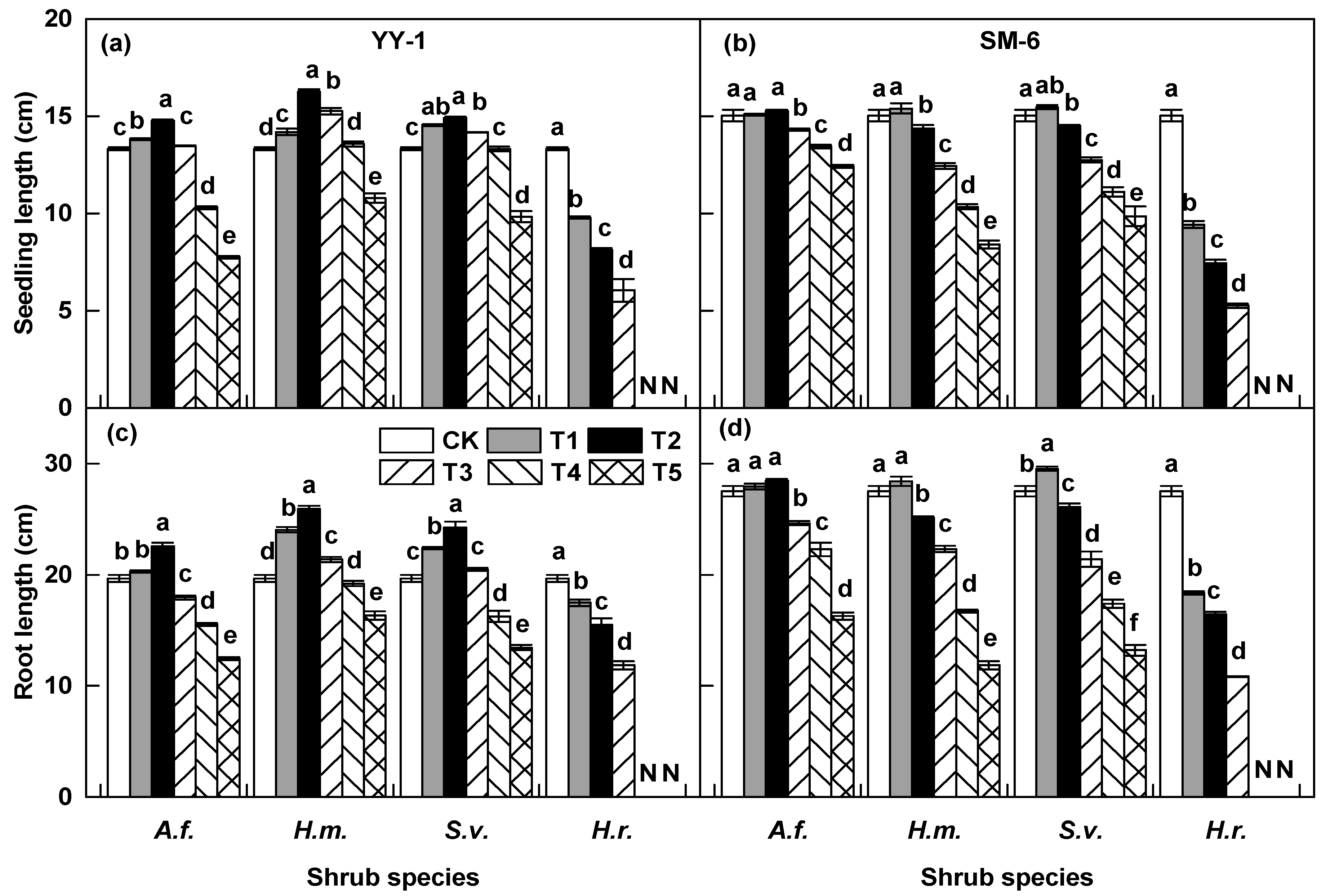
3.2. Effects of Shrub Aqueous Leaf Extracts on A. pedunculata Seedling Growth
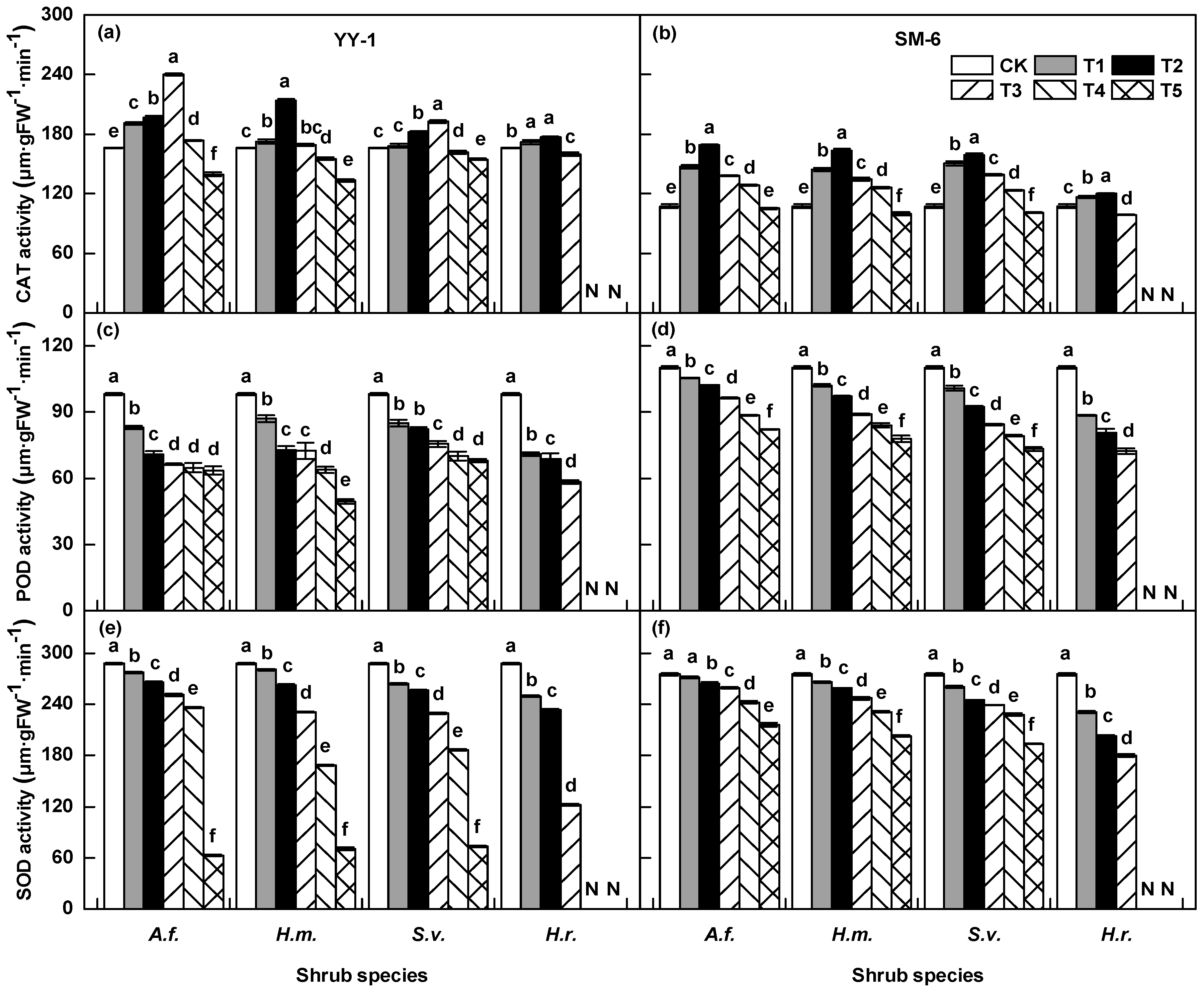
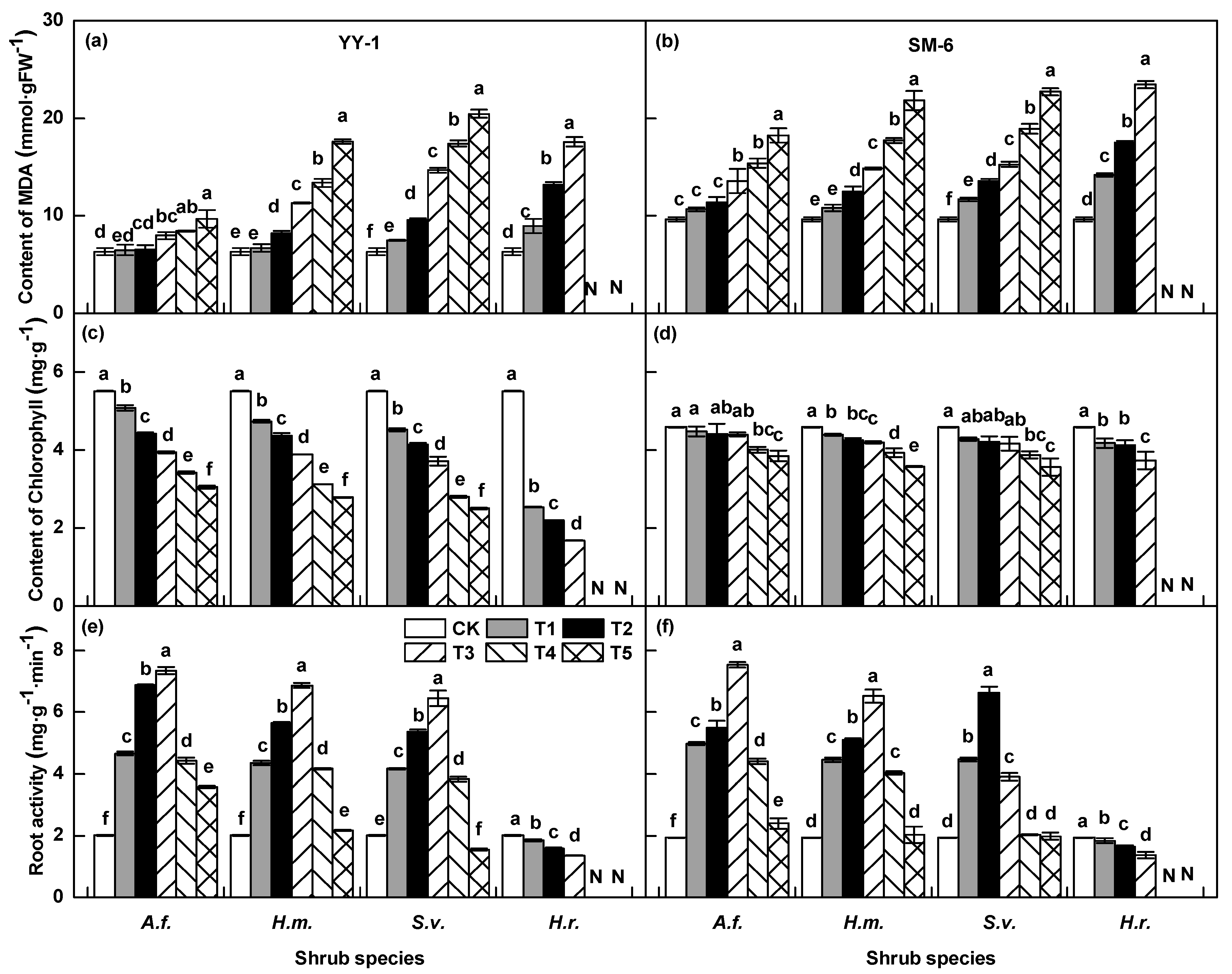
3.3. Effects of Shrub Aqueous Leaf Extracts on Physiological Traits of A. pedunculata Seedlings
3.3.1. Enzyme Activity
3.3.2. Malondialdehyde Content
3.3.3. Chlorophyll Content
3.3.4. Root Activity
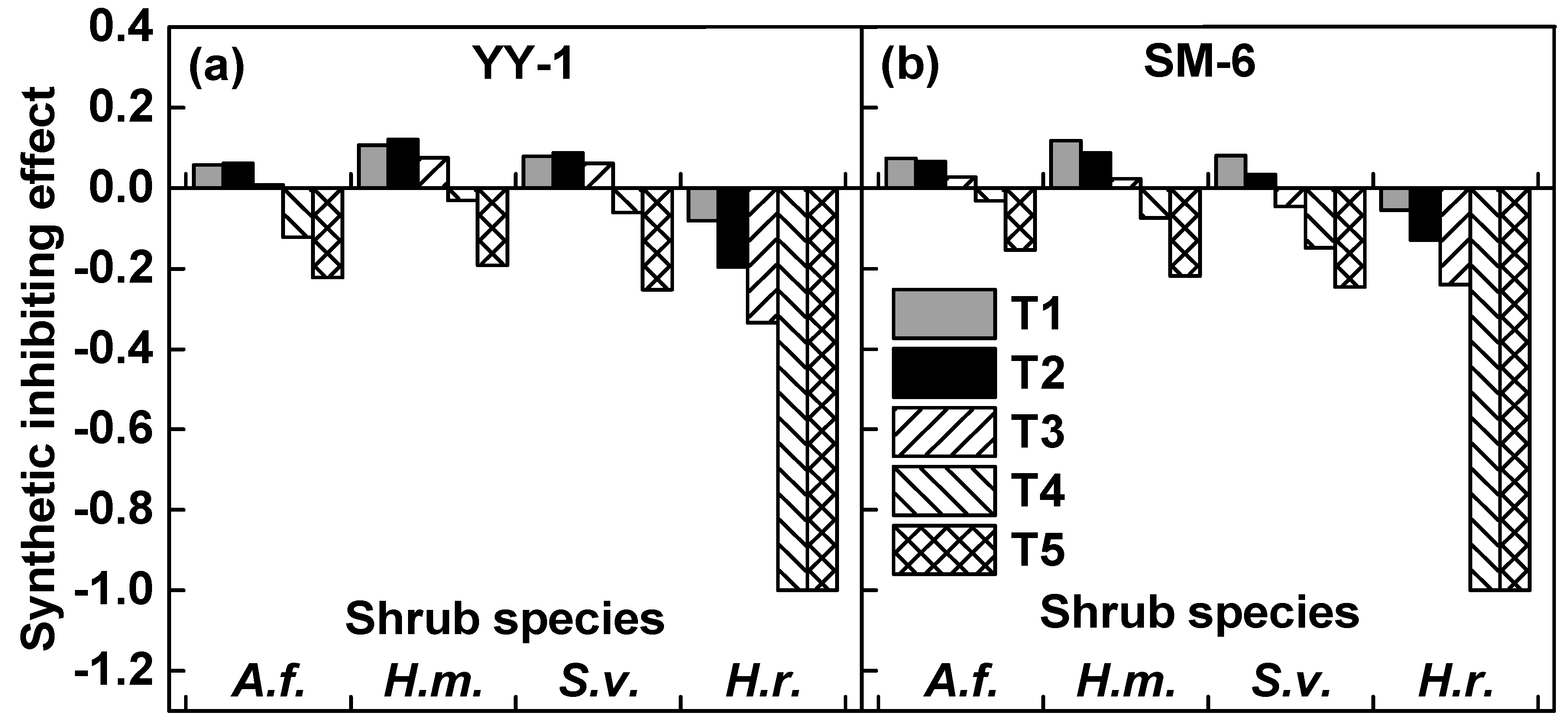
3.4. Synthetic Inhibiting Effect
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ekimova, N.V.; Muratova, E.N.; Silkin, P.P. The role of polyploidy in adaptation and settling of steppe shrubs in Central Asia. Russ. J. Genet. 2012, 2, 105–109. [Google Scholar] [CrossRef]
- Wang, J.; Ren, H.; Yang, L.; Li, D.; Guo, Q. Soil seed banks in four 22-year-old plantations in South China: Implications for restoration. For. Ecol. Manag. 2009, 258, 2000–2006. [Google Scholar] [CrossRef]
- Chu, J.M.; Yang, H.X.; Lu, Q.; Zhang, X.Y. Endemic shrubs in temperate arid and semiarid regions of northern China and their potentials for rangeland restoration. AoB Plants 2015, 7, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Schuler, L.J.; Bugmann, H.; Snell, R.S. From monocultures to mixed-species forests: Is tree diversity key for providing ecosystem services at the landscape scale? Landsc. Ecol. 2017, 32, 1499–1516. [Google Scholar] [CrossRef]
- Fornara, D.A.; Tilman, D. Plant functional composition influences rates of soil carbon and nitrogen accumulation. J. Ecol. 2008, 96, 314–322. [Google Scholar] [CrossRef] [Green Version]
- Kooch, Y.; Bayranvand, M. Composition of tree species can mediate spatial variability of C and N cycles in mixed beech forests. For. Ecol. Manag. 2017, 401, 55–64. [Google Scholar] [CrossRef]
- Nickmans, H.; Collet, C.; Bonal, D.; Verheyen, K.; Ponette, Q. Tree size and local neighborhood affect foliar nutrient content in a mixed plantation of beech (Fagus sylvatica) and maple (Acer pseudoplatanus). For. Ecol. Manag. 2017, 400, 159–172. [Google Scholar] [CrossRef]
- Lamb, E.G.; Kennedy, N.; Siciliano, S.D. Effects of plant species richness and evenness on soil microbial community diversity and function. Plant Soil 2011, 338, 483–495. [Google Scholar] [CrossRef]
- Mallick, N.; Mohn, F.H. Reactive oxygen species: Response of algal cells. J. Plant Physiol. 2000, 157, 183–193. [Google Scholar] [CrossRef]
- Sun, Y.F.; Zhang, Y.Q.; Feng, W.; Qin, S.G.; Liu, Z.; Bai, Y.X.; Yan, R.; Fa, K.Y. Effects of xeric shrubs on soil microbial communities in a desert in northern China. Plant Soil 2016, 414, 281–294. [Google Scholar] [CrossRef]
- Isbell, F.; Adler, P.R.; Eisenhauer, N.; Fornara, D.; Kimmel, K.; Kremen, C.; Letourneau, D.K.; Liebman, M.; Polley, H.W.; Quijas, S.; et al. Benefits of increasing plant diversity in sustainable agroecosystems. J. Ecol. 2017, 105, 871–879. [Google Scholar] [CrossRef] [Green Version]
- Ehbrecht, M.; Schall, P.; Ammer, C.; Seidel, D. Quantifying stand structural complexity and its relationship with forest management, tree species diversity and microclimate. Agric. For. Meteorol. 2017, 242, 1–9. [Google Scholar] [CrossRef]
- Sezen, I.; Ercisli, S.; Cakir, O.; Koc, A.; Temi, E.; Hadziabulic, A. Biodiversity and landscape use of sea buckthorn (Hippophae rhamnoides L.) in the Coruh Valley of Turkey. Erwerbs-Obstbau 2015, 57, 23–28. [Google Scholar] [CrossRef]
- Lai, Z.R.; Zhang, Y.Q.; Liu, J.B.; Wu, B.; Qin, S.G.; Fa, K.Y. Fine-root distribution, production, decomposition, and effect on soil organic carbon of three revegetation shrub species in northwest China. For. Ecol. Manag. 2016, 359, 381–388. [Google Scholar] [CrossRef]
- Wei, Q.; Wei, Y.; Wu, H.W.; Yang, X.Y.; Chen, H.L.; Zhang, H.X. Chemical composition, anti-oxidant, and antimicrobial activities of four saline-tolerant plant seed oils extracted by SFC. J. Am. Oil Chem. Soc. 2016, 93, 1173–1182. [Google Scholar] [CrossRef]
- Tanaka-Oda, A.; Kenzo, T.; Kashimura, S.; Ninomiya, I.; Wang, L.H.; Yoshikawa, K.; Fukuda, K. Physiological and morphological differences in the heterophylly of Sabina vulgaris Ant. in the semi-arid environment of Mu Us Desert, Inner Mongolia, China. J. Arid Environ. 2010, 74, 43–48. [Google Scholar] [CrossRef]
- Requena, N.; Perez-Solis, E.; Azcón-Aguilar, C.; Jeffries, P.; Barea, J.M. Management of indigenous plant-microbe symbioses aids restoration of desertified ecosystems. Appl. Environ. Microbiol. 2001, 67, 495–498. [Google Scholar] [CrossRef] [PubMed]
- Ishii, Y.; Sakamoto, K.; Yamanaka, N.; Wang, L.; Yoshikawa, K. Light acclimation of needle pigment composition in Sabina vulgaris seedlings under nurse plant canopy. J. Arid Environ. 2006, 67, 403–415. [Google Scholar] [CrossRef]
- Rice, E.L. Allelopathy, 2nd ed.; Academic Press: New York, NY, USA, 1984; pp. 1–50, 309–315. [Google Scholar]
- Ninkovic, V. Volatile interaction between undamaged plants: A short cut to coexistence. In Plant Communication from an Ecological Perspective; Springer: Heidelberg, Germany, 2010; pp. 75–86. [Google Scholar]
- Glinwood, R.; Ninkovic, V.; Pettersson, J. Chemical interaction between undamaged plants—Effects on herbivores and natural enemies. Phytochemistry 2011, 72, 1683–1689. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Li, X.; Chen, Y.; Zhao, J.; Wan, S.; Lin, Y.; Fu, S. Effects of Eucalyptus litter and roots on the establishment of native tree species in Eucalyptus plantations in South China. For. Ecol. Manag. 2016, 375, 76–83. [Google Scholar] [CrossRef]
- Peerzada, A.M.; Ali, H.H.; Chauhan, B.S. Weed management in sorghum (Sorghum bicolor (L.) Moench) using crop competition: A review. Crop Prot. 2017, 95, 74–80. [Google Scholar] [CrossRef]
- Siri-Udom, S.; Suwannarach, N.; Lumyong, S. Applications of volatile compounds acquired from Muscodor heveae against white root rot disease in rubber trees (Hevea brasiliensis Müll. Arg.) and relevant allelopathy effects. Fungal Biol. 2017, 121, 573–581. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.X.; Liu, Z.W.; Tian, N.; Luc, N.T.; Zhu, B.C.; Bing, Y.H. Allelopathic effects of decomposed leaf litter from intercropped trees on rape. Turk. J. Agric. For. 2015, 39, 898–908. [Google Scholar] [CrossRef] [Green Version]
- Yan, S.; Zou, G.H.; Li, S.J.; Wang, H.; Liu, H.Q.; Zhai, G.W.; Guo, P.; Song, H.M.; Yan, C.J.; Tao, Y.Z. Seed size is determined by the combinations of the genes controlling different seed characteristics in rice. Theor. Appl. Genet. 2011, 123, 1173–1181. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Lü, B.; Xiao, Z.W.; Yan, J.Z.; Zhang, Y.F. Seed characteristics and hard seed broken methods of Sophora alopecuroides and Thermopsis lanceolata (in Chinese with English abstract). Trans. Chin. Soc. Agric. Eng. 2010, 26, 406–410. [Google Scholar]
- Le Rouzic, B.; Thiébaut, G.; Brient, L. Selective growth inhibition of cyanobacteria species (Planktothrix agardhii) by a riparian tree leaf extracts. Ecol. Eng. 2016, 97, 74–78. [Google Scholar] [CrossRef]
- Wang, C.Y.; Xiao, H.G.; Zhao, L.L.; Liu, J.; Wang, L.; Zhang, F.; Shi, Y.C.; Du, D.L. The allelopathic effects of invasive plant Solidago canadensis on seed germination and growth of Lactuca sativa enhanced by different types of acid deposition. Ecotoxicology 2016, 25, 555–562. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Cai, J.; Jiang, Z.M.; Zhang, S.X. Allelopathic effects of walnut leaves leachate on seed germination, seedling growth of medicinal plants. Allelopath. J. 2010, 26, 235–242. [Google Scholar]
- Wang, C.Z.; Zhang, D.J.; Zang, J.; Ji, T.W.; Tang, Z.Q.; Zhao, Y.Y. Allelopathic effects of volatile compounds from Eucalyptus grandis on Vigna radiata, Raphanus sativus and Lactuca sativa. Allelopath. J. 2015, 36, 273–282. [Google Scholar]
- Fu, J.M.; Huang, B.R. Involvement of antioxidants and lipid peroxidation in the adaptation of two cool-season grasses to localized drought stress. Environ. Exp. Bot. 2001, 45, 105–114. [Google Scholar] [CrossRef]
- Hodges, D.M.; DeLong, J.M.; Forney, C.F.; Prange, R.K. Improving the thiobarbituric acid-reactive-substances assay for estimating lipid peroxidation in plant tissues containing anthocyanin and other interfering compounds. Planta 1999, 207, 604–611. [Google Scholar] [CrossRef]
- Arnon, D.I. Copper enzymes in isolated chloroplast polyphenol oxidase in Beta vulgaris. Plant Physiol. 1949, 24, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Williamson, G.B.; Richardson, D. Bioassays for allelopathy: Measuring treatment responses with independent controls. J. Chem. Ecol. 1988, 14, 181–187. [Google Scholar] [CrossRef] [PubMed]
- Arowosegbe, S.; Afolayan, A.J. Germination and growth inhibitory effects of aqueous extracts of different parts of Bidens pilosa L. var radiata on some vegetable crops. Bull. Pure Appl. Sci. 2013, 32, 11–20. [Google Scholar]
- Mahmoud, A.; Singh, S.D.; Muralikrishna, K.S. Allelopathy in jatropha plantation: Effects on seed germination, growth and yield of wheat in north-west India. Agric. Ecosyst. Environ. 2016, 231, 240–245. [Google Scholar] [CrossRef]
- Aliskan, I.K.; Terzi, I. Allelopathic effects of walnut leaf extracts and juglone on seed germination and seedling growth. J. Hortic. Sci. Biotechnol. 2001, 76, 436–440. [Google Scholar] [CrossRef]
- Thomaszewishi, M.; Thimann, K.V. Interaction of phenolic acids, metallic Ions and chelating agents on auxin-induced growth. Plant Physiol. 1966, 41, 1443–1454. [Google Scholar] [CrossRef]
- Yu, J.Q.; Matsui, Y. Effects of root exudates of cucumber (Cucumis sativus) and allelochemicals on Ion uptake by cucumber seedlings. J. Chem. Ecol. 1997, 23, 817–827. [Google Scholar] [CrossRef]
- Zeng, R.S.; Luo, S.M.; Shi, Y.H.; Shi, M.B.; Tu, C.Y. Physiological and biochemical mechanism of allelopathy of Secalonic acid F on higher plants. Agron. J. 2001, 93, 72–79. [Google Scholar] [CrossRef]
- Roshchina, V.V.; Roshchina, V.D. The Excretory Function Higher Plants; Springer: New York, NY, USA, 1993; pp. 213–215. [Google Scholar]
- Huang, Y.; Bai, Y.; Wang, Y.; Kong, H. Allelopathic effects of the extracts from an invasive species Solidago canadensis L. on Microcystis aeruginosa. Lett. Appl. Microbiol. 2013, 57, 451–458. [Google Scholar] [CrossRef] [PubMed]
- Klaudia, B. Morphological changes in two tobacco and petunia cultivars under exposure to tropospheric ozone. Acta Biol. Crac. Ser. Bot. 2013, 55, 1–9. [Google Scholar] [CrossRef]
- Mallik, A.U.; Biswas, S.R.; Collier, L.C.S. Belowground interactions between Kalmia angustifolia and Picea mariana: Roles of competition, root exudates and ectomycorrhizal association. Plant Soil 2016, 403, 471–483. [Google Scholar] [CrossRef]
- Maxwell, K.; Johnson, G.N. Chlorophyll fluorescence-a practical guide. J. Exp. Bot. 2000, 345, 659–668. [Google Scholar] [CrossRef]
- Tommasion, E.; Griffa, S.; Grunberg, K.A.; Ribotta, A.N.; Colomba, E.L.; Carloni, E.; Quiroga, M.P.; Luna, C. Malondialdehyde content as a potential biochemical indicator of tolerant Cenchrus ciliaris L. genotypes under heat stress treatment. Grass Forage Sci. 2012, 67, 456–459. [Google Scholar] [CrossRef]
- Einhellig, F.A. Mechanism of Action of Allelochemicals in Allelopathy; ACS Publications: Washington, DC, USA, 1995; Volume 582, pp. 96–116. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Habitat | Population Code | Longitude (E) | Latitude (N) | Altitude (m) | AMP 1 (mm) | AMT 2 (°C) |
|---|---|---|---|---|---|---|
| Yuyang County, Shaanxi Province | YY | 108°58′–110°24′ | 37°49′–38°5′ | 1200–1300 | 406.9 | 8.1 |
| Shenmu County, Shaanxi Province | SM | 107°20′–111°30′ | 37°27.5′–39°22.5′ | 1200–1300 | 250–440.8 | 8.5 |
| Species | Thousand-Seed Weight (g) | Thousand-Kernel Weight (g) | Single-Seed Volume (mm3) | Single-Kernel Volume (mm3) | Kernel Rate (%) |
|---|---|---|---|---|---|
| YY-1 | 746.27 ± 0.35 a | 183.76 ± 1.47 a | 1532.09 ± 0.16 a | 371.34 ± 0.10 a | 24.62 ± 0.10 b |
| SM-6 | 448.46 ± 0.97 b | 121.08 ± 0.44 b | 984.99 ± 3.22 b | 250.01 ± 2.04 b | 27.00 ± 0.04 a |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, X.; Wang, J.; Zhang, R.; Huang, Y.; Feng, S.; Ma, X.; Zhang, Y.; Sikdar, A.; Roy, R. Allelopathic Effects of Aqueous Leaf Extracts from Four Shrub Species on Seed Germination and Initial Growth of Amygdalus pedunculata Pall. Forests 2018, 9, 711. https://0-doi-org.brum.beds.ac.uk/10.3390/f9110711
Wang X, Wang J, Zhang R, Huang Y, Feng S, Ma X, Zhang Y, Sikdar A, Roy R. Allelopathic Effects of Aqueous Leaf Extracts from Four Shrub Species on Seed Germination and Initial Growth of Amygdalus pedunculata Pall. Forests. 2018; 9(11):711. https://0-doi-org.brum.beds.ac.uk/10.3390/f9110711
Chicago/Turabian StyleWang, Xiuqing, Jinxin Wang, Ruiqi Zhang, You Huang, Shulin Feng, Xu Ma, Yuyu Zhang, Ashim Sikdar, and Rana Roy. 2018. "Allelopathic Effects of Aqueous Leaf Extracts from Four Shrub Species on Seed Germination and Initial Growth of Amygdalus pedunculata Pall." Forests 9, no. 11: 711. https://0-doi-org.brum.beds.ac.uk/10.3390/f9110711