Changes Occurring in Spontaneous Maize Fermentation: An Overview
by
 and
and
Clemencia Chaves-López
*, 
Chiara Rossi
,
Francesca Maggio
, 
Antonello Paparella
and
Annalisa Serio
*
Faculty of Bioscience and Technology for Food, Agriculture and Environment, University of Teramo, Via R. Balzarini 1, 64100 Teramo, TE, Italy
*
Authors to whom correspondence should be addressed.
Fermentation 2020, 6(1), 36; https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation6010036
Submission received: 26 February 2020
/
Revised: 19 March 2020
/
Accepted: 21 March 2020
/
Published: 23 March 2020
(This article belongs to the Special Issue Fermentation and Bioactive Metabolites 2.0)
Abstract
:Maize and its derived fermented products, as with other cereals, are fundamental for human nutrition in many countries of the world. Mixed cultures, principally constituted by lactic acid bacteria (LAB) and yeasts, are responsible for maize fermentation, thus increasing its nutritional value and extending the products’ shelf-life. Other microorganisms involved, such as molds, acetic acid bacteria, and Bacillus spp. can contribute to the final product characteristics. This review gives an overview of the impact of the activities of this complex microbiota on maize product development and attributes. In particular, starting from amylolytic activity, which is able to increase sugar availability and influence the microbial succession and production of exopolysaccharides, vitamins, and antimicrobial compounds, which improve the nutritional value. Further activities are also considered with positive effects on the safety profile, such as phytates detoxification and mycotoxins reduction.
1. Introduction
Maize or corn (Zea mays) is a graminaceous annual plant whose origin is linked to America. It was introduced into Europe in the sixteenth century, then spread outside the continent, across Africa and Far East Asia, and due to its exceptional geographic adaptability, nowadays, it is considered as one of the most important cereals in the world. Maize is a good source of metabolizable energy, and in spite of the poor protein content, it is also considered as a vital food grain in many countries, and in particular, in Africa, Asia, Central, and Southern America.
In recent years, the number of reports describing the fermentation process around the world, as well as the microorganisms involved in maize-fermented products, particularly from Africa and Latin America, has significantly increased. Table 1 reports the most common maize-based fermented food products and their principal technological characteristics, such as fermentation time and temperature, pH, and ethanol content.
Fermentation is recognized as a natural way to preserve and safeguard foods and beverages, enhancing the nutritional value, improving the digestibility, destroying undesirable components, and inhibiting undesirable microorganisms [66]. However, in artisanal fermented products, biological risks such as pathogenic microorganisms, as well as chemical contaminants and toxic molecules of microbial origin, including mycotoxins, biogenic amines, and cyanogenic glycosides can be found [67]. For this reason, a deep understanding of the role of the different microbial groups developing in spontaneously fermented products is of crucial importance to optimize the final quality and to improve the food safety of these products.
In a fermented product, the metabolic activity reflects the metabolic capabilities of the different species or microbial groups that, together with the technological characteristics of the process, influence the sensorial properties of spontaneously fermented products [9]. In fact, microbial interactions in mixed cultures, taking place in fermented products, occur via multiple mechanisms, and the effects of such interactions on the fitness of the strains involved may be either positive, neutral, or negative [68]. Taking into account that any spatial or temporal change in the community composition can consequently modify this complex ecosystem, then the role of the different species on flavor, rheology, and shelf-life, as well as on the functional/nutritional characteristics, has been a matter of study. In the light of these considerations, in this review, we will focus on important activities that occur during maize fermentation, with particular emphasis on those determining the microbial succession and the development of fermentation, such as the increased sugar availability by the degradation of starch, and the production of exopolysaccharides, vitamins, and antimicrobial compounds. In addition, we will also consider the microbial action that also provides detoxification of phytates and reduction of mycotoxins.
2. Maize Fermentation
Around the world, maize grains are processed and fermented following different traditions to obtain a great diversity of products. In general, fermented maize production initially involves the cleaning of the grains, which are then successively soaked in water until soft and ground when wet, followed by fermentation. Domínguez-Ramírez et al. [69] classify the methods used to prepare fermented maize products into 4 categories according to the methods in which foods are made with: 1) dried kernel, 2) complete maize ears soaked, 3) maize flour and 4) mashed tender maize. Moreover, on the basis of the texture, fermented maize products can be classified into liquid (gruel and porridges), such as ogi, dalaki; chicha, champus, and boza; solid (dough and dumplings), such as masa agria, pozol, kenkey, akidi, and komé; and dry (baked, fried, and steam-cooked granulated products), such as arraw, dégué, masa, and wômi [70].
Before maize fermentation, some traditions provided the use of a pretreatment such as nixtamalization, a process where the maize grains are soaked with an alkali (generally lime), then cooked, dried, and ground to obtain the flour; grain germination, and chewing. These actions lead to physical and chemical changes in the grains, thereby acting as selective agents for the microbiota that guides the fermentation process in this modified substrate. Microorganisms utilize a large number of nutrients present in the grains, and their metabolism is a major driving force in the regulation of microbial diversity and activity on the fermented maize. The fermentation processes modify the grains through various steps, in which endogenous enzymes (amylases, proteases, phytases, etc.) and microbial enzymes (usually from lactic acid bacteria and yeasts) are involved [71]. The microbial activity in maize dough or slurry is a well-defined temporal succession of naturally occurring microorganisms that are usually found in association with each type of fermentation.
In uninoculated fermented products, microorganisms principally derived from the raw materials can also be affected by the phytochemical treatments of maize or by the environmental features, such as temperature, rainfall, or insect attacks during cultivation [36]. In addition, the microbial diversity of this kind of fermented product can generally be contributed to by the water employed during the production, as well as the tools (e.g., spoons, pots, etc.), the contact with wooden tables, or the exposition to the air, and particularly the step of grinding and soaking [55]. The recontamination, after any cooking or hot water treatments, depends mainly on household conditions (e.g., air, storage containers, spoons, etc.) [72]. In some cases, microbial propagation is guaranteed by the addition of a portion of fermented material from a previous batch (back-slopping).
Environmental parameters such as temperature, pH, inoculum quantity (where applicable), and the fermentation time defines the members of the maize microbiota, which are part of a complex consortium. In addition, the maize pretreatment before fermentation also contributes to the prevalence of some species; in this regard, a greater biodiversity of species and in particular of Candida spp. were detected in chicha, a Colombian fermented beverage produced with maize pretreated with chewing, compared to those without any treatment [10].
Therefore, as mentioned previously, the mixture of microorganisms that carries out the fermentation leads to a product with very variable quality and sensory characteristics. On the other hand, the geographical isolation among the different fermented maize products provides significantly different microbial communities so that each maize fermented product can be considered as unique [36].
During the fermentation, a given microorganism, or groups of them, initiates the growth and becomes established during a specific period of time; afterward, the growth decreases due to the accumulation of toxic end-products or other inhibitory factors. In this way, the microorganisms provide the appropriate environment to other species less sensitive to those inhibitory factors. Culture-independent approaches have shown that microbial diversity in maize fermented product microbiomes is highly underestimated [11,16,36,53,73]. However, as evidenced in Table 2 and Table 3, the coexistence of lactic acid bacteria (LAB) and yeasts in fermented maize products is unavoidable. In addition, the presence of fungi, acetic acid bacteria (AAB), and Bacillus species is frequent in several products. Concerning the pathogenic bacteria, very few studies reported the presence of Escherichia coli and Enterobacter aerogenes.
Concerning the most common microorganisms found in fermented maize products, it is well established that LAB frequently produce enzymes able to breakdown polysaccharides or other molecules with high molecular weight, as well as organic acids and some compounds able to kill or reduce the microbial populations, such as bacteriocins and hydrogen peroxide [77]. They are also able to increase the content of free amino acids and B group vitamins, improving the availability of iron, zinc, and calcium by breaking down antinutritional compounds [31]; in addition, they produce gas and other volatile compounds (VOCs) contributing to the sensory properties of the product.
Yeasts, besides providing growth factors such as vitamins and soluble nitrogen to LAB, also produce several extracellular enzymes (lipases, esterases, amylases, and phytases), some of which participate in the formation of fermented maize flavor and aroma [45]. Yeasts produce a wide variety of VOCs, such as alcohols, esters, aldehydes, and ketones, that enrich the sensory characteristics of the maize fermented product and also contribute to reducing mold growth and spore germination, as in the case of ethyl acetate [78]. Recently, studies of Ponomarova et al. [79], combining metabolomics and genetics, evidenced that yeasts enable the growth of LAB through endogenous, multi-component cross-feeding in a readily established community.
On the other hand, the aerobic spore-forming bacteria (Bacillus spp.) secrete a wide range of degradative enzymes, such as amylases and proteases [80], and can also produce antimicrobial compounds such as bacilysin, which is able to inhibit molds and bacteria; and iturin and chloromethane, which inhibit bacteria [59], thus playing an important role in the fermented maize product development.
During maize fermentation, as occurring in other uninoculated fermentations, the competition among species for substrates, acid tolerance, syntrophic interactions, and other physiological properties of microbial populations causes fast variations in the microbiota structure. However, the microbial consortium of dough and beverages from fermented maize is stable, and mutually beneficial interactions among different species can contribute to the coexistence of some of them [81]. For example, this is the case for Lactobacillus plantarum, Lactobacillus fermentum, and Saccharomyces cerevisiae in ogi [82], Lb. plantarum and Acetobacter fabarum in masa agria [36], Pediococcus pentosaceus and Weissella confusa in atole agrio [4] or Lb. fermentum and Lactobacillus delbrueckii subsp. bulgaricus in akamu [2]. Microbiological studies have revealed that during spontaneous fermentation, bacteria and yeasts secrete a diverse array of metabolites that are available for all the community members. Thus, the interactions of the different microorganisms play a significant role during maize fermentation and participate in the changes of the nutritional, rheological, and sensorial treats through modification of the maize composition.
Figure 1 describes the main effect of the metabolic processes of the principal microbial groups involved in maize fermentation, improving their mutual interactions, while Figure 2 depicts the effect of these microbial activities on the characteristics of the final fermented maize. All these activities are detailed in the next paragraphs.
3. Production of Amylolytic Enzymes
Mature maize kernels contain low levels of free sugars, mainly generated by endogenous grain amylases; these sugars support the growth of microorganisms like LAB, which can begin the fermentation process [31]. It is well established that at the beginning of maize fermentation, only a few microorganisms are able to use the starch, and for this reason, the microbial biodiversity is lower than in the second stage. The action of microbial amylases releases other carbon sources (i.e., dextrins and maltose) accessible for a greater number of species, including nonamylolytic strains. Moreover, the organic products formed during fermentation (lactic acid, formic acid, and ethanol) may also serve as carbon sources for microorganisms such as yeasts [56]. Thus, the amylolytic activity during maize fermentation is fundamental because it improves the energy sources for the nonamylolytic microorganisms and plays an important role in the fast reduction of pH values.
The amylolytic activity in LAB species is not a very frequent feature; only a few species exhibit this activity by converting the starch directly into lactic acid in a single step [83]. For this bioconversion, a fundamental role is played by the gene amyA, which encodes for an extracellular α-amylase that is not expressed continuously but transiently [84]. In addition, this amylolytic activity is strain-dependent [5] and can be inhibited by the pH reduction due to the lactobacilli growth [85]. In this context, some prefermentation processes (Figure 3) could contribute to selecting bacteria with high amylolytic activity [58]. According to Petrova et al. [86], bacteria of Lactobacillus, Lactococcus, and Streptococcus genera are able to directly metabolize starch for the production of lactic acid, with lactobacilli (e.g., Lactobacillus amylovorus) as the most efficient.
Amylolytic LAB (ALAB) may be key organisms during the production of the maize fermented food, and this is the case of Lb. fermentum in Mexican pozol [56]. In an integrated vision of how the low amounts of fermenting sugars present in maize can determine microbial diversity and support a high number of lactic acid bacteria in pozol, Diaz-Ruiz et al. [58] pointed out that the presence of ALAB in the first stage of fermentation played a fundamental role in this food ecosystem, although their amylase level was low. The authors suggested that the amylolytic activity of Streptococcus infantarius, which has been established as a predominant LAB during pozol’s fermentation, together with Streptococcus bovis, could provide low-molecular-weight malto-oligosaccharides to the not amylolytic microorganisms during the initial steps of nixtamal dough fermentation. In the same way, Lb. plantarum (CPQBA 087–11 DRM) isolated from Colombian masa agria showed high amylolytic activity [87]. The ALAB amylolytic activity is mainly due to the production of extracellular amylases. According to genomic studies, ALAB produce alpha-amylases, maltogenic amylases, amylopullulanases, pullulanases, neopullulanases, and 6-glucosidases [86].
Starch hydrolysis is performed mainly by bacteria rather than by yeasts [88], however, the amylolytic activity was also reported in yeasts as a strain-dependent feature; in particular, Saccharomyces cerevisiae (2/77) and Candida krusei (2/27), grown in a medium containing amylopectin incorporated with 2% soluble starch, showed high activity [45]. Moreover, Candida famata, C. krusei, and S. cerevisiae isolated from fermented maize in Indonesia and Africa showed amylolytic activity [45,89]. Chicha from the Andes is a source of yeasts with high amylolytic activity, with Cryptococcus flavescens, Cryptococcus magnus, Cryptococcus carnescens, Pichia membranifaciens, Cryptococcus spp, Rhodotorula mucilaginosa, and Wickerhamomyces anomalus as the most potent producers [12,88].
Bacillus spp. bacteria, commonly isolated from fermented maize products, are beneficial as well for the fermentation process due to the production of amylase [90], and particularly of α-amylases, which convert starch to glucose for microorganisms lacking in these enzymes, such as S. cerevisiae [91].
The amylolytic activity of bacteria and yeasts is not only beneficial for microbiota growth, but it can also have an effect on the rheology of the product. Acid and amylase enzymes easily attack the amorphous regions of the starch granules, reducing the molecular mass of amylose and amylopectin [92]. This suggests that the enzymes produced by LAB during the maize fermentation have an effect on the glycosidic bonds in the starch granule, hydrolyzing them and enabling the granules to absorb water faster, thus reducing the viscosity of the fermented slurry and the cohesive structure of the doughs. Moreover, while the viscosity of bulk and starchy weaning gruel is decreased, the nutrient density is increased, thus maintaining an acceptable thickness for feeding young children [93].
4. Production of Exopolysaccharides (EPS)
The exopolysaccharides (EPS) are microbial biopolymers secreted into the extracellular environment in the form of capsules or biofilm. These compounds protect the cell against several environmental stresses occurring during fermentation [94], and this encompasses a wide transcriptional response with many induced or repressed genes [95]. Therefore, the particular condition encountered during maize fermentation could favor the production of EPS and could affect the way in which microorganisms interact with the external environment, whether it is liquid or solid [94].
Several factors influence the formation and the features of the different EPS, such as types of monosaccharides, type of linkages, degree of branching, and molecular weight. From the technological point of view, EPS formed from sucrose by glycansucrase activity during sourdough fermentation influence the viscoelastic properties of the dough and beneficially affects the rheological characteristics and shelf-life (in particular starch retrogradation) of the product [96]. In this context, Falade et al. [97], who investigated the impact of fermentation on the maize dough, found a higher elastic modulus than the viscous modulus and suggested that the doughs are viscoelastic solids, exhibiting more elastic properties than viscous ones. Although the authors attributed the cohesive dough structure in sourdough bread in part to endosperm matrix protein degradation, the contribution of EPS to the increase of the viscoelastic nature of the dough should not be underestimated [98]. Moreover, probably the viscous nature of some uncooked products such as ogi and fufu slurries could be partly due to the excretion of exopolysaccharides by the dominant LAB. In addition, EPS confer beneficial physiological effects on human health, such as antitumor activity and immunomodulating bioactivity [99].
EPS-producing LAB belongs to different genera such as Enterococcus, Lactobacillus, Lactococcus, Leuconostoc, Pediococcus, Streptococcus, and Weissella, but also EPS-producers Bifidobacterium, Acetobacter spp., and Bacillus spp. have been reported [100,101,102,103]. As reported in Table 2 and Table 3, several species of these genera are commonly associated with maize products, and in recent years, the isolation of EPS-producing LAB from spontaneously fermented maize products has gained attention. Some studies suggested that the amount of EPS produced is strain- and species-dependent [100]. On the other hand, Donot et al. [94] suggested that the physiological role of EPS depends on the biotope of the microorganisms producing them.
The roles of EPS produced by LAB have been described by several researchers, nevertheless, the majority of these papers were achieved in lab conditions and using strains growing in sucrose-containing agar culture. In this context, among 88 strains isolated from atole agrio, Lb. plantarum and P. pentosaceus were the major EPS producers [5]. In the same way, in a study on 70 LAB strains isolated from ogi, only a few of them, belonging to the species Lactococcus brevis, Lactococcus mesenteroides, Lactococcus lactis, Lb. fermentum, Lactococcus lactis, and Lb. plantarum, showed potential to produce EPS, with values ranging between 120 and 1,390 mg ml-1. EPS production is a common treat of Lb. plantarum and Lactococcus rhamnosus isolated from Kenyan ikii, with quantities ranging from 298.53 mg l-1 to 431 mg l-1 [21].
Although the exact role of the EPS produced during fermentation on the fermented maize ecosystem is not well elucidated, the presence of EPS in dough could contribute to retaining humidity, delaying water movement through the dough toward the peripheral area, thus reducing the water loss by evaporation. For this reason, the EPS producers could strongly promote the stabilization of dough microbiota.
5. Vitamins and Amino Acids Increase
Maize is often deficient in vitamins (having a very low concentration of vitamins A and B12), and amino acids (lacking arginine and methionine). Some pretreatments of the maize grains, such as germination, contribute to increasing their nutritional value, particularly in terms of peptides, amino acids, vitamins (B1 and E), gamma-aminobutyric acid (GABA), and total phenolic content [104]. Even during fermentation, an increase of some of these compounds occurs. It is generally believed that the yeasts excrete nutrients of which the LAB benefit, such as pyruvate, amino acids, and vitamins. Although LAB are auxotrophic for different growth factors, some strains are able to produce B group vitamins, such as cobalamin (B12), folate (B11), and riboflavin (B2). In particular, some Lb. plantarum, Lactococcus rossiae, Lb. fermentum, Lactococcus buchneri, Lactococcus hilgardii, and Lb. brevis strains were demonstrated to be vitamin B12 producers [105,106,107], while Lactococcus pentosus, Lb. plantarum, and Lactococcus acidophylus were riboflavin producers. Uninoculated maize fermentation generally increases the levels of nutritional compounds such as thiamine (vitamins B1), folate, riboflavin, total carotenoids, vitamin C, and Vitamin E [27,108,109,110]. However, the various steps involved in the process of ogi in Cameroon contribute to reducing thiamine (69%), riboflavin (82%), and β-carotene (66%) in maize fermented according to traditional preparation [111]. On the other hand, the increase in folate content has been reported in many fermented products [112]; in fact, it is well established that some indigenous yeasts species such as S. cerevisiae, Candida milleri, Torulaspora delbruekii, Issatchenkia orientalis, Pichia anomala, Kluyveromyces marxianus, and Candida glabrata produce considerable amounts of folate during fermentation. In particular, in togwa, I. orientalis, P. anomala, S. cerevisiae, K. marxianus, and C. glabrata were able to increase folate concentration after 46 h of fermentation, with C. glabrata as the highest producer (23-fold more) compared to unfermented samples [65]. The same authors also highlighted that folate production is highly culture- and species-dependent, being greater during the exponential phase. On the contrary, folate production by LAB is lower, and some of these bacteria deplete folate during fermentation [113]. The increase of methionine, tryptophan, and folate content in fermented maize has also been attributed to the activity of non-LAB-bacteria such as Bacillus licheniformis and Enterobacter cloacae [114]; again, the ability to synthesize folate may reflect strain differences.
The combined processes of germination and fermentation of maize using Lactobacillus plantarum, Lactococcus lactis, Bacillus subtilis, and Bifidobacterium longum increased the content of GABA five-fold. This amino acid could be produced by glutamate decarboxylation performed by Lactobacillus spp., and it plays a role in regulating neuronal excitability throughout the nervous system of mammals, inducing hypotension, and exerting diuretic and tranquilizer effects [115].
6. Production of Antimicrobial Compounds
As already described, the fermentation occurring in maize products depends on a consortium of several genera and species. Usually, one or different species start to proliferate and settle down during a specific period of time. Successively, the decrease or even cessation of growth as a consequence of the increase of toxic end products or other inhibitory factors paves the way to other species less sensitive to those inhibitory factors. Microbial metabolism during maize fermentation may lead to a series of compounds capable of inhibiting a considerable spectrum of bacteria and fungi. In particular, the production of lactic and acetic acids as end products contributes to reducing the pH, creating a hostile environment for the growth of many microorganisms. While lactic acid is produced mainly by LAB, acetic acid is produced principally by acetic acid bacteria (AAB) and specifically by Acetobacter spp., when they find excess oxygen [31]. The organic acids disrupt the mechanisms responsible for maintaining the membrane potential, thus inhibiting the active transport across the membrane. In this way, many foodborne pathogens could be inhibited during the fermentative process. In addition, other antimicrobial compounds are recognized to be produced during fermentation, including propionic acid, ethanol generated by yeast, and by LAB via the heterofermentative pathway, H2O2 produced during the aerobic growth of LAB, and diacetyl, formed from an excess of citrate-derived pyruvate. Moreover, selected bacteria isolated from fermented maize products display the ability to produce bacteriocins, which are ribosomally synthesized proteinaceous compounds exerting selective antimicrobial activity.
The antimicrobial potential of LAB isolated from fermented maize dough has been studied by Olsen et al. [116], who examined the bacterial interactions during fermentation. They observed a widespread occurrence of antimicrobial compounds, effective against Gram-positive and Gram-negative bacteria. In particular, strains of Lb. plantarum, and Lb. fermentum/reuteri showed antagonism correlated with the synergic effect of acids, as well as with the production of compounds sensitive to proteolytic enzymes, therefore probably bacteriocins.
In general, in fermented maize, LAB are regularly associated members of the genera Lactobacillus, Pediococcus, Leuconostoc, Enterococcus, Lactococcus, and Weissella, and many species have been recognized as bacteriocin-producers. In one of the first studies in this research field, Olasupo et al. [117] observed a reduction of about 4 log UFC mL−1 of E. coli during ogi fermentation, when a bacteriocin-producing Lactobacillus was inoculated. Also Lb. plantarum E2, isolated from chicha de jora (an alcoholic beverage from Peru), produced a bacteriocin limiting the growth of Lb. fermentum Chj4C, another strain isolated from the native beverage [118]. Moreover, 28 strains of Lb. plantarum and 3 Lb. fermentum isolated from poto poto, produced the bacteriocin plantarincin, able to reduce the population of Escherichia coli, Salmonella enterica, Enterobacter aerogenes, Bacillus cereus, Staphylococcus aureus, Listeria monocytogenes, and Enterococcus faecalis [54]. In the same way, one Lb. rhamnosus and several Lb. plantarum strains isolated from sha’a, a typical maize fermented beverage from Cameroon, produced bacteriocins able to inhibit both Gram-positive and Gram-negative bacteria, including species of the genera Lactobacillus, Streptococcus, Bacillus, Staphylococcus, Salmonella, Shigella, Pseudomonas, Klebsiella, and Escherichia, also including multidrug resistant strains of the pathogens E. coli and S. aureus [119]. Furthermore, Lb. lactis, Lb. fermentum, Lb. casei and Lb. plantarum isolated from ogi, showed different antimicrobial potential, particularly against Salmonella Typhimurium and Shigella dysenteriae [120]. The potential of Lb. plantarum ULAG24 isolated from ogi in Nigeria to release bacteriocins during maize fermentation was exploited to inhibit Salmonella in the spot-on-lawn experiment. In this case, the LAB strain expressed all nine genes associated with plantaricin biosynthesis [121]. The inhibition of E. coli and S. aureus was observed using a bacteriocin-producing Lactobacillus isolated from akamu [122].
The high occurrence of bacteriocin-producing Lb. fermentum strains, isolated in different stages of doklu production, were reported by Assohoun-Djeni et al. [15]. The authors also observed the capability of 16 strains belonging to various species and mainly Lb. fermentum, Lb. plantarum, Pediococcus acidilactici, P. pentosaceus, and Weissella cibaria to produce antifungal compounds that inhibited the growth of Eurotium repens, Penicillium corylophilum, Aspergillus niger, Wallemia sebi, and Cladosporium sphaerospermum. Lactic and acetic acids are the most recognized antifungal molecules produced by LAB, however, other acids such as formic, propionic, butyric, phenyllactic, hydroxyphenyllactic, and indole-3-lactic, in addition to peptides of low molecular weight [123,124,125] are produced. All of them are supposed to have the fungal cell wall as a target.
The antibacterial activity of Bacillus spp. CS93 isolated from pozol has been demonstrated [59,126]; this strain produced several antimicrobial substances such as bacilysin, chlorotetaine, and iturin A, whose efficacy was potent against E. coli and S. aureus, thus explaining the efficacy of the traditional medicinal uses of pozol by the Mayan civilization [59].
This aspect is very important, as substances produced by several bacteria to protect themselves and to maintain a competitive advantage on other microorganisms not only contribute to the succession dynamics of the different microbial groups but also improve the microbiological quality and safety traits of this kind of products.
7. Reduction of Phytates
As with other cereal grains, maize contains some antinutritional factors such as phytic acid (PA) or phytate (PI) as salt, polyphenols, and tannins that can cause serious problems to human health. In fact, by forming a complex with minerals and by enzymes inhibition, phytic acid reduces the bioavailability and digestibility of proteins and carbohydrates. PA is myo-inositol 1,2,3,4,5,6-hexakis (dihydrogen phosphate), and it is the principal storage form of phosphorus and inositol in several oil seeds and grains [127]. It accounts for 50–80% of the total phosphorous, particularly in maize, with the greater part of PA (>80%) concentrated in the germ. Untreated PA has the ability to chelate important cations such as Ca2+, Mg2+, Fe2+, and especially Zn2+, and residues of proteins, forming a very insoluble salt in which the minerals are very little bio-available as well. The phytases reduce the hexa form of phytic acid (IP6; myo-inositol 1,2,3,4,5,6-hexakisphosphate) into lower forms, such as IP5, IP4, IP3, IP2, IP1, and myo-inositol [128]. The lower forms of PA have a lower binding capacity for metals such as iron and zinc [129].
Phytic acid not only has negative health effects as it chelates important minerals but also because it impairs the absorption of lipids and proteins due to the inhibition of the enzymes pepsin, amylase, and trypsin [130]. In fact, some food processing can partially degrade PA [131], however, most of the food phytate remains not degraded and reaches the gastrointestinal tract, where gut microbiota is usually not efficient in expressing phytate-degrading enzymes (phytases). In this context, Markiewicz et al. [132] suggested that the efficacy of the phytate degradation improves when the microbiota is already adapted to a high content of phytate, as occurs in the vegetarians’ intestine.
The potential antinutritional effects of PI present in maize is, to some extent, limited by the fermentation process, which is one of the most effective measures to reduce its amount. In fact, during cereal fermentation, endogenous and microbial phytases find optimum pH conditions for their activities, therefore releasing minerals such as manganese (which is an important growth factor for LAB), iron, zinc, and calcium [31]. In particular, natural fermentation of maize can achieve a significant reduction in PI; for example, Ejigui et al. [111] showed a PI reduction of 61% in corn flour after 96 h of fermentation at 30 °C. Recently, Gabaza et al. [133] observed a reduction between 20% and 88% of phytic acid in slurries maize samples from five different locations in Zimbabwe, after 26 h of fermentation. The authors also observed an increase of iron and zinc bioaccessibility, correlated in part with the reduction of phytic acid. The hypothesis that microorganisms involved in maize fermentation constitute the active part in maize detoxification was supported by early studies in vitro [134]. The authors demonstrated that Bacillus subtilis, B. cereus, B. licheniformis, Bacillus megaterium, Pseudomonas maltophyla, and Pseudomonas aeruginosa isolated during maize fermentation were able to reduce the phytate-phosphorous up to 68%. Successively, many authors have reported positive results in the reduction of PI using lactic acid bacteria and yeasts, improving the bioavailability of minerals in fermented maize [135,136]. The reduction of phytic acid by the natural microbiota has also been reported during the fermentation of maize bran by Decimo et al. [137]; in this case a reduction of 50% at the end of the sourdough-like fermentation process (corresponding to the twelfth refreshment), was reported.
It has been suggested that starter cultures are more effective than natural contaminants in uninoculated fermentation in reducing the PI content [138]. The ability of a lactic acid fermenting system in decreasing the phytate amount of fermented maize is dependent on various characteristics, as well as on environmental conditions during growth, harvest and storage of the cereal. Fermentation of maize flour resulted in an 88% reduction of phytic acid, and even lower levels of phytic acid remained when a starter culture (61%) or a germinated flour (71%) were used [139]. In this context, positive results were obtained with Lactobacillus amylovorus that has been demonstrated to be a good phytase producer in the defined MRS medium supplemented with 1% glucose and 24 mg of PI [140], and with L. buchneri M11 that reduced the PI content up to 95.5% after 72 h at 30 °C in kutukutu (from Cameroon). In addition, the levels of calcium, potassium, magnesium, sodium, sulphur, and zinc were significantly (p < 0.05) increased in amaku (from Nigeria) fermented with L. plantarum strains [3]. The capability of selected yeast, isolated from fermented dough and beverages, to reduce the PI content has been also reported, in particular for P. kudriavzevii, S. cerevisiae, Candida tropicalis and Pichia kluyveri [141,142].
Some yeast strains originating from togwa seemed to have developed a high phytase production; particularly Pichia kudriavzevii TY13 and Hanseniaspora guilliermondii TY14 [143] showed a great myo-inositol hexakisphosphate (IP6) degrading capacity during fermentation in maize-based model togwa, being able to degrade about 95% of the initial phytate amount. S. cerevisiae, C. krusei, C. tropicalis, and G. candidum were also able to degrade phytate.
During fermentation, other metabolic pathways are involved in the production of several interesting bioactive molecules. For example, phenolic compounds such as phenolic acids, flavonoids, and tannins, deriving from whole grain cereals, could be metabolized by microorganisms and modified into higher bioactive compounds (i.e., catechin, quercetin, and gallic acid) [144]; these and other microbial-based transformations are worthy of investigation.
8. Reduction of Mycotoxins
Mycotoxins are fungal metabolites commonly occurring in food, which pose a health risk for consumers. Maize is considered the most susceptible crop to mycotoxin contamination. In particular, aflatoxins (AFs), cyclopiazonic acid (CA), fumonisins (FUM), and zearalenone (ZEN) are the most frequent mycotoxins revealed in maize [145], although other mycotoxins such as cereulide and patulin have been reported in maize products [146]. It is well known that maize contamination with mycotoxins can occur either during pre-harvest, when the crop plant is growing, or during post-harvest processing. Mycotoxins are very stable to the physical and chemical treatments used in food processing, thus their elimination is very difficult; nevertheless, some processes, such as cleaning, milling, brewing, fermentation, cooking, baking, frying, roasting, flaking, alkaline cooking, nixtamalization (soaking, cooking in an alkaline solution, and hulling of grains), and extrusion [147] have been shown to reduce the mycotoxin content. During fermentation, maize detoxification can be achieved by microbial binding and/or biotransformation of mycotoxins into less toxic compounds [148]. Several strains of lactic acid bacteria and yeasts exhibit detoxifying properties, and their potential in removing mycotoxins has been reported [149]. This capability has been associated with the noncovalent binding of mycotoxins by fractions of the cell wall skeleton of lactic acid bacteria and yeasts [150]. On the basis of several studies, it appeared that pH and temperature influenced the binding, pH 4 and 37 °C being the most favorable conditions, although with differences among the strains [151]. Some mycotoxins (e.g., AFB1, FB1, and ZEN) were proven to be degraded to various extents by fermentation or biotransformation during maize fermentation processes. In this context, the aflatoxin B1 removal can occur due to the opening of AFB1 lactone ring, resulting in its complete detoxification [152].
The individual or synergistic activity of lactic acid bacteria or yeast strains applied as starter cultures has also been explored to reduce the mycotoxin content in maize products. In general, efficient detoxification can be achieved by the deliberate introduction of lactic acid bacteria strains. Table 4 reports some examples of the species used and the related mycotoxins reduction achieved.
The results of these studies indicate that lactic acid bacteria and some yeast species showed a good performance in reducing the mycotoxin content. In addition, as suggested by Cho et al. [161] and Haskard et al. [162], mycotoxins degradation during fermentation may either be strain-specific or may require synergistic interactions of more than one species/strain. Moreover, other non-LAB species (e.g., Bacillus subtilis) have been implicated in ZEN degradation (up to 99% of 1 mg/kg after 24 h) in liquid medium [75,163]. An excellent review documenting the decontamination of mycotoxins in fermented foods is available in the literature [163] and can be consulted for further information.
9. Conclusions
During maize fermentation, as in other natural fermentation, the competition among species for substrates, the acid tolerance, the syntrophic interactions, and other physiological properties of microbial populations cause variations in the microbiota structure. The studies have revealed that natural microbiota produces a wide assortment of compounds that are made accessible for all the community members, also inducing changes of the nutritional, rheological and sensorial treats of the fermented product, and with undeniable positive effects on the consumers, by the reduction of undesirable compounds and the improvement in vitamin and mineral availability.
Through spontaneous fermentation, the maize, a relatively poor cereal, can be transformed into a rich product, as the basis of the diet for many populations all over the world. However, because of the complexity of the microbiota and of the interactions occurring between microorganisms and maize environment, the role of the single microbial groups on the product is not always very obvious. In fact, although in the last years more attention has been paid to the microbiota of fermented maize products, it is important to deeply study the role particularly of subdominant populations, in order to better understand the processes that shape and drive the composition and dynamics of the maize fermentation as an essential step, not only to improve the process but also to safeguard human health.
Author Contributions
Conceptualization, writing—original draft preparation, C.C.-L.; writing—review and editing, A.S.; data curation, C.R. and F.M.; supervision, A.P. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Nwokoro, O.; Chukwu, B.C. Studies on Akamu, a traditional fermented maize food. Rev. Chil. Nutr. 2012, 39, 180–184. [Google Scholar]
- Obinna-Echem, P.C.; Kuri, V.; Beal, J. Evaluation of the microbial community, acidity and proximate composition of akamu, a fermented maize food. J. Sci. Food Agric. 2014, 94, 331–340. [Google Scholar] [CrossRef] [PubMed]
- Obinna-Echem, P.C.; Beal, J.; Kuri, V. Effect of processing method on the mineral content of Nigerian fermented maize infant complementary food – Akamu. LWT-Food Sci. Technol. 2015, 61, 145–151. [Google Scholar] [CrossRef]
- Pérez-Cataluña, A.; Elizaquível, P.; Carrasco, P.; Espinosa, J.; Reyes, D.; Wacher, C.; Aznar, R. Diversity and dynamics of lactic acid bacteria in Atole agrio, a traditional maize-based fermented beverage from South-Eastern Mexico, analysed by high throughput sequencing and culturing. Antonie van Leeuwenhoek 2018, 111, 385–399. [Google Scholar] [CrossRef] [PubMed]
- Väkeväinen, K.; Valderrama, A.; Espinosa, J.; Centurión, D.; Rizo, J.; Reyes-Duarte, D.; Díaz-Ruiz, G.; von Wright, A.; Elizaquível, P.; Esquivel, K.; et al. Characterization of lactic acid bacteria recovered from atole agrio, a traditional Mexican fermented beverage. LWT-Food Sci. Technol. 2018, 88, 109–118. [Google Scholar] [CrossRef] [Green Version]
- Steinkraus, K.H. Handbook of Indigenous Fermented Food, 2nd ed.; Marcel Dekker: New York, NY, USA, 1996. [Google Scholar]
- Von Mollendorff, J.W.; Vaz-Velho, M.; Todorov, S.D. Boza, a traditional cereal-based fermented beverage: A rich source of probiotics and bacteriocin-producing lactic acid bacteria. In Functional Properties of Traditional Foods; Kristbergsson, K., Ötles, S., Eds.; Springer: Boston, MA, USA, 2016; pp. 157–188. [Google Scholar]
- Okaru, A.O.; Abuga, K.O.; Kibwage, I.O.; Hausler, T.; Luy, B.; Kuballa, T.; Rhem, J.; Lachenmeier, D.W. Aflatoxin contamination in unrecorded beers from Kenya–A health risk beyond ethanol. Food Control 2017, 79, 344–348. [Google Scholar] [CrossRef]
- Osorio-Cadavid, E.; Chaves-López, C.; Tofalo, R.; Paparella, A.; Suzzi, G. Detection and identification of wild yeasts in Champús, a fermented Colombian maize beverage. Food Microbiol. 2008, 25, 771–777. [Google Scholar] [CrossRef]
- Chaves-López, C.; Serio, A.; Grande-Tovar, C.D.; Cuervo-Mulet, R.; Delgado-Ospina, J.; Paparella, A. Traditional fermented foods and beverages from a microbiological and nutritional perspective: The Colombian heritage. Compr. Rev. Food Sci. Food Saf. 2014, 13, 1031–1048. [Google Scholar] [CrossRef] [Green Version]
- Elizaquível, P.; Pérez-Cataluña, A.; Yépez, A.; Aristimuño, C.; Jiménez, E.; Cocconcelli, P.S.; Vignolo, G.; Aznar, R. Pyrosequencing vs. culture-dependent approaches to analyze lactic acid bacteria associated to chicha, a traditional maize-based fermented beverage from Northwestern Argentina. Int. J. Food Microbiol. 2015, 198, 9–18. [Google Scholar] [CrossRef]
- Mendoza, L.M.; Neef, A.; Vignolo, G.; Belloch, C. Yeast diversity during the fermentation of Andean chicha: A comparison of high-throughput sequencing and culture-dependent approaches. Food Microbiol. 2017, 67, 1–10. [Google Scholar] [CrossRef]
- Assohoun, M.C.N.; Djéni, T.N.; N’Guessan, F.K.; Koussemon, M. Preliminary study on antimicrobial properties of Lactic acid bacteria involved in the fermentation of corn dough during doklu processing in Côte D’Ivoire. Food 2012, 6, 65–70. [Google Scholar]
- Assohoun, M.C.; Djeni, T.N.; Koussémon-Camara, M.; Brou, K. Effect of fermentation process on nutritional composition and aflatoxins concentration of doklu, a fermented maize based food. Food Nutr. Sci. 2013, 4, 1120. [Google Scholar]
- Assohoun-Djeni, N.M.C.; Djeni, N.T.; Messaoudi, S.; Lhomme, E.; Koussemon-Camara, M.; Ouassa, T.; Chobert, J.M.; Onno, B.; Dousset, X. Biodiversity, dynamics and antimicrobial activity of lactic acid bacteria involved in the fermentation of maize flour for doklu production in Côte d’Ivoire. Food Control 2016, 62, 397–404. [Google Scholar] [CrossRef]
- Greppi, A.; Rantsiou, K.; Padonou, W.; Hounhouigan, J.; Jespersen, L.; Jakobsen, M.; Cocolin, L. Determination of yeast diversity in ogi, mawè, gowé and tchoukoutou by using culture-dependent and-independent methods. Int. J. Food Microbiol. 2013, 165, 84–88. [Google Scholar] [CrossRef] [PubMed]
- Adinsi, L.; Akissoé, N.H.; Dalodé-Vieira, G.; Anihouvi, V.B.; Fliedel, G.; Mestres, C.; Hounhouigan, J.D. Sensory evaluation and consumer acceptability of a beverage made from malted and fermented cereal: Case of gowe from Benin. Food Sci Nutr 2015, 3, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Adinsi, L.; Mestres, C.; Akissoé, N.; Vieira-Dalodé, G.; Anihouvi, V.; Durand, N.; Hounhouigan, D.J. Comprehensive quality and potential hazards of gowe, a malted and fermented cereal beverage from West Africa. A diagnostic for a future re-engineering. Food Control. 2017, 82, 18–25. [Google Scholar] [CrossRef]
- Gadaga, T.H.; Mutukumira, A.N.; Narvhus, J.A.; Feresu, S.B. A review of traditional fermented foods and beverages of Zimbabwe. Int. J. Food Microbiol. 1999, 53, 1–11. [Google Scholar] [CrossRef]
- Kalui, C.M.; Mathara, J.M.; Kutima, P.M.; Kiiyukia, C.; Wongo, L.E. Partial characterisation and identification of lactic acid bacteria involved in the production of ikii: A traditional fermented maize porridge by the Kamba of Kenya. J. Trop Microbiol. Biotechnol. 2008, 4, 3–15. [Google Scholar] [CrossRef]
- Kalui, C.M.; Mathara, J.M.; Kutima, P.M.; Kiiyukia, C.; Wongo, L.E. Functional characteristics of Lactobacillus plantarum and Lactobacillus rhamnosus from ikii, a Kenyan traditional fermented maize porridge. Afr. J. Biotechnol. 2009, 8, 4363–4373. [Google Scholar]
- Mkandawire, P.; Luginaah, I.; Tobias, J. Landscapes of economic deprivation and locally distilled liquor (kachasu): An emerging milieu of HIV/AIDS risk in urban Northern Malawi. Environ. Plan A. 2011, 43, 2384–2398. [Google Scholar] [CrossRef]
- Jespersen, L.; Halm, M.; Kpodo, K.; Jakobsen, M. Significance of yeasts and moulds occurring in maize dough fermentation for ‘kenkey’production. Int. J. Food Microbiol. 1994, 24, 239–248. [Google Scholar] [CrossRef]
- Nout, M.J.R.; Kok, B.; Vela, E.; Nche, P.F.; Rombouts, F.M. Acceleration of the fermentation of kenkey, an indigenous fermented maize food of Ghana. Food Res. Int. 1996, 28, 599–604. [Google Scholar] [CrossRef]
- Atter, A.; Ofori, H.; Anyebuno, G.A.; Amoo-Gyasi, M.; Amoa-Awua, W.K. Safety of a street vended traditional maize beverage, ice-kenkey, in Ghana. Food Control. 2015, 55, 200–205. [Google Scholar] [CrossRef]
- Oduro-Yeboah, C.; Mestres, C.; Amoa-Awua, W.; Fliedel, G.; Durand, N.; Matignon, B.; Michodjehoun, V.L.; Saalia, F.K.; Sakyi-Dawson, E.; Abbey, L. Steeping time and dough fermentation affect the milling behaviour and quality of white kenkey(nsiho), a sour stiff dumpling prepared from dehulled maize grains. J. Cereal. Sci. 2016, 69, 377–382. [Google Scholar] [CrossRef]
- Andah, A.; Muller, H.G. Studies on koko, a Ghanaian fermented maize porridge. Ghana J Agric Sci 1973, 6, 103–108. [Google Scholar]
- Oranusi, S.; Dahunsi, S.O. Preliminary study on hazards and critical control points of kokoro, a Nigerian indigenous fermented maize snack. SpringerPlus. 2015, 4, 253. [Google Scholar] [CrossRef] [Green Version]
- Bolade, M.K. Physical and organoleptic characteristics of non-sour ‘kokoro’(a Nigerian maize-based snack) as influenced by flour particle size differential. LWT-Food Sci. Technol. 2018, 87, 287–292. [Google Scholar] [CrossRef]
- Tchikoua, R.; Tatsadjieu, N.L.; Mbofung, C.M.F. Effect of selected lactic acid bacteria on growth of Aspergillus flavus and aflatoxin B 1 production in kutukutu. J Microbiol Res 2015, 5, 84–94. [Google Scholar]
- Blandino, A.; Al-Aseeri, M.E.; Pandiella, S.S.; Cantero, D.; Webb, C. Cereal-based fermented foods and beverages. Food Res. Int. 2003, 36, 527–543. [Google Scholar] [CrossRef]
- Prado, F.C.; Parada, J.L.; Pandey, A.; Soccol, C.R. Trends in non-dairy probiotic beverages. Food Res. Int. 2008, 41, 111–123. [Google Scholar] [CrossRef]
- Chelule, P.K.; Mbongwa, H.P.; Carries, S.; Gqaleni, N. Lactic acid fermentation improves the quality of amahewu, a traditional South African maize-based porridge. Food Chem. 2010, 122, 656–661. [Google Scholar] [CrossRef]
- Franz, C.M.; Huch, M.; Mathara, J.M.; Abriouel, H.; Benomar, N.; Reid, G.; Galvez, A.; Holzapfel, W.H. African fermented foods and probiotics. Int. J. Food Microbiol. 2014, 190, 84–96. [Google Scholar] [CrossRef] [PubMed]
- Awobusuyi, T.D.; Siwela, M.; Kolanisi, U.; Amonsou, E.O. Provitamin A retention and sensory acceptability of amahewu, a non-alcoholic cereal-based beverage made with provitamin A-biofortified maize. J. Sci. Food Agric. 2016, 96, 1356–1361. [Google Scholar] [CrossRef] [PubMed]
- Chaves-López, C.; Serio, A.; Delgado-Ospina, J.; Rossi, C.; Grande-Tovar, C.D.; Paparella, A. Exploring the bacterial microbiota of Colombian fermented maize dough “Masa Agria” (Maiz Añejo). Front Microbiol. 2016, 7, 1168. [Google Scholar] [CrossRef]
- Ramos, C.L.; Schwan, R.F. Technological and nutritional aspects of indigenous Latin America fermented foods. Curr. Opin. Food Sci. 2017, 13, 97–102. [Google Scholar] [CrossRef]
- Fowoyo, P.T.; Ogunbanwo, S.T. Phenotypic diversity of lactic acid bacteria isolated from Massa, a fermented maize dough. Afr. J. Microbiol. Res. 2010, 4, 2682–2691. [Google Scholar]
- Hounhouigan, D.J.; Nout, M.J.; Nago, C.M.; Houben, J.H.; Rombouts, F.M. Characterization and frequency distribution of species of lactic acid bacteria involved in the processing of mawé, a fermented maize dough from Benin. Int. J. Food Microbiol. 1993, 18, 279–287. [Google Scholar] [CrossRef]
- Hounhouigan, D.J.; Nout, M.J.; Nago, C.M.; Houben, J.H.; Rombouts, F.M. Composition and microbiological and physical attributes of mawè, a fermented maize dough from Benin. Int. J. Food Sci. Technol. 1993, 28, 513–517. [Google Scholar] [CrossRef]
- Agati, V.; Guyot, J.P.; Morlon-Guyot, J.; Talamond, P.; Hounhouigan, D.J. Isolation and characterization of new amylolytic strains of Lactobacillus fermentum from fermented maize doughs (mawè and ogi) from Benin. J. Appl. Microbiol. 1998, 85, 512–520. [Google Scholar] [CrossRef]
- Zulu, R.M.; Dillon, V.M.; Owens, J.D. Munkoyo beverage, a traditional Zambian fermented maize gruel using Rhynchosia root as amylase source. Int. J. Food Microbiol. 1997, 34, 249–258. [Google Scholar] [CrossRef]
- Foma, R.K.; Destain, J.; Mobinzo, P.K.; Kayisu, K.; Thonart, P. Study of physicochemical parameters and spontaneous fermentation during traditional production of munkoyo, an indigenous beverage produced in Democratic Republic of Congo. Food Control. 2012, 25, 334–341. [Google Scholar] [CrossRef]
- Schoustra, S.E.; Kasase, C.; Toarta, C.; Kassen, R.; Poulain, A.J. Microbial community structure of three traditional Zambian fermented products: Mabisi, chibwantu and munkoyo. PLoS ONE 2013, 8, e63948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Omemu, A.M.; Oyewole, O.B.; Bankole, M.O. Significance of yeasts in the fermentation of maize for ogi production. Food Microbiol. 2007, 24, 571–576. [Google Scholar] [CrossRef] [PubMed]
- Omemu, A.M. Fermentation dynamics during production of ogi, a Nigerian fermented cereal porridge. Rep. Opin. 2011, 3, 8–17. [Google Scholar]
- Bolaji, O.T.; Adepoju, P.A.; Olalusi, A.P. Economic implication of industrialization of a popular weaning food ogi production in Nigeria: A review. Afr. J. Food Sci. 2015, 9, 495–503. [Google Scholar] [CrossRef] [Green Version]
- Izah, S.C.; Kigigha, L.T.; Okowa, I.P. Microbial quality assessment of fermented maize Ogi (a cereal product) and options for overcoming constraints in production. Biotechnol Res. 2016, 2, 81–93. [Google Scholar]
- Henshaw, E.; Akpan, J.; Lennox, J.; Frank, E. Bacterial community dynamics during the production of ogi from millet ‘a Nigerian fermented food’ using culture-dependent approach. Imp. J. Interdiscip. Res. 2016, 2, 621–623. [Google Scholar]
- Ekundayo, J.A. The production of pito, a Nigerian fermented beverage. J. Food Technol. 1969, 4, 217–225. [Google Scholar] [CrossRef]
- Onaghise, E.O.; Izuagbe, Y.S. Improved brewing and preservation of pito, a nigerian alcoholic beverage from maize. Eng. Life Sci. 1989, 9, 137–142. [Google Scholar] [CrossRef]
- Louembé, D.; Kéléké, S.; Kobawila, S.C.; Nzoussi, J.P. Variabilité et amélioration de la technologie traditionnelle de production de la pâte fermentée de maïs au Congo. Tropicultura 2004, 22, 211–218. [Google Scholar]
- Abriouel, H.; Omar, N.B.; López, R.L.; Martínez-Cañamero, M.; Keleke, S.; Gálvez, A. Culture-independent analysis of the microbial composition of the African traditional fermented foods poto poto and dégué by using three different DNA extraction methods. Int. J. Food Microbiol. 2006, 111, 228–233. [Google Scholar] [CrossRef] [PubMed]
- Ben-Omar, N.; Abriouel, H.; Keleke, S.; Valenzuela, A.S.; Martínez-Cañamero, M.; López, R.L.; Ortega, E.; Gálvez, A. Bacteriocin-producing Lactobacillus strains isolated from poto poto, a Congolese fermented maize product, and genetic fingerprinting of their plantaricin operons. Int. J. Food Microbiol. 2008, 127, 18–25. [Google Scholar] [CrossRef] [PubMed]
- Wacher, C.; Canas, A.; Cook, P.E.; Barzana, E.; Owens, J.D. Sources of microorganisms in pozol, a traditional Mexican fermented maize dough. World J. Microbiol. Biotechnol. 1993, 9, 269–274. [Google Scholar] [CrossRef] [PubMed]
- Ben-Omar, N.; Ampe, F. Microbial community dynamics during production of the Mexican fermented maize dough pozol. Appl. Environ. Microbiol. 2000, 66, 3664–3673. [Google Scholar] [CrossRef] [Green Version]
- Escalante, A.; Wacher, C.; Farres, A. Lactic acid bacterial diversity in the traditional Mexican fermented dough pozol as determined by 16S rDNA sequence analysis. Int. J. Food Microbiol. 2001, 64, 21–31. [Google Scholar] [CrossRef]
- Diaz-Ruiz, G.; Guyot, J.P.; Ruiz-Teran, F.; Morlon-Guyot, J.; Wacher, C. Microbial and physiological characterization of weakly amylolytic but fast-growing lactic acid bacteria: A functional role in supporting microbial diversity in pozol, a Mexican fermented maize beverage. Appl. Environ. Microbiol. 2003, 69, 4367–4374. [Google Scholar] [CrossRef] [Green Version]
- Phister, T.G.; O’Sullivan, D.J.; McKay, L.L. Identification of bacilysin, chlorotetaine, and iturin A produced by Bacillus sp. strain CS93 isolated from pozol, a Mexican fermented maize dough. Appl. Environ. Microbiol. 2004, 70, 631–634. [Google Scholar] [CrossRef] [Green Version]
- Wacher-Rodarte, M.D.C.; Trejo-Muñúzuri, T.P.; Montiel-Aguirre, J.F.; Drago-Serrano, M.E.; Gutiérrez-Lucas, R.L.; Castañeda-Sánchez, J.I.; Sainz-Espuñes, T. Antibiotic resistance and multidrug-resistant efflux pumps expression in lactic acid bacteria isolated from pozol, a nonalcoholic Mayan maize fermented beverage. Food Sci. Nutr. 2015, 4, 423–430. [Google Scholar] [CrossRef] [Green Version]
- Sanni, A.I. Chemical studies on sekete beer. Food Chem. 1989, 33, 187–191. [Google Scholar] [CrossRef]
- Adegoke, G.O.; Nwaigwe, R.N.; Oguntimein, G.B. Microbiological and biochemical changes during the production of sekete- A fermented beverage made from maize. J. Food Sci. Technol. 1995, 32, 516–518. [Google Scholar]
- Sangwan, S.; Kumar, S.; Goyal, S. Maize utilisation in food bioprocessing: An overview. In Maize: Nutrition Dynamics and Novel Uses; Chaudhary, D.P., Kumar, S., Langyan, S., Eds.; Springer: New Delhi, India, 2014; pp. 119–134. [Google Scholar]
- Mugula, J.K.; Nnko, S.A.M.; Narvhus, J.A.; Sørhaug, T. Microbiological and fermentation characteristics of togwa, a Tanzanian fermented food. Int. J. Food Microbiol. 2003, 80, 187–199. [Google Scholar] [CrossRef]
- Hjortmo, S.B.; Hellström, A.M.; Andlid, T.A. Production of folates by yeasts in Tanzanian fermented togwa. FEMS Yeast Res. 2008, 8, 781–787. [Google Scholar] [CrossRef] [Green Version]
- Marshall, E.; Mejia, D. Traditional Fermented Food and Beverages for Improved Livelihoods; Food and Agriculture Organization of the United Nations: Rome, Italy, 2012; pp. 1–79. [Google Scholar]
- Capozzi, V.; Fragrasso, M.; Russo, P. Microbiological safety and the management of microbial resources in artisanal foods and beverages: The need for a transdisciplinary assessment to conciliate actual trends and risks avoidance. Microorganisms 2020, 8, 306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaves-López, C.; Serio, A.; Rossi, C.; Pepe, A.; Compagnone, E.; Paparella, A. Interaction between Galactomyces geotrichum KL20B, Lactobacillus plantarum LAT3 and Enterococcus faecalis KE06 during milk fermentation. Fermentation 2017, 3, 52. [Google Scholar] [CrossRef] [Green Version]
- Domínguez-Ramírez, L.L.; Díaz-Ruiz, G.; Wacher, C. Maize (Zea mays L. subsp. mays) Fermentation. In Fermented Foods, Part II: Technological Interventions; Ray, R.C., Montet, D., Eds.; Taylor & Francis: Abingdon-on-Thames, UK, 2017. [Google Scholar]
- Soro-Yao, A.A.; Brou, K.; Amani, G.; Thonart, P.; Djè, K.M. The use of lactic acid bacteria starter cultures during the processing of fermented cereal-based foods in West Africa: A review. Trop Life Sci. Res. 2014, 25, 81–100. [Google Scholar] [PubMed]
- Achi, O.K.; Ukwuru, M. Cereal-based fermented foods of Africa as functional foods. Int. J. Microbiol. Appl. 2015, 2, 71–83. [Google Scholar]
- Kunene, N.F.; Geornaras, I.; von Holy, A.; Hastings, J.W. Characterization and determination of origin of lactic acid bacteria from a sorghum-based fermented weaning food by analysis of soluble proteins and amplified fragment lenght polymorphism fingerprinting. Appl. Environl. Microbiol. 2000, 66, 1084–1092. [Google Scholar] [CrossRef] [Green Version]
- Ampe, F.; Miambi, E. Cluster analysis, richness and biodiversity indexes derived from denaturing gradient gel electrophoresis fingerprints of bacterial communities demonstrate that traditional maize fermentations are driven by the transformation process. Int. J. Food Microbiol. 2000, 60, 91–97. [Google Scholar] [CrossRef]
- Bassi, D.; Orrù, L.; Vasquez, J.C.; Cocconcelli, P.S.; Fontana, C. Peruvian chica: A focus on the microbial populations of this ancient maize-based fermented beverage. Microorganisms 2020, 8, 93. [Google Scholar] [CrossRef] [Green Version]
- Okeke, C.A.; Ezekiel, C.N.; Nwangburuka, C.C.; Sulyok, M.; Ezeamagu, C.O.; Adeleke, R.A.; Dike, S.K.; Krska, R. Bacterial diversity and mycotoxin reduction during maize fermentation (steeping) for ogi production. Front Microbiol. 2015, 6, 1402. [Google Scholar] [CrossRef] [Green Version]
- Obatolu, V.A.; Adeniyi, P.O.; Ashaye, O.A. Nutritional, sensory and storage quality of Sekete from Zea mays. Int J Food Sci Nutr Eng 2016, 6, 73–80. [Google Scholar]
- Mokoena, M.P.; Mutanda, T.; Olaniran, A.O. Perspectives on the probiotic potential of lactic acid bacteria from African traditional fermented foods and beverages. Food Nutr. Res. 2016, 60, 29630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fredlund, E.; Druvefors, U.Ä.; Olstorpe, M.N.; Passoth, V.; Schnürer, J. Influence of ethyl acetate production and ploidy on the anti-mould activity of Pichia anomala. FEMS Microbiol. Lett. 2004, 238, 133–137. [Google Scholar] [PubMed]
- Ponomarova, O.; Gabrielli, N.; Sévin, D.C.; Mülleder, M.; Zirngibl, K.; Bulyha, K.; Andrejev, S.; Kafkia, E.; Typas, A.; Sauer, U.; et al. Yeast creates a niche for symbiotic lactic acid bacteria through nitrogen overflow. Cell Syst. 2017, 5, 345–357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Almeida, E.G.; Rachid, C.C.; Schwan, R.F. Microbial population present in fermented beverage ‘cauim’produced by Brazilian Amerindians. Int. J. Food Microbiol. 2007, 120, 146–151. [Google Scholar] [CrossRef] [PubMed]
- De Vuyst, L.; Neysens, P. The sourdough microflora: Biodiversity and metabolic interactions. Trends Food Sci. Tech. 2005, 16, 43–56. [Google Scholar] [CrossRef]
- Teniola, O.D.; Odunfa, S.A.; Holzapfel, W.H. Selection, use and the influence of starter cultures in the nutrition and processing improvement of Ogi. In Proceedings of the 2nd international workshop: Food-based approaches for a healthy nutrition; Brouwer, I.D., Traore, A.S., Treche, S., Eds.; Universitaires de Ouagadougou: Ouagadougou, Burkina Faso, 2005; pp. 697–708. [Google Scholar]
- Reddy, G.; Altaf, M.D.; Naveena, B.J.; Venkateshwar, M.; Kumar, E.V. Amylolytic bacterial lactic acid fermentation-a review. Biotechnol. Adv. 2008, 26, 22–34. [Google Scholar] [CrossRef]
- Humblot, C.; Turpin, W.; Chevalier, F.; Picq, C.; Rochette, I.; Guyot, J.P. Determination of expression and activity of genes involved in starch metabolism in Lactobacillus plantarum A6 during fermentation of a cereal-based gruel. Int. J. Food Microbiol. 2014, 185, 103–111. [Google Scholar] [CrossRef]
- Mestres, C.; Nguyen, T.C.; Adinsi, L.; Hounhouigan, J.D.; Fliedel, G.; Loiseau, G. The interaction between starch hydrolysis and acidification kinetic determines the quality of a malted and fermented sorghum beverage. J. Cereal Sci. 2015, 63, 8–13. [Google Scholar] [CrossRef]
- Petrova, P.; Petrov, K.; Stoyancheva, G. Starch-modifying enzymes of lactic acid bacteria – structures, properties and applications. Strach Stärke 2013, 65, 34–47. [Google Scholar] [CrossRef]
- Betancourt Botero, S.P.; Bolívar Escobar, G.A.; Ramírez Toro, C. Fermentación de maíz de alta calidad proteica con Lactobacillus plantarum (CPQBA 087-11 DRM) aislado en Colombia de masas tradicionales fermentadas. Rev Argent Microbiol 2013, 45, 282–283. [Google Scholar] [CrossRef] [Green Version]
- Grijalva-Vallejos, N.; Aranda, A.; Matallana, E. Evaluation of yeasts from Ecuadorian chicha by their performance as starters for alcoholic fermentations in the food industry. Int. J. Food Microbiol. 2020, 317, 108462. [Google Scholar] [CrossRef] [PubMed]
- Rahmawati, J.; Dewanti-Hariyadi, R.; Hariyadi, P.; Fardiaz, D.; Richana, N. Isolation and identification of microorganisms during spontaneous fermentation of maize. J. Teknol. Dan. Industri. Pangan. 2013, 24, 33–39. [Google Scholar]
- Resende, L.V.; Pinheiro, L.K.; Miguel, M.G.D.C.P.; Ramos, C.L.; Vilela, D.M.; Schwan, R.F. Microbial community and physicochemical dynamics during the production of ‘Chicha’, a traditional beverage of Indigenous people of Brazil. World J. Microbiol. Biotechnol. 2018, 34, 46. [Google Scholar] [CrossRef]
- Sachdev, S.; Ojha, S.K.; Mishra, S. Bacillus spp. amylase: Production, isolation, characterisation and its application. Int. J. Appl. Sci. Biotechnol. 2016, 4, 3–14. [Google Scholar] [CrossRef] [Green Version]
- Putri, W.D.R.; Marseno, D.W.; Cahyanto, M.N. Role of lactic acid bacteria on structural and physicochemical properties of sour cassava starch. APCBEE Procedia. 2012, 2, 104–109. [Google Scholar] [CrossRef] [Green Version]
- FAO/WHO. Fermentation: Assessment and research. Report of a Joint FAO/WHO Workshop on fermentation as household technology to improve food safety; WHO Document WHO/FNU/FOS/96.1; World Health Organization: Geneva, Switzerland, 1996; pp. 1–79. [Google Scholar]
- Donot, F.; Fontana, A.; Baccou, J.C.; Schorr-Galindo, S. Microbial exopolysaccharides: Main examples of synthesis, excretion, genetics and extraction. Carbohydr. Polym. 2012, 87, 951–962. [Google Scholar] [CrossRef]
- Serrazanetti, D.I.; Guerzoni, M.E.; Corsetti, A.; Vogel, R. Metabolic impact and potential exploitation of the stress reactions in lactobacilli. Food Microbiol. 2009, 26, 700–711. [Google Scholar] [CrossRef]
- Galle, S.; Arendt, E.K. Exopolysaccharides from sourdough lactic acid bacteria. Crit. Rev. Food Sci. Nutr. 2014, 54, 891–901. [Google Scholar] [CrossRef]
- Falade, A.T.; Emmambux, M.N.; Buys, E.M.; Taylor, J.R. Improvement of maize bread quality through modification of dough rheological properties by lactic acid bacteria fermentation. J. Cereal Sci. 2014, 60, 471–476. [Google Scholar] [CrossRef] [Green Version]
- Tamani, R.J.; Goh, K.K.T.; Brennan, C.S. Physico-chemical properties of sourdough bread production using selected lactobacilli starter cultures. J. Food Qual. 2013, 36, 245–252. [Google Scholar] [CrossRef]
- Patel, S.; Majumder, A.; Goyal, A. Potentials of exopolysaccharides from lactic acid bacteria. Indian J. Microbiol. 2012, 52, 3–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adebayo-tayo, B.C.; Onilude, A.A. Screening of lactic acid bacteria strains isolated from some Nigerian fermented foods for EPS production. World Appl. Sci. J. 2008, 4, 741–747. [Google Scholar]
- Gänzle, M.; Schwab, C. Ecology of exopolysaccharide formation by lactic acid bacteria: Sucrose utilization, stress tolerance, and biofilm formation. In Bacterial Polysaccharides, Current Innovations and Future Trends; Ullrich, M., Ed.; Caister Academic Press: Norfolk, UK, 2009; pp. 263–278. [Google Scholar]
- Galle, S.; Schwab, C.; Arendt, E.; Gänzle, M. Exopolysaccharide-forming Weissella strains as starter cultures for sorghum and wheat sourdoughs. J. Agric. Food Chem. 2010, 58, 5834–5841. [Google Scholar] [CrossRef] [PubMed]
- Ua-Arak, T.; Jakob, F.; Vogel, R.F. Characterization of growth and exopolysaccharide production of selected acetic acid bacteria in buckwheat sourdoughs. Int. J. Food Microbiol. 2016, 239, 103–112. [Google Scholar] [CrossRef]
- Hiran, P.; Kerdchoechuen, O.; Laohakunjit, N. Combined effects of fermentation and germination on nutritional compositions, functional properties and volatiles of maize seeds. J. Cereal Sci. 2016, 71, 207–216. [Google Scholar] [CrossRef]
- Madhu, A.N.; Giribhattanavar, P.; Narayan, M.S.; Prapulla, S.G. Probiotic lactic acid bacterium from kanjika as a potential source of vitamin B12: Evidence from LC-MS, immunological and microbiological techniques. Biotechnol. Lett. 2010, 32, 503–506. [Google Scholar] [CrossRef]
- Masuda, M.; Ide, M.; Utsumi, H.; Niiro, T.; Shimamura, Y.; Murata, M. Production potency of folate, vitamin B12, and thiamine by lactic acid bacteria isolated from japanese pickles. Biosci. Biotechnol. Biochem. 2012, 76, 2061–2067. [Google Scholar] [CrossRef] [Green Version]
- De Angelis, M.; Bottacini, F.; Fosso, B.; Kelleher, P.; Calasso, M.; Di Cagno, R.; Ventura, M.; Picardi, E.; van Sinderen, D.; Gobbetti, M. Lactobacillus rossiae, a vitamin B12 producer, represents a metabolically versatile species within the genus Lactobacillus. PLoS ONE 2014, 9, e107232. [Google Scholar] [CrossRef] [Green Version]
- Murdock, F.A.; Fields, M.L. B-vitamin content of natural lactic acid fermented cornmeal. J. Food Sci. 1984, 49, 373–375. [Google Scholar] [CrossRef]
- Oladeji, B.S.; Akanbi, C.T.; Gbadamosi, S.O. Effects of fermentation on antioxidant properties of flours of a normal endosperm and quality protein maize varrieties. J. Food Meas. Caract. 2017, 11, 1148–1158. [Google Scholar] [CrossRef]
- Yépez, A.; Russo, P.; Spano, G.; Khomenko, I.; Biasoli, F.; Capozzi, V.; Aznar, R. In situ riboflavin fortification of different kefir-like cereal-based beverages using selected Andean LAB. Food Microbiol. 2019, 77, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Ejigui, J.; Savoie, L.; Marin, J.; Desrosiers, T. Beneficial changes and drawbacks of a traditional fermentation process on chemical composition and antinutritional factors of yellow maize (Zea mays). J. Biol. Sci. 2005, 5, 590–596. [Google Scholar]
- Saubade, F.; Hemery, Y.M.; Guyot, J.P.; Humblot, C. Lactic acid fermentation as a tool for increasing the folate content of foods. Crit. Rev. Food Sci. Nutr. 2017, 57, 3894–3910. [Google Scholar] [CrossRef] [PubMed]
- Kariluoto, S.; Aittamaa, M.; Korhola, M.; Salovaara, H.; Vahteristo, L.; Piironen, V. Effects of yeasts and bacteria on the levels of folates in rye sourdoughs. Int. J. Food Microbiol. 2006, 106, 137–143. [Google Scholar] [CrossRef] [PubMed]
- Fields, M.L.; Yoa, F.G. Nutritional improvement of corn meal by fermentation by Bacillus licheniformis and Enterobacter cloacae. J. Food Prot. 1990, 53, 245–248. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, M.; Maemura, K.; Oki, K.; Shiraishi, N.; Shibayama, Y.; Katsu, K. Gamma-aminobutyric acid GABA and cell proliferation, focus on cancer cells. Histol. Histopathol. 2006, 21, 1135–1141. [Google Scholar]
- Olsen, A.; Halm, M.; Jakobsen, M. The antimicrobial activity of lactic acid bacteria from fermented maize (kenkey) and their interactions during fermentation. J. Appl. Microbiol. 1995, 79, 506–512. [Google Scholar] [CrossRef]
- Olasupo, N.A.; Olukoya, D.K.; Odunfa, S.A. Assessment of a bacteriocin-producing Lactobacillus strain in the control of spoilage of a cereal-based African fermented food. Folia. Microbiol. 1997, 42, 31–34. [Google Scholar] [CrossRef]
- Quillama, E. Producción de bacteriocinas por cepas de Lactobacillus aisladas de chicha de jora. Master’s Thesis, Universidad Nacional Mayor de San Marcos, Lima, Peru, 1998. [Google Scholar]
- Kaktcham, P.M.; Zambou, N.F.; Tchouanguep, F.M.; El-Soda, M.; Choudhary, M.I. Antimicrobial and safety properties of Lactobacilli isolated from two Cameroonian traditional fermented foods. Sci. Pharm. 2012, 80, 189–203. [Google Scholar] [CrossRef] [Green Version]
- Onwuakor, C.E.; Nwaugo, V.O.; Nnadi, C.J.; Emetole, J.M. Effect of varied culture conditions on crude supernatant (bacteriocin) production from four Lactobacillus species isolated from locally fermented maize (Ogi). Am. J. Microbiol. Res. 2014, 2, 125–130. [Google Scholar] [CrossRef] [Green Version]
- Oguntoyinbo, F.A.; Narbad, A. Multifunctional properties of Lactobacillus plantarum strains isolated from fermented cereal foods. J. Funct. Foods 2015, 17, 621–631. [Google Scholar] [CrossRef] [Green Version]
- Ekwem, O.H. Isolation of antimicrobial producing lactobacilli from akamu (a Nigerian fermented cereal gruel). Afr. J. Microbiol. Res. 2014, 8, 718–720. [Google Scholar] [CrossRef] [Green Version]
- Lavermicocca, P.; Valerio, F.; Evidente, A.; Lazzaroni, S.; Corsetti, A.; Gobbetti, M. Purification and characterization of novel antifungal compounds from the sourdough Lactobacillus plantarum strain 21B. Appl. Environ. Microbiol. 2000, 66, 4084–4090. [Google Scholar] [CrossRef] [Green Version]
- Rizzello, C.G.; Cassone, A.; Coda, R.; Gobbetti, M. Antifungal activity of sourdough fermented wheat germ used as an ingredient for bread making. Food Chem. 2011, 127, 952–959. [Google Scholar] [CrossRef]
- Guimarães, A.; Santiago, A.; Teixeira, J.A.; Venâncio, A.; Abrunhosa, L. Anti-aflatoxigenic effect of organic acids produced by Lactobacillus plantarum. Int. J. Food Microbiol. 2018, 264, 31–38. [Google Scholar] [CrossRef] [Green Version]
- Moran, S.; Robertson, K.; Paradisi, F.; Rai, D.K.; Murphy, C.D. Production of lipopeptides in Bacillus sp. CS93 isolated from Pozol. FEMS Microbiol. Lett. 2010, 304, 69–73. [Google Scholar] [CrossRef] [Green Version]
- Vats, P.; Banerjee, U.C. Production studies and catalytic properties of phytases (myo-inositolhexakisphosphate phosphohydrolases): An overview. Enzyme Microb. Technol. 2004, 35, 3–14. [Google Scholar] [CrossRef]
- Ragon, M.; Aumelas, A.; Chemardin, P.; Santiago, S.; Moulin, G.; Boze, H. Complete hydrolysis of myo-inositol hexakisphosphate by a novel phytase from Debaryomyces castellii CBS 2923. Appl. Microbiol. Biotechnol. 2008, 78, 47–53. [Google Scholar] [CrossRef]
- Agte, V.V.; Gokhale, M.K.; Chiplonkar, S.A. Effect of natural fermentation on in vitro zinc bioavailability in cereal-legume mixture. Int. J. Food Sci. Tech. 1997, 31, 29–32. [Google Scholar] [CrossRef]
- Coulibaly, A.; Kouakou, B.; Chen, J. Phytic acid in cereal grains: Structure, healthy or harmful ways to reduce phytic acid in cereal grains and their effects on nutritional quality. Amer J. Plant Nutr. Fertil. Tech. 2011, 1, 1–22. [Google Scholar] [CrossRef] [Green Version]
- Afify, A.E.M.M.R.; El-Beltagi, H.S.; Abd El-Salam, S.M.; Omran, A.A. Bioavailability of iron, zinc, phytate and phytase activity during soaking and germination of white sorghum varieties. PLoS ONE 2011, 6, e25512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Markiewicz, L.H.; Honke, J.; Haros, M.; Świątecka, D.; Wróblewska, B. Diet shapes the ability of human intestinal microbiota to degrade phytate–in vitro studies. J. Appl. Microbiol. 2013, 115, 247–259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gabaza, M.; Shumoy, H.; Muchuweti, M.; Vandamme, P.; Raes, K. Iron and zinc bioaccessibility of fermented maize, sorghum and millets from five locations in Zimbabwe. Food Res. Int. 2018, 103, 361–370. [Google Scholar] [CrossRef] [PubMed]
- Lopez, Y.; Gordon, D.T.; Fields, M.L. Release of phosphorus from phytate by natural lactic acid fermentation. J. Food Sci. 1983, 48, 953–954. [Google Scholar] [CrossRef]
- Sandberg, A.S.; Andlid, T. Phytogenic and microbial phytases in human nutrition. Int. J. Food Sci. Technol. 2002, 37, 823–833. [Google Scholar] [CrossRef]
- Gernah, D.I.; Ariahu, C.C.; Ingbian, E.K. Effects of malting and lactic fermentation on some chemical and functional properties of maize (Zea mays). Am. J. Food Technol. 2011, 6, 404–412. [Google Scholar] [CrossRef] [Green Version]
- Decimo, M.; Quattrini, M.; Ricci, G.; Fortina, M.G.; Brasca, M.; Silvetti, T.; Manini, F.; Erba, D.; Criscuoli, F.; Casiraghi, M.C. Evaluation of microbial consortia and chemical changes in spontaneous maize bran fermentation. AMB Express. 2017, 7, 205. [Google Scholar] [CrossRef]
- Murali, A.; Kapoor, R. Effect of natural and pure culture fermentation of finger millet on zinc availability as predicted from HCl extractability and molar ratios. J. Food Sci. Technol. 2003, 40, 112–114. [Google Scholar]
- Hotz, C.; Gibson, R.S. Assessment of home-based processing methods to reduce the phytate content and phytate/zinc molar ratio of white maize (Zea mays). J. Agric. Food Chem. 2001, 49, 692–698. [Google Scholar] [CrossRef]
- Sreeramulu, G.; Srinivasa, D.S.; Nand, K.; Joseph, R. Lactobacillus amylovorus as a phytase producer in submerged culture. Lett. Appl. Microbiol. 1996, 23, 385–388. [Google Scholar] [CrossRef]
- Nuobariene, L.; Hansen, A.S.; Jespersen, L.; Arneborg, N. Phytase-active yeasts from grain-based food and beer. J. Appl. Microbiol. 2011, 110, 1370–1380. [Google Scholar] [CrossRef] [PubMed]
- Ogunremi, O.R.; Sanni, A.I.; Agrawal, R. Probiotic potentials of yeasts isolated from some cereal-based Nigerian traditional fermented food products. J. Appl. Microbiol. 2015, 119, 797–808. [Google Scholar] [CrossRef]
- Hellström, A.M.; Almgren, A.; Carlsson, N.G.; Svanberg, U.; Andlid, T.A. Degradation of phytate by Pichia kudriavzevii TY13 and Hanseniaspora guilliermondii TY14 in Tanzanian togwa. Int. J. Food. Microbiol. 2012, 153, 73–77. [Google Scholar] [CrossRef] [PubMed]
- Adebo, O.A.; Gabiela-Medina-Meza, I. Impact of fermentation on the phenolic compounds and antioxidant activity of whole cereal grains: A mini review. Molecules 2020, 25, 927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marin, S.; Ramos, A.J.; Cano-Sancho, G.; Sanchis, V. Mycotoxins: Occurrence, toxicology, and exposure assessment. Food Chem. Toxicol. 2013, 60, 218–237. [Google Scholar] [CrossRef] [PubMed]
- Abia, W.A.; Warth, B.; Ezekiel, C.N.; Sarkanj, B.; Turner, P.C.; Marko, D.; Krska, R.; Sulyok, M. Uncommon toxic microbial metabolite patterns in traditionally home-processed maize dish (fufu) consumed in rural Cameroon. Food Chem. Toxicol. 2017, 107, 10–19. [Google Scholar] [CrossRef] [PubMed]
- Karlovsky, P.; Suman, M.; Berthiller, F.; De Meester, J.; Eisenbrand, G.; Perrin, I.; Oswald, I.P.; Speijers, G.; Chiodini, A.; Recker, T.; et al. Impact of food processing and detoxification treatments on mycotoxin contamination. Mycotoxin Res. 2016, 32, 179–205. [Google Scholar] [CrossRef]
- Styriak, I.; Conková, E. Microbial binding and biodegradation of mycotoxins. Vet. Hum. Toxicol. 2002, 44, 358–361. [Google Scholar] [PubMed]
- Shetty, P.H.; Jespersen, L. Saccharomyces cerevisiae and lactic acid bacteria as potential mycotoxin decontaminating agents. Trends Food Sci. Technol. 2006, 17, 48–55. [Google Scholar] [CrossRef]
- Dawlal, P.; Brabet, C.; Thantsha, M.S.; Buys, E.M. Potential of lactic acid bacteria for the reduction of fumonisin exposure in African fermented maize based foods. World Mycotoxin. J. 2017, 10, 309–318. [Google Scholar] [CrossRef] [Green Version]
- Nout, M.J.R. Fermented foods and food safety. Food Res. Int. 1994, 27, 291–298. [Google Scholar] [CrossRef]
- Mokoena, M.P.; Chelule, P.K.; Gqaleni, N. The toxicity and decreased concentration of aflatoxin B1 in natural lactic acid fermented maize meal. J. Appl. Microbiol. 2006, 100, 773–777. [Google Scholar] [CrossRef] [PubMed]
- Fandohan, P.; Zoumenou, D.; Hounhouigan, D.J.; Marasas, W.F.O.; Wingfield, M.J.; Hell, K. Fate of aflatoxins and fumonisins during the processing of maize into food products in Benin. Int. J. Food Microbiol. 2005, 98, 249–259. [Google Scholar] [CrossRef]
- Wacoo, A.P.; Mukisa, I.M.; Meeme, R.; Byakika, S.; Wendiro, D.; Sybesma, W.; Kort, R. Probiotic enrichment and reduction of aflatoxins in a traditional African maize-based fermented food. Nutrients 2019, 11, 265. [Google Scholar] [CrossRef] [Green Version]
- Oluwafemi, F.; Da-Silva, F.A. Removal of aflatoxins by viable and heat-killed Lactobacillus species isolated from fermented maize. J. Appl. Biosci. 2009, 16, 871–876. [Google Scholar]
- Ezekiel, C.N.; Abia, W.A.; Ogara, I.M.; Sulyok, M.; Warth, B.; Krska, R. Fate of mycotoxins in two popular traditional cereal-based beverages (kunu-zaki and pito) from rural Nigeria. LWT-Food Sci. Technol. 2015, 60, 137–141. [Google Scholar] [CrossRef]
- Nyamete, F.A.; Mourice, B.; Mugula, J.K. Fumonisin B 1 reduction in lactic acid bacteria fermentation of maize porridges. Tanzania. J. Agric. Sci. 2016, 15, 13–20. [Google Scholar]
- Nyamete, F.A.; Bennink, M.; Mugula, J.K. Potential of lactic acid fermentation in reducing aflatoxin B1 in Tanzania maize-based gruel. Afr. J. Food Agric. Nutr. Dev. 2016, 16, 11139–11151. [Google Scholar] [CrossRef]
- Mokoena, M.P.; Chelule, P.K.; Gqaleni, N. Reduction of fumonisin B1 and zearalenone by lactic acid bacteria in fermented maize meal. J. Food Prot. 2005, 68, 2095–2099. [Google Scholar] [CrossRef]
- Niderkorn, V.; Boudra, H.; Morgavi, D.P. Binding of Fusarium mycotoxins by fermentative bacteria in vitro. J. Appl. Microbiol. 2006, 101, 849–856. [Google Scholar] [CrossRef] [PubMed]
- Cho, K.J.; Kang, J.S.; Cho, W.T.; Lee, C.H.; Ha, J.K.; Song, K.B. In vitro degradation of zearalenone by Bacillus subtilis. Biotechnol. Lett. 2010, 32, 1921–1924. [Google Scholar] [CrossRef] [PubMed]
- Haskard, C.A.; El-Nezami, H.S.; Kankaanpää, P.E.; Salminen, S.; Ahokas, J.T. Surface binding of aflatoxin B1 by lactic acid bacteria. Appl. Environ. Microbiol. 2001, 67, 3086–3091. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adebiyi, J.A.; Kayitesi, E.; Adebo, O.A.; Changwa, R.; Njobeh, P.B. Food fermentation and mycotoxin detoxification: An African perspective. Food Control. 2019, 106, 106731. [Google Scholar] [CrossRef]
Figure 1.
The interactions among the principal microbial groups in maize fermentation.

Figure 2.
The effect of microbial activities on the characteristics of the final fermented maize product.
Figure 2.
The effect of microbial activities on the characteristics of the final fermented maize product.

Figure 3.
Traditional maize pretreatments.


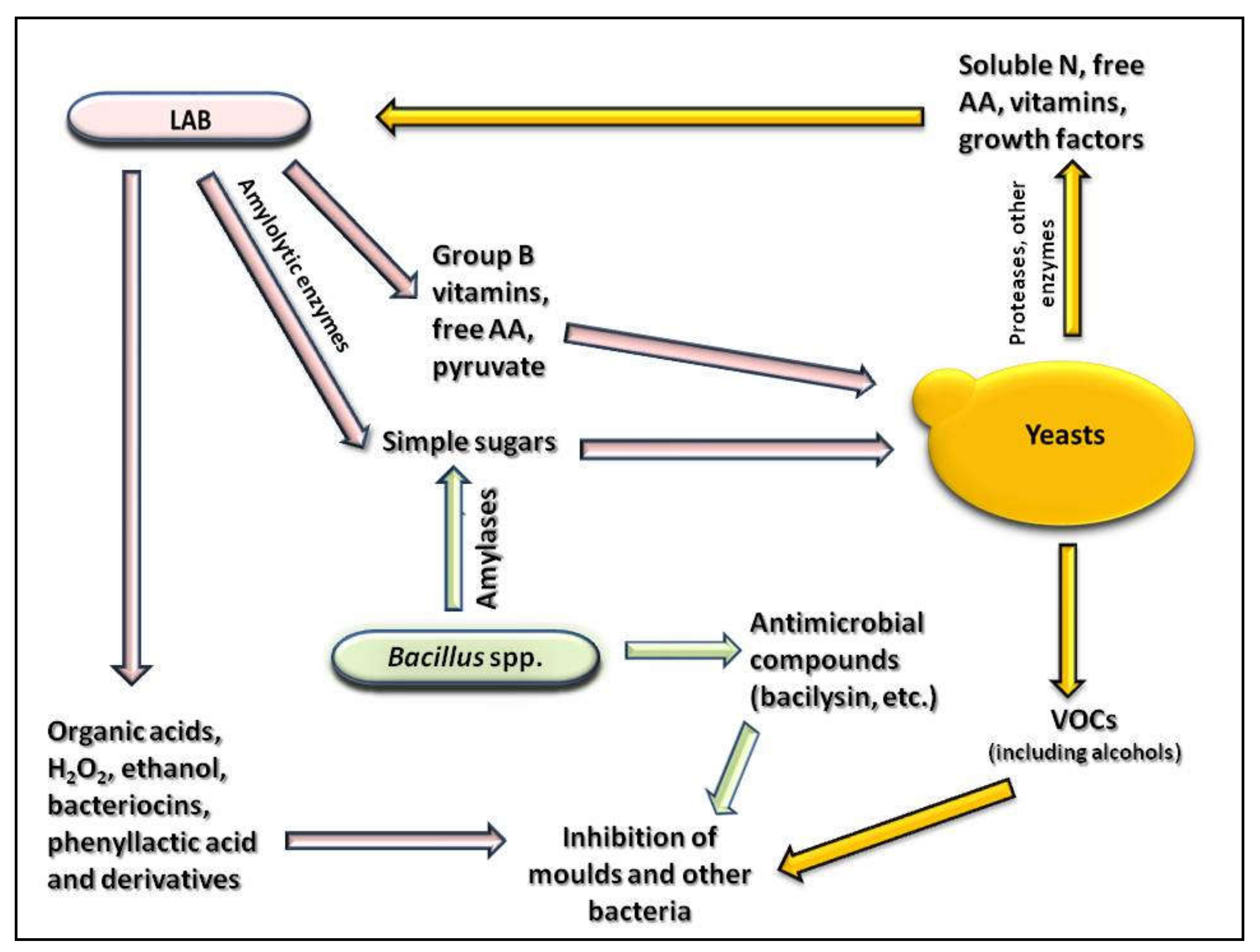{kind=link}
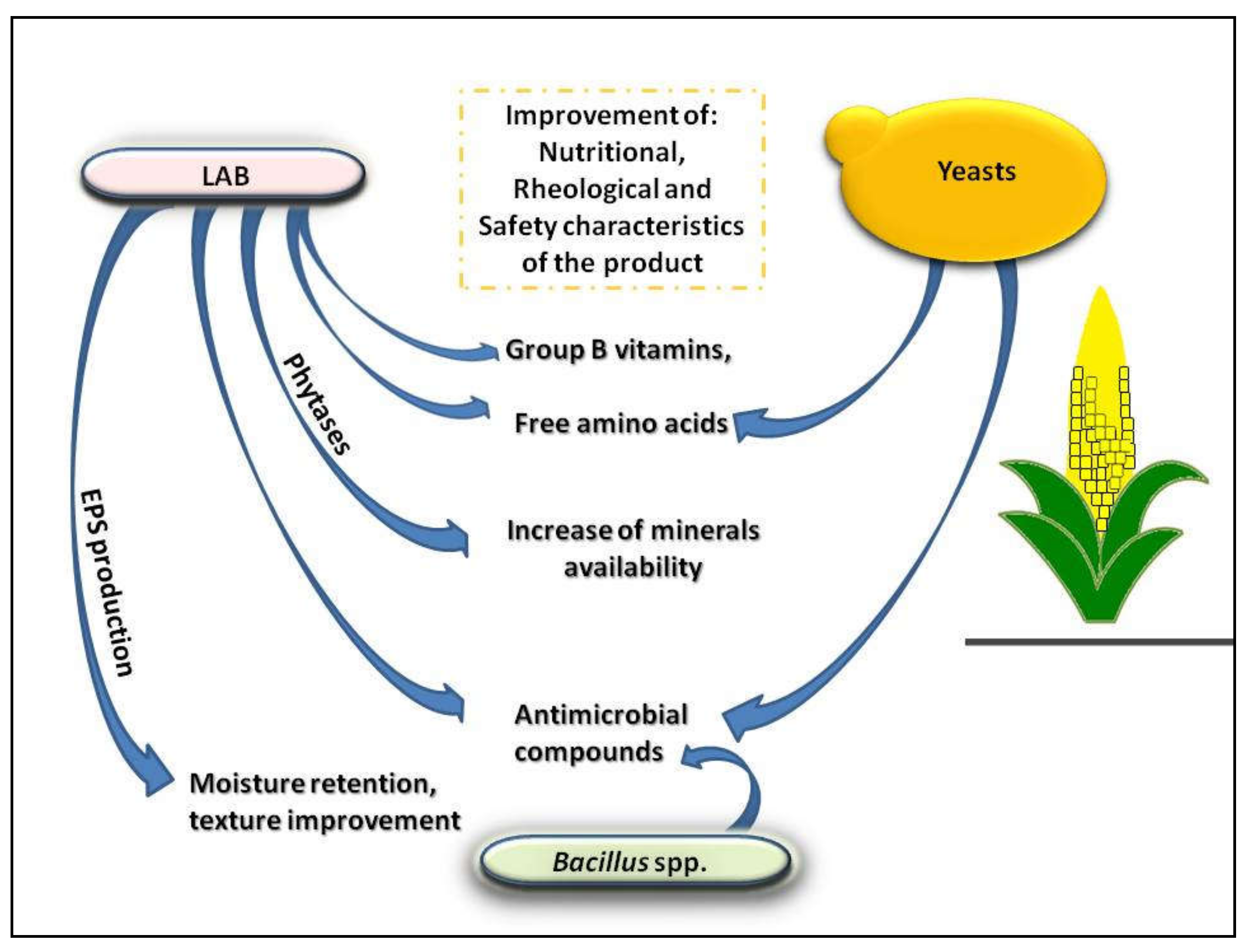{kind=link}
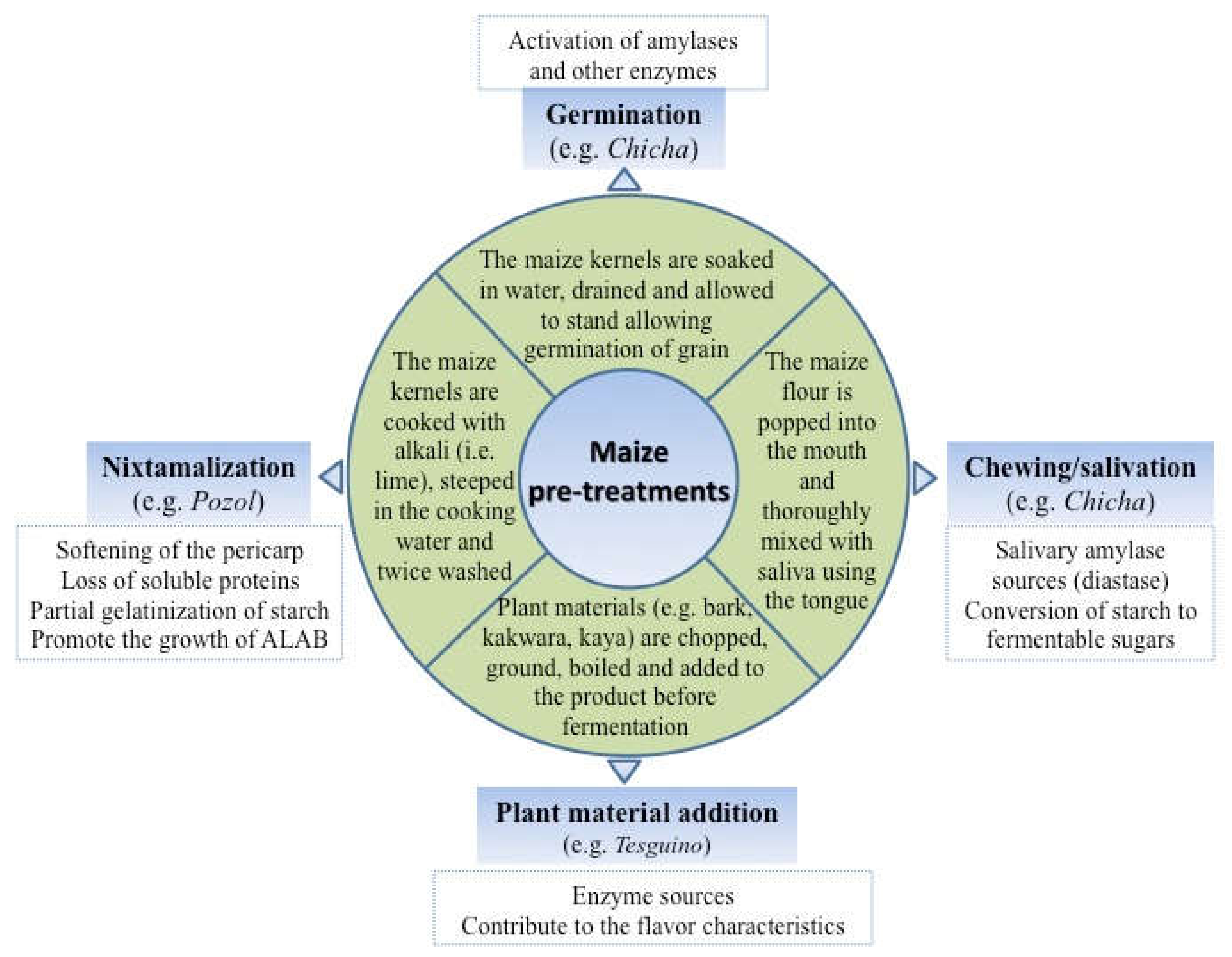{kind=link}
Table 1.
Maize-based fermented food products and their technological characteristics.
| Name of Product | Country | Food Category | Fermentation Time–Temperature | pH | Ethanol Content (v/v %) | References |
|---|---|---|---|---|---|---|
| Akamu | Nigeria | Porridge | 72 h, 28–30 °C | 3.2–3.9 | - | [1,2,3] |
| Atole agrio | Mexico | Beverage | 6–12 h, 34 °C | 3.9 | - | [4,5] |
| Busaa | Kenya, Nigeria, Ghana | Beverage | 5–7 d + 2–3 d, room T | 3.5–4.0 | 3.5–4.8 | [6,7,8] |
| Champús | Colombia, Ecuador, Perù | Beverage | 24–48 h, 12–15 °C | 3.5–4.0 | 2.5–4.2 | [9,10] |
| Chicha | Argentine | Beverage | 3–7 d + 2 d, room T | 3.6–3.8 | 2.0–12.0 | [11,12] |
| Chicha | Colombia | Beverage | 2–6 d, 18–32 °C | 3.5–4.6 | 2.0–12.0 | [10] |
| Doklu | Côte d’Ivoire | Dough | 2–4 d, room T | 2.2–3.8 | - | [13,14,15] |
| Gowé | Benin | Dough | 16 h, room T | 3.6–4.1 | - | [16,17,18] |
| Ilambazi lokubilisa | Zimbabwe | Porridge | 2–4 d, room T | n.a. | - | [19] |
| Ikii | Kenya | Porridge | n.a. | 3.9 | - | [20,21] |
| Kachasu | Zimbabwe | Beverage | 4–7 d, room T | n.a. | 9.0–41.0 | [7,19,22] |
| Kenkey | Ghana | Dough | 2–4 d, room T | 3.7 | - | [23,24,25,26] |
| Koko | Ghana | Porridge | 2 d, room T | n.a. | - | [7,27] |
| Kokoro | Nigeria | Snack | 24 h, 27–31 °C | 6.2 | - | [28,29] |
| Kutukutu | Cameroon | Dough | 120 h, 25–30 °C | 2.7–3.1 | - | [30] |
| Mahewu/Amahewu | South Africa, Arabian gulf countries | Beverage | 24–72 h, room T | 3.5–3.6 | - | [31,32,33,34,35] |
| Masa agria | Colombia | Dough | 3–5 d, 35–40 °C | 3.1–4.4 | - | [10,36,37] |
| Massa | Nigeria | Snack | 12–24 h, room T | n.a. | - | [38] |
| Mawè | Benin, Togo | Dough | 72 h, 28–32 °C | 3.8–4.2 | - | [16,39,40,41] |
| Munkoyo | Katanga, Zambia, Southern Democratic Republic of Congo | Beverage | 24–48 h, 25–30 °C | 3.5–4.0 | 0.0–2.7 | [42,43,44] |
| Mutwiwa | Zimbabwe | Porridge | room T | n.a. | - | [19] |
| Ogi | Nigeria | Porridge | 48–72 h + 24–48 h, 28–30 °C | 3.8–4.1 | - | [16,45,46,47,48,49] |
| Pito | Nigeria | Beverage | 12 h + 12 h, room T | 4.9 | 3.0–4.0 | [50,51] |
| Poto poto | Congo | Dough | 55 h + 10–11 h, room T | 3.7–3.8 | - | [52,53,54] |
| Pozol | Mexico | Dough | 2–7 d, room T | 4.2–4.6 | - | [55,56,57,58,59,60] |
| Sekete | Nigeria | Beverage | 2–3 d, room T | 2.8–4.3 | 0.9–4.0 | [61,62] |
| Tesguino | Mexico | Beverage | 2–3 d, room T | n.a. | 3.7 | [63] |
| Togwa | Tanzania | Beverage | 12–24 h, room T | 3.1–3.3 | - | [64,65] |
n.a., not available.
Table 2.
Lactic acid bacteria and yeasts involved in the fermentation of maize food products.
| Name of Product | Lactic Acid Bacteria | Yeasts | References |
|---|---|---|---|
| Akamu | L. lactis, Lb. acidophilus, Lb. amylovorus, Lb. delbrueckii, Lb. fermentum, Lb. helveticus, Lb. plantarum | C. albicans, C. tropicalis, Candida utilis, Clavispora lusitaniae, Rhodotorula glutinis, Saccharomyces paradoxus | [1,2] |
| Atole agrio | E. asini, E. casseliflavus, E. faecium, E. hirae, E. mundtii, L. lactis, L. piscium, Lb. aviarius, Lb. brevis, Lb. casei, Lb. composti, Lb. coryniformis, Lb. curvatus, Lb. dixtrinicus, Lb. mali, Lb. fabifermentans, Lb. paracasei, Lb. paraplantarum, Lb. plantarum, Lb. pentosus, Lb. rhamnosus, Lc. garlicum, Lc. mesenteroides, Lc. pseudomesenteroides, W. cibaria, W. confusa, W. paramesenteroides, P. stilesii, S. equines, W. hellenica, W. oryzae, P. pentosaceus | u.s. | [4,5] |
| Busaa | Lb. brevis, Lb. buchneri, Lb. casei, Lb. helveticus, Lb. plantarum, Lb. salivarius, Lb. viridescens, P. damnosus, P. parvulus | Candida krusei, Saccharomyces cerevisiae | [6,7] |
| Champús | u.s. | Galactomyces geotrichum, Hanseniaspora sp., Issatchenkia orientalis, Pichia fermentans, P. kluyveri, Saccharomyces cerevisiae, Torulospora delbruekii, Zygosaccharomyces fermentati | [9,10] |
| Chicha | E. durans, E. faecalis, E. faecium, E. gallinarum, E. hirae, E. lactis, E. mundtii, L. lactis, Lb. acidophilus, Lb. aviarius, Lb. brevis, Lb. casei, Lb. composti, Lb. crispatus, Lb. diolivorans, Lb. fabifermentans, Lb. farraginis, Lb. fermentum, Lb. harbinensis, Lb. helveticus, Lb. murinus, Lb. odoratitofui, Lb. paracasei, Lb. paraplantarum, Lb. plantarum, Lb. reuteri, Lb. rossiae, Lb. suebicus, Lb. vaccinostercus, Lc. citreum, Lc. lactis, Lc. mesenteroides, Lc. pseudomesenteroides, S. equinus, S. gallolyticus W. cibaria, W. confusa, W.hellenica, W. viridescens | Candida parapsilosis, C. zeylanoides, Cryptococcus carnescens, Cry. flavescens, Cry. magnus, Cry. nemorosus, Hanseniaspora uvarum, Debaryomyces hansenii, Kluyveromyces lactis, K. marxianus, Meyerozyma guilliermondii, Pichia sp., P. fermentans, P. membranifaciens, Rhodotorula mucilaginosa, R. slooffiae, S. cerevisiae, Torulaspora delbrueckii, Wickerhamomyces anomalus, Trichosporon domesticum | [11,12] |
| Chicha | Lactobacillus sp., Leuconostoc sp. | Candida ethanolica, C. oleophila, C. parapsilosis, C. pomicola, C. railenensis, C. sergipensis, C. spandovensis, Hanseniaspora opuntiae, H. uvarum, Issatchenkia sp., Kazachstania exigua, Kodamaea ohmeri, Lodderomyces elongisporus, Metschnikowia koreensis, Monilia candida, Mycoderma vini, Oidium lactis, Pichia sp., P. guilliermondii, Saccharomyces cerevisiae, S. pastorianus, Wickerhamomyces anomalus, W. pijperi | [10] |
| Chica | Lactobacillus plantarum, Lb. fermentum, Weissella cibaria, Leuconostoc sp., Lactococcus sp., S. luteciae, S. alactolyticus | [74] | |
| Doklu | Enterococcus sp., Lactobacillus sp., Lb. fermentum, Lb. plantarum, Pediococcus sp., P. acidilactici, P. pentosaceus, Streptococcus sp., Weissella sp., W. cibaria | u.s. | [13,15] |
| Gowé | u.s. | u.s. | [18] |
| Ikii | Lb. confusus, Lb. fermentum, Lb. plantarum, Lb. rhamnosus, Pediococcus sp. | u.s. | [20,21] |
| Kachasu | - | u.s. | [7] |
| Kenkey | Lb. fermentum, Lb. reuteri | Candida kefir, C. krusei, C. mycoderma, C. tropicalis, Saccharomyces cerevisiae | [23] |
| Koko | Lb. brevis, Lb. plantarum | Saccharomyces cerevisiae | [7] |
| Kokoro | Lactobacillus sp. | - | [28] |
| Mahewu/amahewu | Leuconostoc spp., L. lactis, Lb. delbrueckii, S. lactis | - | [31,32,34] |
| Masa agria | Lactococcus sp., L. lactis, Lactobacillus sp., Lb. amylolyticus, Lb. brevis, Lb. coleohominis, Lb. delbrueckii, Lb. crustorum, Lb. curvatus, Lb. fermentum, Lb. gallinarum, Lb. helveticus, Lb. nagelii, Lb. nantensis, Lb. panis, Lb. plantarum, Lb. pontis, Lb. rossiae, Lb. siliginis, Lb. vaccinostercus, Lc. citreum, P. argentinicus, Streptococcus sp., Weissella sp., W. beninensis, W. confusa, W. fabalis, W. fabaria, W. salipiscis | u.s. | [10,36] |
| Massa | Lb. fermentum, Lb. lactis, Lb. plantarum, Lc. mesenteroides, Pediococcus acidilactici | u.s. | [38] |
| Mawè | L. lactis, Lb. brevis, Lb. buchneri, Lb. confusus, Lb. curvatus, Lb. fermentum, Lb. reuteri, Lb. salivarius, Lc. mesenteroides, Pediococcus acidilactici, P. pentosaceus | Candida krusei, Clavispora lusitaniae, Saccharomyces cerevisiae | [16,39,41] |
| Munkoyo | Lb. brevis, Lb. delbruekii, Lb. fermentum, Lb. plantarum, Lb. rossiae, W. cibaria, W. confusa | Saccharomyces cerevisiae | [43,44] |
| Mutwiwa | Pediococcus pentosaceus | u.s. | [19] |
| Ogi | E. faecalis, Lb. acidophilus, Lb. brevis, Lb. cellobiosus, Lb. fermentum, Lb. paraplantarum, Lb. plantarum, Lc. lactis, Lc. paramesenteroides, P. acidilactici, P. claussenii, P. pentosaceus, S. lactis | Candida albicans, C. krusei, C. tropicalis, C. utilis, Clavispora lusitaniae, Geotrichum candidum, G. fermentans, Rhodotorula glutinis, R. graminis, Saccharomyces cerevisiae, S. pastorianus | [16,45,46,48,49,75] |
| Pito | Lactobacillus sp. | Candida sp., Geotrichum candidum | [50] |
| Poto poto | Enterococcus sp., Lb. acidophilus, Lb. casei, Lb. delbrueckii, Lb. fermentum, Lb. gasseri, Lb. plantarum, Lb. reuteri | - | [53,54] |
| Pozol | Bifidobacterium minimum, Enterococcus sp., E. saccharolyticus, E. sulfureus, Lactococcus sp., L. lactis, Lactobacillus sp., Lb. alimentarius, Lb. casei, Lb. delbrueckii, Lb. fermentum, Lb. plantarum, Leuconostoc sp., Streptococcus sp., S. bovis, S. macedonicus, S. suis, Weissella sp. | u.s. | [55,56,57,58,59,60] |
| Sekete | L. lactis, Lb. brevis, Lb. delbrueckii, Lb. plantarum, Lc. mesenteroides, P. cerevisiae, Streptococcus spp. | Geotrichum sp., Saccharomyces spp., S. cerevisiae | [7,62,76] |
| Tesguino | Lactobacillus sp., Leuconostoc sp., Pediococcus sp., Streptococcus sp. | Brettanomyces sp., Candida guilliermondii, Cryptococcus sp., Geotrichum sp., Hansenula anomala, Pichia sp., Saccharomyces cerevisiae, S. kluyveri | [63] |
| Togwa | Lb. brevis, Lb., cellobiosus, Lb. fermentum, Lb. plantarum, P. pentosaceus, W. confusa | Candida glabrata, C. pelliculosa, C. tropicalis, Issatchenkia orientalis, Kluyveromyces marxianus, Pichia anomala, Saccharomyces cerevisiae | [64,65] |
u.s., unidentified species.
Table 3.
Enterobacteriaceae, molds and others bacteria involved in the fermentation of maize food products.
Table 3.
Enterobacteriaceae, molds and others bacteria involved in the fermentation of maize food products.
| Name of Product | Acetic Acid Bacteria | Enterobacteriaceae | Molds | Others | References |
|---|---|---|---|---|---|
| Akamu | - | Enterobacter aerogenes, Escherichia coli, Proteus sp., Serratia sp. | Aspergillus niger, A. oryzae, Mucor circinelloides, Penicillium citrinum, Rhizopus microsporus, R. oligosporus | Bacillus cereus, B. licheniformis, B. subtilis, Pseudomonas alkaligenes, P. aeruginosa | [1] |
| Atole agrio | Acetobacter estunensis, A. indonesiensis, A. pasteurianus, A. tropicalis, Gluconacetobacter sp., Gluconobacter sp., G. frateurii, Kozakia sp. | u.s. | u.s. | - | [4,5] |
| Chicha | - | - | Penicillium sp. | - | [12] |
| Chicha | Acetobacter sp. | - | Aspergillus sp., Penicillium sp. | - | [10] |
| Koko | - | Enterobacter cloacae | - | Acinetobacter sp. | [7] |
| Kokoro | - | Klebsiella sp., Proteus sp. | Alternaria sp., Aspergillus sp., Cephalosporium sp., Fusarium sp., Mucor sp., Penicillium sp., Rhizopus sp. | Bacillus sp., Pseudomonas sp., Staphylococcus sp. | [28] |
| Masa agria | Acetobacter sp., A. cibinongensis, A. fabarum, A. lovaniensis, A. orientalis, Gluconobacter oxydans | Enterobacter aerogenes, Escherichia sp., Pantoea agglomerans, Serratia sp. | - | Acinetobacter sp., A. junii, A. ursingii, Bacteroides sp., Comamonas terrigena, Dechloromonas sp., Delftia, Frateuria aurantia, Gemmata sp., Pseudomonas sp., Sphingobium sp., Sphingomonas sp., Stenotrophomonas bacterium, Sugarcane phytoplasma | [10,36] |
| Munkoyo | - | - | - | Bacillus licheniformis | [44] |
| Ogi | - | Citrobacter sp., Enterobacter sp., Escherichia coli, Klebsiella spp. | Aspergillus flavus, A. niger, A. oryzae, Fusarium subglutinans, Mucor circinelloides, Penicillium citrinum, Rhizopus microsporus, R. nigrigans, R. oligosporus, R. stolonifer | Acinetobacter berezinae, Aerobacter sp., Alcaligenes faecalis, Bacillus cereus, B. licheniformis, B. mycoides, B. subtilis, B. thuringiensis, Bordetella avium, B. bronchisepta, Corynebacterium sp., Micrococcus luteus, Moorella glycerini, Myroides marinus, Pseudomonas aeruginosa, P. hibiscicola, S. aureus | [46,48,49,75] |
| Pito | - | - | Aspergillus versicolor, Penicillium purpurogenum, P. simplicissimum | - | [50] |
| Poto poto | - | Escherichia coli | - | Bacillus sp. | [53] |
| Pozol | - | u.s. | u.s. | Bacillus sp., Clostridium sp, Exiguobacterium acetylicum, E. aurantiacum, Oxalophagus oxalicus | [55,56,57,59,60] |
| Sekete | - | - | Aspergillus flavus, A. niger, Mucor rouxii | Bacilus subtilis, Propionibacterium spp. | [7,62,76] |
u.s., unidentified species.
Table 4.
Reduction of mycotoxins (%) during fermentation.
| Mycotoxin | Detoxifying Microorganism | Reduction (%) | Strain Origin | Place of Fermentation | Reference |
|---|---|---|---|---|---|
| Aflatoxin B1 | Indigenous microbial communities | 40–60.8 | Ogi | Ogi | [55] |
| 27.5 | Maize meal | Maize meal | [153] | ||
| Lb. brevis | 63 | Kutukutu | Kutukutu | [30] | |
| Lb. buchneri | 64.2 | Kutukutu | Kutukutu | [30] | |
| Lb. rhamnosus and S. thermophilus | 92–100 | Commercial strains | Kwete | [154] | |
| S. lactis and Lb. delbrueckii | 75 | Commercial strains | Maize meal | [145] | |
| Aflatoxin B2 | Indigenous microbial communities | 68–82.8 | Ogi | Ogi | [55] |
| Lb. rhamnosus and S. thermophilus | 91.8–100 | Commercial strains | Kwete | [154] | |
| Aflatoxin M1 | Indigenous microbial communities | 100 | Ogi | Ogi | [55] |
| Aflatoxins | Indigenous microbial communities | 80 | Ogi | Ogi | [153] |
| ≥91 | Mawe | Mawe | [153] | ||
| Lb. acidophilus | 37.5 | Ogi | Maize | [155] | |
| Lb. brevis | 75 | Ogi | Maize | [155] | |
| Lb. casei | 62.5 | Ogi | Maize | [155] | |
| Lb. delbrueckii | 56.25 | Ogi | Maize | [155] | |
| Lb. plantarum | 95 | Ogi | Maize | [155] | |
| Alternariol | Indigenous microbial communities | 96.7 | Kunu-zaki | Kunu-zaki | [156] |
| Alternariolmethylether | Indigenous microbial communities | 96 | Kunu-zaki | Kunu-zaki | [156] |
| Beauvericin | Indigenous microbial communities | 99.9 | Kunu-zaki | Kunu-zaki | [156] |
| Citrinin | Indigenous microbial communities | 33–100 | Ogi | Ogi | [75] |
| Cyclopiazonic acid | Indigenous microbial communities | 98.1–100 | Ogi | Ogi | [75] |
| Deoxynivalenol | Indigenous microbial communities | 98.9 | Kunu-zaki | Kunu-zaki | [156] |
| Enniatins | Indigenous microbial communities | 94.7 | Kunu-zaki | Kunu-zaki | [156] |
| Fumonisin B1 | Back slopped* | 30 | Maize based porridge | Maize based porridge | [157] |
| 68 | Togwa | Togwa | [158] | ||
| Indigenous microbial communities | 20 | Maize based porridge | Maize based porridge | [157] | |
| 55 | Togwa | Togwa | [158] | ||
| 13–88.8 | Ogi | Ogi | [75] | ||
| 99.4 | Kunu-zaki | Kunu-zaki | [156] | ||
| 24.4 | Maize meal | Maize meal | [159] | ||
| Lb. casei | 17 | n.a. | Maize based porridge | [157] | |
| 52 | n.a. | Togwa | [158] | ||
| Lb. delbrueckii ssp. delbrueckii | 69 | Ogi and mahewu | in vitro: 24 h, 30°C, pH 4 | [150] | |
| 29 | Ogi and mahewu | in vitro: 6 d, 30°C, pH 4 | [150] | ||
| Lb. fermentum | 17 | n.a. | Maize based porridge | [157] | |
| 55 | n.a. | Togwa | [158] | ||
| Lb. plantarum | 73 | Ogi and mahewu | in vitro: 24 h, 30°C, pH 4 | [150] | |
| 8 | Ogi and mahewu | in vitro: 6 d, 30°C, pH 4 | [150] | ||
| 24 | n.a. | Maize based porridge | [157] | ||
| 55 | n.a. | Togwa | [158] | ||
| Pediococcus pentosaceus | 43 | Ogi and mahewu | in vitro: 24 h, 30°C, pH 4 | [150] | |
| 19 | Ogi and mahewu | in vitro: 6 d, 30°C, pH 4 | [150] | ||
| 24 | n.a. | Maize based porridge | [157] | ||
| 45 | n.a. | Togwa | [158] | ||
| S. lactis and Lb. delbrueckii | 74.6 | Commercial strains | Maize meal | [159] | |
| Fumonisin B2 | Indigenous microbial communities | 44–80 | Ogi | Ogi | [75] |
| Lb. delbrueckii ssp. delbrueckii | 95 | Ogi and mahewu | in vitro: 24 h, 30 °C, pH 4 | [150] | |
| 6 | Ogi and mahewu | in vitro: 6 d, 30 °C, pH 4 | [150] | ||
| Lb. delbrueckii ssp. bulgaricus | 55 | n.a. | in vitro: corn infusion, 24 h, 25°C, pH 4 | [152] | |
| Lb. plantarum | 95 | Ogi and mahewu | in vitro: 24 h, 30 °C, pH 4 | [143] | |
| 21 | Ogi and mahewu | in vitro: 6 d, 30 °C, pH 4 | [143] | ||
| Lb. rhamnosus | 80 | n.a. | in vitro: corn infusion, 24 h, 25 °C, pH 4 | [160] | |
| Leuconostoc mesenteroides | 65 | n.a. | in vitro: corn infusion, 24 h, 25 °C, pH 4 | [160] | |
| Pediococcus pentosaceus | 89 | Ogi and mahewu | in vitro: 24 h, 30 °C, pH 4 | [150] | |
| 67 | Ogi and mahewu | in vitro: 6 d, 30 °C, pH 4 | [150] | ||
| Fumonisin B3 | Indigenous microbial communities | 46–95 | Ogi | Ogi | [55] |
| Fumonisins | Indigenous microbial communities | 99.5 | Kunu-zaki | Kunu-zaki | [148] |
| 29 | Ogi | Ogi | [146] | ||
| ≥87 | Mawe | Mawe | [146] | ||
| Fusaproliferin | Indigenous microbial communities | 97.8 | Kunu-zaki | Kunu-zaki | [148] |
| Moniliformin | Indigenous microbial communities | 98.3 | Kunu-zaki | Kunu-zaki | [148] |
| Zearalenone | Indigenous microbial communities | 79–100 | Ogi | Ogi | [75] |
| 76.2 | Kunu-zaki | Kunu-zaki | [156] | ||
| 34.3 | Maize meal | Maize meal | [159] | ||
| Lb. delbrueckii ssp. bulgaricus | 75 | n.a. | in vitro: corn infusion, 24 h, 25 °C, pH 4 | [160] | |
| Lb. rhamnosus | 80 | n.a. | in vitro: corn infusion, 24 h, 25 °C, pH 4 | [160] | |
| Leuconostoc mesenteroides | 75 | n.a. | in vitro: corn infusion, 24 h, 25 °C, pH 4 | [160] | |
| S. lactis and Lb. delbrueckii | 68.2 | Commercial strains | Maize meal | [159] |
* back-slopped is referred to practice based on the inoculation of material coming from a previous batch culture. n.a., not available.
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Chaves-López, C.; Rossi, C.; Maggio, F.; Paparella, A.; Serio, A. Changes Occurring in Spontaneous Maize Fermentation: An Overview. Fermentation 2020, 6, 36. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation6010036
AMA Style
Chaves-López C, Rossi C, Maggio F, Paparella A, Serio A. Changes Occurring in Spontaneous Maize Fermentation: An Overview. Fermentation. 2020; 6(1):36. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation6010036
Chicago/Turabian StyleChaves-López, Clemencia, Chiara Rossi, Francesca Maggio, Antonello Paparella, and Annalisa Serio. 2020. "Changes Occurring in Spontaneous Maize Fermentation: An Overview" Fermentation 6, no. 1: 36. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation6010036
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.