Aromatic Potential of Diverse Non-Conventional Yeast Species for Winemaking and Brewing


1
NIZO, P.O. Box 20, 6710 BA Ede, The Netherlands
2
Food Technology Area, Faculty of Pharmacy, Universitat de València, Avda. Vicent Andrés Estellés s/n, Burjassot, 46100 Valencia, Spain
3
FerMentor, Ede, 6718 ZB Ede, The Netherlands
4
Wageningen Food & Biobased Research (WFBR), Wageningen University & Research, Bornse Weilanden 9, 6708 WG Wageningen, The Netherlands
*
Author to whom correspondence should be addressed.
Fermentation 2020, 6(2), 50; https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation6020050
Submission received: 8 April 2020
/
Revised: 4 May 2020
/
Accepted: 8 May 2020
/
Published: 11 May 2020
(This article belongs to the Special Issue Non-Saccharomyces Yeasts as Aroma Enhancers in Fermented Products)
Abstract
:Traditionally, Saccharomyces species are those used to conduct industrial alcoholic fermentations. Recently, an increasing interest has arisen with respect to the potential of so-called non-conventional yeasts to improve wine and beer aroma profiles, keeping the particular terroir of each region or for the development of craft beers. In this study, the potential of diverse non-conventional yeasts to improve aroma in winemaking and brewing was investigated, testing several pure and mixed culture combinations. In addition, a comparison between microscale and labscale was carried out in order to assess the value of microwine and microbeer as screening tools. The results indicated that non-Saccharomyces yeasts were good candidates to enhance or diversify aroma profiles in alcoholic beverages, especially regarding acetate ester yield and fruity aromas. However, mixed cultures with Saccharomyces spp. are normally required to achieve a successful fermentation. The adjustment of pithing ratios is crucial for this purpose. Microscale is presented as an effective and efficient screening tool to compare different culture combinations, although scaling-up will always be necessary in order to get results closer to real winemaking or brewing processes.
Keywords:
non-conventional yeasts; aromas; bioflavouring; wine; beer; microscale; alcoholic fermentation; screening1. Introduction
The most common starter cultures employed in alcoholic fermentation are Saccharomyces species. These species combine several desirable characteristics, such as efficiency in fermenting sugars and producing ethanol, synthesis of aromas, and high ethanol tolerance [1]. However, in the early stages of wine fermentation, as well as in spontaneously fermented wines, several non-Saccharomyces species also participate in the fermentation process. These species may have a positive impact on desirable organoleptic characteristics of the wine such as improved mouthfeel and more complex flavour profiles [2].
Recently there has been an increasing interest in both winemaking and brewing towards non-Saccharomyces yeasts for their potential for aroma enhancement while maintaining the particular terroir of each region or for the development of craft beers.
In the case of wine, application studies with a selection of these so-called non-conventional yeasts indeed improve wine aroma; for example, the sequential fermentation of Ribolla Gialla grape juice employing commercial Saccharomyces cerevisiae Lalvin T73 and Zygosaccharomyces kombuchaensis CBS8849 or Kazachstania gamospora CBS 10400 resulted in positive organoleptic properties and suitable fermentation dynamics, rapid sugar consumption and industrial strain compatibility [3]. In addition, Jood et al. [4] showed that mixed-culture fermentations involving S. cerevisiae and different Kazachstania species have very distinct aroma profiles, whereas Varela et al. [5] showed that wines fermented with a combination of Metschnikowia pulcherrima and S. uvarum were characterised by increased concentrations of 2-phenylethanol and 2-phenylethyl acetate, both associated with positive sensory attributes. In simultaneous and sequential inoculations of selected Rhodotorula mucilaginosa and S.cerevisiae strains, both the variety and levels of fermentative aromacompounds were improved, especially that of (Z)-3-hexene-1-ol, nerol oxide, certain acetates and ethyl esters. These changes are associated with an improved perception of citrus, sweet fruit, acid fruit, berry, and floral aroma [6].
In a similar approach, the use of non-Saccharomyces yeasts in brewing has been explored [7]. Torulaspora delbrueckii, for example, was studied in single-strain fermentations for the production of low-alcohol beer and for enhancing beer flavour profiles. Some strains fermented all the wort sugars, whereas others did not, but all strains tested displayed improved flavour-forming properties [8]. In another study, the use of pure cultures of T. delbrueckii and mixed cultures of T. delbrueckii and S. cerevisiae for brewing resulted in a well noticeable change in flavour profile due to T. delbrueckii. Distinctive sensory profiles with pure cultures of T. Delbrueckii were characterised by ‘fruit/citric’ notes and ‘full-bodied’ attributes whereas some mixed cultures enhanced the ‘fruity/ester’ and ‘hop’ aromas. At the same time, these beers showed enhanced clarity and a more persistent and compact foam [2]. Another approach involving non-Saccharomyces yeasts to produce low-alcohol beers was carried out by van Rijswijck et al. [9]. These authors found that the non-conventional yeasts Cyberlindnera fabianii and Pichia kudriavzevii both in pure and mixed cultures with S. cerevisiae yielded beer with lower ethanol content and higher levels of esters. Brettanomyces custersii, isolated from Lambic beer, showed higher β-glucosidase activity than S. cerevisiae, thus a higher potential for aroma release from precursors. Fermentation with pure cultures of a Brettanomyces custersii strain or in a co-culture with a S. cerevisiae resulted in enhanced release of volatiles from hop glycosides [2]. In a more recent study, Holt et al. [10] screened 17 non-conventional yeast species in sequential brewing fermentation, followed by inoculation of S. cerevisiae. In these fermentations, it was possible to enhance banana flavour (isoamyl acetate) by Pichia kluyverii, spicy notes (phenolic compounds) by Brettanomyces species and clove-like aroma (4-vinylguaiacol) by T. delbrueckii.
Furthermore, Gutiérrez et al. [11] explored the potential of 99 non-Saccharomyces yeasts to produce pleasant fruity aromas in three industrial media (beer, wine and cider) through an olfactory assay. Of these, 21 yeasts were further evaluated for their aroma profile and fermentation capacity using wort, grape and apple juice. These authors reached the conclusion that the choice of yeast has more impact than the medium composition. The yeasts with the best opportunities to tune beer flavour belong to the species Galactomyces geotrichum, Kazachstania zonata, Kluyveromyces lactis, Lindnera meyerae, Pichia kluyveri, Starmera caribaea, Yarrowia lipolytica and Saccharomycodes ludwigii. Finally, Ravasio et al. [12] explored the potential of 60 non-conventional yeasts to improve beer aroma and found that the species Wickerhamomyces anomalus was a good candidate to combine with lager yeast to increase fruity aroma.
In this paper, we analyse the fermentation performance and the aroma potential in microscale of 10 non-Saccharomyces species employed as pure and mixed cultures co-inoculated in different ratios with one Saccharomyces strain on both wort and wine must, showing that variation in pitching rates is a valid option to control diversification as well as the additional value in analysing large amounts of culture combinations using microwine and microbeer. In addition, a comparison between microscale and labscale was carried out to analyse the correspondence in fermentation performance and aroma profiles.
2. Materials and Methods
2.1. Strains
Ten non-Saccharomyces strains were included in this study and are listed in Table 1 (Westerdijk Fungal Biodiversity Institute-CBS-KNAW, Utrecht, The Netherlands). In addition, two commercial Saccharomyces species were included as references: S.cerevisiae Lalvin T73 (Lallemand) in wine and S. Pastorianus WS34/70 (Fermentis, Lesaffre) in beer.
2.2. Microscale Fermentations
Strains were pre-cultured overnight in GPY medium at 30 °C. After centrifugation (1000× g, 5 min), cells were resuspended in sterile Syrah must (Sofralab, Magenta, France) or enzymatically treated glucose wort (14.5°P) with no hops (Carlsberg, Copenhagen, Denmark). Wine must contained 250 g/L of sugars and was sterilized overnight using 1 mL/L dimethyl dicarbonate (Sigma-Aldrich, St. Louis, MO, USA) whereas wort was autoclaved.
Fermentations were carried out in triplicate employing 24-well microplates (MP), where each well contained 5 mL of must or wort. The MP were covered with a breathable seal (Sigma-Aldrich, St. Louis, MO, USA) and incubated at 25 °C for 10 days in anaerobic jars. The strains were inoculated in MP as pure cultures (OD600 nm of 0.2) or mixed cultures in combination with S. cerevisiae Lalvin T73 (microwine) or WS34/70 (microbeer) in proportion 1:1 (each strain OD600 nm of 0.1). In addition, extra microbeer experiments were carried out employing OD600 nm of 0.2 of non-Saccharomyces yeasts and OD600 nm of 0.04 and 0.01 of WS34/70 (5:1 and 20:1 proportions, respectively). These extra experiments were done due to cultures mixed in 1:1 ratio resulted in microbeers similar as those produced by pure S. cerevisiae culture, indicating the outcompeting growth rate of the commercial strain.
2.3. Residual Sugars and Ethanol Analysis by High-Performance Liquid Chromatography (HPLC)
Mono/di-saccharides (glucose, fructose, maltose, sucrose, maltotriose) and ethanol were determined by HPLC (Thermo Fisher Scientific, USA) equipped with a refractive index detector Shodex RI 101. Prior to analyses, proteins and fat in the samples were precipitated by Grimbely Biggs reagents and removed after centrifugation.
The HPLC system was equipped with a Sil-10 AD vp injector, a LC-10ATvp pump and a CTO-6A oven. Sample volume of 25 µL was injected onto the mixed bed guard AG50W-X4 400 mesh (20 × 9 mm, packed with H+, Bio-Rad, Hercules, CA, USA) and AG3-X4A, 200–400 mesh (Bio-Rad, Hercules, CA, USA). The column (Aminex HPX-87P, 300 × 7.8 mm; BioRad, Hercules, CA, USA), was held at 80 °C. Mono/disaccharides and ethanol were eluted isocratically with miliQ water, flowing at a rate of 0.4 mL/min. The autosampler temperature was 10 °C. The detection was carried out by a refractive index detector ERC-7510 (Erma, Japan). Data analysis was conducted with Chromeleon software version 6.60 (Thermo Fisher Scientific, Waltham, MA, USA). External standards (97%–99% purity) for the indicated mono/di-saccharides and ethanol were obtained by Sigma-Aldrich (St. Louis, MO, USA) and a 4-point calibration curve was constructed.
2.4. Aroma Analysis by GC-MS
Aroma compounds, higher alcohols, esters and vicinal diketones (Table 2) were extracted by headspace solid-phase microextraction (HS-SPME) employing a grey fibre DVB/CAR/PDMS (Divinylbenzene/Carboxen/Polydimethylsiloxane) and determined by GC-MS. The fibre was exposed to the headspace of the samples during 15 min at 60 °C and desorbed during 2 min in the GC injector at 250 °C. The injection device was a Combi PAL autosampler (CTC Analytics, Zwingen, Switzerland). The injected compounds were refocused at the beginning of the GC column by cryo-trapping at −110 °C (ThermoQuest Ce Instruments, Wigan, UK). Subsequently, the cryo-trap was heated to 200 °C at a rate of 50 °C/s.GC–MS analyses were carried out with a Finnigan Trace GC/MS (Thermo Fisher Scientific, Waltham, MA, USA) equipped with a 30 m × 0.25 µm VF-wax ms, df = 0.5 µm, capillary column (Varian, Palo Alto, CA, USA). The chromatographic oven was initially set at 40 °C for 1 min, then raised to 250 °C at 20 °C/min and then kept at 250 °C for 3 min. The carrier gas Helium was kept at a constant flow of 1.5 mL/min. The total runtime was 14.5 min. Mass spectral data was collected over a range of m/z 35–250 in full scan mode (scan time 0.25 s). Quantification of the compounds was done using calibration curves employing external standards of each aroma compound purchased from Sigma-Aldrich (St. Louis, MO, USA). Data analysis was performed by the Xcalibur 2.1 software (Thermo Fisher Scientific, Inc., Waltham, MA, USA).
2.5. Comparison of Microscale to Labscale
S. cerevisiae NIZO 900 was pre-cultured overnight at 30 °C in GPY (2% glucose, ScharlauChemie, Spain; 1% bacteriological peptone, Oxoid, UK; 0.5% yeast extract, Oxoid, UK). This strain was selected due to its good performance in fermenting alcoholic beverages. Afterwards, yeast cultures were centrifuged at 1000× g for 5 min, the medium was decanted, and the cells were subsequently re-suspended in triplicate in either 200 mL (bottles-lab scale) or 5 mL (microplates, MTPs-microscale) of Syrah must (Sofralab, Magenta, France) at a final OD600 nm of 0.1. Priortoinoculation, wine must was sterilized overnight with 1 mL/L dimethyl dicarbonate (Sigma-Aldrich, St. Louis, MO, USA). Samples were taken after 1, 3, 7 and 10 days of fermentation, centrifuged (4 °C, 1000× g, 5 min) and frozen immediately at −20 °C, except for OD measurements.
2.5.1. Biomass Analysis
Optical density measurement at 600 nm (OD600 nm) was carried out in a UV/visible spectrophotometer (Ultra-spec 2000, Amersham Pharmacia Biotech, Little Chalfont, UK). To correct for non-linearity of the assay, samples were pre-diluted in medium to obtain reads between 0.2 and 0.4.
2.5.2. Fermentation Kinetics and Aroma Profiles
Residual sugars and ethanol in the resulting fermentations were analysed by HPLC as described in Section 2.3.
Aroma profiles were determined by headspace solid-phase dynamic extraction (HS-SPDE) in combination with GC-MS. The HS-SPDE extraction was carried out using a 2.5 mL HS-syringe with a polydimethylsiloxane active charcoal coated needle (PDMS/AC). After the equilibration of the samples at 60 °C for 15 min, 1 mL of the headspace was injected into the GC column (1 min at 250 °C in splitless mode). GC and MS conditions were the same as described in Section 2.4.
2.6. Statistical Analysis
Multifactorial and one-way ANOVA were performed at 95% of confidence level (p value ≤ 0.05) with the results of aroma production and fermentation kinetics in all the experiments. Average comparison was done using Fisher (microwine) or Tukey’s (microbeer) tests. The statistical analyses were carried out by XLSTAT (Microsoft, Redmond, WA, USA).
3. Results and Discussion
3.1. Microwine
Microwine fermentations were carried out by 10 non-Saccharomyces strains (Table 1) in pure and mixed culture together with S. cerevisiae Lalvin T73 in proportion 1:1. Their fermentation performance and aroma potential were evaluated.
3.1.1. Fermentation Performance
Multifactorial ANOVA shows that non-conventional strain and type of culture, pure or mixed, were significant factors determining the residual sugar amount. However, just the type of culture was significantly influencing ethanol yield, indicating that ethanol synthesis was similar among the non-conventional yeasts employed in this study. Residual sugars and ethanol production of the resulting microwines are presented in Table 3.
One-way ANOVA points out that, in general, fermentations carried out by pure cultures of non-conventional yeasts left significantly higher amounts of residual sugars and produced lower quantities of ethanol compared to their corresponding mixed cultures and reference S. cerevisiae Lalvin T73. Ethanol was not even detected in case of Starmera caribaea CBS 7692 and Barnettozyma california CBS 8860 as practically no fermentation of the sugars occurred. One exception to this pattern was Wickerhamomyces subpelliculosus CBS 5552, which showed a fermentation performance as good as the reference strain even when used as a pure culture, leaving hardly any residual sugars and producing high ethanol levels.
Its fermentation efficiency was in accordance with [16], who observed similar ethanol production by this strain compared to S. cerevisiae baker’s yeast when bread fermentations were carried out and may indicate that this strain presented an intense Crabtree effect similar to S. cerevisiae.
The difference in ethanol production between pure and mixed cultures can be explained by the fact that it has been demonstrated that yeast in mixed cultures do not simply coexist, but rather interact, and as a result, oenological traits can be modulated [17]. The finding that single cultures of non-Saccharomyces yeasts are, in general, poor fermenters and sensitive to ethanol is in accordance to several authors [17,18,19]. S. cerevisiae, as consequence of Crabtree effect, prioritises fermentative metabolism over aerobic respiration when sugar levels are higher than 10 g/L. Nevertheless, many non-Saccharomyces yeasts do not show this effect and are able to grow using oxygen regardless of sugar concentration and thus, divert carbon into other metabolites, which reduces ethanol production [20]. In this way, fermentation performance improves when non-Saccharomyces yeasts are together with S. cerevisiae, with some exceptions. For instance, Dashko et al. [3] presented Kazachstania gamospora CBS 10400, the strain employed in this study, as Crabtree positive, observing a fermentation performance similar to S. cerevisiae in terms of residual sugars and ethanol yield when employed in sequential inoculation together to S. cerevisiae. In our study, CBS 10400 when used in mixed fermentation resulted in less residual sugars and much higher ethanol content, approaching that produced by S. cerevisiae.
On the other hand, the limited ethanol synthesis can also be considered an advantage for the wine sensory profile, since excessive ethanol amounts can increase the perception of bitterness and astringency and mask some volatile compounds [20]. Furthermore, this approach leads to the development of healthier wines and has been proposed as a strategy for counteracting climatic change and the resulting ethanol increase in wines [20]. Another interesting aspect is that it has been demonstrated that non-Saccharomyces strains that show poor efficiency in fermenting sugars may produce higher glycerol quantities, which is a highly desirable trait in wines, especially in red [11].
On the other hand, mixed cultures were similar to the reference strain in terms of residual sugars and ethanol yield. Torulaspora microellipsoides CBS 6641 in mixed culture even produced significantly more ethanol than the reference strain, being two-fold that produced in their corresponding pure culture. However, previous literature indicates that this strain produced low ethanol levels in sequential fermentations in wine, as well as when fermenting other substrates such as cashew apple juice [3,21]. Nevertheless, sequential and mixed fermentations can result in a different outcome and cashew apple juice present high quantity of phenolic compounds, which can exert microbial inhibitory effects.
3.1.2. Aroma Profiles
Table 4 shows the aroma profile of resulting microwines. The impact on aroma synthesis of non-Saccharomyces strains as a pure and mixed culture was studied employing multifactorial ANOVA. The type of culture, either pure or mixed, affected the production of all the aroma compounds except for ethyl caprate. In addition, the impact of non-conventional strains on acetate esters, phenylethyl alcohol and ethyl caprylate was significant.
Higher Alcohols
Higher alcohols contribute to floral, fruity or herbal aromas depending on their synergistic effects with other flavour-active compounds. In higher concentrations they could also influence flavour release and contribute to mouthfeel. Some of the most important higher alcohols in alcoholic beverages are isobutyl alcohol (solvent, fusel, spiritous), isoamyl alcohol (harsh, nail polish, banana) and phenylethyl alcohol (floral, rose, sweet), presenting, respectively, 40, 30 and 10 mg/L sensory thresholds in wine [13]. Excessive amounts of these compounds can be detrimental to wine quality; however, moderate amounts increase wine complexity [18].
Multifactorial ANOVA showed that the type of culture (pure or mixed) was a significant factor regarding higher alcohol yield. Contrarily, the non-Saccharomyces yeast participating in the fermentation did not influence the quantity of higher alcohols, except in the case of phenylethyl ethanol, where differences of even 8-fold were observed.
Isobutyl alcohol was above its sensory threshold in all of the mixed fermentations whereas isoamyl alcohol only in some of them (Table 4). In all these cases, the amount of the two alcohols were higher than in the corresponding pure non-Saccharomyces cultures and even higher than in the reference microwine, where the amounts of these alcohols were below their sensory thresholds (Table 4). Mixed fermentations carried out with Hanseniaspora guilliermondii CBS 2567, T. microellipsoides CBS 6641, Zygosaccharomyces kombuchaensis CBS 8849 and Zygosaccharomyces rouxii CBS 9716 had much higher isobutyl alcohol levels and Barnettozyma californica CBS 8860 had higher isobutyl and isoamyl alcohols amounts compared to the reference culture (Table 4). In previous works, intensive production of isobutyl and isoamyl alcohols was already seen in some of the non-Saccharomyces strains tested here, such as H. Guilliermondii CBS 2567 and T. Microellipsoides CBS 6641 when used as pure cultures in standard growth medium [22]. In this previous work, H. guilliermondii yielded 27 and 2.3 times the average quantity of isobutyl alcohol compared to Saccharomyces strains and W. subpelliculosus CBS 5552 produced 8 times the average quantity of isoamyl alcohol compared to Saccharomyces strains. Furthermore, the high production of alcohols by Z. kombuchaensis CBS 8849 has already been observed by other authors in pilot-scale sequential fermentations [3], suggesting a proper match between microscale and pilot scale regarding aroma profile.
It is worth mentioning the synergistic effect between reference strain and strains Starmera caribaea CBS 7692 and B. californica CBS 8860 regarding isoamyl alcohol production, since the amount of this alcohol in mixed fermentations was higher than the addition of quantities yielded by each strain separately (Table 5).
On the contrary, phenylethyl alcohol levels were higher in pure cultures, being above its sensory threshold in all cases except for S. caribaea CBS 7692 and with these differences being significant in the case of Galactomyces geotrichum CBS 772.71, W. Subpelliculosus CBS 5552, T. microellipsoides CBS 6641, S. caribaea CBS 7692 and K. gamospora CBS 10400 (Table 5). In mixed cultures, the amounts of this alcohol were below its sensory threshold with the exception of CBS 8031 (Hanseniaspora vineae). However, the reference microwine contained the highest phenylethyl alcohol level (Table 5).
Acetate Esters
Acetate esters are desirable aroma compounds presenting a wide range of fruity flavours and are of special interest inthe case of young wines produced from neutral grape varieties [18]. Among the most important acetate esters present in alcoholic beverages are ethyl acetate (nail polish, fruity; sensory threshold: 7.5 mg/L), isoamyl acetate (banana, pear; sensory threshold: 0.03mg/L) and phenylethyl acetate (flowery, rose, fruity, honey; sensory threshold: 0.25 mg/L) [13].
Multifactorial ANOVA points out that non-Saccharomyces strain and type of culture, mixed or pure, significantly affected acetate ester yields.
Reference microwine did not excel in the amounts of any acetate ester and isoamyl acetate was missing in most of the mixed cultures (Table 4), indicating a negative interaction between Saccharomyces and non-Saccharomyces strains regarding this aroma compound.
Despite being the main acetate ester in wine, ethyl acetate only appeared above its sensory threshold in four pure cultures: H. guilliermondii CBS 2567, W. subpelliculosus CBS 5552, S. caribaea CBS 7692 and K. gamospora CBS 10400 (Table 5). Excessive acetate ester production, a trait traditionally linked to non-Saccharomyces strains, can negatively affect the aroma profile, giving “oxidised” notes [17,18,23]. However, in our study this seems not to be the case.
Similarly, in the case of isoamyl and phenylethyl acetate, pure cultures yielded significantly higher amounts (Table 4), indicating high activity of the acetyltransferases, which must be inhibited to a certain degree in mixed cultures. A remarkable example in this sense is the strain S. caribaea CBS 7692, since its huge acetate ester production was not shown when combined with S. cerevisiae (Table 4). This extraordinary acetate ester synthesis of S. caribaea CBS 7692 when used as pure culture was already described in previous works, where this strain and H. guilliermondii CBS 2567 produced from 44- to 84-fold the levels found in reference S. cerevisiae pure culture [11,22]. H. guilliermondii CBS 2567 also excelled in the production of phenylethyl acetate and the precursor alcohol when fermenting cashew apple juice [21]. In our study, acetate ester production by H. guilliermondii CBS 2567 was not so huge, but still remarkable (Table 4).
It is worth mentioning the high isoamyl acetate production by pure cultures of S. caribaea CBS 7692 and K. gamospora CBS 10400, which probably resultsin a strong banana flavour, as already observed in bread fermented by the latter [16]. These strains also present high synthesis of phenylethyl acetate, which was already seen by Dashko et al. [3] in sequential pilot scale fermentations. The strains G. geotrichum CBS 772.72, Hanseniaspora vineae CBS 8031 and Z. kombuchaensis CBS 8849 also showed high production of phenylethyl acetate (Table 4), which probably resulted in a remarkable rose aroma. Jolly et al. [18] described high production of this ester in H. vineae and H. guilliermondii, which was in accordance with our results, especially in case of the former (see CBS 8031 in Table 4).
Ethyl Esters
Ethyl esters, as acetate esters, are desirable aroma compounds presenting fruity and flowery flavours. Some of the most important ethyl esters in alcoholic beverages are ethyl caproate (green apple, aniseed aroma; sensory threshold: 0.05 mg/L), ethyl caprylate (sour apple, sweet soap aroma; sensory threshold: 0.02 mg/L) and ethyl caprate (apple, floral, soap aroma; sensory threshold: 0.20 mg/L) [13].
Multifactorial ANOVA shows that the type of culture carrying out the fermentation, pure or mixed, significantly influenced ethyl caproate and ethyl caprylate yield, being the latter also affected by the non-Saccharomyces strain. On the contrary, none of the factors were significantly important to determine ethyl caprate levels, which even appeared below its sensory threshold in some pure cultures.
Despite not being a significant difference probably due to high standard deviations linked to the low amounts detected, the following trend could be observed: ethyl caproate levels were equal or higher in mixed cultures compared to their corresponding pure cultures and the reference microwine (Table 4).
Regarding ethyl caprylate and ethyl caprate, only the microwine produced by pure culture of Z. rouxii CBS 9716 presented higher amounts compared to reference microwine (Table 4). Pure culture of W. subpelliculosus CBS 5552 also showed high ethyl caprylate quantity (Table 4). The relatively high ethyl ester production by S. cerevisiae compared to other Saccharomyces and non-Saccharomyces strains is a typical trait extensively described in the literature [3,22,24].
3.2. Microbeer
Microbeer fermentations were carried out with ten non-Saccharomyces strains (Table 1) in pure and mixed cultures together with S. pastorianus WS34/70 in proportions 1:1, 5:1 and 20:1 (non-Saccharomyces: S. pastorianus). Cultures mixed in 1:1 ratio, resulted in fermentation characteristics matching a pure Saccharomyces culture, indicating the outcompeting growth rate of the commercial strain. Thus, only the results of the 5:1 and 20:1 fermentations are shown.
3.2.1. Fermentation Performance
Multifactorial ANOVA shows that the quantity of residual sugars is affected by the adjunct strains, but the levels of ethanol were similar in all fermentations (Table 5). In more detail, most of the yeast combinations showed good fermentation performance, consuming practically all the sugars present in wort (Table 5). However, there were some exceptions to this pattern: small sugar amounts (5–8 g/L) were left by non-conventional yeasts H. guilliermondii CBS 2567, Z. kombuchaensis CBS 8849 and Z. rouxii CBS 9716 when used in combination to WS34/70 in proportion 5:1 and T. microellipsoides CBS 6641 in proportion 20:1 (Table 5). Different sugar consumption efficiency was already seen in previous works by these strains [11,21], especially by Z. rouxii, which is not able to properly ferment maltose [7]. Surprisingly, T. microellipsoides CBS 6641 left more residual sugars when used in higher proportion, indicating an interaction between yeasts regarding their fermentation abilities already seen in mixed fermentations by other authors [17]. These differences in sugar consumption were not translated into significant differences in ethanol yield, which was around 6% in all the fermentations (Table 5). This fact could be explained by the bifurcation of metabolic flux to glycerol production by certain strains which would use part of the sugars to this end, thus reducing the expected ethanol yield and as a strategy to cope with hyperosmotic stressful conditions, which has been extensively studied in Saccharomyces species [25,26].
3.2.2. Aroma Profiles
Table 6 shows the aroma profiles of the different microbeers. In this study, the impact of two factors on the aroma of the resulting beer were investigated, non-Saccharomyces strain and the pitching ratio. Statistical analyses showed that the strain employed is affecting the yield of all the aroma compounds except for isoamyl acetate whereas the pitching ratio especially impacted on vicinal diketones and certain esters, phenylethyl acetate, ethyl caproate and ethyl caprylate. Finally, strain and pitching ratio combined significantly influenced the amounts of vicinal diketones, phenylethyl acetate and ethyl caprate in line with previous studies [27,28].
Vicinal Diketones
Vicinal diketones are carbonyl compounds that are undesirable when they appear in high concentrations in beer. Diacetyl adds a buttery flavour to beer if found above its sensory threshold (0.10–0.15 mg/L) and 2,3-pentanedione its known for its toffee flavour (sensory threshold 0.90 mg/L) [14]. However, it is known that other aroma compounds could strongly influence the sensory threshold and perception of diacetyl in alcoholic beverages [29].
Both the adjunct strains, as well as dosage, significantly influenced the final quantity of vicinal diketones at the end of fermentation according to multifactorial ANOVA. The interaction of these two factors made a significant impact, as well. Diacetyl appears above its sensory threshold in most of the resulting microbeers (even in the reference microbeer), contrarily to the case of 2,3-pentanedione, whose quantity was below its sensory threshold in all of the cases (Table 6). However, it has to be taken into account that microbeers did not undergo maturation in this study. In the maturation process, yeast take up diacetyl and reduce it to 2,3-butanediol, which does not have an undesirable flavour [14].
Most of the 5:1 fermentations presented diacetyl levels near its threshold and similar to reference microbeer with the exception of H. guilliermondii CBS 2567, W. subpelliculosus CBS 5552, S. caribaea CBS 7692 and B. californica CBS 8860, which showed higher levels (Table 6). This fact was already observed in some of these non-conventional yeasts in previous works [22].
On the other hand, diacetyl levels, on average, increase with increasing dosage of adjunct strain, indicating that these non-conventional yeasts can produce more diacetyl, thus making necessary longer maturation times if used industrially. However, a few strains form an exception to the above and result in lower (W. subpelliculosus CBS 5552) or similar (H. guilliermondii CBS 2567, Z. rouxii CBS 9716 and K. gamospora CBS 10400) amounts of diacetyl when higher proportion of non-conventional yeast is employed.
Higher Alcohols
Multifactorial ANOVA showed that the non-Saccharomyces strain participating in the fermentation was a significant factor regarding higher alcohol yield. Contrarily, the pitching ratio did not influence the quantity of higher alcohols, which was in accordance to [30], who compared the brewing performance of Brettanomyces bruxellensis as pure culture at different pitching ratios and the only aroma compound presenting different levels was 4-vinylguaiacol. This pattern was just the opposite as in the case of microwine, evidencing the importance of the fermentative substrate employed to carry out the fermentation process.
In general, adjunct cultures resulted in equal or lower concentration of higher alcohols compared to the reference microbeer (Table 6). In all microbeers, isoamyl alcohol was the only alcohol present in concentrations above its sensory threshold (Table 6). Ravasio et al. [12] obtained similar results when screening 60 non-conventional yeast for aroma production in brewing as they only detected isoamyl alcohol and 2-phenylethanol as higher alcohols produced by all the yeasts. In our data set, strains Z. rouxii CBS 9716 and W. subpelliculosus CBS 5552 resulted in 2-phenylethyl alcohol levels higher than that found in the reference microbeer (Table 6). The high production of alcohols in by Z. rouxii has already been reported by other authors for beer as well as for other food products such as soy source [14].
Acetate Esters
Except for ethyl acetate, none of the acetate esters investigated reached its sensory threshold (Table 6).
In general, levels of ethyl acetate and isoamyl acetate did not vary between the reference microbeer and the adjunct strains nor between pitching ratios, except for ethyl acetate, which appeared slightly above its sensory threshold in just two strains, W. subpelliculosus CBS 5552 and Z. rouxii CBS 9716, not being significant differences between pitching ratios (Table 6). High ethyl acetate synthesis by Z. rouxii was observed in soy sauce fermentation and beer [7,14], whereas high levels of acetate esters, especially ethyl acetate, were previously described in other species of the genus Wickerhamomyces, W. anomalus [7,12].
In contrast to ethyl acetate and isoamyl acetate, both adjunct strain and pitching ratio were crucial in case of 2-phenylethyl acetate as multifactorial ANOVA showed. In some mixed fermentations, higher amounts of 2-phenylethyl acetate than the reference strain S. pastorianus WS34/70 were observed and higher levels corresponded with pitching ratio. These strains were H. guilliermondii CBS 2567, W. subpelliculosus CBS 5552, K. gamospora CBS 10400 and H. vineae CBS 8031, being the latter able to synthesize higher amounts of this compound even in 5:1 fermentation (Table 6). Gamero et al. [22] observed high acetate ester synthesis by H. guilliermondii, whereas Gutierrez et al. [11] described a strong fruity flavour with W. subpelliculosus, which was even able to mask a high production of phenolic compounds.
Ethyl Esters
Multifactorial ANOVA showed that the non-Saccharomyces strains as well as their pitching ratio were significant factors regarding ethyl ester yield with the exception of ethyl caprate, which was not affected by the strain proportion. However, none of the ethyl esters studied reached its sensory threshold in any of the resulting microbeers. However, together they could be perceived if they have nearly the same characteristics and receptors as a result of additive or synergistic effects.
In general, ethyl ester levels decrease with increasing pitching ratios of the non-conventional yeasts (Table 6). This is in line with several studies where Saccharomyces yeasts have pointed out their huge capacity to synthesise ethyl esters, which has been positively correlated with the ethanol yield and fermentation capacity [10,22]. The exceptionsto this pattern were microbeers involving B. californica CBS 8860 (5:1) and K. gamospora CBS 10400 (20:1), where a significantly higher amount of ethyl caprylate or ethyl caprate was found, respectively (Table 6).
3.3. Comparison Microscale vs.Labscale
To assess the effect of scaling-up, we compare microscale and labscale fermentations. Specifically, comparison of yeast growth, fermentation performance and aroma profiles were carried out between microwine and wine produced at labscale.
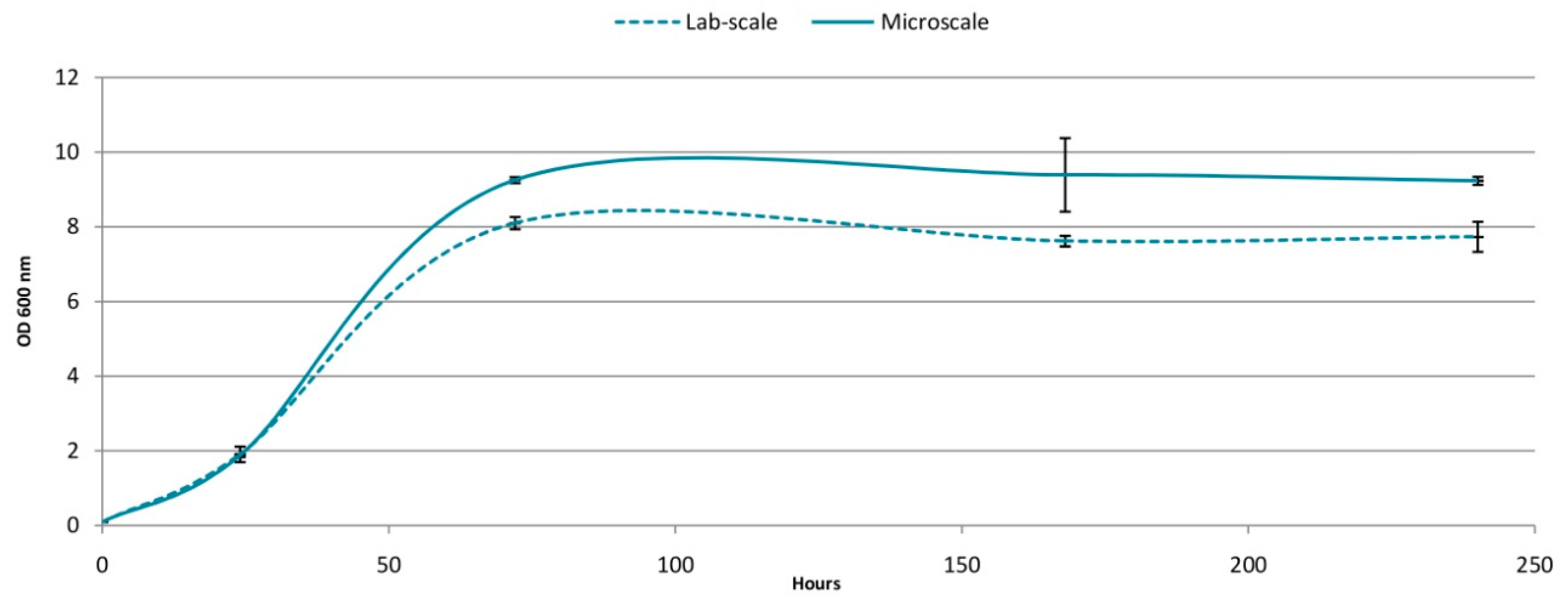
Similar trends were observed regarding yeast growth and fermentation kinetics despite appearing significant differences in the absolute values, which were expected as a consequence of the differences in oxygen levels, resulting of the differences in volume. The lower the volume, the less anaerobic environment is created. Similar behaviour would happen when scaling-up between lab and pilot scales or between pilot and industrial scales.
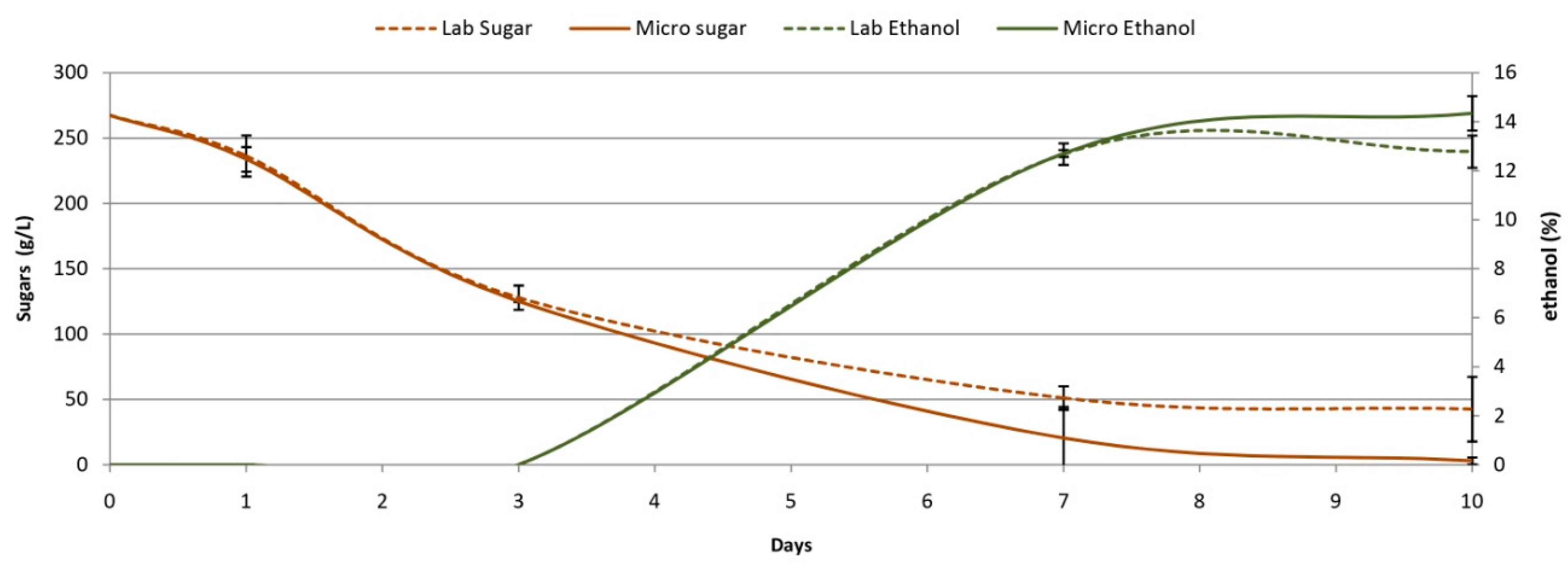
Sugar consumption and ethanol production (Figure 1) were similar up to day 3, and thereafter the microscale was marginally faster, resulting in a full sugar depletion at the end of fermentation. This is in line with the slightly higher biomass in the microwine system (Figure 2) and was according to [31], who observed accelerated sugar consumption in fermentations carried out in MTPs compared to those in Erlenmeyer flasks. However, these authors experienced lower ethanol levels in MTPs at the end of fermentation due to evaporation loss and despite being the same levels at early fermentation stages. In our study, this fact did not occur (Figure 1).
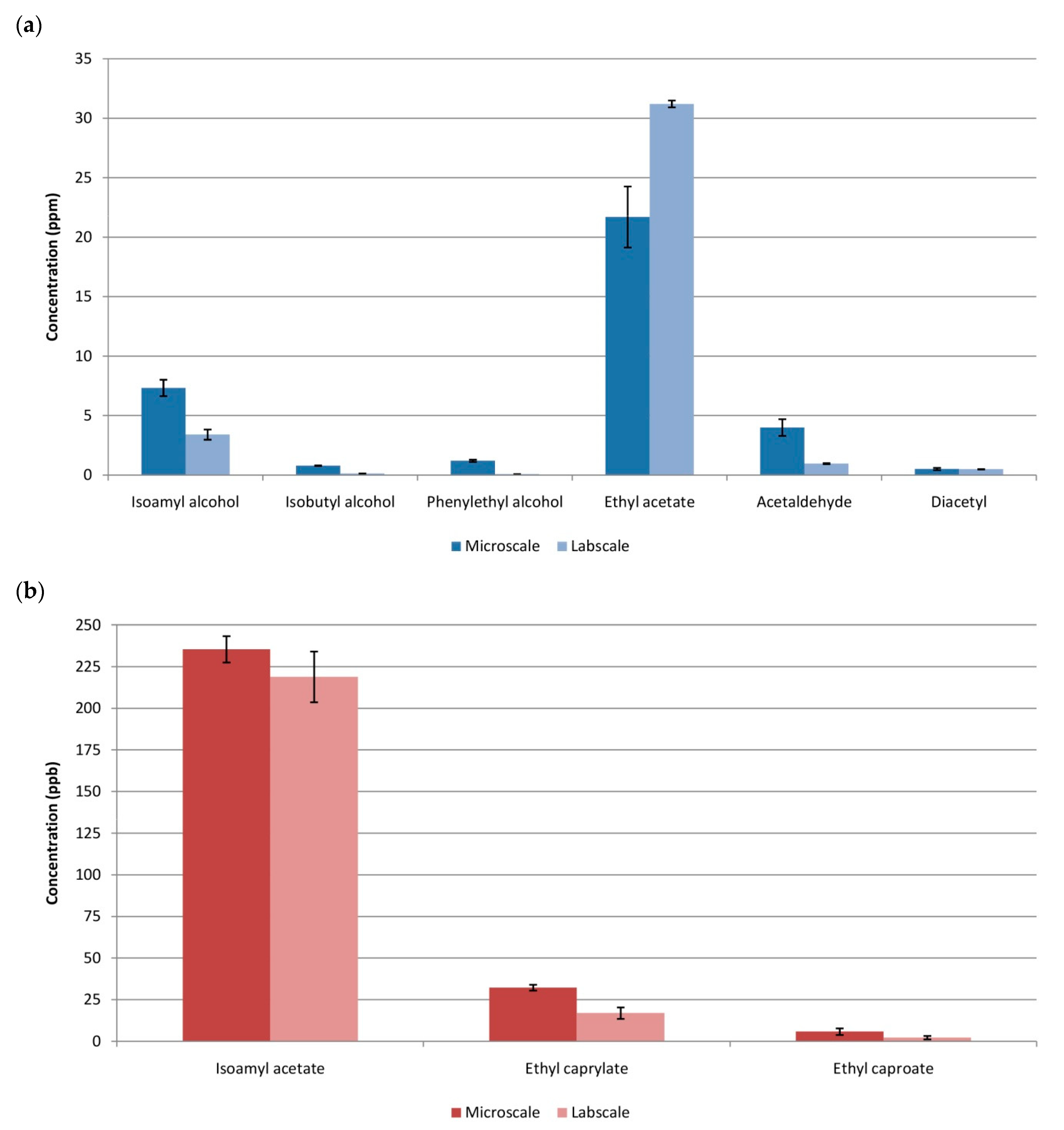
Most relevant levels of aroma compounds are presented in Figure 3. The significant differences that appeared in practically all the aroma compounds could also be explained by the levels of oxygen present in each scale. It has been demonstrated that differences in aeration causes differences in the yield of aroma compounds [32]. The initial oxygenation of the musts causes, in general, higher levels of alcohols and esters [32], which could explain the higher amounts of these aromas found in microscale compared to labscale. This was the case of isobutyl alcohol, isoamyl alcohol, phenylethanol, isoamyl acetate, ethyl caproate and ethyl caprylate. For instance, in the case of higher alcohols, these differences range from 4- to 20-fold [32]. One exception is ethyl acetate, which seems not to be affected by the levels of oxygen according to [32]. In our study, just slightly lower quantities of this compound were found in microscale. Regarding acetaldehyde and diacetyl, the former increased due to the higher presence of oxygen whereas the latter was not affected at all, being the same in both scales.
So the actual flavour levels in the microsystem were systematically higher than in the lab system. Besides oxygen, this matches with the slightly higher biomass levels and sugar consumption and does not compromise the screening power of the microsystem, although upscaling will always require fine tuning. Our results indicate that microsystems provide a useful and economical platform for the screening of interesting traits in industrial strains, as [31] affirmed, since the results are valid for comparison purposes. However, scaling-up will always be necessary in order to get closer to real winemaking or brewing processes.
4. Conclusions
Non-conventional yeasts show an enormous potential to improve and diversify aroma profiles of fermented beverages as wine and beer, especially regarding acetate ester yields and fruity aromas However, it has to be taken into account that mixed cultures of these species together to fermentative strains, Saccharomyces spp., is normally required to achieve successful fermentations. The adjustment of pitching ratio is crucial in these cases to avoid Saccharomyces spp. to take over and allow non-Saccharomyces yeasts to exploit all their aromatic potential.
Microsystems have proven their value as a tool for screening yeasts for key aromas in both wine and beer, giving the possibility of testing multiple combinations in a fast and simple way. However, scaling-up with the strains showing the highest potential will always be necessary in order to get closer to real winemaking or brewing outputs.
Author Contributions
Conceptualization, A.G., A.D., B.S. and C.d.J.; methodology, A.G., A.D., B.S. and C.d.J.; formal analysis, A.G.; investigation, A.G and A.D.; resources, C.d.J.; writing—original draft preparation, A.G.; writing—review & editing, A.G., A.D., B.S. and C.d.J.; visualization, A.G.; supervision, A.G. and C.d.J.; project administration, A.G. and C.d.J.; funding acquisition, A.G., C.d.J. All authors have read and agreed to the published version of the manuscript.
Funding
This research was founded by the European Comission—Marie Curie Initial Training Network CORNUCOPIA (FP7-PEOPLE-2010-ITN, grant agreement nr. 264717).
Acknowledgments
The authors thank the European Comission for the funding; Westerdijk Fungal Biodiversity Institute-CBS-KNAW for providing most of the strains; Amparo Querol (IATA-CSIC) for providing commercial wine strain and Carlsberg Research Laboratory for providing wort for microbeer experiments.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Aslankoohi, E.; Herrera-Malaver, B.; Rezaei, M.N.; Steensels, J.; Courtin, C.M.; Verstrepen, K.J. Non-Conventional Yeast Strains Increase the Aroma Complexity of Bread. PLoS ONE 2016, 11, e0165126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varela, C. The impact of non-Saccharomyces yeasts in the production of alcoholic beverages. Appl. Microbiol. Biotechnol. 2016, 100, 9861–9874. [Google Scholar] [CrossRef] [PubMed]
- Dashko, S.; Zhou, N.; Tinta, T.; Sivilotti, P.; Lemut, M.S.; Trost, K.; Gamero, A.; Boekhout, T.; Butinar, L.; Vrhovsek, U.; et al. Use of non-conventional yeast improves the wine aroma profile of Ribolla Gialla. J. Ind. Microbiol. Biotechnol. 2015, 42, 997–1010. [Google Scholar] [CrossRef] [PubMed]
- Jood, I.; Hoff, J.W.; Setati, M.E. Evaluating fermentation characteristics of Kazachstania spp. and their potential influence on wine quality. World J. Microbiol. Biotechnol. 2017, 33, 129. [Google Scholar] [CrossRef] [PubMed]
- Varela, C.; Sengler, F.; Solomon, M.; Curtin, C. Volatile flavour profile of reduced alcohol wines fermented with the non-conventional yeast species Metschnikowia pulcherrima and Saccharomyces uvarum. Food Chem. 2016, 209, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.-C.; Li, A.-H.; Dizy, M.; Ullah, N.; Sun, W.-X.; Tao, Y. Evaluation of aroma enhancement for “Ecolly” dry white wines by mixed inoculation of selected Rhodotorula mucilaginosa and Saccharomyces cerevisiae. Food Chem. 2017, 228, 550–559. [Google Scholar] [CrossRef]
- Capece, A.; Romaniello, R.; Siesto, G.; Romano, P. Conventional and Non-Conventional Yeasts in Beer Production. Fermentation 2018, 4, 38. [Google Scholar] [CrossRef] [Green Version]
- Michel, M.; Kopecká, J.; Meier-Dörnberg, T.; Zarnkow, M.; Jacob, F.; Hutzler, M. Screening for new brewing yeasts in the non-Saccharomyces sector with Torulaspora delbrueckii as model. Yeast 2016, 33, 129–144. [Google Scholar] [CrossRef] [Green Version]
- Rijswijck, I.M.H.; Rooijackers, J.C.M.W.; Abee, T.; Smid, E.J. Performance of non-conventional yeasts in co-culture with brewers’ yeast for steering ethanol and aroma production. Microb. Biotechnol. 2017, 10, 1591–1602. [Google Scholar] [CrossRef] [Green Version]
- Holt, S.; Mukherjee, V.; Lievens, B.; Verstrepen, K.J.; Thevelein, J.M. Bioflavoring by non-conventional yeasts in sequential beer fermentations. Food Microbiol. 2018, 72, 55–66. [Google Scholar] [CrossRef]
- Gutiérrez, A.; Boekhout, T.; Gojkovic, Z.; Katz, M. Evaluation of non-Saccharomyces yeasts in the fermentation of wine, beer and cider for the development of new beverages. J. Inst. Brew. 2018, 124, 389–402. [Google Scholar] [CrossRef]
- Ravasio, D.; Carlin, S.; Boekhout, T.; Groenewald, M.; Vrhovsek, U.; Walther, A.; Wendland, J. Adding Flavor to Beverages with Non-Conventional Yeasts. Fermentation 2018, 4, 15. [Google Scholar] [CrossRef] [Green Version]
- Swiegers, J.H.; Bartowsky, E.; Henschke, P.; Pretorius, I. Yeast and bacterial modulation of wine aroma and flavour. Aust. J. Grape Wine Res. 2005, 11, 139–173. [Google Scholar] [CrossRef]
- Michel, M.; Meier-Dörnberg, T.; Jacob, F.; Methner, F.J.; Wagner, S.; Hutzler, M. Review: Pure non-Saccharomyces starter cultures for beer fermentation with a focus on secondary metabolites and practical applications. J. Inst. Brew. 2016, 122, 569–587. [Google Scholar] [CrossRef]
- Šmogrovičová, D.; Dömény, Z. Beer volatile by-product formation at different fermentation temperature using immobilised yeasts. Process. Biochem. 1999, 34, 785–794. [Google Scholar] [CrossRef]
- Zhou, N.; Schifferdecker, A.J.; Gamero, A.; Compagno, C.; Boekhout, T.; Piškur, J.; Knecht, W. Kazachstania gamospora and Wickerhamomyces subpelliculosus: Two alternative baker’s yeasts in the modern bakery. Int. J. Food Microbiol. 2017, 250, 45–58. [Google Scholar] [CrossRef]
- Comitini, F.; Gobbi, M.; Domizio, P.; Romani, C.; Lencioni, L.; Mannazzu, I.; Ciani, M. Selected non-Saccharomyces wine yeasts in controlled multistarter fermentations with Saccharomyces cerevisiae. Food Microbiol. 2011, 28, 873–882. [Google Scholar] [CrossRef]
- Jolly, N.P.; Varela, C.; Pretorius, I.S. Not your ordinary yeast: Non-Saccharomyces yeasts in wine production uncovered. FEMS Yeast Res. 2013, 14, 215–237. [Google Scholar] [CrossRef] [Green Version]
- Minnaar, P.; Du Plessis, H.; Jolly, N.; Van Der Rijst, M.; Du Toit, M. Non-Saccharomyces yeast and lactic acid bacteria in co-inoculated fermentations with two Saccharomyces cerevisiae yeast strains: A strategy to improve the phenolic content of Syrah wine. Food Chem. 2019, 4, 100070. [Google Scholar] [CrossRef]
- Canonico, L.; Solomon, M.; Comitini, F.; Ciani, M.; Varela, C. Volatile profile of reduced alcohol wines fermented with selected non-Saccharomyces yeasts under different aeration conditions. Food Microbiol. 2019, 84, 103247. [Google Scholar] [CrossRef]
- Gamero, A.; Ren, X.; Lamboni, Y.; de Jong, C.; Smid, E.J.; Linnemann, A.R. Development of a low-alcoholic fermented beverage employing cashew apple Juice and non-conventional yeasts. Fermentation 2019, 5, 71. [Google Scholar] [CrossRef] [Green Version]
- Gamero, A.; Quintilla, R.; Groenewald, M.; Alkema, W.; Boekhout, T.; Hazelwood, L. High-throughput screening of a large collection of non-conventional yeasts reveals their potential for aroma formation in food fermentation. Food Microbiol. 2016, 60, 147–159. [Google Scholar] [CrossRef] [PubMed]
- Sabel, A.; Martens, S.; Petri, A.; König, H.; Claus, H. Wickerhamomyces anomalus AS1: A new strain with potential to improve wine aroma. Ann. Microbiol. 2013, 64, 483–491. [Google Scholar] [CrossRef]
- Gamero, A.; Tronchoni, J.; Querol, A.; Belloch, C. Production of aroma compounds by cryotolerant Saccharomyces species and hybrids at low and moderate fermentation temperatures. J. Appl. Microbiol. 2013, 114, 1405–1414. [Google Scholar] [CrossRef] [PubMed]
- Arroyo-Lopez, F.N.; Torrado, R.P.; Querol, A.; Barrio, E. Modulation of the glycerol and ethanol syntheses in the yeast Saccharomyces kudriavzevii differs from that exhibited by Saccharomyces cerevisiae and their hybrid. Food Microbiol. 2010, 27, 628–637. [Google Scholar] [CrossRef]
- Torrado, R.P.; Oliveira, B.M.; Zemančíková, J.; Sychrova, H.; Querol, A. Alternative Glycerol Balance Strategies among Saccharomyces Species in Response to Winemaking Stress. Front. Microbiol. 2016, 7, 628. [Google Scholar] [CrossRef] [Green Version]
- Erten, H.; Tangüler, H.; Cakiroz, H. The Effect of Pitching Rate on Fermentation and Flavour Compounds in High Gravity Brewing. J. Inst. Brew. 2007, 113, 75–79. [Google Scholar] [CrossRef]
- Verbelen, P.J.; Dekoninck, T.M.L.; Saerens, S.M.G.; Van Mulders, S.E.; Thevelein, J.M.; Delvaux, F.R. Impact of pitching rate on yeast fermentation performance and beer flavour. Appl. Microbiol. Biotechnol. 2009, 82, 155–167. [Google Scholar] [CrossRef]
- Bartowsky, E.J.; Henschke, P.A. The ‘buttery’ attribute of wine—Diacetyl—Desirability, spoilage and beyond. Int. J. Food Microbiol. 2004, 96, 235–252. [Google Scholar] [CrossRef]
- Yakobson, C. Pure Culture Fermentation Characteristics of Brettanomyces Yeast Species and Their Use in the Brewing Industry. Ph.D. Thesis, Heriot-Watt University, Edinburgh, UK, 2010. [Google Scholar]
- Liccioli, T.; Tran, T.M.T.; Cozzolino, D.; Jiranek, V.; Chambers, P.J.; Schmidt, S.A. Microvinification—How small can we go? Appl. Microbiol. Biotechnol. 2010, 89, 1621–1628. [Google Scholar] [CrossRef]
- Valero, E.; Moyano, L.; Millan, M.; Medina, M.; Ortega, J. Higher alcohols and esters production by Saccharomyces cerevisiae. Influence of the initial oxygenation of the grape must. Food Chem. 2002, 78, 57–61. [Google Scholar] [CrossRef]
Figure 1.
Fermentation performance in labscale wine and microwine.

Figure 2.
Yeast growth in labscale wine and microwine.

Figure 3.
Aroma profiles at microwine and labscale. (a) Major compounds; (b) Minor compounds.


{kind=link}
{kind=link}
{kind=link}
Table 1.
Non-Saccharomyces yeasts used in this study.
| Strain | Species | Substrate | Origin |
|---|---|---|---|
| CBS 772.71 | Galactomyces geotrichum | Soil | Puerto Rico |
| CBS 2567 | Hanseniaspora guilliermondii | Juice | Israel |
| CBS 5552 | Wickerhamomyces subpelliculosus | Sugar | Unknown |
| CBS 6641 | Torulaspora microellipsoides | Plant | USA |
| CBS 7692 | Starmera caribaea | Plant | West Indies |
| CBS 8031 | Hanseniaspora vineae | Plant | Canada |
| CBS 8849 | Zygosaccharomyces kombuchaensis | Fungus | Russia |
| CBS 8860 | Barnettozyma californica | Fruit | Russia |
| CBS 9716 | Zygosaccharomyces rouxii | Insect | Germany |
| CBS 10400 | Kazachstania gamospora | Unknown | Japan |
Table 2.
Sensory descriptors and thresholds of the aroma compounds investigated in the microfermentations.
Table 2.
Sensory descriptors and thresholds of the aroma compounds investigated in the microfermentations.
| Compound | Aroma | ST Wine * (mg/L) | ST Beer ** (mg/L) |
|---|---|---|---|
| Vicinal diketones | |||
| Diacetyl | buttery | - | 0.10–0.15 |
| 2,3-Pentanedione | toffee | - | 0.90 |
| Aldehydes | |||
| Acetaldehyde | herbaceous, sherry, nutty, bruised apple | 100 | 20–25 |
| Higheralcohols | |||
| Isobutyl alcohol | solvent, fusel, spiritous | 40 | 100 |
| Isoamyl alcohol | harsh, nail polish, banana | 30 | 50–65 |
| Phenylethyl alcohol | floral, rose, sweet | 10 | 40 |
| Acetateesters | |||
| Ethyl acetate | nail polish, fruity | 7.50 | 30–33 |
| Isoamyl acetate | banana, pear | 0.03 | 1.60–2.00 |
| Phenylethyl acetate | flowery, rose, fruity, honey | 0.25 | 3.80 |
| Ethyl Esters | |||
| Ethyl caproate | green apple, aniseed | 0.05 | 0.23 |
| Ethyl caprylate | sour apple, sweet soap | 0.02 | 0.50–0.90 |
| Ethyl caprate | apple, floral, soap | 0.20 | 1.50 |
Table 3.
Residual sugars and ethanol in the resulting microwines.
| Strain 1 | Strain 2 | Ratio | Sugars (g/L) | Ethanol% (v/v) |
|---|---|---|---|---|
| Lalvin T73 | 11 ± 4.4 f | 20 ± 0.0 b | ||
| CBS 772.71 | 163 ± 7.8 a–d | 13 ± 0.3 d | ||
| CBS 772.71 | Lalvin T73 | 1:1 | 63 ± 1.7 d–f | 17 ± 1.1 b,c |
| CBS 2567 | 170 ± 1.4 a–c | 11 ± 0.1 d,e | ||
| CBS 2567 | Lalvin T73 | 1:1 | 30 ± 1.3 e,f | 21 ± 0.8 a,b |
| CBS 5552 | 20 ± 2.1 f | 18 ± 1.1 b,c | ||
| CBS 5552 | Lalvin T73 | 1:1 | 32 ± 0.6 e,f | 18 ± 0.3 b,c |
| CBS 6641 | 157 ± 2.7 a–d | 12 ± 0.7 d | ||
| CBS 6641 | Lalvin T73 | 1:1 | 34 ± 1.4 e,f | 24 ± 0.2 a |
| CBS 7692 | 235 ± 20 a | nd | ||
| CBS 7692 | Lalvin T73 | 1:1 | 59 ± 7.3 c–f | 19 ± 0.5 b,c |
| CBS 8031 | 173 ± 5.7 a,b | 13 ± 0.3 d | ||
| CBS 8031 | Lalvin T73 | 1:1 | 73 ± 2.3 b–f | 18 ± 0.8 b,c |
| CBS 8849 | 135 ± 1.7 b–f | 13 ± 0.0 d | ||
| CBS 8849 | Lalvin T73 | 1:1 | 30 ± 3.2 e,f | 21 ± 1.2 a,b |
| CBS 8860 | 250 ± 6.2 a | nd | ||
| CBS 8860 | Lalvin T73 | 1:1 | 75 ± 5.4 b–f | 22 ± 0.3 a,b |
| CBS 9716 | 138 ± 24.3 a–e | 15 ± 1.0 c,d | ||
| CBS 9716 | Lalvin T73 | 1:1 | 78 ± 2.0 b–f | 22 ± 0.6 a,b |
| CBS 10400 | 170 ± 4.5 a,b | 11 ± 0.4 d,e | ||
| CBS 10400 | Lalvin T73 | 1:1 | 74 ± 2.0 b–f | 18 ± 0.1 b,c |
Letters indicate grouping according to ANOVA.
Table 4.
Aroma compounds (mg/L) in the resulting microwines.
| Strain 1 | Strain 2 | Ratio | Isobutyl Alcohol | Isoamyl Alcohol | Phenylethyl Alcohol | Ethyl acetate | Isoamyl Acetate | Phenylethyl Acetate | Ethyl Caproate | Ethyl Caprylate | Ethyl Caprate |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Lalvin T73 | 26 a–c | 24 a–c | 33i | 3 a | 1.54bc | 1.98a | 0.55a,b | 1.43e,f | 0.57c,d | ||
| CBS 772.71 | 12 a | 23 a–c | 17f,g | 4 a–c | 0.81ab | 23.79c | 0.12a,b | 0.22a,b | 0.26a–d | ||
| CBS 772.71 | Lalvin T73 | 1:1 | 78b–f | 30a–d | 3 a,b | nd | 0.09a | 2.49a | 0.87b | 0.29a,b | 0.42a–d |
| CBS 2567 | 18 a,b | 11 a | 10b–e | 11c,d | 1.55bc | 13.35b | 0.06a | 0.10a | 0.08a | ||
| CBS 2567 | Lalvin T73 | 1:1 | 126e–g | 21 a–c | 6 a–d | nd | nd | 1.41a | 0.53a,b | 0.21a,b | 0.30a–d |
| CBS 5552 | 13 a | 19 a–c | 22g,h | 10b–d | 0.44ab | 11.88b | 0.22a,b | 1.26d–f | 0.60d | ||
| CBS 5552 | Lalvin T73 | 1:1 | 82c–g | 24 a–d | 6 a–d | nd | nd | 1.36a | 0.71a,b | 0.21a,b | 0.25a–d |
| CBS 6641 | 10 a | 16 a,b | 19g,h | 3 a,b | 0.25a | 0.95a | 0.14a,b | 0.36a–c | 0.30a–d | ||
| CBS 6641 | Lalvin T73 | 1:1 | 141g | 39b–d | 3 a | nd | nd | 2.19a | 0.78a,b | 0.23a,b | 0.40a–d |
| CBS 7692 | 14 a | 12 a | 9 a–d | 20e | 4.87d | 28.53d | 0.25a,b | 0.75b–d | 0.11a,b | ||
| CBS 7692 | Lalvin T73 | 1:1 | 76b–e | 41c,d | 3 a | nd | nd | 1.99a | 0.79a,b | 0.37a–c | 0.52a–d |
| CBS 8031 | 11 a | 22 a–c | 20g,h | 4 a,b | 0.76ab | 26.24c,d | 0.11a,b | 0.14a | 0.29a–d | ||
| CBS 8031 | Lalvin T73 | 1:1 | 75b–e | 29 a–d | 17f,g | nd | nd | 1.72a | 0.88b | 0.29a,b | 0.54b–d |
| CBS 8849 | 18 a,b | 25a–d | 25 h | 5 a–c | 0.88ab | 28.76d | 0.10a,b | 0.11a | 0.12a–c | ||
| CBS 8849 | Lalvin T73 | 1:1 | 138f,g | 26 a–d | 5 a–d | nd | nd | 2.42a | 0.67a,b | 0.22a,b | 0.29a–d |
| CBS 8860 | 20 a,b | 14 a | 11c–f | 4 a,b | 2.25c | 1.53a | 0.29a,b | 0.60a–c | 0.23a–d | ||
| CBS 8860 | Lalvin T73 | 1:1 | 125e–g | 48d | 4 a–c | nd | nd | 2.35a | 0.88b | 0.44a–c | 0.53a–d |
| CBS 9716 | 29 a–d | 25 a–d | 11d–f | 2 a | 1.56b,c | 1.49a | 0.39a,b | 1.61f | 0.69d | ||
| CBS 9716 | Lalvin T73 | 1:1 | 87d–g | 33a–d | 3 a | nd | nd | 1.85a | 0.81a,b | 0.28a,b | 0.59d |
| CBS 10400 | 10 a | 14 a | 16e–g | 14d | 3.85d | 33.71 e | 0.21a,b | 0.89c–e | 0.41a–d | ||
| CBS 10400 | Lalvin T73 | 1:1 | 76b–e | 32a–d | 7 a–d | nd | 0.06a | 2.71a | 0.83a,b | 0.29a,b | 0.58c,d |
Results in bold, above their sensory thresholds; letters indicate grouping according to ANOVA; standard deviations were below 20% in all of the cases.
Table 5.
Residual sugars and ethanol in the resulting microbeers.
| Strain 1 | Strain 2 | Ratio | Sugars (g/L) | Ethanol% (v/v) |
|---|---|---|---|---|
| WS34/70 | nd d | 5.9 ± 0.6 a | ||
| CBS 772.71 | WS34/70 | 5:1 | nd d | 5.6 ± 1.4 a |
| CBS 772.71 | WS34/70 | 20:1 | nd d | 6.5 ± 0.4 a |
| CBS 2567 | WS34/70 | 5:1 | 5.4 ± 1.4 a–c | 6.2 ± 1.1 a |
| CBS 2567 | WS34/70 | 20:1 | 2.4 ± 0.7 c,d | 6.0 ± 1.4 a |
| CBS 5552 | WS34/70 | 5:1 | nd d | 6.4 ± 0.3 a |
| CBS 5552 | WS34/70 | 20:1 | nd d | 5.4 ± 1.3 a |
| CBS 6641 | WS34/70 | 5:1 | 2.2 ± 0.4 c,d | 6.0 ± 0.6 a |
| CBS 6641 | WS34/70 | 20:1 | 5.2 ± 1.0 a–c | 6.4 ± 0.5 a |
| CBS 7692 | WS34/70 | 5:1 | 0.9 ± 1.2 d | 6.8 ± 0.3 a |
| CBS 7692 | WS34/70 | 20:1 | 3.5 ± 1.9 b–d | 6.2 ± 1.4 a |
| CBS 8031 | WS34/70 | 5:1 | nd d | 6.9 ± 1.2 a |
| CBS 8031 | WS34/70 | 20:1 | 1.0 ± 1.4 d | 6.2 ± 0.6 a |
| CBS 8849 | WS34/70 | 5:1 | 7.1 ± 0.6 a,b | 5.5 ± 1.2 a |
| CBS 8849 | WS34/70 | 20:1 | nd d | 6.5 ± 0.6 a |
| CBS 8860 | WS34/70 | 5:1 | nd d | 6.0 ± 0.4 a |
| CBS 8860 | WS34/70 | 20:1 | 0.8 ± 1.1 d | 6.8 ± 0.1 a |
| CBS 9716 | WS34/70 | 5:1 | 7.7 ± 0.4 a | 5.7 ± 1.1 a |
| CBS 9716 | WS34/70 | 20:1 | 3.9 ± 1.6 a–d | 6.6 ± 0.3 a |
| CBS 10400 | WS34/70 | 5:1 | nd d | 7.3 ± 0.4 a |
| CBS 10400 | WS34/70 | 20:1 | 1.6 ± 0.5 c,d | 6.6 ± 0.4 a |
Letters indicate grouping according to ANOVA.
Table 6.
Aroma compounds (mg/L) in the resulting microbeers.
| Strain 1 | Strain 2 | Ratio | Diacetyl | 2,3-Pentadione | Isobutyl Alcohol | Isoamyl Alcohol | Phenylethyl Alcohol | Ethyl Acetate | Isoamyl Acetate | Phenylethyl Acetate | Ethyl Caproate | Ethyl Caprylate | Ethyl Caprate |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| WS34/70 | 0.14a | 0.02 b,c | 32 d | 74e | 27 d–f | 27 a | 0.52 a | 0.84 a–e | 0.04 d | 0.12 g | 0.06 e,f | ||
| CBS 772.71 | WS34/70 | 5:1 | 0.13a | 0.02 a,b | 29 a–d | 67a–e | 23 a–e | 23 a | 0.41 a | 0.86 a–e | 0.04 c,d | 0.11 f,g | 0.05 c–f |
| CBS 772.71 | WS34/70 | 20:1 | 0.41c | 0.10 h | 29 a–d | 68b–e | 20 a–c | 20 a | 0.37 a | 1.22 e–g | 0.03 a–d | 0.10 e–g | 0.04 b–e |
| CBS 2567 | WS34/70 | 5:1 | 0.65d | 0.06 f,g | 27 a–c | 62a–d | 19 a–c | 19 a | 0.35 a | 1.04 b–f | 0.03 a–c | 0.03 a–c | 0.02 a–d |
| CBS 2567 | WS34/70 | 20:1 | 0.62d | 0.05 e,f | 25 a | 60a,b | 18 a,b | 18 a | 0.38 a | 1.53 f,g | 0.02 a,b | 0.02 a | 0.01 a |
| CBS 5552 | WS34/70 | 5:1 | 0.30b | 0.04 d | 27 a–c | 64a–d | 34 g,h | 34a | 0.45 a | 0.59 a,b | 0.03 b–d | 0.09 d–g | 0.02 a–d |
| CBS 5552 | WS34/70 | 20:1 | 0.08 a | 0.01 a,b | 30 b–d | 63a–d | 34 g,h | 34a | 0.33 a | 1.68 g | 0.03 a–c | 0.04 a–d | 0.01 a,b |
| CBS 6641 | WS34/70 | 5:1 | 0.09 a | 0.01 a,b | 28 a–d | 67a–e | 16 a | 16 a | 0.62 a | 0.61 a–c | 0.04 c,d | 0.12 g,h | 0.05 d–f |
| CBS 6641 | WS34/70 | 20:1 | 0.33b,c | 0.03 d | 28 a–d | 66a–e | 19 a–c | 19 a | 0.47 a | 1.20 d–g | 0.03 a–c | 0.03 a–c | 0.01 a,b |
| CBS 7692 | WS34/70 | 5:1 | 0.41c | 0.04 d,e | 25 a,b | 62a–d | 25 b–e | 25 a | 0.36 a | 1.01 b–e | 0.03 a–d | 0.09 e–g | 0.03 a–e |
| CBS 7692 | WS34/70 | 20:1 | 0.60d | 0.07 g | 27 a–c | 61a–c | 19 a,b | 19 a | 0.53 a | 1.12 c–f | 0.03 a–d | 0.04 a–c | 0.02 a–d |
| CBS 8031 | WS34/70 | 5:1 | 0.08 a | 0.02 a,b | 26 a–c | 60a,b | 29 e–g | 29 a | 0.43 a | 2.25 h | 0.03 a–c | 0.02 ab | 0.01 abc |
| CBS 8031 | WS34/70 | 20:1 | 0.30b | 0.03 c,d | 26 a–c | 60a,b | 32 f–h | 32a | 0.23 a | 3.03 i | 0.02 a | 0.01 a | 0.01 a,b |
| CBS 8849 | WS34/70 | 5:1 | 0.08 a | 0.02 a,b | 30 b–d | 68b–e | 27 c–f | 27 a | 0.44 a | 0.72 a–e | 0.04 c,d | 0.10 e–g | 0.04 b–e |
| CBS 8849 | WS34/70 | 20:1 | 0.56d | 0.05 f | 29 a–d | 69c–e | 22 a–d | 22 a | 0.31 a | 0.43 a | 0.03 a–d | 0.04 a–c | 0.01 a,b |
| CBS 8860 | WS34/70 | 5:1 | 0.27b | 0.03 d | 29 a–d | 69c–e | 26 c–f | 26 a | 0.37 a | 0.74 a–e | 0.04 d | 0.16 h | 0.07 f |
| CBS 8860 | WS34/70 | 20:1 | 0.42c | 0.11 h | 31 c,d | 68b–e | 24 b–e | 24 a | 0.43 a | 0.80 a–e | 0.03 b–d | 0.10 e–g | 0.06 e,f |
| CBS 9716 | WS34/70 | 5:1 | 0.05 a | 0.01 a,b | 29 a–d | 70d,e | 35 g,h | 35a | 0.40 a | 0.60 a,b | 0.04 c,d | 0.10 e–g | 0.02 a–d |
| CBS 9716 | WS34/70 | 20:1 | 0.10a | 0.02 a,b | 28 a–d | 69b–e | 38 h | 38a | 0.38 a | 0.66 a–d | 0.03 a–d | 0.07 b–e | 0.01 a,b |
| CBS 10400 | WS34/70 | 5:1 | 0.10a | 0.02 a,b | 27 a–d | 61a–d | 20 a–c | 20 a | 0.24 a | 1.06 b–f | 0.03 a–d | 0.06 b–e | 0.05 d–f |
| CBS 10400 | WS34/70 | 20:1 | 0.05 a | 0.01 a | 25 a,b | 59a | 25 b–f | 25 a | 0.28 a | 1.66 g | 0.03 a–d | 0.07 c–f | 0.12 g |
Results in bold, above sensory thresholds; letters indicate grouping according to ANOVA; standard deviations were below 20% in all of the cases.
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Gamero, A.; Dijkstra, A.; Smit, B.; de Jong, C. Aromatic Potential of Diverse Non-Conventional Yeast Species for Winemaking and Brewing. Fermentation 2020, 6, 50. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation6020050
AMA Style
Gamero A, Dijkstra A, Smit B, de Jong C. Aromatic Potential of Diverse Non-Conventional Yeast Species for Winemaking and Brewing. Fermentation. 2020; 6(2):50. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation6020050
Chicago/Turabian StyleGamero, Amparo, Annereinou Dijkstra, Bart Smit, and Catrienus de Jong. 2020. "Aromatic Potential of Diverse Non-Conventional Yeast Species for Winemaking and Brewing" Fermentation 6, no. 2: 50. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation6020050
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.