Saccharomyces cerevisiae Strain Diversity Associated with Spontaneous Fermentations in Organic Wineries from Galicia (NW Spain)


Abstract
:1. Introduction
2. Materials and Methods
2.1. Wineries and Sampling
2.2. Sample Processing and Yeast Isolation
2.3. Identification of Saccharomyces cerevisiae Yeast Strains
2.4. Determination of Yeast Killer Activity
2.5. Data Analysis
3. Results
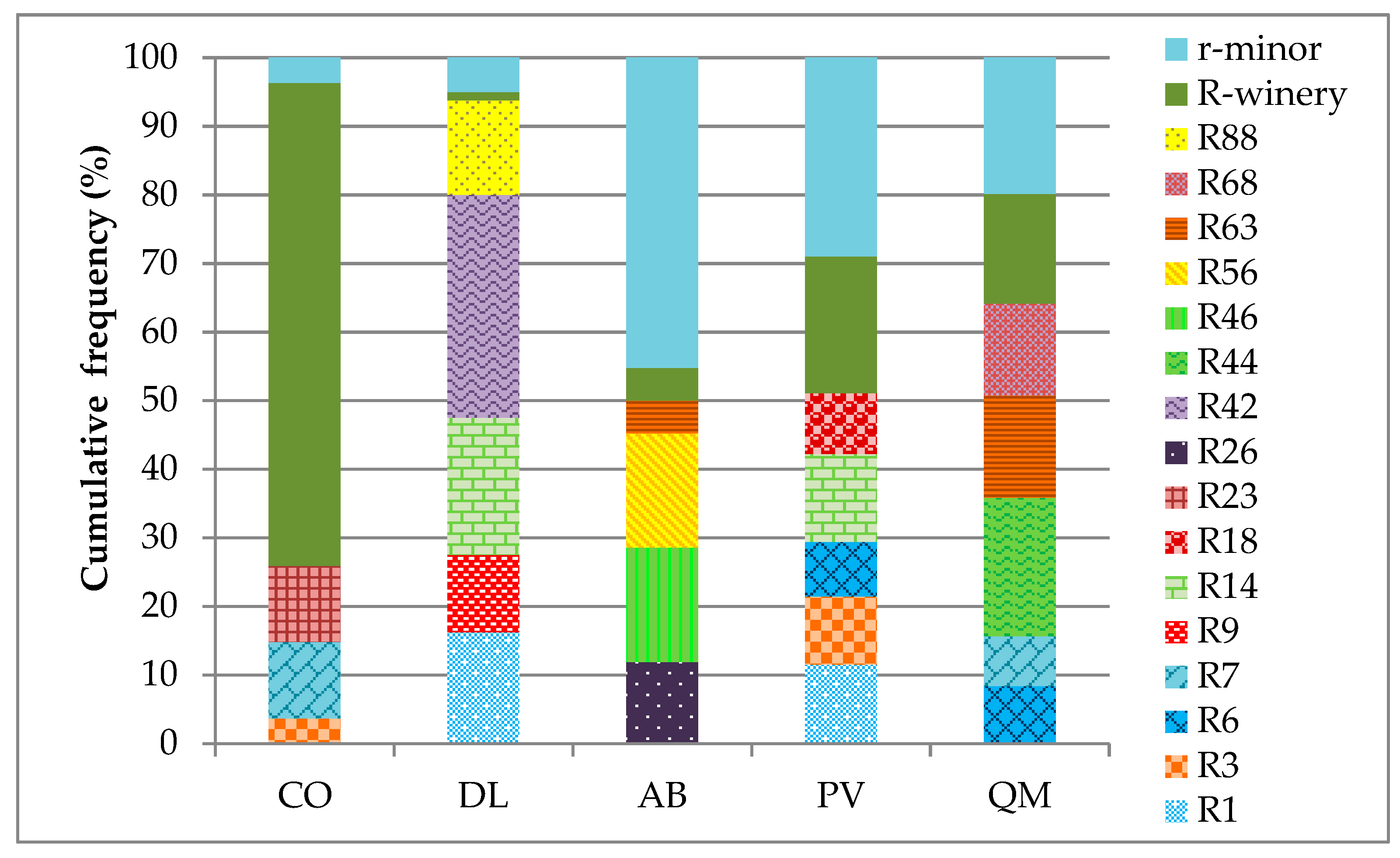
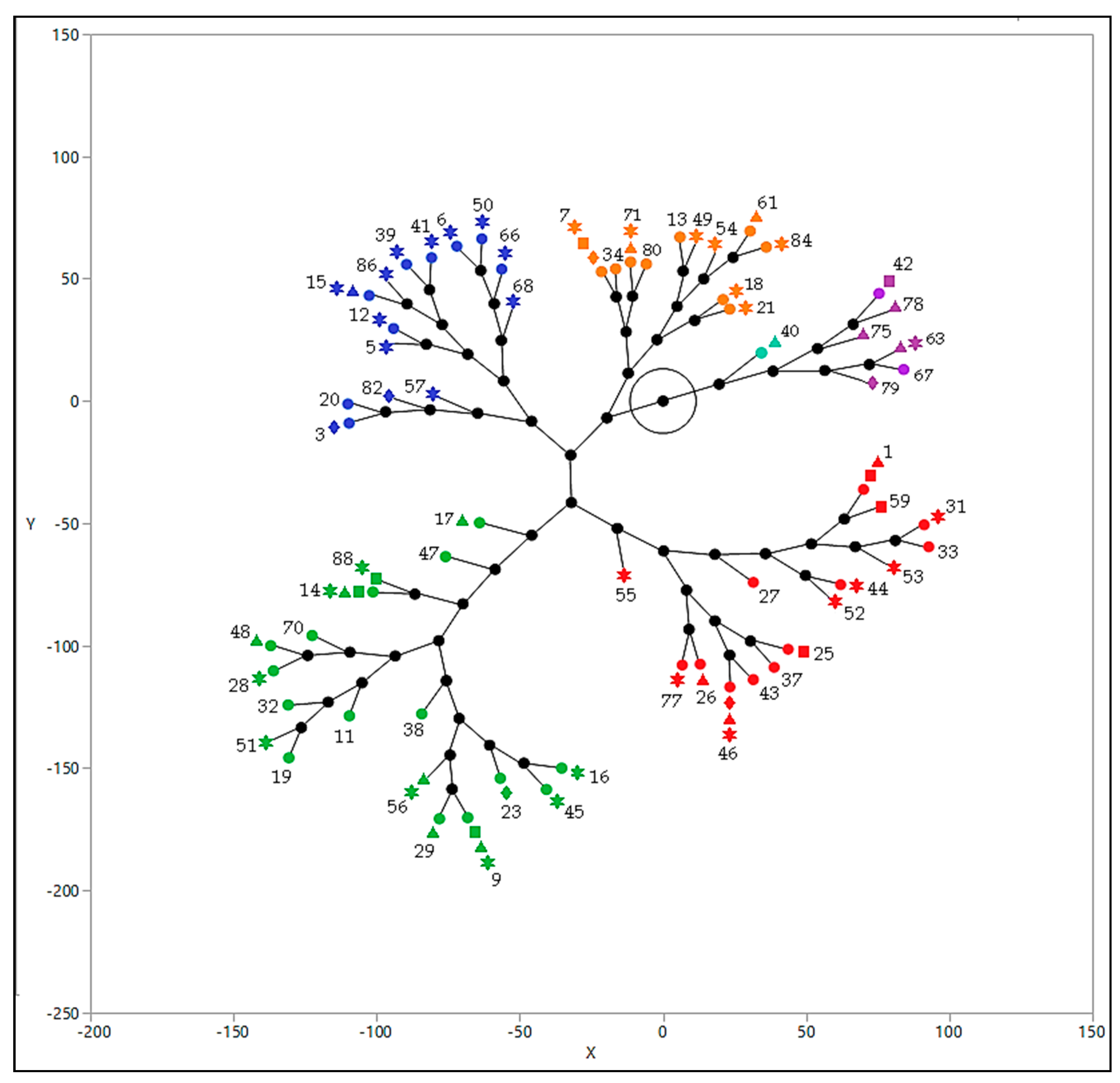
3.1. Diversity of S. cerevisiae Strains in Galician Organic Wineries
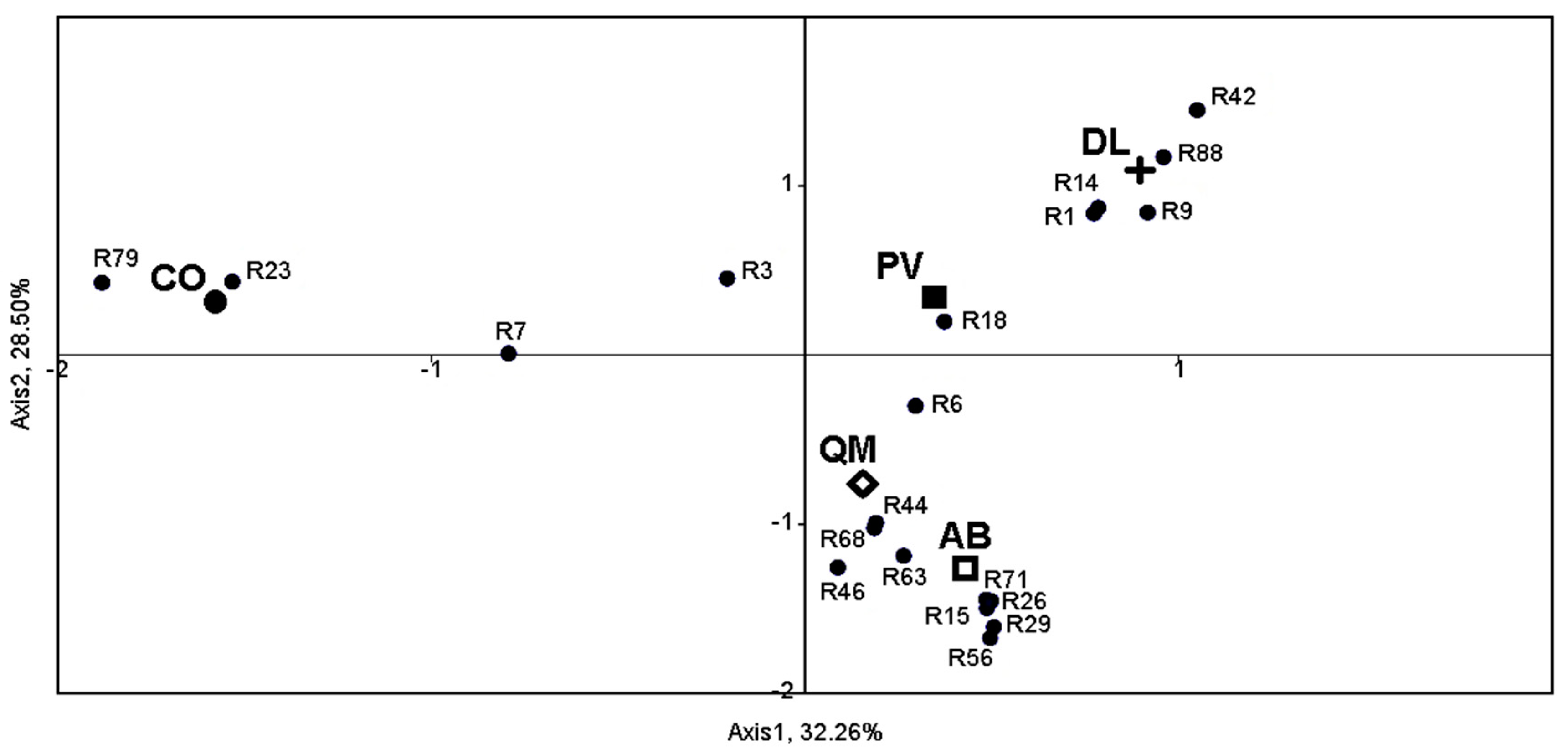
3.2. Occurrence of S. cerevisiae Strains in Different Wineries
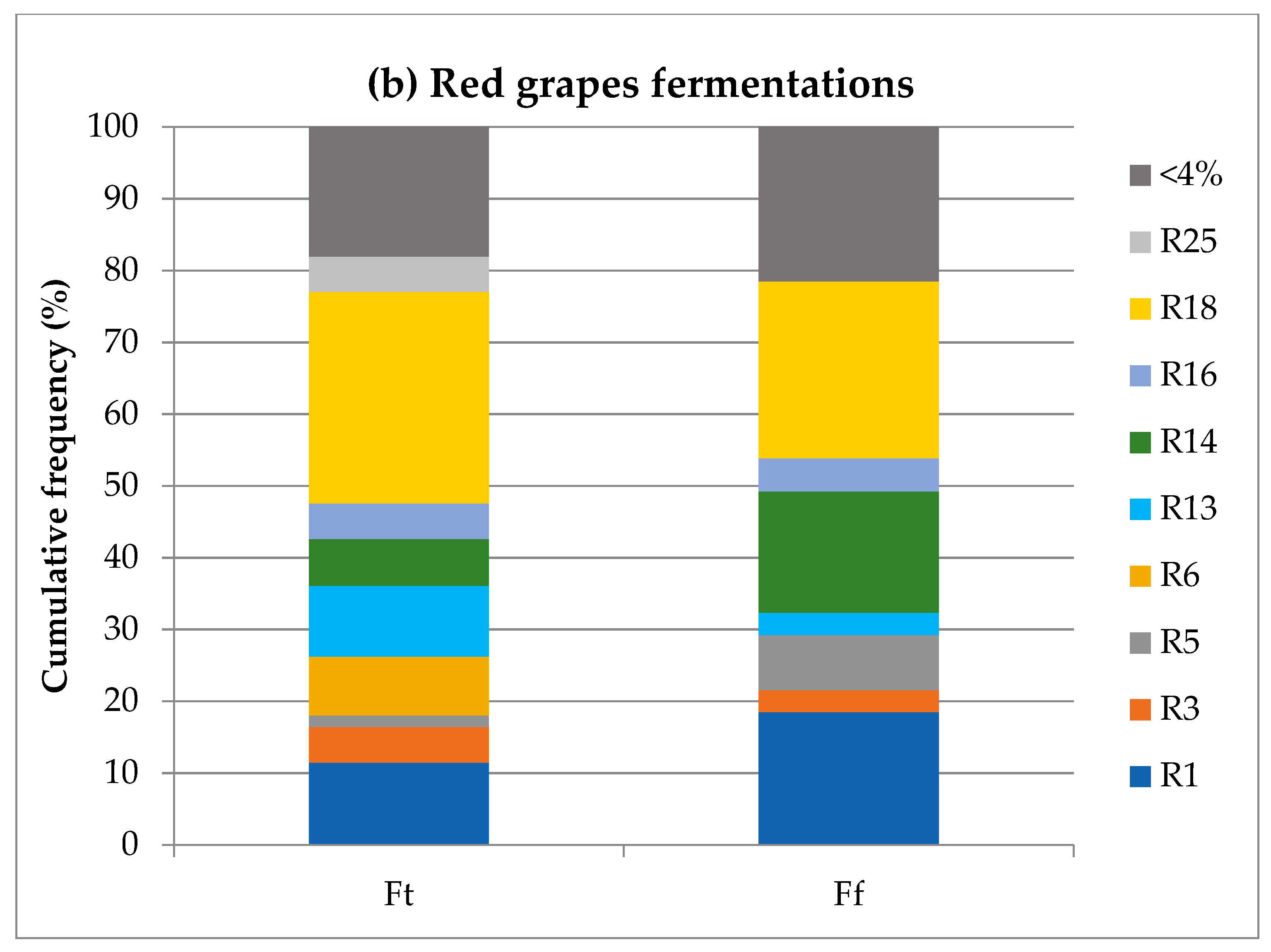
3.3. Incidence of S. cerevisiae Strains per Campaign
3.4. S. cerevisiae Population Dynamics during Fermentation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- MAPA (Ministerio de Agricultura y Pesca). Agricultura Ecológica, Estadísticas 2018; MAPA: Madrid, Spain, 2019; pp. 1–175.
- Willer, H.; Lernoud, J. The World of Organic Agriculture. Statistics and Emerging Trends 2018. Fibl, Fricks, and Ifoam-Organics International; Research Institute of Organic Agriculture (FiBL): Frick, Switzerland; IFOAM – Organics International: Bonn, Germany, 2018; ISBN 9783037360675. [Google Scholar]
- Council of the European Union Commission regulation (EC). No 889/2008 of 5 September 2008 laying down detailed rules for the implementation of Council Regulation (EC) No 834/2007 on organic production and labelling of organic products with regard to organic production, labelling and control. Off. J. Eur. Union 2008, 250, 1–84. [Google Scholar]
- Council of the European Union Council Regulation (EC). No 834/2007 of 28 June 2007 on organic production and labelling of organic products and repealing Regulation (EEC) No 2092/91. Off. J. Eur. Union 2007, 189, 1–23. [Google Scholar]
- Council of the European Union Commission regulation (EC). No 203/2012 of 8 March 2012 determining rules on organic wine production. Off. J. Eur. Union 2012, 71, 42–47. [Google Scholar]
- Blanco, P.; Mirás-Avalos, J.M.; Suárez, V.; Orriols, I. Inoculation of treixadura musts with autochthonous Saccharomyces cerevisiae strains: Fermentative performance and influence on the wine characteristics. Food Sci. Technol. Int. 2013, 19, 177–186. [Google Scholar] [CrossRef]
- Tello, J.; Cordero-Bueso, G.; Aporta, I.; Cabellos, J.M.; Arroyo, T. Genetic diversity in commercial wineries: Effects of the farming system and vinification management on wine yeasts. J. Appl. Microbiol. 2011, 112, 302–315. [Google Scholar] [CrossRef]
- Martiniuk, J.T.; Pacheco, B.; Russell, G.; Tong, S.; Backstrom, I.; Measday, V. Impact of commercial strain use on saccharomyces cerevisiae population structure and dynamics in pinot noir vineyards and spontaneous fermentations of a Canadian winery. PLoS ONE 2016, 11, e0160259. [Google Scholar] [CrossRef] [Green Version]
- Guzzon, R.; Widmann, G.; Settanni, L.; Malacarne, M.; Francesca, N.; Larcher, R. Evolution of yeast populations during different biodynamic winemaking processes. S. Afr. J. Enol. Vitic. 2011, 32, 242–250. [Google Scholar] [CrossRef] [Green Version]
- Cocolin, L.; Heisey, A.; Mills, D.A. Direct identification of the indigenous yeasts in commercial wine fermentations. Am. J. Enol. Vitic. 2001, 52, 49–53. [Google Scholar]
- Fleet, G.H. Wine yeasts for the future. FEMS Yeast Res. 2008, 8, 979–995. [Google Scholar] [CrossRef] [Green Version]
- Tofalo, R.; Perpetuini, G.; Schirone, M.; Fasoli, G.; Aguzzi, I.; Corsetti, A.; Suzzi, G. Biogeographical characterization of Saccharomyces cerevisiae wine yeast by molecular methods. Front. Microbiol. 2013, 4, 166. [Google Scholar] [CrossRef] [Green Version]
- Capece, A.; Romaniello, R.; Siesto, G.; Romano, P. Diversity of saccharomyces cerevisiae yeasts associated to spontaneously fermenting grapes from an Italian “heroic vine-growing area”. Food Microbiol. 2012, 31, 159–166. [Google Scholar] [CrossRef] [PubMed]
- Lambrechts, M.G.; Pretorius, I.S. Yeast and its importance to wine aromaA review. S. Afr. J. Enol. Vitic. 2000, 21, 97–129. [Google Scholar]
- Fleet, G.H. Yeast interactions and wine flavour. Int. J. Food Microbiol. 2003, 86, 11–22. [Google Scholar] [CrossRef]
- Romano, P.; Fiore, C.; Paraggio, M.; Caruso, M.; Capece, A. Function of yeast species and strains in wine flavour. Int. J. Food Microbiol. 2003, 86, 169–180. [Google Scholar] [CrossRef]
- Sannino, C.; Francesca, N.; Corona, O.; Settanni, L.; Cruciata, M.; Moschetti, G. Effect of the natural winemaking process applied at industrial level on the microbiological and chemical characteristics of wine. J. Biosci. Bioeng. 2013, 116, 347–356. [Google Scholar] [CrossRef] [Green Version]
- Tufariello, M.; Chiriatti, M.A.; Grieco, F.; Perrotta, C.; Capone, S.; Rampino, P.; Tristezza, M.; Mita, G.; Grieco, F. Influence of autochthonous Saccharomyces cerevisiae strains on volatile profile of Negroamaro wines. LWT Food Sci. Technol. 2014, 58, 35–48. [Google Scholar] [CrossRef]
- Tofalo, R.; Patrignani, F.; Lanciotti, R.; Perpetuini, G.; Schirone, M.; Di Gianvito, P.; Pizzoni, D.; Arfelli, G.; Suzzi, G. Aroma profile of montepulciano d’abruzzo wine fermented by single and co-culture starters of autochthonous Saccharomyces and non-Saccharomyces yeasts. Front. Microbiol. 2016, 7, 610. [Google Scholar] [CrossRef]
- Nisiotou, A.; Sgouros, G.; Mallouchos, A.; Nisiotis, C.S.; Michaelidis, C.; Tassou, C.; Banilas, G. The use of indigenous Saccharomyces cerevisiae and Starmerella bacillaris strains as a tool to create chemical complexity in local wines. Food Res. Int. 2018, 111, 498–508. [Google Scholar] [CrossRef]
- Drumonde-Neves, J.; Franco-Duarte, R.; Vieira, E.; Mendes, I.; Lima, T.; Schuller, D.; Pais, C. Differentiation of saccharomyces cerevisiae populations from vineyards of the azores archipelago: Geography vs Ecology. Food Microbiol. 2018, 74, 151–162. [Google Scholar] [CrossRef]
- Goddard, M.R.; Anfang, N.; Tang, R.; Gardner, R.C.; Jun, C. A distinct population of Saccharomyces cerevisiae in New Zealand: Evidence for local dispersal by insects and human-aided global dispersal in oak barrels. Environ. Microbiol. 2010, 12, 63–73. [Google Scholar] [CrossRef]
- Beltran, G.; Torija, M.J.; Novo, M.; Ferrer, N.; Poblet, M.; Guillamón, J.M.; Rozs, N.; Mas, A. Analysis of yeast populations during alcoholic fermentation: A six year follow-up study. Syst. Appl. Microbiol. 2002, 25, 287–293. [Google Scholar] [CrossRef]
- Santamaría, P.; Garijo, P.; López, R.; Tenorio, C.; Gutiérrez, A.R. Analysis of yeast population during spontaneous alcoholic fermentation: Effect of the age of the cellar and the practice of inoculation. Int. J. Food Microbiol. 2005, 103, 49–56. [Google Scholar] [CrossRef]
- Scholl, C.M.; Morgan, S.C.; Stone, M.L.; Tantikachornkiat, M.; Neuner, M.; Durall, D.M. Composition of Saccharomyces cerevisiae strains in spontaneous fermentations of Pinot Noir and Chardonnay. Aust. J. Grape Wine Res. 2016, 22, 384–390. [Google Scholar] [CrossRef]
- Morgan, S.C.; Scholl, C.M.; Benson, N.L.; Stone, M.L.; Durall, D.M. Sulfur dioxide addition at crush alters Saccharomyces cerevisiae strain composition in spontaneous fermentations at two Canadian wineries. Int. J. Food Microbiol. 2017, 244, 96–102. [Google Scholar] [CrossRef] [PubMed]
- Blanco, P.; Orriols, I.; Losada, A. Survival of commercial yeasts in the winery environment and their prevalence during spontaneous fermentations. J. Ind. Microbiol. Biotechnol. 2011, 38, 235–239. [Google Scholar] [CrossRef] [PubMed]
- Hall, B.; Durall, D.M.; Stanley, G. Population dynamics of Saccharomyces cerevisiae during spontaneous fermentation at a British Columbia winery. Am. J. Enol. Vitic. 2011, 62, 66–72. [Google Scholar] [CrossRef]
- Lange, J.N.; Faasse, E.; Tantikachornkiat, M.; Gustafsson, F.S.; Halvorsen, L.C.; Kluftinger, A.; Ledderhof, D.; Durall, D.M. Implantation and persistence of yeast inoculum in Pinot noir fermentations at three Canadian wineries. Int. J. Food Microbiol. 2014, 180, 56–61. [Google Scholar] [CrossRef]
- de Celis, M.; Ruiz, J.; Martín-Santamaría, M.; Alonso, A.; Marquina, D.; Navascués, E.; Gómez-Flechoso, M.; Belda, I.; Santos, A. Diversity of Saccharomyces cerevisiae yeasts associated to spontaneous and inoculated fermenting grapes from Spanish vineyards. Lett. Appl. Microbiol. 2019, 68, 580–588. [Google Scholar] [CrossRef]
- Schuller, D.; Alves, H.; Dequin, S.; Casal, M. Ecological survey of Saccharomyces cerevisiae strains from vineyards in the Vinho Verde Region of Portugal. FEMS Microbiol. Ecol. 2005, 51, 167–177. [Google Scholar] [CrossRef] [Green Version]
- Schuller, D.; Cardoso, F.; Sousa, S.; Gomes, P.; Gomes, A.C.; Santos, M.A.S.; Casal, M. Genetic diversity and population structure of Saccharomyces cerevisiae strains isolated from different grape varieties and winemaking regions. PLoS ONE 2012, 7, e32507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Maio, S.; Polizzotto, G.; Di Gangi, E.; Foresta, G.; Genna, G.; Verzera, A.; Scacco, A.; Amore, G.; Oliva, D. Biodiversity of indigenous Saccharomyces populations from old wineries of South-Eastern Sicily (Italy): Preservation and economic potential. PLoS ONE 2012, 7, e30428. [Google Scholar] [CrossRef]
- Viel, A.; Legras, J.L.; Nadai, C.; Carlot, M.; Lombardi, A.; Crespan, M.; Migliaro, D.; Giacomini, A.; Corich, V. The geographic distribution of Saccharomyces cerevisiae isolates within three Italian neighboring winemaking regions reveals strong differences in yeast abundance, genetic diversity and industrial strain dissemination. Front. Microbiol. 2017, 8, 1595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valero, E.; Cambon, B.; Schuller, D.; Casal, M.; Dequin, S. Biodiversity of saccharomyces yeast strains from grape berries of wine-producing areas using starter commercial yeasts. FEMS Yeast Res. 2007, 7, 317–329. [Google Scholar] [CrossRef] [Green Version]
- Versavaud, A.; Courcoux, P.; Roulland, C.; Dulau, L.; Hallet, J.N. Genetic diversity and geographical distribution of wild Saccharomyces cerevisiae strains from the wine-producing area of Charentes, France. Appl. Environ. Microbiol. 1995, 61, 3521–3529. [Google Scholar] [CrossRef] [Green Version]
- Gayevskiy, V.; Goddard, M.R. Geographic delineations of yeast communities and populations associated with vines and wines in New Zealand. ISME J. 2012, 6, 1281–1290. [Google Scholar] [CrossRef] [Green Version]
- Knight, S.; Goddard, M.R. Quantifying separation and similarity in a Saccharomyces cerevisiae metapopulation. ISME J. 2015, 9, 361–370. [Google Scholar] [CrossRef] [Green Version]
- Morrison-Whittle, P.; Goddard, M.R. From vineyard to winery: A source map of microbial diversity driving wine fermentation. Environ. Microbiol. 2018, 20, 75–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bokulich, N.A.; Thorngate, J.H.; Richardson, P.M.; Mills, D.A. Microbial biogeography of wine grapes is conditioned by cultivar, vintage, and climate. Proc. Natl. Acad. Sci. USA 2014, 111, E139–E148. [Google Scholar] [CrossRef] [Green Version]
- Bokulich, N.; Collins, T.; Masarweh, C.; Allen, G.; Heymann, H.; Ebeler, S.E.; Mills, D.A. Associations among wine grape microbiome, metabolome, and fermentation behavior suggest microbial contribution to regional wine characteristics. MBio 2016, 7, e00631-16. [Google Scholar] [CrossRef] [Green Version]
- Pinto, C.; Pinho, D.; Cardoso, R.; Custódio, V.; Fernandes, J.; Sousa, S.; Pinheiro, M.; Egas, C.; Gomes, A.C. Wine fermentation microbiome: A landscape from different Portuguese wine appellations. Front. Microbiol. 2015, 6, 905. [Google Scholar] [CrossRef] [Green Version]
- CRAEGA Estadísticas. Memoria Anual de Actividades do ano 2018 do Consello Regulador da Agricultura Ecolóxica de Galicia; CRAEGA: Lugo, Spain, 2019; pp. 1–16. [Google Scholar]
- Longo, E.; Cansado, J.; Agrelo, D.; Villa, T.G. Effect of climatic conditions on yeast diversity in grape musts from Northwest Spain. Am. J. Enol. Vitic. 1991, 42, 141–144. [Google Scholar]
- Lema, C.; Garcia-Jares, C.; Orriols, I.; Angulo, L. Contribution of Saccharomyces and non-Saccharomyces populations to the production of some components of Albariño wine aroma. Am. J. Enol. Vitic. 1996, 47, 206–216. [Google Scholar]
- Blanco, P.; Ramilo, A.; Cerdeira, M.; Orriols, I. Genetic diversity of wine Saccharomyces cerevisiae strains in an experimental winery from Galicia (NW Spain). Antonie Van Leeuwenhoek Int. J. Gen. Mol. Microbiol. 2006, 89, 351–357. [Google Scholar] [CrossRef]
- Castrillo, D.; Rabuñal, E.; Neira, N.; Blanco, P. Yeast diversity on grapes from Galicia, NW Spain: Biogeographical patterns and the influence of the farming system. Oeno One 2019, 53, 573–587. [Google Scholar] [CrossRef]
- Pallmann, C.L.; Brown, J.A.; Olineka, T.L.; Cocolin, L.; Mills, D.A.; Bisson, L.F. Use of WL medium to profile native flora fermentations. Am. J. Enol. Vitic. 2001, 52, 198–203. [Google Scholar]
- Heard, G.M.; Fleet, G.H. Evaluation of selective media for enumeration of yeasts during wine fermentation. J. Appl. Bacteriol. 1986, 60, 477–481. [Google Scholar] [CrossRef]
- Querol, A.; Barrio, E.; Huerta, T.; Ramon, D. Molecular monitoring of wine fermentations conducted by active dry yeast strains. Appl. Environ. Microbiol. 1992, 58, 2948–2953. [Google Scholar] [CrossRef] [Green Version]
- Maqueda, M.; Zamora, E.; Álvarez, M.L.; Ramírez, M. Characterization, ecological distribution, and population dynamics of saccharomyces sensu stricto killer yeasts in the spontaneous grape must fermentations of Southwestern Spain. Appl. Environ. Microbiol. 2012, 78, 735–743. [Google Scholar] [CrossRef] [Green Version]
- Shannon, C.E. A mathematical theory of communication. SIGMOBILE Mob. Comput. Commun. Rev. 2001, 5, 3–55. [Google Scholar] [CrossRef]
- Simpson, E.H. Measurement of diversity. Nature 1949, 163, 688. [Google Scholar] [CrossRef]
- Pielou, E.C. An Introduction to Mathematical Ecology; Wiley: New York, NY, USA, 1969. [Google Scholar]
- Anderson, M.J.; Walsh, D.C.I. PERMANOVA, ANOSIM, and the Mantel test in the face of heterogeneous dispersions: What null hypothesis are you testing? Published by: Ecological Society of America linked references are available on JSTOR for this article: Your use of the JSTOR archi. Ecol. Monogr. 2013, 83, 557–574. [Google Scholar] [CrossRef]
- Clarke, K.R. Non-parametric multivariate analyses of changes in community structure. Aust. J. Ecol. 1993, 18, 117–143. [Google Scholar] [CrossRef]
- Anderson, M.J. A new method for non-parametric multivariate analysis of variance. Austral Ecol. 2001, 26, 32–46. [Google Scholar]
- Torija, M.J.; Rozès, N.; Poblet, M.; Guillamón, J.M.; Mas, A. Yeast population dynamics in spontaneous fermentations: Comparison between two different wine-producing areas over a period of three years. Antonie Van Leeuwenhoek Int. J. Gen. Mol. Microbiol. 2001, 79, 345–352. [Google Scholar] [CrossRef]
- Cordero-Bueso, G.; Arroyo, T.; Serrano, A.; Valero, E. Influence of different floor management strategies of the vineyard on the natural yeast population associated with grape berries. Int. J. Food Microbiol. 2011, 148, 23–29. [Google Scholar] [CrossRef]
- Börlin, M.; Venet, P.; Claisse, O.; Salin, F.; Legras, J.L.; Masneuf-Pomarede, I. Cellar-associated Saccharomyces cerevisiae population structure revealed high-level diversity and perennial persistence at Sauternes wine estates. Appl. Environ. Microbiol. 2016, 82, 2909–2918. [Google Scholar] [CrossRef] [Green Version]
- Santamaría, P.; López, R.; López, E.; Garijo, P.; Gutiérrez, A.R. Permanence of yeast inocula in the winery ecosystem and presence in spontaneous fermentations. Eur. Food Res. Technol. 2008, 227, 1563–1567. [Google Scholar] [CrossRef]
- Zhang, H.Y.; Lee, S.A.; Bradbury, J.E.; Warren, R.N.; Sheth, H.; Hooks, D.O.; Richards, K.D.; Gardner, R.C. Yeasts isolated from New Zealand vineyards and wineries. Aust. J. Grape Wine Res. 2010, 16, 491–496. [Google Scholar] [CrossRef]
- Blanco, P.; Mirás-Avalos, J.M.; Orriols, I. Effect of must characteristics on the diversity of Saccharomyces strains and their prevalence in spontaneous fermentations. J. Appl. Microbiol. 2012, 112, 936–944. [Google Scholar] [CrossRef]
- Ciani, M.; Capece, A.; Comitini, F.; Canonico, L.; Siesto, G.; Romano, P. Yeast interactions in inoculated wine fermentation. Front. Microbiol. 2016, 7, 555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pérez, F.; Ramírez, M.; Regodón, J.A. Influence of killer strains of Saccharomyces cerevisiae on wine fermentation. Antonie Van Leeuwenhoek Int. J. Gen. Mol. Microbiol. 2001, 79, 393–399. [Google Scholar] [CrossRef]
- Cocolin, L.; Pepe, V.; Comitini, F.; Comi, G.; Ciani, M. Enological and genetic traits of Saccharomyces cerevisiae isolated from former and modern wineries. FEMS Yeast Res. 2004, 5, 237–245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ciani, M.; Mannazzu, I.; Marinangeli, P.; Clementi, F.; Martini, A. Contribution of winery-resident Saccharomyces cerevisiae strains to spontaneous grape must fermentation. Antonie Van Leeuwenhoek Int. J. Gen. Mol. Microbiol. 2004, 85, 159–164. [Google Scholar] [CrossRef]
- Knight, S.; Klaere, S.; Fedrizzi, B.; Goddard, M.R. Regional microbial signatures positively correlate with differential wine phenotypes: Evidence for a microbial aspect to terroir. Sci. Rep. 2015, 5, 114233. [Google Scholar] [CrossRef] [Green Version]
- Vigentini, I.; De Lorenzis, G.; Fabrizio, V.; Valdetara, F.; Faccincani, M.; Panont, C.A.; Picozzi, C.; Imazio, S.; Failla, O.; Foschino, R. The vintage effect overcomes the terroir effect: A three year survey on the wine yeast biodiversity in Franciacorta and Oltrepò Pavese, two northern Italian vine-growing areas. Microbiol. 2015, 161, 362–373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sabate, J.; Cano, J.; Querol, A.; Guillamón, J.M. Diversity of Saccharomyces strains in wine fermentations: Analysis for two consecutive years. Lett. Appl. Microbiol. 1998, 26, 452–455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castrillo Cachón, D. Estudio de la diversidad de levaduras en uva de cultivo ecológico y convencional en Galicia: Patrones biogeográficos e influencia en las características químicas y sensoriales del vino. Ph. D. Thesis, University of León, León, Spain, 2018. [Google Scholar]
- Callejon, R.M.; Clavijo, A.; Ortigueira, P.; Troncoso, A.M.; Paneque, P.; Morales, M.L. Volatile and sensory profile of organic red wines produced by different selected autochthonous and commercial Saccharomyces cerevisiae strains. Anal. Chim. Acta 2010, 660, 68–75. [Google Scholar] [CrossRef]
- Blanco, P.; Mirás-Avalos, J.M.; Pereira, E.; Orriols, I. Fermentative aroma compounds and sensory profiles of Godello and Albariño wines as influenced by Saccharomyces cerevisiae yeast strains. J. Sci. Food Agric. 2013, 93, 2849–2857. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
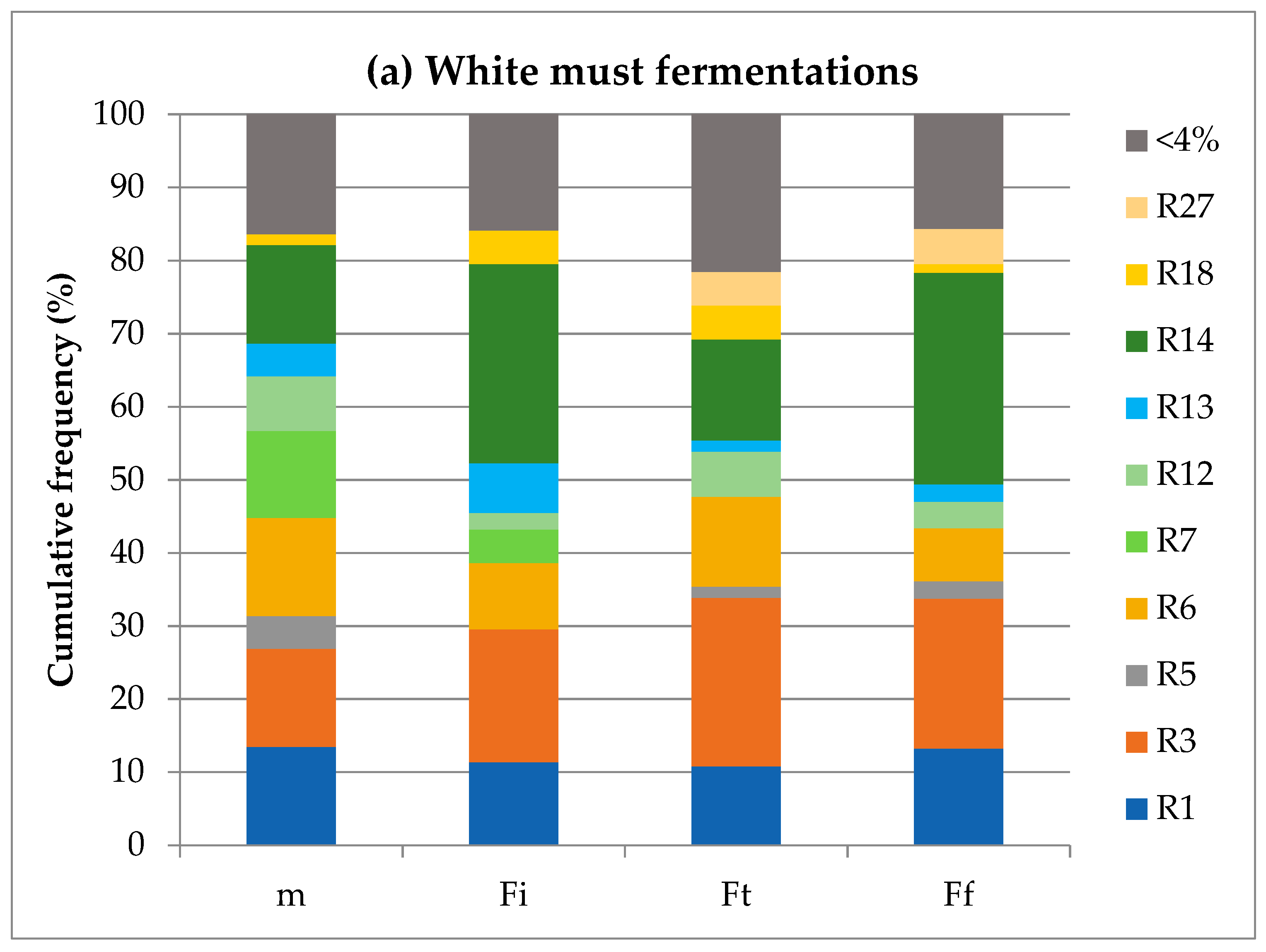{kind=link}
{kind=link}
| Winery (Code) | Denomination of Origin (DO) | Vintages | Number of Fermentations | |
|---|---|---|---|---|
| White | Red | |||
| Corisca (CO) * | Rías Baixas | 2 | 4 (3 + 1) | - |
| Diego de Lemos (DL) | Ribeira Sacra | 3 | 3 (1 + 1 + 1) | 6 (2 + 2 + 2) |
| Adega Beatriz (AB) * | Ribeiro | 2 | 2 (1 + 1) | 2 (1 + 1) |
| Pazo de Vieite (PV) ** | Ribeiro | 3 | 8 (1 + 1 + 6) | 6 (1 + 1 + 4) |
| Quinta da Muradella (QM) | Monterrei | 3 | 5 (2 + 2 + 1) | 13 (1 + 5 + 7) |
| Winery | Number of Fermentations | Isolates Analyzed | Richness * (%) | Diversity Indexes | ||
|---|---|---|---|---|---|---|
| S | H′ | 1-D | E | |||
| Corisca | 4 | 27 | 6 (9%) | 1.12 | 0.53 | 0.63 |
| Diego de Lemos | 9 | 80 | 8 (12%) | 1.73 | 0.79 | 0.89 |
| Adega Beatriz | 4 | 42 | 16 (24%) | 2.51 | 0.90 | 0.91 |
| Pazo de Vieite | 14 | 452 | 46 (70%) | 3.14 | 0.94 | 0.82 |
| Quinta da Muradella | 18 | 262 | 34 (52%) | 2.70 | 0.90 | 0.77 |
| Total | 49 | 863 | 66 (100%) | 3.56 | 0.96 | 0.85 |
| Pairwise | |||||||
|---|---|---|---|---|---|---|---|
| ANOSIM | CO | DL | AB | PV | QM | ||
| Permutation N | 9999 | CO | - | 0.9167 | 1 | 1 | 1 |
| Mean rank within | 26.13 | DL | 0.1037 | - | 0.3333 | 0.4444 | 0.3704 |
| Mean rank between | 57.83 | AB | 0.1002 | 0.1018 | - | 0.5926 | 0.5556 |
| R | 0.604 | PV | 0.0662 | 0.0256 | 0.0296 | - | 0.5556 |
| p (global; same) | 0.0001 | QM | 0.1045 | 0.1021 | 0.1008 | 0.0269 | - |
| PERMANOVA | CO | DL | AB | PV | QM | ||
| Permutation N | 9999 | CO | - | 4.272 | 11.38 | 13.37 | 7.909 |
| Total sum of squares | 3.533 | DL | 0.0995 | - | 2.029 | 1.807 | 1.705 |
| Within-group sum of squares | 1.406 | AB | 0.1003 | 0.102 | - | 2.77 | 2.623 |
| F | 3.781 | PV | 0.0688 | 0.0813 | 0.0298 | - | 2.513 |
| p (global; same) | 0.0001 | QM | 0.0951 | 0.2008 | 0.0974 | 0.0285 | - |
| Strain * | Campaign | ||
|---|---|---|---|
| 2013 | 2014 | 2015 | |
| R1 | 1.14 | 1.36 | 10.46 |
| R3 | 0.00 | 2.27 | 6.91 |
| R6 | 2.27 | 2.27 | 8.60 |
| R14 | 3.41 | 7.73 | 9.44 |
| R18 | 0.00 | 2.27 | 6.75 |
| R23 | 0.00 | 5.00 | 0.17 |
| R26 | 0.00 | 5.00 | 0.17 |
| R28 | 7.95 | 0.91 | 2.02 |
| R42 | 7.95 | 0.45 | 3.71 |
| R44 | 7.95 | 5.45 | 6.07 |
| R56 | 7.95 | 0.00 | 0.34 |
| R63 | 3.41 | 1.36 | 5.90 |
| R68 | 0.00 | 15.00 | 0.34 |
| R79 | 20.45 | 0.00 | 0.00 |
| R88 | 12.50 | 17.27 | 0.84 |
| Total number of strains | 21 | 38 | 54 |
| % major strains | 75.00 | 66.36 | 61.72 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Castrillo, D.; Neira, N.; Blanco, P. Saccharomyces cerevisiae Strain Diversity Associated with Spontaneous Fermentations in Organic Wineries from Galicia (NW Spain). Fermentation 2020, 6, 89. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation6030089
Castrillo D, Neira N, Blanco P. Saccharomyces cerevisiae Strain Diversity Associated with Spontaneous Fermentations in Organic Wineries from Galicia (NW Spain). Fermentation. 2020; 6(3):89. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation6030089
Chicago/Turabian StyleCastrillo, David, Noemi Neira, and Pilar Blanco. 2020. "Saccharomyces cerevisiae Strain Diversity Associated with Spontaneous Fermentations in Organic Wineries from Galicia (NW Spain)" Fermentation 6, no. 3: 89. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation6030089