Snails as Taxis for a Large Yeast Biodiversity
by

Madina Akan
1,2, 
Florian Michling
1,
Katrin Matti
1,
Sinje Krause
1,
Judith Muno-Bender
1 and
Jürgen Wendland
1,2,* 1
Department of Microbiology and Biochemistry, Hochschule Geisenheim University, Von-Lade-Strasse 1, D-65366 Geisenheim, Germany
2
Research Group of Microbiology (MICR)—Functional Yeast Genomics, Vrije Universiteit Brussel, Pleinlaan 2, BE-1050 Brussels, Belgium
*
Author to whom correspondence should be addressed.
Fermentation 2020, 6(3), 90; https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation6030090
Submission received: 24 August 2020
/
Revised: 11 September 2020
/
Accepted: 15 September 2020
/
Published: 18 September 2020
(This article belongs to the Special Issue Yeast Biotechnology 4.0)
Abstract
:Yeasts are unicellular fungi that harbour a large biodiversity of thousands of species, of which particularly ascomycetous yeasts are instrumental to human food and beverage production. There is already a large body of evidence showing that insects play an important role for yeast ecology, for their dispersal to new habitats and for breeding and overwintering opportunities. Here, we sought to investigate a potential role of the terrestrial snails Cepaea hortensis and C. nemoralis, which in Europe are often found in association with human settlements and gardens, in yeast ecology. Surprisingly, even in a relatively limited culture-dependent sampling size of over 150 isolates, we found a variety of yeast genera, including species frequently isolated from grape must such as Hanseniaspora, Metschnikowia, Meyerozyma and Pichia in snail excrements. We typed the isolates using standard ITS-PCR-sequencing, sequenced the genomes of three non-conventional yeasts H. uvarum, Meyerozyma guilliermondii and P. kudriavzevii and characterized the fermentation performance of these three strains in grape must highlighting their potential to contribute to novel beverage fermentations. Aggravatingly, however, we also retrieved several human fungal pathogen isolates from snail excrements belonging to the Candida clade, namely Ca. glabrata and Ca. lusitaniae. Overall, our results indicate that diverse yeasts can utilise snails as taxis for dispersal. This courier service may be largely non-selective and thus depend on the diet available to the snails.
1. Introduction
Yeasts, in particular Saccharomyces cerevisiae, have been associated with human activities for millennia. It could be argued that this yeast was our first domesticated species, probably long before humans built permanent settlements. S. cerevisiae has been studied very intensively over decades and is the model eukaryote [1]. Yet, knowledge on yeast ecology is only beginning to emerge. We can now distinguish between several thousand yeast species, primarily amongst ascomycetes, although yeasts, i.e., unicellular fungi that proliferate preferably by budding, also exist in other fungal classes. There is a large body of evidence on yeasts and their world-wide distribution, which is certainly aided by large scale sequencing approaches that also lead to the identification of hundreds of novel yeast species every year [2].
One can distinguish between persistent species that reside in a given habitat and transient species that are only associated with a given habitat for a short while. The ecology of yeasts their relationship with other organisms needs to be studied and understood as well.
Yeasts require suitable substrates that supply them with the required nutrients. Thus, a large biodiversity is often found with flowers, fruits or plant exudates [3,4]. However, this displays an intrinsic problem, as yeast cells require a mode of transportation to reach these food sources. A distribution by water or wind is not targeted and provides little success and on the other hand, yeasts themselves are non-motile. Yeasts are often found in tri-trophic interactions including a yeast, a plant and an insect vector the yeast cells can depend upon for dispersal [5,6,7].
The association of yeast and animals could be transient or maintained over longer periods. The potential benefits for the yeast are: (i) transport to a new nutrient source, (ii) getting actual access to the nutrient source, e.g., by stinging sucking insects that break an otherwise insurmountable barrier and (iii) finding a place e.g., in the insect gut to remain protected through periods of low nutrient availability, particularly over winter [8].
The potential benefits for the insect are: (i) presentation of a food source as yeasts may indicate them via the production of specific volatiles meant to attract a specific insect species (this is like calling a cab) [9], (ii) offering of yeasts themselves as a protein rich food source (that is the price to pay for the cab), (iii) provision of essential components required, e.g., for larval development (to ensure services also in the future) [10] and (iv) to obtain microorganisms that have probiotic functions in the insect gut.
For S. cerevisiae, wasps have been identified as a key animal vector and a natural reservoir [8]. Wasps are capable of opening grapes by biting through the grape skin and thereby inoculating yeast cells into the sugar rich juice. In addition, wasp queens provide shelter for S. cerevisiae while overwintering. It was also shown that the insect gut serves as a platform to promote outbreeding and hybridization of different Saccharomyces species [11]. S. cerevisiae can attract Drosophila by producing acetate esters (volatile aroma compounds) and ATF1, a gene coding for an acetyltransferase, provides a crucial enzymatic activity for their production [12]. Yeasts in a Drosophila diet can also promote their reproduction [13,14,15].
These tri-trophic interactions go beyond simple short-lived interactions but often approach evolved relationships, i.e., symbiosis [2]. These evolved relationships may have had further consequences in that insects provide special organs, mycetocytes/mycetomes, to harbour their symbionts [16].
These are just two examples of the symbiotic benefits of a yeast–insect relationship and many more wait to be uncovered. Studying tri-trophic interactions may also be of importance for industrial microbiology. In the case of Ashbya gossypii, it appears that stinging-sucking insects (Heteroptera) use Ashbya for opening a new food source (oleander) to them. These insects can apparently only feed on oleander if Ashbya detoxifies plant compounds by overproducing riboflavin [17,18,19]. To adapt to this niche Ashbya evolved into an efficient riboflavin overproducer, a trait that is of strong commercial interest [20]. Thus, studying the yeast biodiversity may help in discovering novel platform strains as producers of diverse chemical compounds [21,22].
Yeasts are present ubiquitously. Thus, yeasts most certainly will be associated with other animals and utilise them as vectors or habitats, which could of course also include pathogenic interactions [23].
Snails and slugs do have a love for beer. This is evident from the vast body of entries in the public domain mostly circulating around how to remove snails form the garden (e.g., https://www.wikihow.com/Use-Beer-to-Get-Rid-of-Snails-in-Your-Garden). Yet, there has not been a scientific follow-up. Snail juice is known for its antimicrobial properties due to the production of cell wall lytic enzymes [24,25,26,27,28]. Thus, yeasts may be attractive as a food source and, based on their aroma volatile production, may actually indicate food sources of potential interest to snails. This resembles yeast–insect relationships, as there could be benefits also for yeasts in that snails could present taxis for circulating yeasts in the environment.
Here, we used two well-known snail species, which are abundant in Europe and have also started to invade North America, namely the closely related land snails Cepaea nemoralis and C. hortensis [29]. Cepaea is closely associated with human settlements [30]. We asked the following questions: (i) Are yeasts associated with these land snails? (ii) Can yeasts use snails as taxis to spread them in the environment? (iii) Which yeast species are associated with these snails? (iv) Do such yeasts have a potential biotechnological value, i.e., can they be successfully used e.g., in grape must fermentations?
2. Materials and Methods
2.1. Strains and Media
Yeast strains identified and used in this study are shown in Supplemental Table S1. Yeast strains were isolated and subcultured in YPD (1% yeast extract, 2% peptone, 2% glucose) with the addition of kanamycin (50 µg/mL) and ampicillin (100 µg/mL) to inhibit bacterial growth. Yeast were generally grown at 30 °C.
2.2. Yeast Isolation
Snails were collected in the early morning hours or in the late afternoon according to their activity rhythms. Snails were quarantined in sterile 50 mL Falcon tubes until they generated excrements. Afterwards, all snails were released into their environment and no snail was sacrificed during the course of this study. Excrements were suspended in 500 µL sterile water and aliquots were spread on YPD petri plates, incubated overnight and microscopically inspected for growth. Small yeast colonies (amongst a background of filamentous fungal growth and sometimes also bacterial growth) were isolated via micromanipulation (Singer Instruments, Somerset, UK) while larger colonies (upon longer incubation) were picked manually.
2.3. Molecular Typing of Yeast Isolates
The determination of genus/species assignments of yeast isolates was performed by ITS-PCR (ITS, Internal Transcribed Spacer) using standard fungal specific ITS1 (5′-CTTGGTCATTTAGAGGAAGTAA-3′) and ITS4 (5′-TCCTCCGCTTATTGATATGC-3′) primers on genomic DNA of yeast strains. Direct sequencing of the respective PCR products (after column purification) was done at Starseq, Mainz, Germany, and at the VIB Core Facilities, Leuven, Belgium.
ITS sequences were identified by MegaBlast search against a local blast database using NCBI blast+ suite version 2.10.1, evaluating the first 100 matches with pairwise identity >90%. The reference database comprised 810,396 unique fungal DNA sequences retrieved from the NCBI partially redundant nucleotide collection (ftp://ftp.ncbi.nlm.nih.gov/blast/db/), retrieved September 5, 2020, using query expression ‘ribosomal internal transcribed spacer AND fungi [filter]’. The nucleotide sequences obtained in this study have been submitted to GenBank (accession numbers MT988151–MT988312).
2.4. Whole Genome Shotgun Sequencing
Whole genome shotgun sequencing data was generated for isolates B079 (Hanseniaspora uvarum), B143 (Pichia kudriavzevii) and B144 (Meyerozyma guilliermondii) using Illumina MiSeq V3 technology and 2× 150 bp read length (LGC Genomics GmbH, Berlin, Germany).
Quality trimming, error-correction and decontamination were performed using BBMap version 38.84 (https://sourceforge.net/projects/bbmap/). Quality-trimmed and error-corrected reads were assembled using SPAdes version 3.14.1 [31] without additional error correction and in ‘careful’ mode for a range of kmer lengths; contigs shorter than <1 kb were discarded. For each isolate, multiple assemblies with high L50 and low N50 values were investigated further.
Locally Collinear Blocks (LCB) were detected using MCM (Mauve Contig Mover, [32]) with automatic seed weight and LCB score determination as implemented in Geneious 2019.2.1 (Biomatters Ltd., NZL-1010 Auckland). Based on the MCM alignment, dissimilarity between query and reference genome was calculated for consecutive 1 kb windows using function dist.dna from package ape version 5.3 [32] in R version 3.6.3. Divergence was estimated using distance metric ‘T92’ [33]. Figure 4 was generated with package circlize version 0.4.10 in R version 3.6.3 [34].
The reads were mapped against sorted contigs (i.e., in order as determined by MCM, see above) and variant call was performed using BBMap. Variant frequencies were tabulated and plotted to estimated isolate ploidy.
The Whole Genome Shotgun projects have been deposited at DDBJ/ENA/GenBank under the accessions JACRSB000000000, JACRSC000000000 and JACRSD000000000. The versions described in this paper are versions JACRSB010000000, JACRSC010000000 and JACRSD010000000.
2.5. Fermentation Conditions
Non-conventional yeasts isolated from snails were subjected to lab-scale fermentations of standard pasteurized white wine must containing sugars amounting to 72° Oechsle. The must was supplemented by adding Fermaid E (inactivated yeast) according to the supplier’s specifications (Lallemand, Vienna, Austria). The cells were inoculated with an OD600 = 0.5. Fermentations were run at 18 °C under constant stirring at 300 rpm conveyed by a magnetic stirrer. Fermentations with the wine yeast EC1118 were done using Ankom devices as described [35].
2.6. Analytical Methods
Chemical analysis at the end of the fermentations included HPLC (High-Pressure Liquid Chromatography) quantification of several compounds including fructose, glucose, ethanol and organic acids; aroma compounds were analysed by GC (Gas Chromatography) as described in Matti et al. [35].
3. Results
3.1. Isolating Yeast Strains from Snails
Some snail species are well-known in Europe and occur in close proximity to human settlements. This may be due to the gardens and food but also because of the shelter we provide and for protection from thrushes and mice, their major predators [30,36]. The brown-lipped grove snail Cepaea nemoralis (and its close relative C. hortensis) are the most common land snails in Europe, which—because of their beauty and colour polymorphisms, i.e., banding patterns—have actually been introduced in North America on purpose [29,37]. Therefore, we chose these species as test subjects.
Our starting hypothesis was that snails act as vectors for yeasts and can taxi them from one food source to the other (Figure 1A). This was also motivated by finding these snails on rich sugar sources such as grapes (Figure 1B).
The collection of snails is dependent on the weather conditions and seasons as snails stay hidden in dry weather over summer and hibernate from late fall until early spring. Thus, our collection efforts were concentrated in the spring and summer months. Collecting snails is simple and is most successful in the early morning and early evening hours or after rainfall. We sampled snails from several regions in central Europe, mostly from Hoeilaart near Brussels, Belgium (Latitude/Longitude: 50.7674846, 4.474407), but also from Geisenheim (49.9863945, 7.96852), Heidelberg (49.4093582, 8.694724) and from Butzbach/Hoch-Weisel (50.4029544, 8.63934) in Germany.
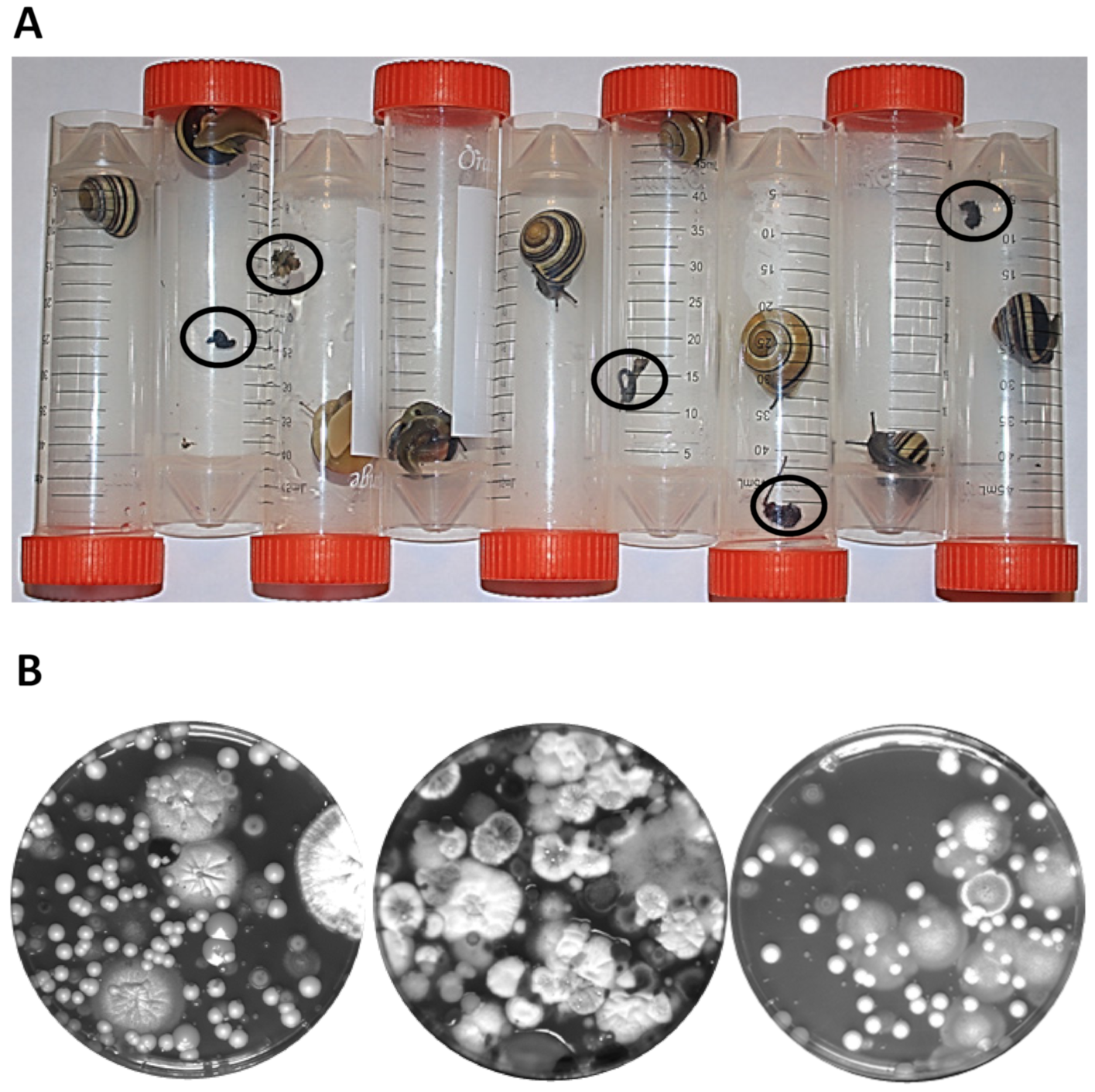
The snails were quarantined in sterile 50 mL tubes until they deposited their excrements, which could occur within a few hours, but their quarantine was regularly carried out overnight (Figure 2A). Afterwards, the snails were released into their environment and no snail was sacrificed or killed in action. Excrements were transferred to Eppendorf tubes and suspended in 500 µL sterile water. Microscopic inspection already indicated the presence of yeast like microorganisms (not shown). Thus, dilutions were plated on rich medium plates (limiting bacterial growth with antibiotics). The plates were incubated over-night or longer and revealed a large biodiversity of yeasts and filamentous fungi (Figure 2B).
Yeasts were isolated by manual picking or by micromanipulation and restreaked to obtain pure culture isolates.
3.2. Typing of Yeast Isolates
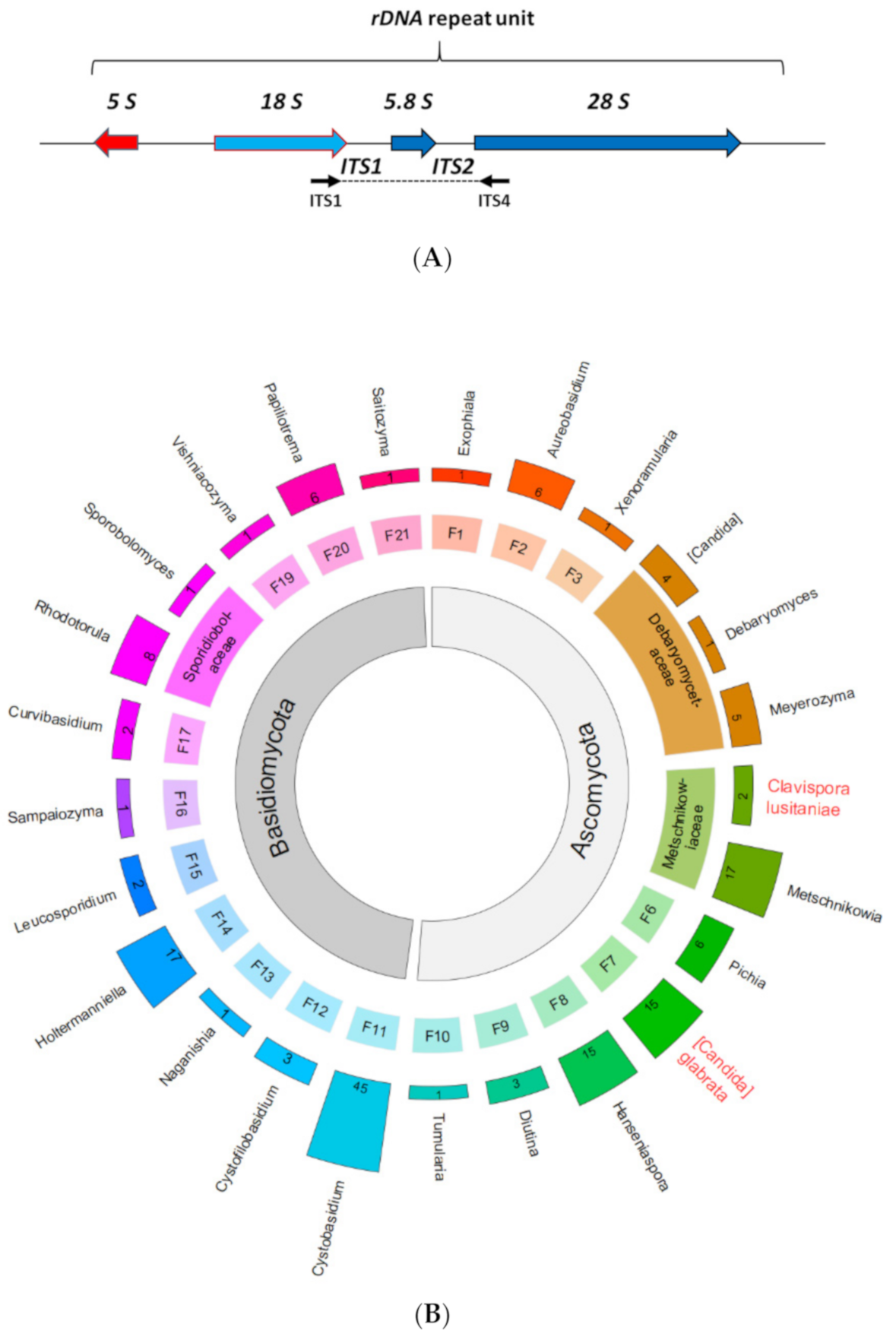
We did not sample bacterial or filamentous fungal biodiversity and did not approach the topic with metagenomics. Instead, we used culture-dependent isolation of yeast strains to secure these strains for downstream analyses such as the fermentation and flavour analyses described below. Genus or species level determination was carried out by regular ITS-typing. For this, genomic DNA was prepared and standard fungal ITS-primers were used to amplify a region from the rDNA repeat unit encompassing the end of the 18S, ITS1, 5.8S, ITS2 and the start of the 28S regions (Figure 3A). The obtained PCR-products were sequenced, and these sequences were compared to the databases to identify the most closely related species and group the isolates at least to the genus level (Figure 3B). We isolated typed >150 strains (Table S1).
This provided the following insight: (i) Yeast-like fungi were found belonging to both ascomycetes and basidiomycetes. This indicated that even with a relatively small sampling size a wide spectrum of yeasts could be identified. (ii) Several ascomycetous isolates grouped within the Candida clade harbouring human fungal pathogens. These were identified as Ca. glabrata and Ca. lusitaniae, while the other isolates were grouped with emerging opportunistic fungal pathogens e.g., belonging to the genus Rhodotorula [38,39]. (iii) There was a substantial overlap in the biodiversity uncovered from snails with that found in vineyards, particularly with yeasts of the genera Aureobasidium, Hanseniaspora, Metschnikowia, Pichia and Rhodotorula [40,41]. (iv) In general, our data indicate that diverse yeasts—despite the lytic enzymes snails can produce—survived passage through the gut and could thus be disseminated with snail excrements and deposited in potentially favourable environments.
3.3. Genome Sequencing
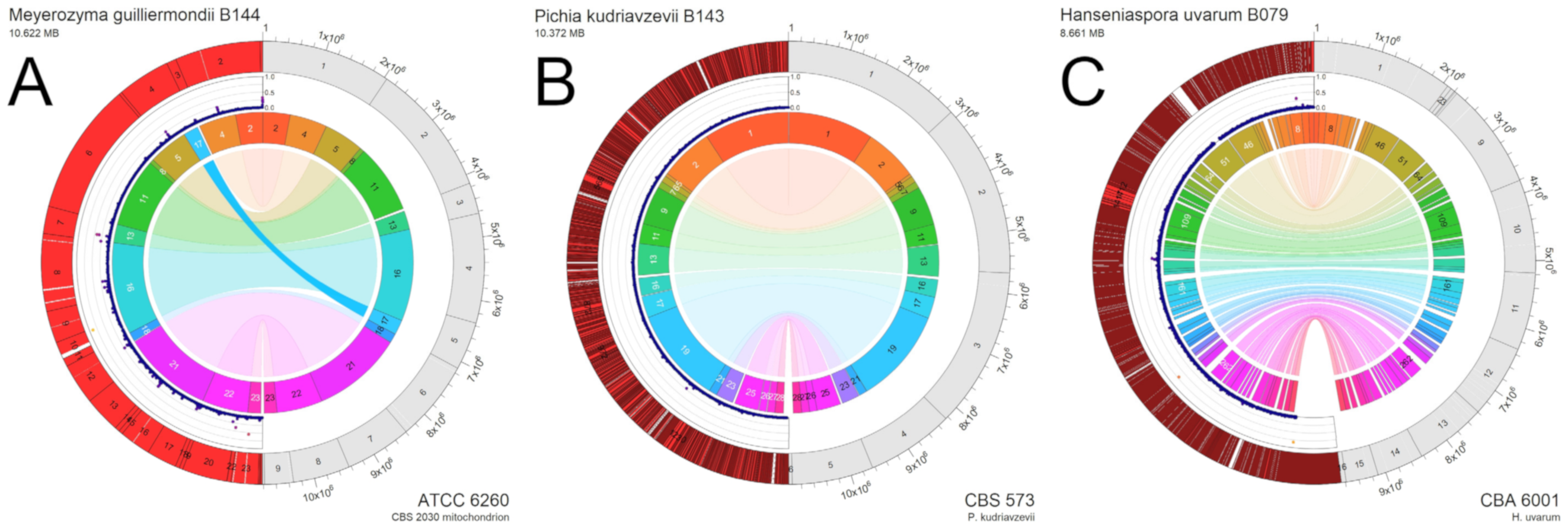
Interestingly, we recovered several yeast species from snails that are often isolated as non-conventional yeasts from grapes or found in grape must. For three of these yeast strains, B079 (Hanseniaspora uvarum), B143 (Pichia kudriavzevii) and B144 (Meyerozyma guilliermondii), we obtained draft genome sequences using the Illumina sequencing platform (see Materials and Methods). Sequencing statistics are detailed in Supplemental Tables S2 and S3. A highly contiguous draft genome was assembled for M. guilliermondii B144. The assemblies for Hanseniaspora uvarum B079 and Pichia kudriavzevii B143 produced noticeably shorter scaffolds (and thus a larger number of contigs and gaps). Variant frequency distributions suggested B079 and B143 are diploid strains, whereas B144 was haploid. We compared the genomic assemblies to their respective reference genomes. For all draft genomes, collinearity and sequence identity to the respective reference genome was very high, indicating that our species assignment was correct and for M. guilliermondii we did not detect major rearrangements, except for one approx. 346 kb translocation in B144. While the other two genomes could be aligned with the reference genomes as well, we cannot provide data on genome rearrangements due to the large number of contigs generated in these assemblies (Figure 4).
3.4. Fermentation Characteristics of Non-Conventional Yeasts
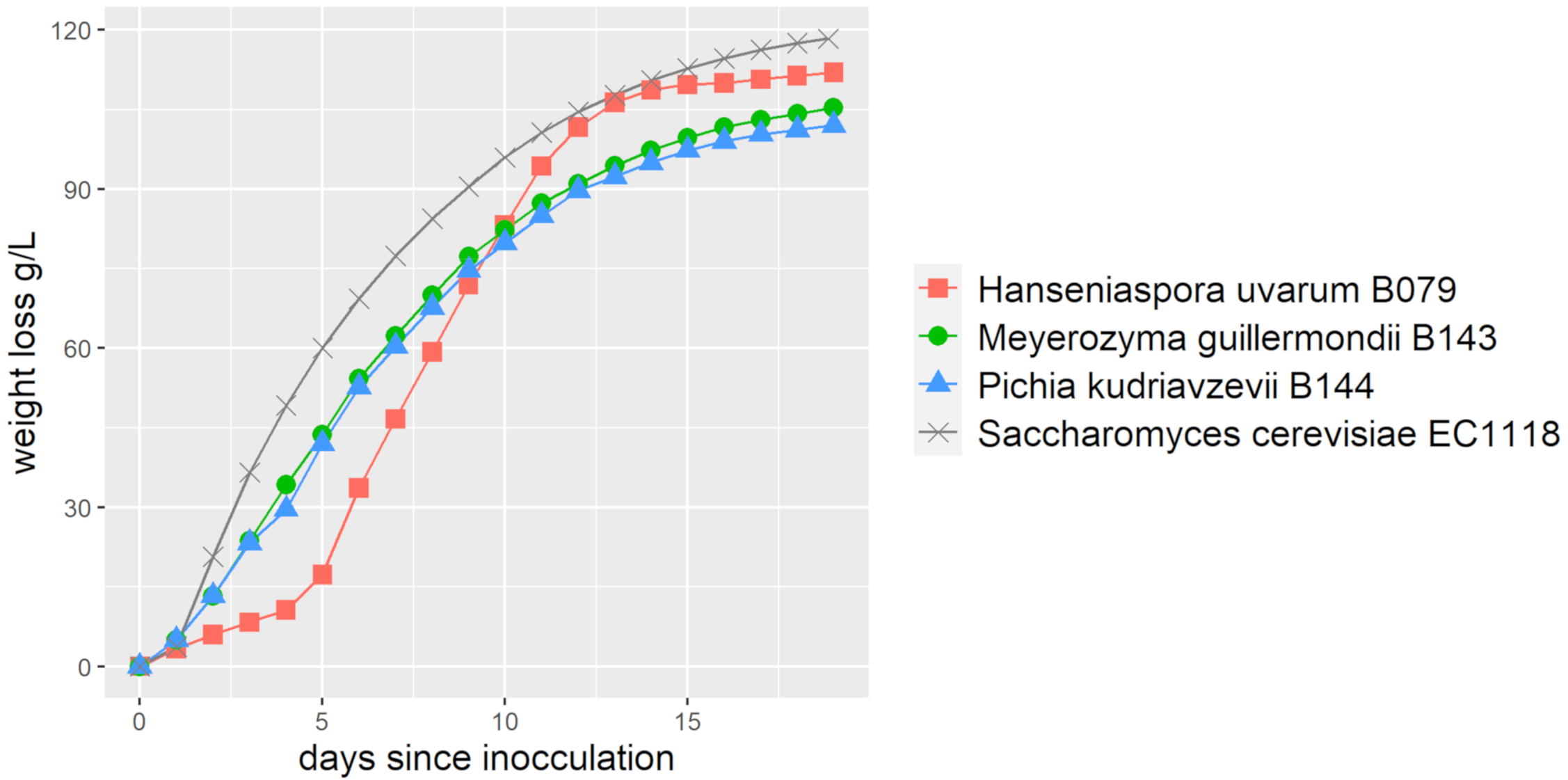
Next, we determined the usability of these three yeast strains in grape must fermentations. We employed a regular white wine must (Müller Thurgau) with 72° Oechsle sugar content. This must was supplemented with an inactivated yeast preparation to provide additional nitrogen (Fermaid E, Lallemand). The fermentations were carried out in triplicate over three weeks and their progress was monitored by measuring weight loss (CO2-release). Oxygen supply for fermentations with non-conventional yeasts was realised by large headspace volumes, which is in contrast to the anaerobic fermentations conducted with the wine yeast EC1118. Substantial weight loss was observed with all strains, concurrent with the utilization of must sugars (Figure 5). While H. uvarum showed a longer lag phase in these experiments, it overtook the other two strains mid-way and finished its fermentations fastest. M guilliermondii and P. kudriavzevii did not reach their end of fermentation in the three-week period, which was mainly due to their inability to ferment fructose completely (Table 1). Besides residual sugar levels, we determined organic acids and ethanol content at the end of fermentation. The ethanol levels produced by these strains were substantial thus raising an interest in these strains for low-alcohol fermentations. Interestingly, the amount of malic acid was found to be lower (less than half) and thus more preferable compared to what is normally found in fermentations carried out with wine yeasts (see [35] for comparison).
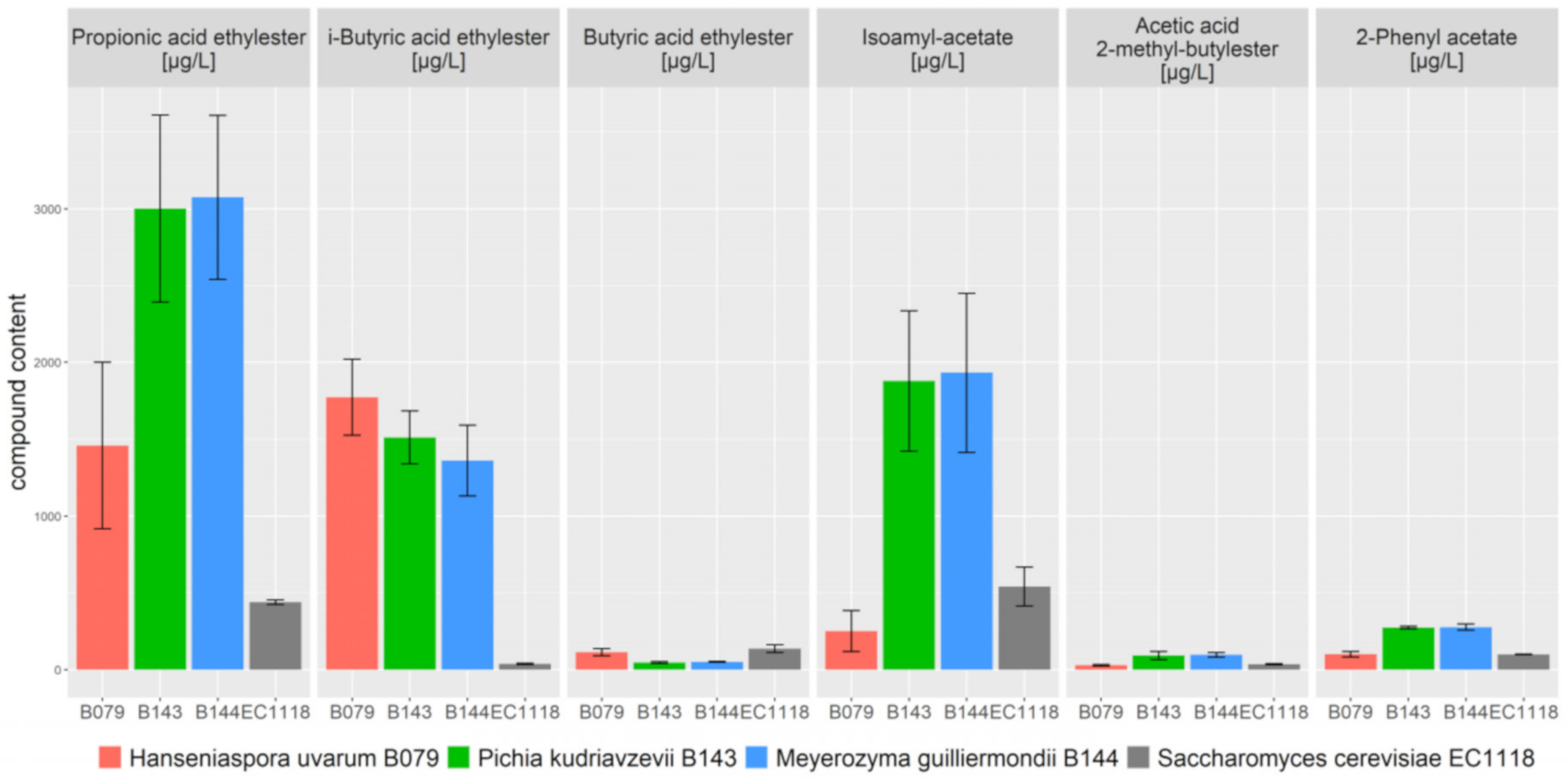
We also analysed several aroma compounds via gas chromatography. Here, we found that the amounts of ethyl acetate (acetic acid ethyl ester) were substantially higher (>tenfold) in the fermentations of these non-conventional yeasts compared to oxygen limited fermentations with wine yeasts (Table 2). While these levels of ethyl acetate are generally perceived as a wine fault in S. cerevisiae wine yeast mediated fermentations, it was noted that these three non-conventional strains were actually good ester producers that are able provide increased fruitiness to alcoholic beverages (Figure 6). The genera Hanseniaspora and Pichia have also previously been noted for their potentially beneficial ester production capabilities in oenological settings [42].
4. Discussion
Our study provided a multidisciplinary approach to yeast ecology and fermentation research that extends into genomics and volatile flavour analysis. Here, we presented the most common land snails of Europe, Cepaea nemoralis, which is now also present in North America, as reservoir for a large yeast biodiversity. This adds to the long-established mutualistic interaction between yeasts and insects and in particular to the well-established connection between S. cerevisiae and wasps [5,8,11,15,43,44,45].
In yeast–insect relationships, benefits are bestowed on both yeasts and their insect partners, which on the animal side may include promotion of larval development, immunity and longevity [46,47]. Benefits for yeasts are the offer of transportation to new food sources and potentially the provision of shelter during off-seasons with limited fruit availability.
Previously, snails were thought to digest yeast cells and feed on them due to their production of lytic enzymes [24]. Our data convincingly show that yeasts of different genera survive passage through the snail gut. Thus, these animals represent a previously neglected reservoir for both ascomycete and basidiomycete yeasts. Currently, our knowledge on this relationship is limited to the taxi component providing benefits for yeasts in helping in their dispersal and providing benefits for snails through the potential attraction to food sources by yeast volatiles. The taxi component may not always be ideal, as it is not guaranteed for the yeasts to reach a sugar rich destination when confined in snail deposits. If so, a second messenger may be required, e.g., a fly that can provide another ride to a final destination that promotes new growth and proliferation. Also, yeasts may be transported by snails via smearing along their paths. In this study we have not investigated snails as reservoirs over longer periods of unfavourable conditions. During dry spells in summer snails stay hidden and of course between late fall and early spring (October to March) snails hibernate. It is not clear at this time if snails provide shelter for yeasts over any of these periods as has been shown for queen wasps during hibernation [8]. The retention time in the snail gut, however, is experimentally accessible, as snails do not need to hibernate in the lab (simply based on temperature) and can thus be subjected to defined dry spells.
We isolated only a few yeast strains per snail; however, these strains could occur in quite large numbers per snail (see Figure 2). The biodiversity view could only be established by collecting scores of snails. This indicates that in contrast to wasps, snails may be quite promiscuous and thus constitute rather non-selective taxis for yeasts. We did not identify a S. cerevisiae strain in our collection. This may simply be due to the small scale of our approach or due to the digestive enzymes that do not allow this yeast’s survival. To test this, we hosted several snails in boxes and fed them vegetables spiked with S. cerevisiae. With this approach, we were then successful in isolating S. cerevisiae from snail excrements.
In our collection, we also identified two human fungal pathogenic Candida species, of which Ca. glabrata is the second most prevalent Candida pathogen for humans [48]. This is interesting since the natural niche for Ca. glabrata has not been defined. Ca. glabrata as wells a Ca. albicans are commensals in the human body [49]. Recently, Ca. albicans was isolated also from oak trees and may thus have a natural niche there. A close relative of C. nemoralis is the grapevine snail Helix pomatia. During our collection efforts, we observed that a large number of these snails sought out shelter on stems of oak trees during the day in one specific valley in the Sonian Forest of Hoeilaart, Belgium. There, the snails crawled up oak trees (one snail per tree only) to about 1 m height, apparently to evade predators. Thus, in a follow-up study a closer look at the occurrence of Ca. albicans in forest snails is warranted.
C. nemoralis is closely associated with human settlements. Thus, this represents a potential infection route with human fungal pathogens via Cepaea depositing these fungal cells in our garden on vegetables destined for raw consumption. This should be monitored in more detail in the future. In another study, Ca. glabrata was also isolated from gulls, which indicated another reservoir of this human fungal pathogen [50].
The biodiversity discovered in Cepaea offered a substantial overlap with yeast genera generally found in the vineyard. This could be due to the locations chosen for sampling, which are closely associated with grape growing. Geisenheim in the Rheingau is a world-famous wine making region. Hoeilaart, on the other hand, is a place where vines are grown in greenhouses to produce table grapes and recently the wine production in Belgium is increasing due to the warming climate, while Hoch-Weisel harbours a variety of different apple trees [51]. We chose to study three yeast strains in more detail and analysed their fermentation behaviour.
These strains showed useful properties and could also enter in co-fermentations with S. cerevisiae. H. uvarum has already been tested in a sequential co-fermentations with a wine yeast and found to decrease volatile acidity, which is in line with our observations [52]. While M. guilliermondii has been found in traditional rice wine fermentations, P. kudriavzevii was actually used in 1:1 co-fermentations with S. cerevisiae to produce wines form overripe Cabernet Sauvignon grapes [53,54]. This demonstrates the potential of non-conventional yeasts in spicing up regular wine yeast fermentations [55]. Some non-conventional yeast species may also show potential as human fungal pathogens. P. kudriavzevii (regarded as non-pathogenic) has been found to be the same species as Issatchenkia orientalis, Ca. glycerinogenes and Ca. krusei (one of the prevalent causes of clinical yeast infections) [56,57]. M. guilliermondii known also as Pichia guilliermondii, can be utilised as biocontrol fungus against spoilage fungi but has also emerged as human fungal pathogen [58].
Our genome analyses indicate that our isolates were substantially similar to previously published genomes. This also revealed that our H. uvarum strain isolate is a member of the fast evolving Hanseniaspora lineage with a low GC content [59].
5. Conclusions
Our study revealed that snails of the genus Cepaea, which are closely associated with human settlements, serve as distributors of yeasts encompassing a large biodiversity. This has long been overlooked and provides a new aspect in yeast ecology. It will be interesting to determine if snails can also serve to shelter yeasts over dry periods in summer or during hibernation in winter.
Interestingly, snails are also a reservoir for human fungal pathogens. This warrants more detailed analyses preferably employing culture-independent next-generation metagenomics sequencing tools and an expansion of the species sampled.
Several of the yeast isolates of this study have also been identified in other biodiversity studies indicating that yeasts are quite industrious in finding means of dispersal. Snails may also be a reservoir for non-conventional yeasts with a potential of being used as starter cultures for alcoholic beverage fermentations.
Supplementary Materials
The following are available online at https://0-www-mdpi-com.brum.beds.ac.uk/2311-5637/6/3/90/s1, Table S1: List of yeast strains isolated in this study, Table S2: Draft Genome statistics for 3 yeast isolates obtained from snail excrement, Table S3: Draft Genome statistics collinearity to reference genome.
Author Contributions
Conceptualization, J.W.; methodology, J.W., F.M., M.A., K.M., S.K.; validation, J.W., F.M., M.A., K.M.; formal analysis, J.W., M.A., F.M., K.M.; investigation, J.W., M.A., F.M., K.M., S.K.; resources, J.W., data curation, J.W., F.M., M.A., K.M., S.K.; writing—original draft preparation, J.W.; writing—review and editing, J.W., M.A., F.M.; visualization, J.W., F.M., J.M.-B., K.M., S.K.; supervision, J.W.; project administration, J.W.; funding acquisition, J.W. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded in part by the European Union Marie Curie Innovative Training Network Aromagenesis 764364 (http://www.aromagenesis.eu). F.M. was funded by the Hessen State Ministry of Higher Education, Research and the Arts (HMWK) via the LOEWE cluster AROMAplus (https://www.hs-geisenheim.de/aromaplus/).
Acknowledgments
We thank the Analytic Team of the Department, particularly Silvia Brezina and Heike Semmler for HPLC and GC analyses and Stefanie Fritsch for calibrations of the analytical equipment and Mattner for excellent technical service.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Botstein, D.; Fink, G.R. Yeast: An experimental organism for 21st Century biology. Genetics 2011, 189, 695–704. [Google Scholar] [CrossRef] [Green Version]
- Starmer, W.T.; Lachance, M.A. Yeast ecology. In The Yeasts: A Taxonomic Study; Kurtzman, C.P., Fell, J.W., Boekhout, T., Eds.; Elsevier Science: London, UK; Burlington, MA, USA, 2011; pp. 65–83. [Google Scholar]
- Hilber-Bodmer, M.; Schmid, M.; Ahrens, C.H.; Freimoser, F.M. Competition assays and physiological experiments of soil and phyllosphere yeasts identify Candida subhashii as a novel antagonist of filamentous fungi. BMC Microbiol. 2017, 17, 4. [Google Scholar] [CrossRef] [Green Version]
- Chappell, C.R.; Fukami, T. Nectar yeasts: A natural microcosm for ecology. Yeast 2018, 35, 417–423. [Google Scholar] [CrossRef] [Green Version]
- Ganter, P.F. Yeast and invertebrate associations. In Biodiversity and Ecophysiology of Yeasts; Rosa, C.A., Peter, G., Eds.; Springer: Berlin/Heidelberg, Germany, 2006; pp. 303–370. [Google Scholar]
- Madden, A.A.; Epps, M.J.; Fukami, T.; Irwin, R.E.; Sheppard, J.; Sorger, D.M.; Dunn, R.R. The ecology of insect-yeast relationships and its relevance to human industry. Proc. Biol. Sci. 2018, 285. [Google Scholar] [CrossRef] [Green Version]
- Klaps, J.; Lievens, B.; Alvarez-Perez, S. Towards a better understanding of the role of nectar-inhabiting yeasts in plant-animal interactions. Fungal. Biol. Biotechnol. 2020, 7, 1. [Google Scholar] [CrossRef] [Green Version]
- Stefanini, I.; Dapporto, L.; Legras, J.L.; Calabretta, A.; Di Paola, M.; De Filippo, C.; Viola, R.; Capretti, P.; Polsinelli, M.; Turillazzi, S.; et al. Role of social wasps in Saccharomyces cerevisiae ecology and evolution. Proc. Natl. Acad. Sci. USA 2012, 109, 13398–13403. [Google Scholar] [CrossRef] [Green Version]
- Palanca, L.; Gaskett, A.C.; Gunther, C.S.; Newcomb, R.D.; Goddard, M.R. Quantifying variation in the ability of yeasts to attract Drosophila melanogaster. PLoS ONE 2013, 8, e75332. [Google Scholar] [CrossRef] [Green Version]
- Murgier, J.; Everaerts, C.; Farine, J.P.; Ferveur, J.F. Live yeast in juvenile diet induces species-specific effects on Drosophila adult behaviour and fitness. Sci. Rep. 2019, 9, 8873. [Google Scholar] [CrossRef] [Green Version]
- Stefanini, I.; Dapporto, L.; Berna, L.; Polsinelli, M.; Turillazzi, S.; Cavalieri, D. Social wasps are a Saccharomyces mating nest. Proc. Natl. Acad. Sci. USA 2016, 113, 2247–2251. [Google Scholar] [CrossRef] [Green Version]
- Christiaens, J.F.; Franco, L.M.; Cools, T.L.; De Meester, L.; Michiels, J.; Wenseleers, T.; Hassan, B.A.; Yaksi, E.; Verstrepen, K.J. The fungal aroma gene ATF1 promotes dispersal of yeast cells through insect vectors. Cell Rep. 2014, 9, 425–432. [Google Scholar] [CrossRef] [Green Version]
- Witzgall, P.; Proffit, M.; Rozpedowska, E.; Becher, P.G.; Andreadis, S.; Coracini, M.; Lindblom, T.U.; Ream, L.J.; Hagman, A.; Bengtsson, M.; et al. “This is not an apple”-yeast mutualism in codling moth. J. Chem. Ecol. 2012, 38, 949–957. [Google Scholar] [CrossRef] [PubMed]
- Tatum, E.L. Nutritional requirements of drosophila melanogaster. Proc. Natl. Acad. Sci. USA 1939, 25, 490–497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stefanini, I. Yeast-insect associations: It takes guts. Yeast 2018, 35, 315–330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suh, S.O.; Noda, H.; Blackwell, M. Insect symbiosis: Derivation of yeast-like endosymbionts within an entomopathogenic filamentous lineage. Mol. Biol. Evol. 2001, 18, 995–1000. [Google Scholar] [CrossRef]
- Walther, A.; Wendland, J. Yap1-dependent oxidative stress response provides a link to riboflavin production in Ashbya gossypii. Fungal Genet. Biol. 2012, 49, 697–707. [Google Scholar] [CrossRef]
- Dietrich, F.S.; Voegeli, S.; Kuo, S.; Philippsen, P. Genomes of Ashbya fungi isolated from insects reveal four mating-type loci, numerous translocations, lack of transposons, and distinct gene duplications. G3 (Bethesda) 2013, 3, 1225–1239. [Google Scholar] [CrossRef] [Green Version]
- Aguiar, T.Q.; Silva, R.; Domingues, L. Ashbya gossypii beyond industrial riboflavin production: A historical perspective and emerging biotechnological applications. Biotechnol. Adv. 2015, 33, 1774–1786. [Google Scholar] [CrossRef] [Green Version]
- Revuelta, J.L.; Ledesma-Amaro, R.; Lozano-Martinez, P.; Diaz-Fernandez, D.; Buey, R.M.; Jimenez, A. Bioproduction of riboflavin: A bright yellow history. J. Ind. Microbiol. Biotechnol. 2017, 44, 659–665. [Google Scholar] [CrossRef]
- Wendland, J. Special issue: Non-conventional yeasts: Genomics and biotechnology. Microorganisms 2019, 8, 21. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.; Alper, H.S. Non-conventional hosts for the production of fuels and chemicals. Curr. Opin. Chem. Biol. 2020, 59, 15–22. [Google Scholar] [CrossRef]
- Reis, E.J.C.; Buscariolo, F.; Siqueira, J.P.Z.; Castilho, E.M.; Almeida, M.T.G. Agapornis sp. pet birds: Source of dissemination of azole-resistant yeasts. Med. Mycol. 2019, 57, 515–518. [Google Scholar] [CrossRef] [PubMed]
- Eddy, A.A.; Williamson, D.A. Method of isolating Protoplasts from Yeast. Nature 1957, 179, 1252–1253. [Google Scholar] [CrossRef]
- Johnston, J.R.; Mortimer, R.K. Use of snail digestive juice in isolation of yeast spore tetrads. J. Bacteriol. 1959, 78, 292. [Google Scholar] [CrossRef] [Green Version]
- Millbank, J.W.; Macrae, R.M. Degradation of yeast cell wall by fractionated snail gut enzyme. Nature 1964, 201, 1347. [Google Scholar] [CrossRef]
- Anderson, F.B.; Millbank, J.W. Protoplast formation and yeast cell-wall structure. The action of the enzymes of the snail, Helix pomatia. Biochem. J. 1966, 99, 682–687. [Google Scholar] [CrossRef] [Green Version]
- Cilia, G.; Fratini, F. Antimicrobial properties of terrestrial snail and slug mucus. J. Complement. Integr. Med. 2018, 15. [Google Scholar] [CrossRef]
- Whitson, M. Cepaea nemoralis (Gastropoda, Helicidae): The invited invader. J. Ky. Acad. Sci. 2005, 66, 82–88. [Google Scholar] [CrossRef]
- Rosin, Z.M.; Kwiecinski, Z.; Lesicki, A.; Skorka, P.; Kobak, J.; Szymanska, A.; Osiejuk, T.S.; Kaluski, T.; Jaskulska, M.; Tryjanowski, P. Shell colour, temperature, (micro)habitat structure and predator pressure affect the behaviour of Cepaea nemoralis. Naturwissenschaften 2018, 105, 35. [Google Scholar] [CrossRef] [Green Version]
- Nurk, S.; Bankevich, A.; Antipov, D.; Gurevich, A.A.; Korobeynikov, A.; Lapidus, A.; Prjibelski, A.D.; Pyshkin, A.; Sirotkin, A.; Sirotkin, Y.; et al. Assembling single-cell genomes and mini-metagenomes from chimeric MDA products. J. Comput. Biol. 2013, 20, 714–737. [Google Scholar] [CrossRef] [Green Version]
- Rissman, A.I.; Mau, B.; Biehl, B.S.; Darling, A.E.; Glasner, J.D.; Perna, N.T. Reordering contigs of draft genomes using the Mauve aligner. Bioinformatics 2009, 25, 2071–2073. [Google Scholar] [CrossRef]
- Tamura, K. Estimation of the number of nucleotide substitutions when there are strong transition-transversion and G+C-content biases. Mol. Biol. Evol. 1992, 9, 678–687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, Z.; Gu, L.; Eils, R.; Schlesner, M.; Brors, B. circlize Implements and enhances circular visualization in R. Bioinformatics 2014, 30, 2811–2812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matti, K.; Bernardi, B.; Brezina, S.; Semmler, H.; von Wallbrunn, C.; Rauhut, D.; Wendland, J. Characterization of old wine yeasts kept for decades under a zero-emission maintenance regime. Fermentation 2020, 6, 9. [Google Scholar] [CrossRef] [Green Version]
- Surmacki, A.; Ozarowska-Nowicka, A.; Rosin, Z.M. Color polymorphism in a land snail Cepaea nemoralis (Pulmonata: Helicidae) as viewed by potential avian predators. Naturwissenschaften 2013, 100, 533–540. [Google Scholar] [CrossRef] [Green Version]
- Davison, A.; Jackson, H.J.; Murphy, E.W.; Reader, T. Discrete or indiscrete? Redefining the colour polymorphism of the land snail Cepaea nemoralis. Heredity 2019, 123, 162–175. [Google Scholar] [CrossRef] [Green Version]
- Wirth, F.; Goldani, L.Z. Epidemiology of Rhodotorula: An emerging pathogen. Interdiscip. Perspect. Infect. Dis. 2012, 2012, 465717. [Google Scholar] [CrossRef] [Green Version]
- Cleveland, A.A.; Harrison, L.H.; Farley, M.M.; Hollick, R.; Stein, B.; Chiller, T.M.; Lockhart, S.R.; Park, B.J. Declining incidence of candidemia and the shifting epidemiology of Candida resistance in two US metropolitan areas, 2008–2013: Results from population-based surveillance. PLoS ONE 2015, 10, e0120452. [Google Scholar] [CrossRef] [Green Version]
- Setati, M.E.; Jacobson, D.; Andong, U.C.; Bauer, F.F. The vineyard yeast microbiome, a mixed model microbial map. PLoS ONE 2012, 7, e52609. [Google Scholar] [CrossRef] [Green Version]
- Varela, C.; Borneman, A.R. Yeasts found in vineyards and wineries. Yeast 2017, 34, 111–128. [Google Scholar] [CrossRef]
- Viana, F.; Gil, J.V.; Genoves, S.; Valles, S.; Manzanares, P. Rational selection of non-Saccharomyces wine yeasts for mixed starters based on ester formation and enological traits. Food Microbiol. 2008, 25, 778–785. [Google Scholar] [CrossRef]
- Brysch-Herzberg, M. Ecology of yeasts in plant-bumblebee mutualism in Central Europe. FEMS Microbiol. Ecol. 2004, 50, 87–100. [Google Scholar] [CrossRef]
- Guzman, B.; Lachance, M.A.; Herrera, C.M. Phylogenetic analysis of the angiosperm-floricolous insect-yeast association: Have yeast and angiosperm lineages co-diversified? Mol. Phylogenet. Evol. 2013, 68, 161–175. [Google Scholar] [CrossRef] [Green Version]
- Blackwell, M. Made for Each Other: Ascomycete Yeasts and Insects. Microbiol. Spectr. 2017, 5. [Google Scholar] [CrossRef]
- Keebaugh, E.S.; Yamada, R.; Ja, W.W. The nutritional environment influences the impact of microbes on drosophila melanogaster life span. mBio 2019, 10. [Google Scholar] [CrossRef] [Green Version]
- Qiao, H.; Keesey, I.W.; Hansson, B.S.; Knaden, M. Gut microbiota affects development and olfactory behavior in Drosophila melanogaster. J. Exp. Biol. 2019, 222. [Google Scholar] [CrossRef] [Green Version]
- Gabaldon, T.; Fairhead, C. Genomes shed light on the secret life of Candida glabrata: Not so asexual, not so commensal. Curr. Genet. 2019, 65, 93–98. [Google Scholar] [CrossRef] [Green Version]
- Opulente, D.A.; Langdon, Q.K.; Buh, K.V.; Haase, M.A.B.; Sylvester, K.; Moriarty, R.V.; Jarzyna, M.; Considine, S.L.; Schneider, R.M.; Hittinger, C.T. Pathogenic budding yeasts isolated outside of clinical settings. FEMS Yeast Res. 2019, 19. [Google Scholar] [CrossRef]
- Al-Yasiri, M.H.; Normand, A.C.; L’Ollivier, C.; Lachaud, L.; Bourgeois, N.; Rebaudet, S.; Piarroux, R.; Mauffrey, J.F.; Ranque, S. Opportunistic fungal pathogen Candida glabrata circulates between humans and yellow-legged gulls. Sci. Rep. 2016, 6, 36157. [Google Scholar] [CrossRef] [Green Version]
- Chenot, C.; Briffoz, L.; Lomartire, A.; Collin, S. Occurrence of ehrlich-derived and varietal polyfunctional thiols in belgian white wines made from chardonnay and solaris grapes. J. Agric. Food Chem. 2019. [Google Scholar] [CrossRef]
- Tristezza, M.; Tufariello, M.; Capozzi, V.; Spano, G.; Mita, G.; Grieco, F. The oenological potential of hanseniaspora uvarum in simultaneous and sequential co-fermentation with saccharomyces cerevisiae for industrial wine production. Front. Microbiol. 2016, 7, 670. [Google Scholar] [CrossRef] [Green Version]
- Bora, S.S.; Keot, J.; Das, S.; Sarma, K.; Barooah, M. Metagenomics analysis of microbial communities associated with a traditional rice wine starter culture (Xaj-pitha) of Assam, India. 3 Biotech. 2016, 6, 153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, L.X.; Wang, G.Q.; Aihaiti, A. Combined indigenous yeast strains produced local wine from over ripen Cabernet Sauvignon grape in Xinjiang. World J. Microbiol. Biotechnol. 2020, 36, 122. [Google Scholar] [CrossRef] [PubMed]
- Bekatorou, A.; Psarianos, C.; Koutinas, A.A. Production of food grade yeasts. Food Technol. Biotechnol. 2006, 44, 407–415. [Google Scholar]
- Douglass, A.P.; Offei, B.; Braun-Galleani, S.; Coughlan, A.Y.; Martos, A.A.R.; Ortiz-Merino, R.A.; Byrne, K.P.; Wolfe, K.H. Population genomics shows no distinction between pathogenic Candida krusei and environmental Pichia kudriavzevii: One species, four names. PLoS Pathog. 2018, 14, e1007138. [Google Scholar] [CrossRef]
- Kieliszek, M.; Kot, A.M.; Bzducha-Wróbel, A.; BŁażejak, S.; Gientka, I.; Kurcz, A. Biotechnological use of Candida yeasts in the food industry: A review. Fungal Biol. Rev. 2017, 31, 185–198. [Google Scholar] [CrossRef]
- Papon, N.; Courdavault, V.; Clastre, M.; Bennett, R.J. Emerging and emerged pathogenic Candida species: Beyond the Candida albicans paradigm. PLoS Pathog. 2013, 9, e1003550. [Google Scholar] [CrossRef] [Green Version]
- Steenwyk, J.L.; Opulente, D.A.; Kominek, J.; Shen, X.X.; Zhou, X.; Labella, A.L.; Bradley, N.P.; Eichman, B.F.; Cadez, N.; Libkind, D.; et al. Extensive loss of cell-cycle and DNA repair genes in an ancient lineage of bipolar budding yeasts. PLoS Biol. 2019, 17, e3000255. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Cepaea snails promoting yeast dispersal in the environment. (A). A specimen of Cepaea on a cab. (B) A Cepaea snail found in the vineyard on ripe grapes.
Figure 1.
Cepaea snails promoting yeast dispersal in the environment. (A). A specimen of Cepaea on a cab. (B) A Cepaea snail found in the vineyard on ripe grapes.

Figure 2.
Isolation of yeasts from Cepaea snails. (A) Snails were isolated in Falcon tubes until they delivered excrements (encircled). Their confinement lasted for several hours up to overnight. (B) Dilution plating of solubilized snail excrements on YPD petri plates. Different sample plates are shown. Either yeasts could be picked or—due to overgrowth by filamentous fungi—cells were micromanipulated from small yeast colonies after overnight incubation at 30 °C.
Figure 2.
Isolation of yeasts from Cepaea snails. (A) Snails were isolated in Falcon tubes until they delivered excrements (encircled). Their confinement lasted for several hours up to overnight. (B) Dilution plating of solubilized snail excrements on YPD petri plates. Different sample plates are shown. Either yeasts could be picked or—due to overgrowth by filamentous fungi—cells were micromanipulated from small yeast colonies after overnight incubation at 30 °C.

Figure 3.
Fungal diversity obtained from snail excrements. (A) Fungal isolates were identified by ITS-PCR sequencing. (B) 165 fungal isolates were obtained from snail (Cepaea spp.) excrements among them 77 ascomycetous and 88 basidiomycetous yeasts. Human pathogens identified are highlighted in red. Outermost track: Bar height is proportionate to the numbers of isolates (shown inside the bars) obtained for each genus. Species names are provided when only a single species was identified for a genus. Middle track: Families; segment height is proportionate to the number of associated genera; key to families: F1—Herpotrichiellaceae; F2—Saccotheciaceae; F—Mycosphaerellaceae; F4—Debaryomyceta-ceae; F5—Metschnikowiaceae; F6—Pichiaceae; F7—Saccharomycetaceae; F8—Saccharomycodaceae; F9—unplaced (Diutina); F10—unplaced (Tumularia); F11—Cystobasidiaceae; F12—Cystofilobasidiaceae; F13—Filobasidiaceae; F14—Holtermanniella; F15—Leucosporidium; F16—Chrysozymaceae; F17—Curvibasidium; F18—Sporidiobolaceae; F19—Bulleribasidiaceae; F20—Rhynchogastremataceae; F21—Trimorphomycetaceae. Innermost track: phyla. The graphs of Figure 3B and Figure 4 were generated using package circlize version 0.4.10 in R version 3.6.3 [34]. Taxonomy follows NCBI’s taxonimic backbone (as of September 5, 2020): Ca. lusitaniae thus appears in genus Clavispora (Metschnikowiaceae).
Figure 3.
Fungal diversity obtained from snail excrements. (A) Fungal isolates were identified by ITS-PCR sequencing. (B) 165 fungal isolates were obtained from snail (Cepaea spp.) excrements among them 77 ascomycetous and 88 basidiomycetous yeasts. Human pathogens identified are highlighted in red. Outermost track: Bar height is proportionate to the numbers of isolates (shown inside the bars) obtained for each genus. Species names are provided when only a single species was identified for a genus. Middle track: Families; segment height is proportionate to the number of associated genera; key to families: F1—Herpotrichiellaceae; F2—Saccotheciaceae; F—Mycosphaerellaceae; F4—Debaryomyceta-ceae; F5—Metschnikowiaceae; F6—Pichiaceae; F7—Saccharomycetaceae; F8—Saccharomycodaceae; F9—unplaced (Diutina); F10—unplaced (Tumularia); F11—Cystobasidiaceae; F12—Cystofilobasidiaceae; F13—Filobasidiaceae; F14—Holtermanniella; F15—Leucosporidium; F16—Chrysozymaceae; F17—Curvibasidium; F18—Sporidiobolaceae; F19—Bulleribasidiaceae; F20—Rhynchogastremataceae; F21—Trimorphomycetaceae. Innermost track: phyla. The graphs of Figure 3B and Figure 4 were generated using package circlize version 0.4.10 in R version 3.6.3 [34]. Taxonomy follows NCBI’s taxonimic backbone (as of September 5, 2020): Ca. lusitaniae thus appears in genus Clavispora (Metschnikowiaceae).

Figure 4.
(A–C). Genomic alignments of three non-conventional yeast isolates, Hanseniaspora uvarum B079, Pichia kudriavzevii B143, Meyerozyma guilliermondii B144, against suitable reference genomes. The left part (red and very dark red) of the outer ring represents the snail yeast isolates, while the right parts (in grey) represent scaffolds of the reference genomes. The draft genomes were ordered using Mauve Contig Mover (MCM); for snail yeast genomes the extent of contigs >10 kb are shown in bright red; contigs >10 kb (100 kb for B079) are identified by numbers; alignment gaps are shown if they were >2.5 kb. In the middle ring, the divergence of the draft genomes to reference genomes (calculated based on consecutive 1 kb windows) is shown (indicating low divergence, i.e., high sequence identity). Only windows with at least 90% nucleotide coverage (900 nt) are shown; points are also colour-coded with higher-divergence tending towards yellow. The Inner ring shows Locally Collinear Blocks (LCB) with a minimum length of 50 kb. LCBs are colour-coded and enumerated; transparency of genomic links connecting LCBs on either genome decreases as mismatches between relative positions on each genome increases (i.e., translocation events are less transparent).
Figure 4.
(A–C). Genomic alignments of three non-conventional yeast isolates, Hanseniaspora uvarum B079, Pichia kudriavzevii B143, Meyerozyma guilliermondii B144, against suitable reference genomes. The left part (red and very dark red) of the outer ring represents the snail yeast isolates, while the right parts (in grey) represent scaffolds of the reference genomes. The draft genomes were ordered using Mauve Contig Mover (MCM); for snail yeast genomes the extent of contigs >10 kb are shown in bright red; contigs >10 kb (100 kb for B079) are identified by numbers; alignment gaps are shown if they were >2.5 kb. In the middle ring, the divergence of the draft genomes to reference genomes (calculated based on consecutive 1 kb windows) is shown (indicating low divergence, i.e., high sequence identity). Only windows with at least 90% nucleotide coverage (900 nt) are shown; points are also colour-coded with higher-divergence tending towards yellow. The Inner ring shows Locally Collinear Blocks (LCB) with a minimum length of 50 kb. LCBs are colour-coded and enumerated; transparency of genomic links connecting LCBs on either genome decreases as mismatches between relative positions on each genome increases (i.e., translocation events are less transparent).

Figure 5.
Fermentation curves based on CO2 release/weight loss (measured daily) of non-conventional yeasts (H. uvarum, M. guilliermondii, P. kudriavzevii) and the EC1118 wine yeast plotted over time.
Figure 5.
Fermentation curves based on CO2 release/weight loss (measured daily) of non-conventional yeasts (H. uvarum, M. guilliermondii, P. kudriavzevii) and the EC1118 wine yeast plotted over time.

Figure 6.
Ester aroma compound formation of non-conventional yeasts compared to the EC1118 wine yeast. Aroma compounds were quantified by gas chromatography at the end of fermentation (see Figure 5).
Figure 6.
Ester aroma compound formation of non-conventional yeasts compared to the EC1118 wine yeast. Aroma compounds were quantified by gas chromatography at the end of fermentation (see Figure 5).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
HPLC analyses of residual sugars, organic acids and total alcohol.
| Glucose | Fructose | Total Sugars | Tartaric Acid | Malate | Shikimic Acid | Lactate | Acetate | Citric Acid | Ethanol | Ethanol | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| strain | [g/L] | [g/L] | [g/L] | [g/L] | [g/L] | [mg/L] | [g/L] | [g/L] | [g/L] | [g/L] | [%] |
| B079 (H. uvarum) | <1 | <1 | 0 | 4.1 ± 0.1 | 2.4 ± 0.1 | 28.3 ± 0.6 | 0.2 ± 0 | 0.2 ± 0.1 | 0.2 ± 0 | 79.7 ± 0.4 | 10.1 ± 0.1 |
| B143 (P. kudriavzevii) | <1 | 16.1 ± 2.1 | 16.1 ± 2.1 | 4.2 ± 0.1 | 1.7 ± 0 | 32 ± 0 | 0.2 ± 0 | 0.8 ± 0.1 | 0.2 ± 0 | 66.7 ± 0.4 | 8.4 ± 0.1 |
| B144 (M. guilliermondii) | 1 | 18.5 ± 6.5 | 18.8 ± 7.1 | 4.2 ± 0 | 1.8 ± 0.1 | 32 ± 0 | 0.2 ± 0 | 0.7 ± 0.1 | 0.2 ± 0 | 66.2 ± 3.4 | 8.4 ± 0.5 |
| EC1118 (S. cerevisiae) | 1.6 | 18.6 | 20.2 | 3.7 | 2.7 | 31 | 0.12 | 0.39 | 0.13 | 77.7 | 9.9 |
Table 2.
Volatile aroma analysis of alcohols and esters.
| Strain | Acetic Acid Ethylester [mg/L] | i-Butanol [mg/L] | Propionic Acid Ethylester [µg/L] | Isoamyl-Alcohol [mg/L] | 2-Methyl-Butanol [mg/L] | i-Butyric Acid Ethylester [µg/L] | Butyric Acid Ethylester [µg/L] | i-Valeric Acid [µg/L] | Hexanol [µg/L] | Isoamyl-Acetate [µg/L] | Acetic Acid 2-Methyl-Butylester [µg/L] | 2-Phenyl-Ethanol [mg/L] | 2-Phenyl Acetate [µg/L] |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| B079 (H. uvarum) | 425 ± 51 | 251 ± 61 | 1459 ± 541 | 375 ± 60 | 89 ± 19 | 1773 ± 247 | 114 ± 24 | 2074 ± 15 | 884 ± 93 | 251 ± 132 | 28 ± 6 | 48 ± 12 | 100 ± 18 |
| B143 (P. kudriavzevii) | 868 ± 213 | 494 ± 64 | 3002 ± 609 | 361 ± 47 | 104 ± 11 | 1512 ± 173 | 46 ± 7 | 2479 ± 195 | 436 ± 39 | 1878 ± 457 | 92 ± 27 | 58 ± 7 | 275 ± 9 |
| B144 (M. guilliermondii) | 883 ± 68 | 497 ± 62 | 3075 ± 534 | 355 ± 32 | 104 ± 8 | 1361 ± 231 | 51 ± 3 | 2090 ± 90 | 438 ± 0 | 1932 ± 519 | 97 ± 15 | 56 ± 3 | 278 ± 21 |
| EC1118 (S. cerevisiae) | 36 ± 4 | 60 ± 5 | 438 ± 15 | 402 ± 35 | 71 ± 4 | 38 ± 4 | 137 ± 25 | 1668 ± 103 | 1542 ± 29 | 540 ± 127 | 36 ± 4 | 57 ± 4 | 98 ± 2 |
For B079, B143 and B144, fermentations were run as triplicates; for EC1118 fermentations were run as duplicates. Highlighted ester compounds are also shown in Figure 6.
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Akan, M.; Michling, F.; Matti, K.; Krause, S.; Muno-Bender, J.; Wendland, J. Snails as Taxis for a Large Yeast Biodiversity. Fermentation 2020, 6, 90. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation6030090
AMA Style
Akan M, Michling F, Matti K, Krause S, Muno-Bender J, Wendland J. Snails as Taxis for a Large Yeast Biodiversity. Fermentation. 2020; 6(3):90. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation6030090
Chicago/Turabian StyleAkan, Madina, Florian Michling, Katrin Matti, Sinje Krause, Judith Muno-Bender, and Jürgen Wendland. 2020. "Snails as Taxis for a Large Yeast Biodiversity" Fermentation 6, no. 3: 90. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation6030090
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.