
Supplementing Glycerol to Inoculum Induces Changes in pH, SCFA Profiles, and Microbiota Composition in In-Vitro Batch Fermentation

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Substrates
2.2. Preparation of Inoculum and Standardization
2.3. Medium Solution
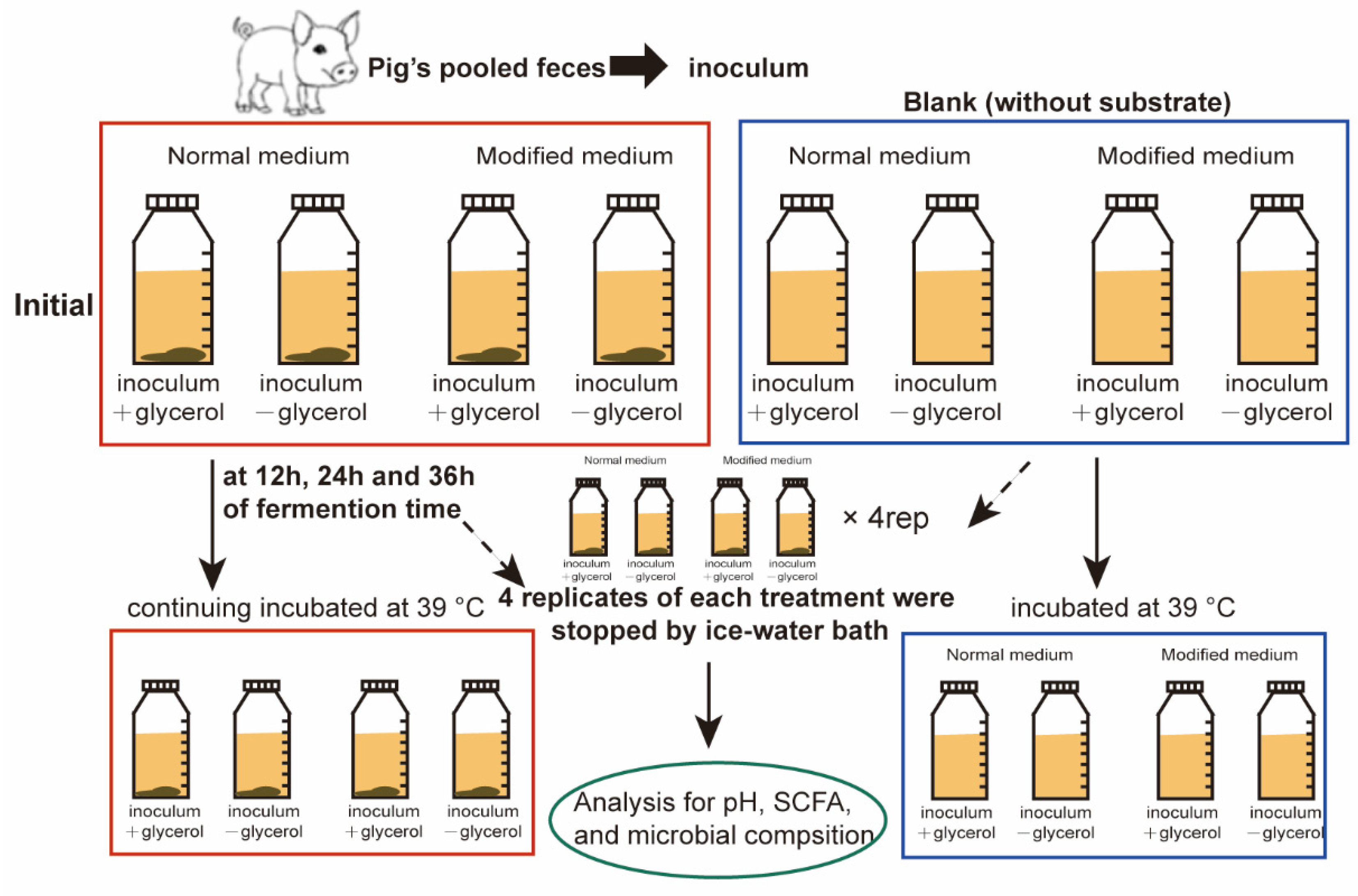
2.4. Experimental Design and In Vitro Fermentation Procedures
2.5. Sample Detection
2.5.1. Chemical Analysis
2.5.2. Measurement of SCFA Production
2.5.3. Microbiota Composition Analysis
2.6. Statistical Analysis
3. Results
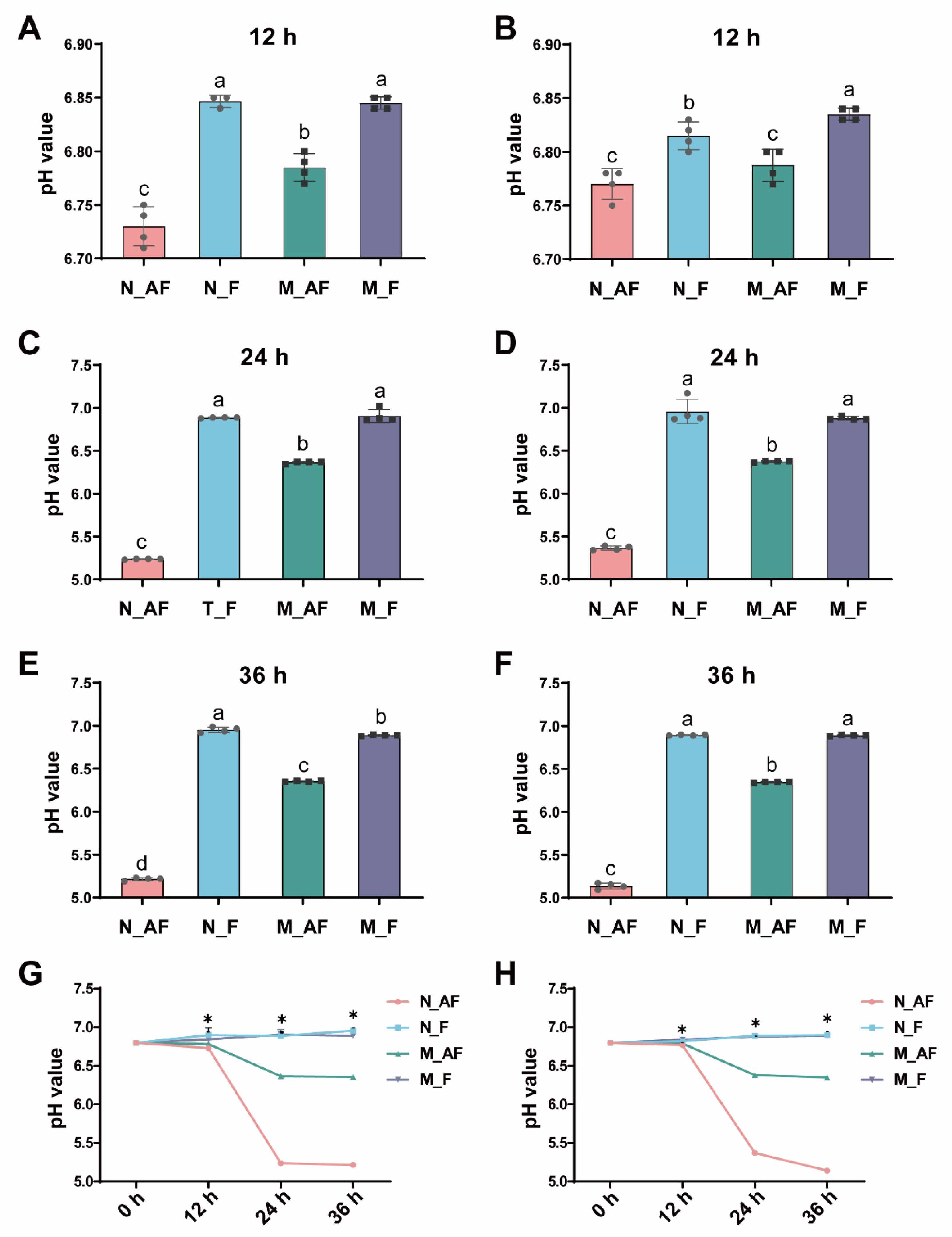
3.1. Changes of pH Values in Fermentation Broth during In-Vitro Fermentation
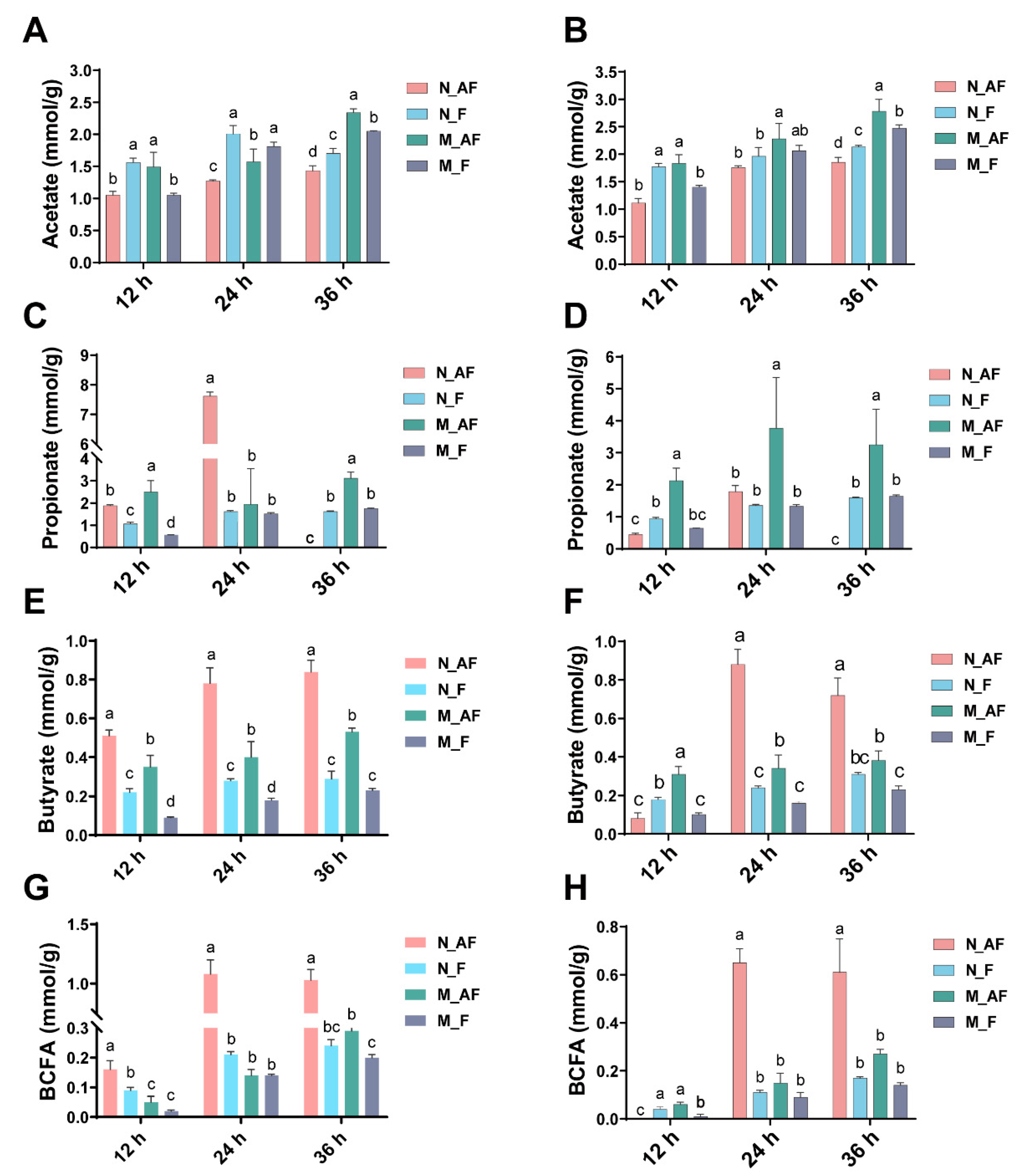
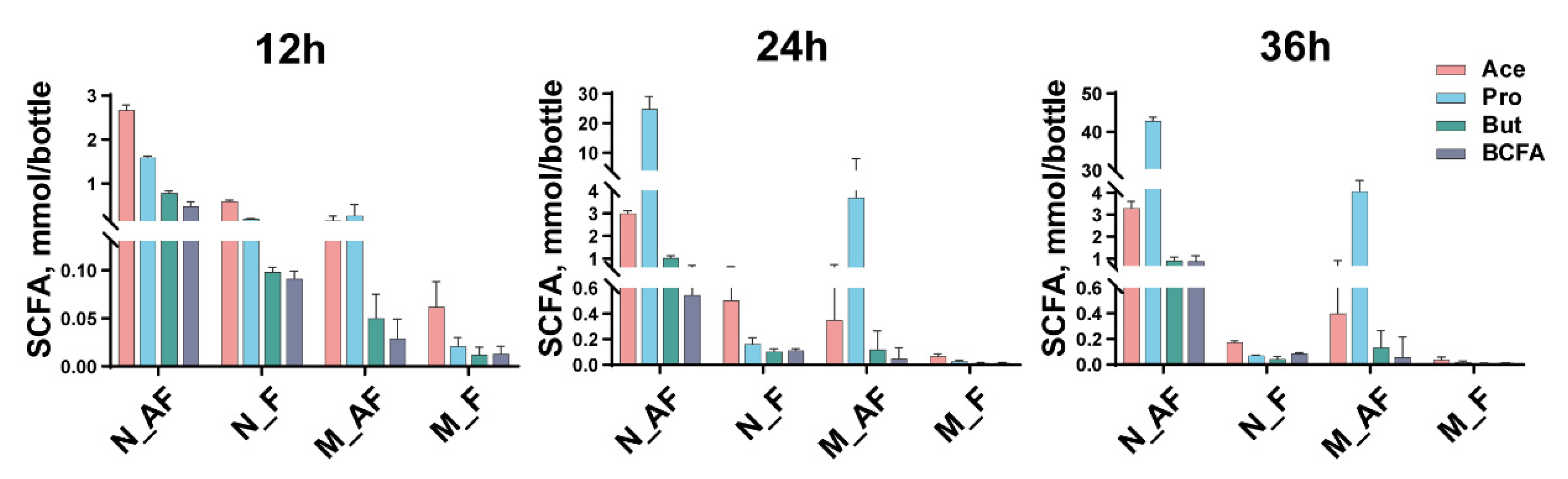
3.2. SCFAs Profiles during Fermentation among Different Medium and Inoculum Treatments
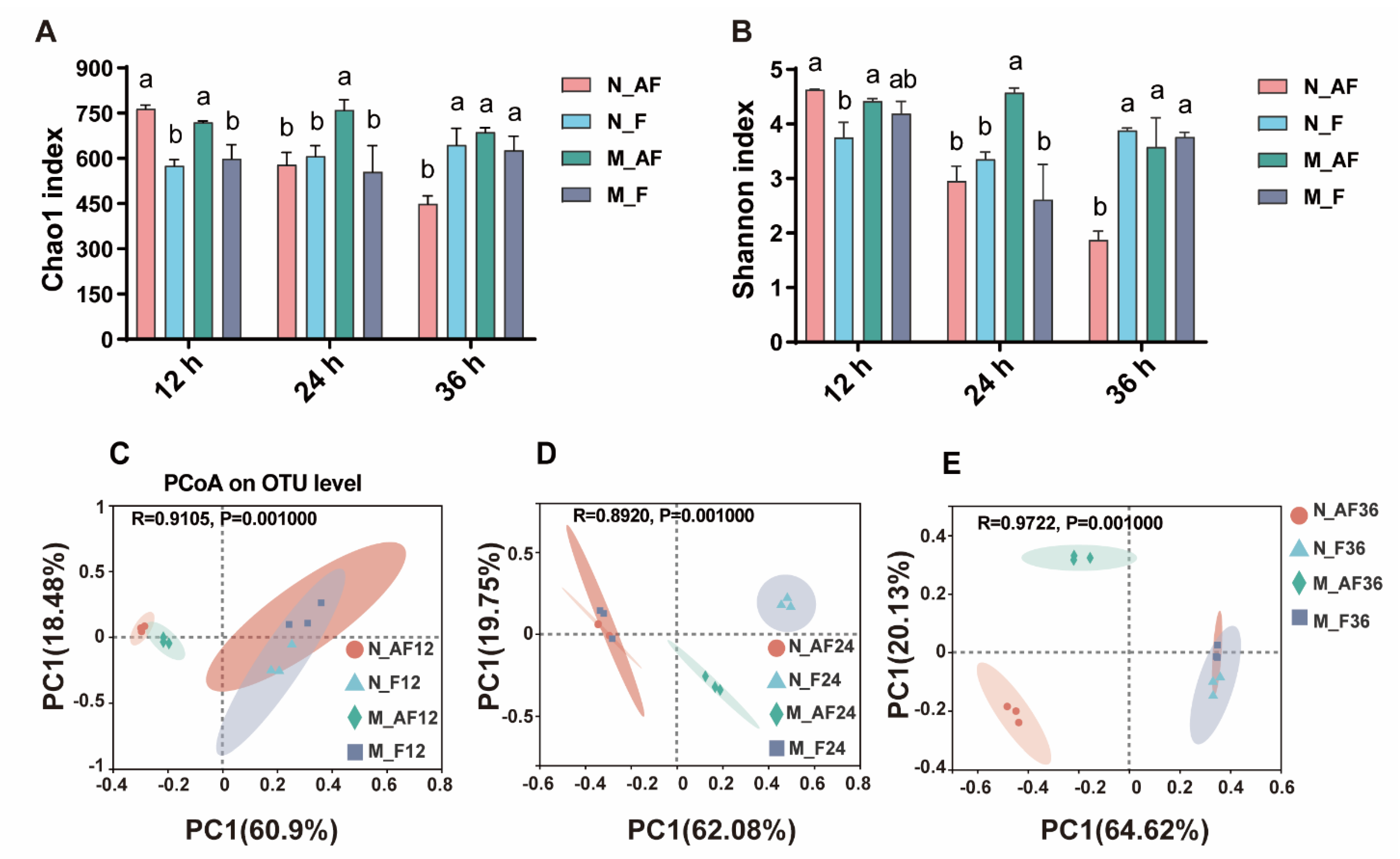
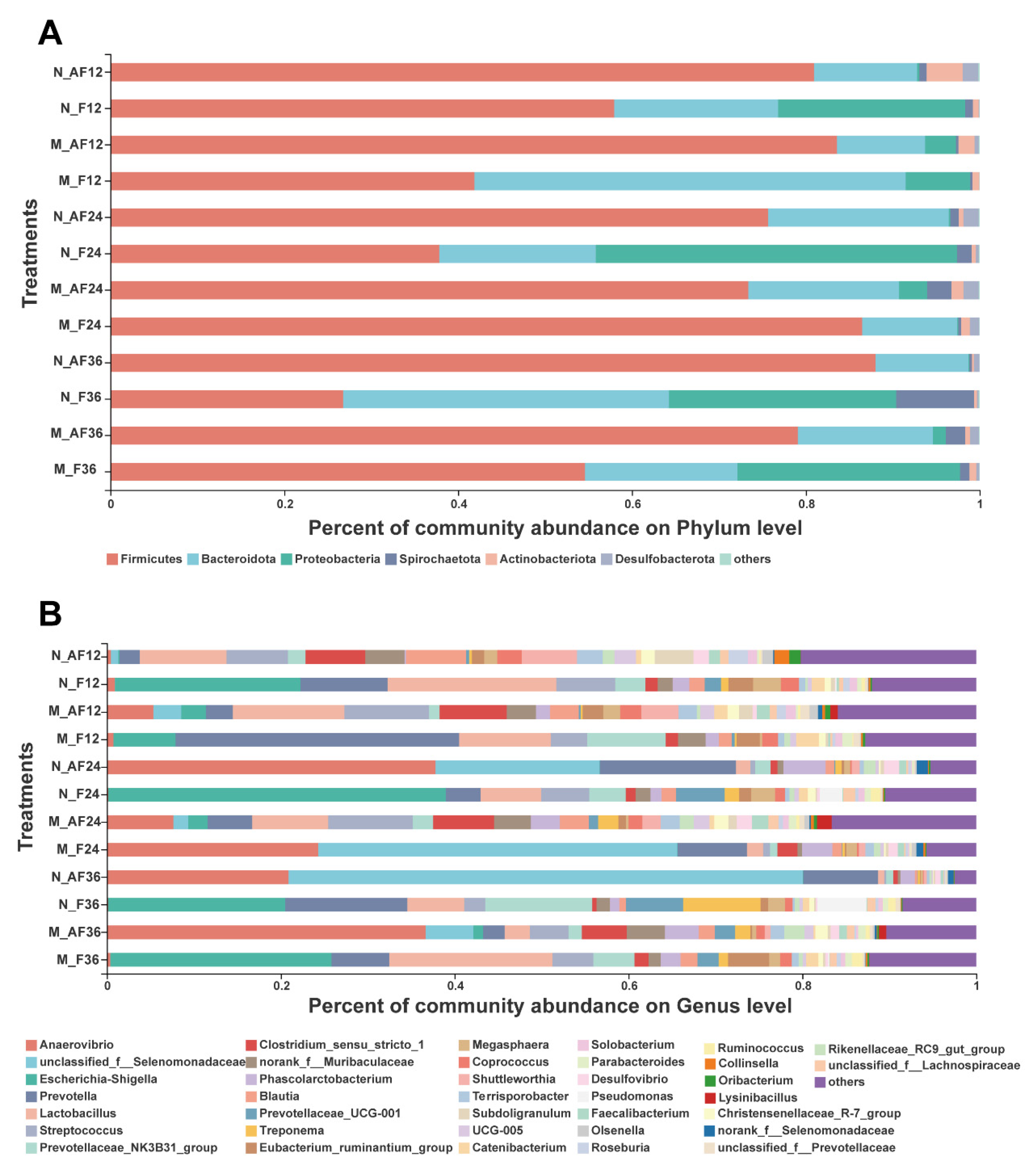
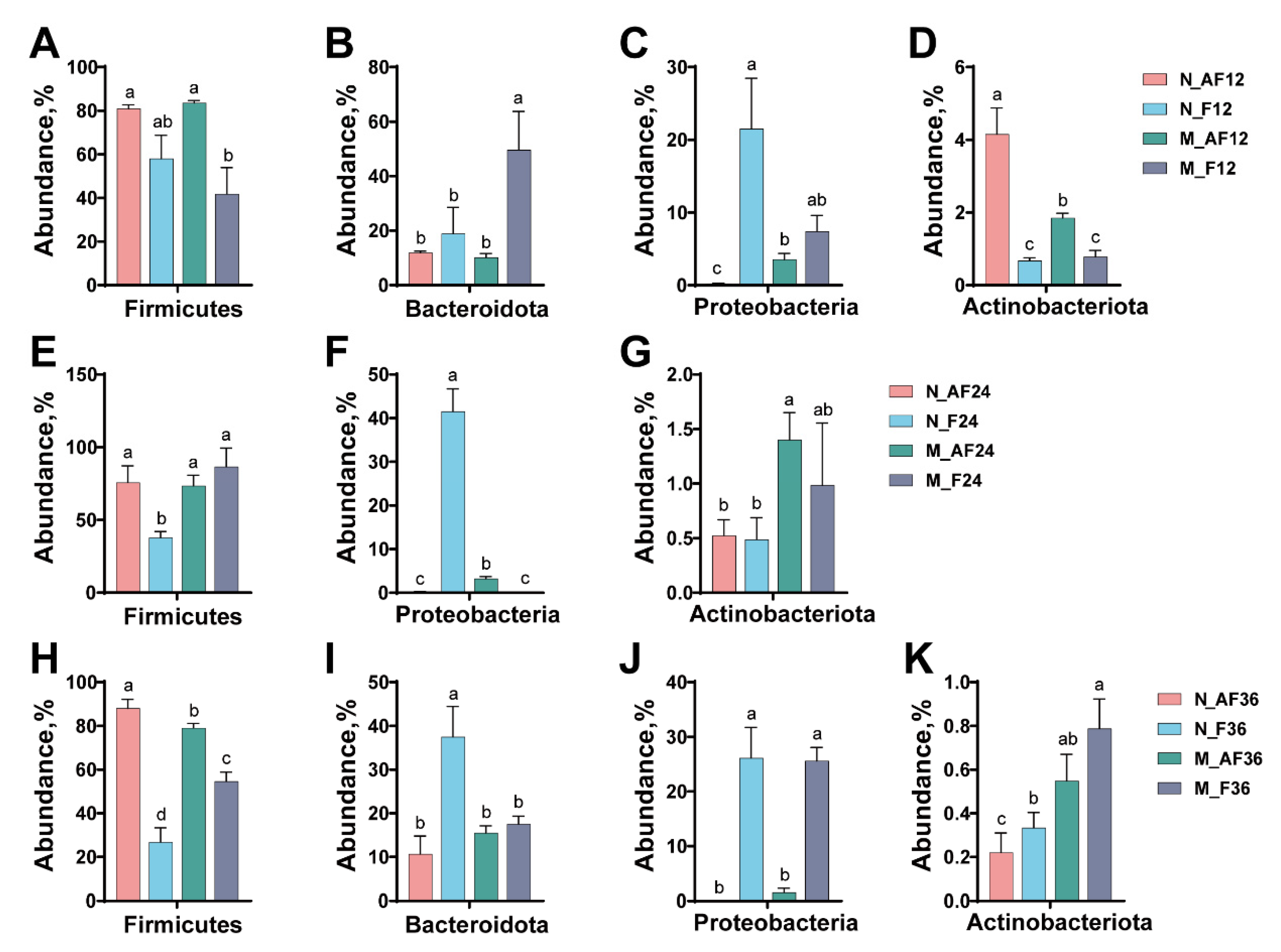
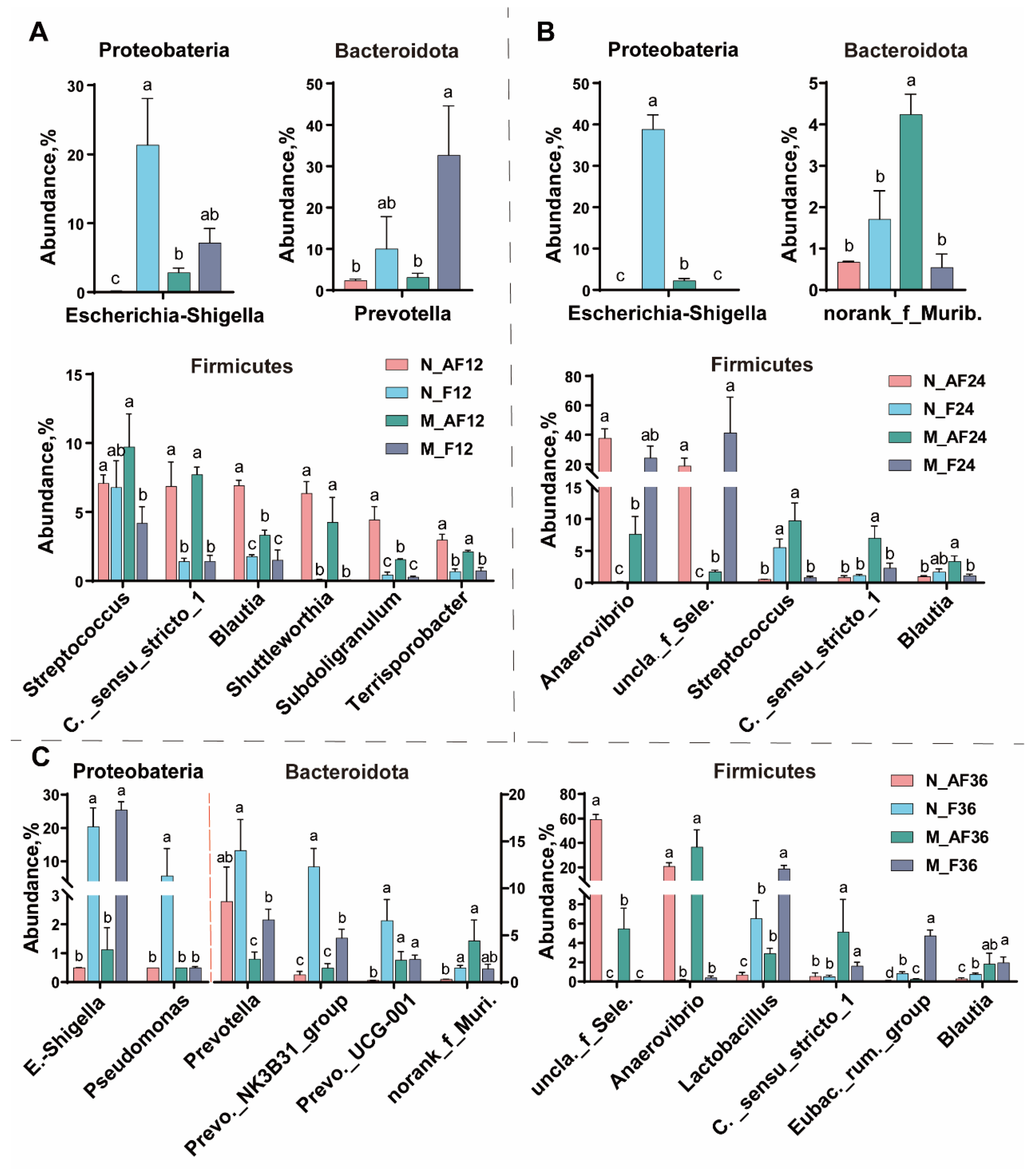
3.3. The Changes in the Microbial Composition during Fermentation
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- de Leeuw, J.A.; Bolhuis, J.E.; Bosch, G.; Gerrits, W.J. Effects of dietary fibre on behaviour and satiety in pigs. Proc. Nutr. Soc. 2008, 67, 334–342. [Google Scholar] [CrossRef] [Green Version]
- Tan, C.; Wei, H.; Zhao, X.; Xu, C.; Zhou, Y.; Peng, J. Soluble Fiber with High Water-Binding Capacity, Swelling Capacity, and Fermentability Reduces Food Intake by Promoting Satiety Rather Than Satiation in Rats. Nutrients 2016, 8, 615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, W.; Xie, J.; Zhang, H. Dietary fibers influence the intestinal SCFAs and plasma metabolites profiling in growing pigs. Food Funct. 2016, 7, 4644–4654. [Google Scholar] [CrossRef]
- Seo, Y.S.; Lee, H.-B.; Kim, Y.; Park, H.-Y. Dietary Carbohydrate Constituents Related to Gut Dysbiosis and Health. Microorganisms 2020, 8, 427. [Google Scholar] [CrossRef] [Green Version]
- Gill, S.K.; Rossi, M.; Bajka, B.; Whelan, K. Dietary fibre in gastrointestinal health and disease. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 101–116. [Google Scholar] [CrossRef]
- Lynd, L.R.; Weimer, P.J.; van Zyl, W.H.; Pretorius, I.S. Microbial cellulose utilization: Fundamentals and biotechnology. Microbiol. Mol. Biol. Rev. 2002, 66, 506–577. [Google Scholar] [CrossRef] [Green Version]
- Bayer, E.A.; Belaich, J.P.; Shoham, Y.; Lamed, R. The cellulosomes: Multienzyme machines for degradation of plant cell wall polysaccharides. Annu. Rev. Microbiol. 2004, 58, 521–554. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Bai, Y.; Tao, S.; Zhang, G.; Wang, J.; Liu, L.; Zhang, S. Fiber-rich foods affected gut bacterial community and short-chain fatty acids production in pig model. J. Funct. Foods 2019, 57, 266–274. [Google Scholar] [CrossRef]
- Rymer, C.; Huntington, J.; Williams, B.; Givens, D. In vitro cumulative gas production techniques: History, methodological considerations and challenges. Anim. Feed Sci. Technol. 2005, 123, 9–30. [Google Scholar] [CrossRef]
- Wang, Y.M. Effects and Mechanisms of Dietary Crude Protein Level on Intestinal Nutrient Digestion and Metabolism in Pigs. Ph.D. Thesis, China Agriculture University, Beijing, China, 2020. [Google Scholar]
- Bai, Y.; Zhou, X.; Li, N.; Zhao, J.; Ye, H.; Zhang, S.; Yang, H.; Pi, Y.; Tao, S.; Han, D.; et al. In vitro fermentation characteristics and fiber-degrading enzyme kinetics of cellulose, arabinoxylan, β-glucan and glucomannan by pig fecal microbiota. Microorganisms 2021, 9, 1071. [Google Scholar] [CrossRef]
- Chen, Y.; Wang, T.; Shen, N.; Zhang, F.; Zeng, R.J. High-purity propionate production from glycerol in mixed culture fermentation. Bioresour. Technol. 2016, 219, 659–667. [Google Scholar] [CrossRef] [PubMed]
- Trabue, S.; Scoggin, K.; Tjandrakusuma, S.; Rasmussen, M.A.; Reilly, P.J. Ruminal fermentation of propylene glycol and glycerol. J. Agric. Food Chem. 2007, 55, 7043–7051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Cleef, E.H.C.B.; Uwituze, S.; Alvarado-Gilis, C.A.; Miller, K.A.; Van Bibber-Krueger, C.L.; Aperce, C.C.; Drouillard, J.S. Elevated concentrations of crude glycerin in diets for beef cattle: Feedlot performance, carcass traits, and ruminal metabolism1. J. Anim. Sci. 2019, 97, 4341–4348. [Google Scholar] [CrossRef] [PubMed]
- Menke, K.H.; Steingass, H. Estimation of the energetic feed value obtained from chemical analysis and in vitro gas production using rumen fluid. Anim. Res. Dev. 1988, 28, 7–55. [Google Scholar]
- Bai, Y.; Zhao, J.B.; Tao, S.Y.; Zhou, X.J.; Pi, Y.; Gerrits, W.J.; Johnston, L.J.; Zhang, S.Y.; Yang, H.J.; Liu, L.; et al. Effect of dietary fiber fermentation on short-chain fatty acid production and microbial composition in vitro. J. Sci. Food Agric. 2020, 100, 4282–4291. [Google Scholar] [CrossRef]
- Yu, J.; Fu, Y.; Deng, Z.; Fan, Y.; Li, H. Effects of soluble dietary fiber from soybean residue fermented by Neurospora crassa on the intestinal flora in rats. Food Funct. 2020, 11, 7433–7445. [Google Scholar] [CrossRef]
- Wong, J.M.; de Souza, R.; Kendall, C.W.; Emam, A.; Jenkins, D.J. Colonic health: Fermentation and short chain fatty acids. J. Clin. Gastroenterol. 2006, 40, 235–243. [Google Scholar] [CrossRef]
- Jha, R.; Rossnagel, B.; Pieper, R.; Van Kessel, A.; Leterme, P. Barley and oat cultivars with diverse carbohydrate composition alter ileal and total tract nutrient digestibility and fermentation metabolites in weaned piglets. Animal 2010, 4, 724–731. [Google Scholar] [CrossRef] [Green Version]
- Tan, Z.K. Study on the Correlation between Roughage-Resistance and Intestinal Microecological Factors of Tibetan Pigs. Ph.D. Thesis, Tibet University, Xizang, China, 2021. [Google Scholar]
- Louis, P.; Flint, H.J. Formation of propionate and butyrate by the human colonic microbiota. Environ. Microbiol. 2017, 19, 29–41. [Google Scholar] [CrossRef] [Green Version]
- Zeng, H.; Umar, S.; Rust, B.; Lazarova, D.; Bordonaro, M. Secondary Bile Acids and Short Chain Fatty Acids in the Colon: A Focus on Colonic Microbiome, Cell Proliferation, Inflammation, and Cancer. Int. J. Mol. Sci. 2019, 20, 1214. [Google Scholar] [CrossRef] [Green Version]
- Marsono, Y.; Illman, R.J.; Clarke, J.M.; Trimble, R.P.; Topping, D.L. Plasma lipids and large bowel volatile fatty acids in pigs fed on white rice, brown rice and rice bran. Br. J. Nutr. 1993, 70, 503–513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uerlings, J.; Schroyen, M.; Bautil, A.; Courtin, C.; Richel, A.; Sureda, E.A.; Bruggeman, G.; Tanghe, S.; Willems, E.; Bindelle, J.; et al. In vitro prebiotic potential of agricultural by-products on intestinal fermentation, gut barrier and inflammatory status of piglets. Br. J. Nutr. 2020, 123, 293–307. [Google Scholar] [CrossRef]
- Bindelle, J.; Buldgen, A.; Boudry, C.; Leterme, P. Effect of inoculum and pepsin–pancreatin hydrolysis on fibre fermentation measured by the gas production technique in pigs. Anim. Feed Sci. Technol. 2007, 132, 111–122. [Google Scholar] [CrossRef]
- Jonathan, M.C.; van den Borne, J.J.; van Wiechen, P.; da Silva, C.S.; Schols, H.A.; Gruppen, H. In vitro fermentation of 12 dietary fibres by faecal inoculum from pigs and humans. Food Chem. 2012, 133, 889–897. [Google Scholar] [CrossRef]
- Liu, Y.; Li, Y.; Ke, Y.; Li, C.; Zhang, Z.; Wu, Y.; Hu, B.; Liu, A.; Luo, Q.; Wu, W. In vitro saliva-gastrointestinal digestion and fecal fermentation of Oudemansiella radicata polysaccharides reveal its digestion profile and effect on the modulation of the gut microbiota. Carbohydr. Polym. 2021, 251, 117041. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y. Study on Determination of Protein Availability in Cereal Grains Based on Simulating Gastro-Small Intestinal Digestion Process for Growing Pigs. Master’s Thesis, Chinese Academy of Agricultural Sciences, Beijing, China, 2019. [Google Scholar]
- Gao, Q.T.; Zhao, F.; Zhang, H.; Wang, Y.; Liu, B.L. Effect of dietary fiber levels and feeding time on passage rate of digesta in the intestinal tract of growing pigs. Chin. J. Anim. Nutr. 2018, 30, 469–476. [Google Scholar]
- AOAC. Official Methods of Analysis, 18th ed.; Association of Official Analytical Chemists: Arlington, TX, USA, 2007. [Google Scholar]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- Wu, W.; Zhang, L.; Xia, B.; Tang, S.; Xie, J.; Zhang, H. Modulation of Pectin on Mucosal Innate Immune Function in Pigs Mediated by Gut Microbiota. Microorganisms 2020, 8, 535. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naïve Bayesian Classifier for Rapid Assignment of rRNA Sequences into the New Bacterial Taxonomy. App.l Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [Green Version]
- Knudsen, K.B.; Jensen, B.B.; Andersen, J.; Hansen, I. Gastrointestinal implications in pigs of wheat and oat fractions: 2. Microbial activity in the gastrointestinal tract. Br. J. Nutr. 1991, 65, 233–248. [Google Scholar] [CrossRef] [Green Version]
- Collins, S.M.; Gibson, G.R.; Kennedy, O.B.; Walton, G.; Rowland, I.; Commane, D.M. Development of a prebiotic blend to influence in vitro fermentation effects, with a focus on propionate, in the gut. FEMS Microbiol. Ecol. 2021, 97, 101–111. [Google Scholar] [CrossRef]
- Montoya, C.A.; Rutherfurd, S.M.; Moughan, P.J. Ileal Digesta Nondietary Substrates from Cannulated Pigs Are Major Contributors to In Vitro Human Hindgut Short-Chain Fatty Acid Production. J. Nutr. 2017, 147, 264–271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tran, T.H.T.; Boudry, C.; Everaert, N.; Théwis, A.; Portetelle, D.; Daube, G.; Nezer, C.; Taminiau, B.; Bindelle, J. Adding mucins to an in vitro batch fermentation model of the large intestine induces changes in microbial population isolated from porcine feces depending on the substrate. FEMS Microbiol. Ecol. 2016, 92, fiv165. [Google Scholar] [CrossRef] [PubMed]
- Macfarlane, S.; Woodmansey, E.J.; Macfarlane, G.T. Colonization of mucin by human intestinal bacteria and establishment of biofilm communities in a two-stage continuous culture system. Appl. Environ. Microbiol. 2005, 71, 7483–7492. [Google Scholar] [CrossRef] [Green Version]
- Van Tassell, M.L.; Miller, M.J. Lactobacillus adhesion to mucus. Nutrients 2011, 3, 613–636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, C.; Tahir, N.; Li, W.; Zhang, Z.; Jiang, D.; Guo, S.; Wang, J.; Wang, K.; Zhang, Q. Enhanced buffer capacity of fermentation broth and biohydrogen production from corn stalk with NaHPO/NaHPO. Bioresour. Technol. 2020, 313, 783–788. [Google Scholar] [CrossRef]
- Mu, Y.; Teng, H.; Zhang, D.-J.; Wang, W.; Xiu, Z.L. Microbial production of 1,3-propanediol by Klebsiella pneumoniae using crude glycerol from biodiesel preparations. Biotechnol. Lett. 2006, 28, 1755–1759. [Google Scholar] [CrossRef]
- Bergner, H.; Kijora, C.; Ceresnakova, Z.; Szakacs, J. In vitro studies on glycerol transformation by rumen microorganisms. Arch. Tierernahr. 1995, 48, 245–256. [Google Scholar] [CrossRef]
- Ye, G.; Zhu, Y.; Liu, J.; Chen, X.; Huang, K. Preparation of glycerol-enriched yeast culture and its effect on blood metabolites and ruminal fermentation in goats. PLoS ONE 2014, 9, 410–418. [Google Scholar] [CrossRef] [Green Version]
- Ramayo-Caldas, Y.; Mach, N.; Lepage, P.; Levenez, F.; Denis, C.; Lemonnier, G.; Leplat, J.-J.; Billon, Y.; Berri, M.; Doré, J.; et al. Phylogenetic network analysis applied to pig gut microbiota identifies an ecosystem structure linked with growth traits. ISME J. 2016, 10, 2973–2977. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Chen, G.; Chen, D.; Ye, H.; Sun, Y.; Zeng, X.; Liu, Z. Purified fraction of polysaccharides from Fuzhuan brick tea modulates the composition and metabolism of gut microbiota in anaerobic fermentation in vitro. Int. J. Biol. Macromol. 2019, 140, 858–870. [Google Scholar] [CrossRef]
- Rui, Y.; Wan, P.; Chen, G.; Xie, M.; Sun, Y.; Zeng, X.; Liu, Z. Simulated digestion and fermentation in vitro by human gut microbiota of intra- and extra-cellular polysaccharides from Aspergillus cristatus. LWT-Food Sci. Technol. 2019, 116, 108508. [Google Scholar] [CrossRef]
- Long, C.; de Vries, S.; Venema, K. Differently Pre-treated Rapeseed Meals Affect in vitro Swine Gut Microbiota Composition. Front. Microbiol. 2020, 11, 570985. [Google Scholar] [CrossRef]
- Han, K.H.; Enomoto, M.; Pelpolage, S.; Nagata, R.; Fukuma, N.; Fukushima, M. In vitro fermentation potential of the residue of Korean red ginseng root in a mixed culture of swine faecal bacteria. Food Funct. 2020, 11, 6202–6214. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.H.; Steiner, T.; Petrof, E.O.; Smieja, M.; Roscoe, D.; Nematallah, A.; Weese, J.S.; Collins, S.; Moayyedi, P.; Crowther, M.; et al. Frozen vs Fresh Fecal Microbiota Transplantation and Clinical Resolution of Diarrhea in Patients With Recurrent Clostridium difficile Infection: A Randomized Clinical Trial. JAMA 2016, 315, 142–149. [Google Scholar] [CrossRef] [Green Version]
- Hu, J.; Chen, L.; Tang, Y.; Xie, C.; Xu, B.; Shi, M.; Zheng, W.; Zhou, S.; Wang, X.; Liu, L.; et al. Standardized Preparation for Fecal Microbiota Transplantation in Pigs. Front. Microbiol. 2018, 9, 1328–1337. [Google Scholar] [CrossRef] [PubMed]
- Fu, X.C.; Chen, A.L.; Zhang, Y.J.; Pang, X.Y. Protective effect of cryoprotectant on viable bacteria in fecal suspension. Genom. Appl. Biol. 2020, 39, 5586–5592. [Google Scholar]
- Zhang, L.; Xing, D.F.; Jiang, Q.Q.; Sun, R.; Ren, N.Q. Properties of cellulose degrading hydrogen producing bacteria flora in rumen fluid at different pH. J. Dalian Polytech. Univ. 2017, 36, 328–332. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item | DM | CP | NDF | ADF | ADL | TDF | IDF | SDF |
|---|---|---|---|---|---|---|---|---|
| Digesta of Barley diet | 92.67 | 10.70 | 38.58 | 14.23 | 2.55 | 46.64 | 36.93 | 9.71 |
| Digesta of Wheat shorts diet | 93.31 | 8.97 | 36.33 | 11.86 | 3.41 | 45.53 | 37.92 | 7.61 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, Q.; Li, K.; Zhong, R.; Long, C.; Liu, L.; Chen, L.; Zhang, H. Supplementing Glycerol to Inoculum Induces Changes in pH, SCFA Profiles, and Microbiota Composition in In-Vitro Batch Fermentation. Fermentation 2022, 8, 18. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation8010018
Gao Q, Li K, Zhong R, Long C, Liu L, Chen L, Zhang H. Supplementing Glycerol to Inoculum Induces Changes in pH, SCFA Profiles, and Microbiota Composition in In-Vitro Batch Fermentation. Fermentation. 2022; 8(1):18. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation8010018
Chicago/Turabian StyleGao, Qingtao, Kai Li, Ruqing Zhong, Cheng Long, Lei Liu, Liang Chen, and Hongfu Zhang. 2022. "Supplementing Glycerol to Inoculum Induces Changes in pH, SCFA Profiles, and Microbiota Composition in In-Vitro Batch Fermentation" Fermentation 8, no. 1: 18. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation8010018