Preliminary Estimates of Age, Growth and Natural Mortality of Margate, Haemulon album, and Black Margate, Anisotremus surinamensis, from the Southeastern United States

Abstract
:1. Introduction
2. Results
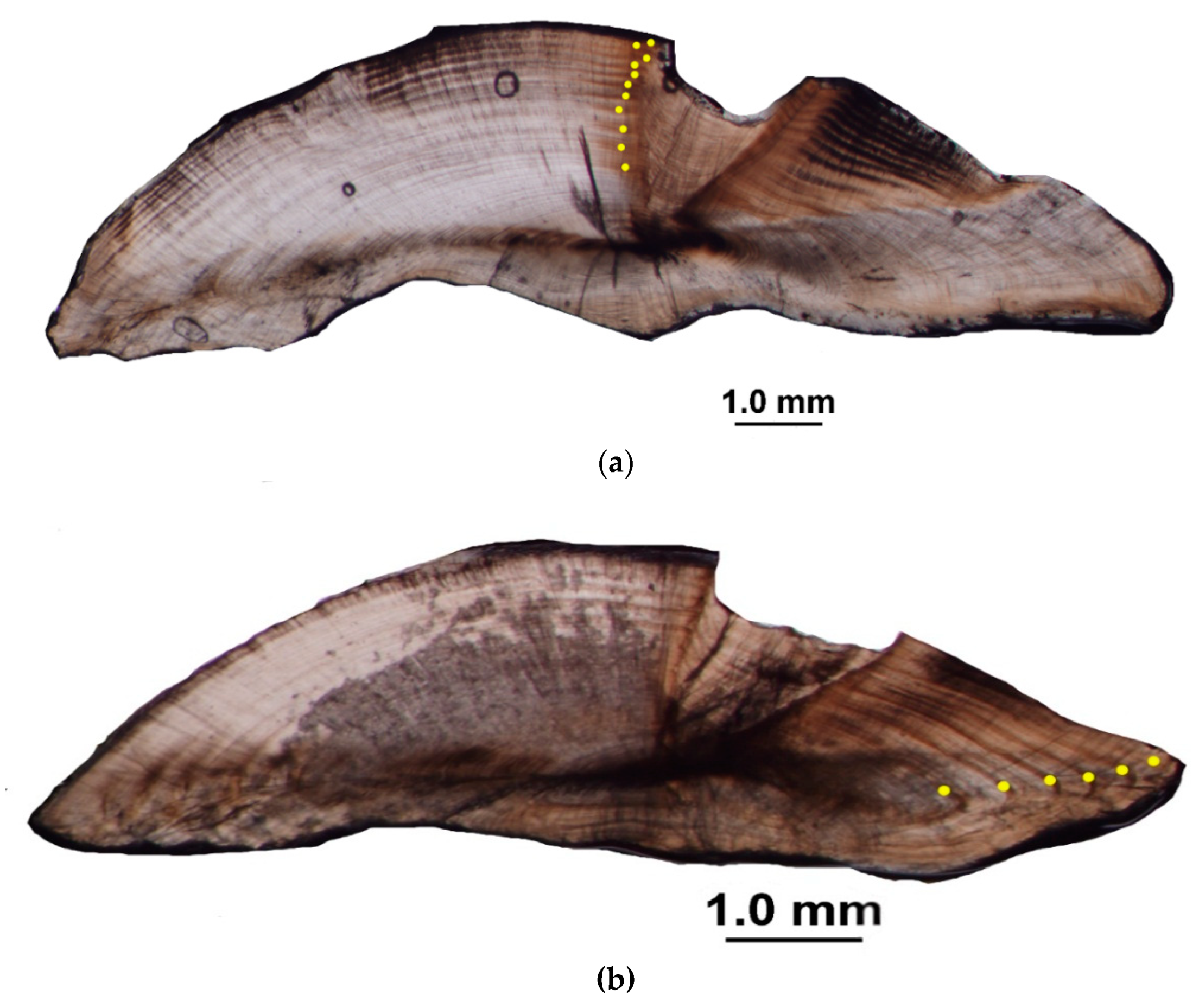
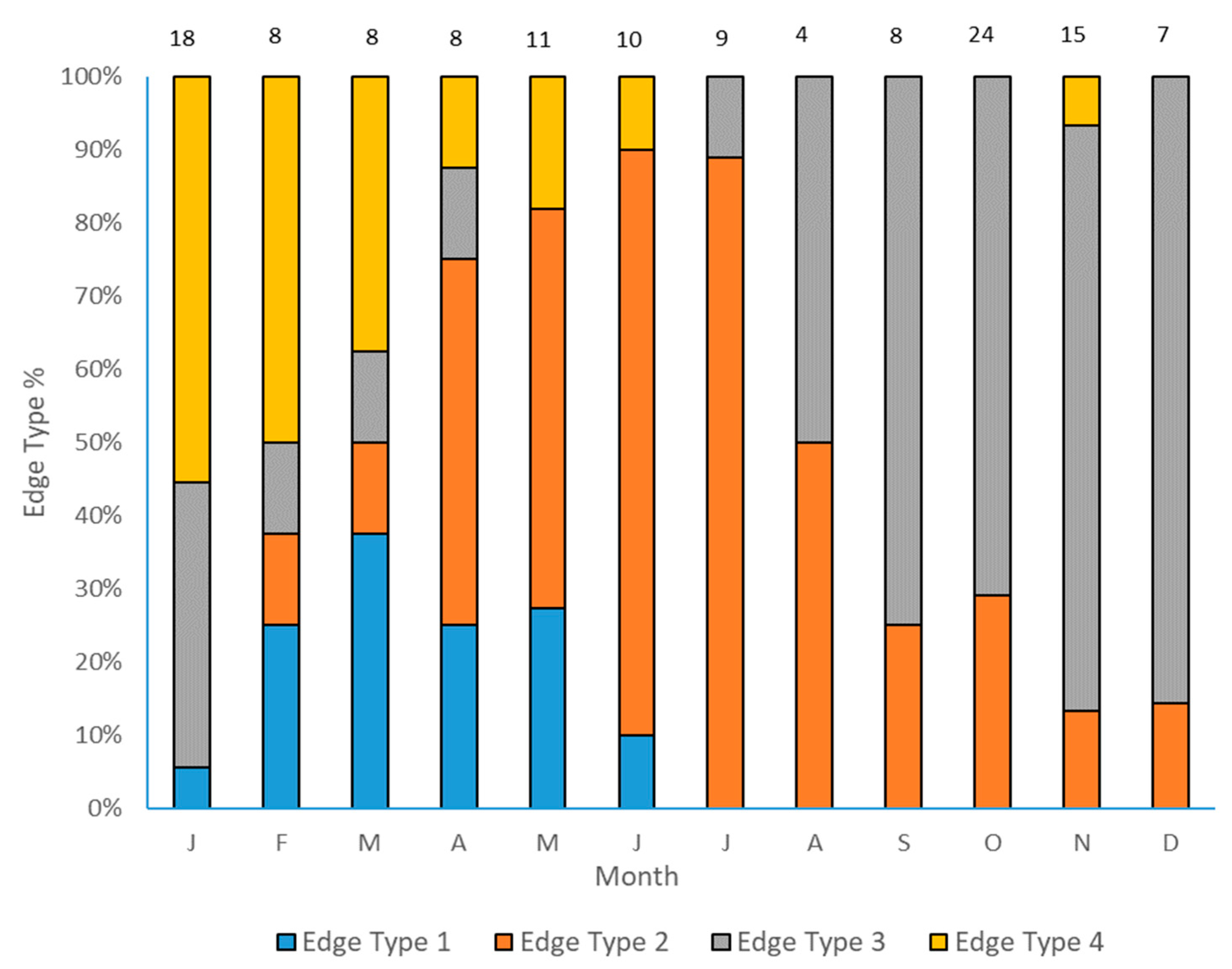
2.1. Age Determination and Timing of Opaque Zone Formation
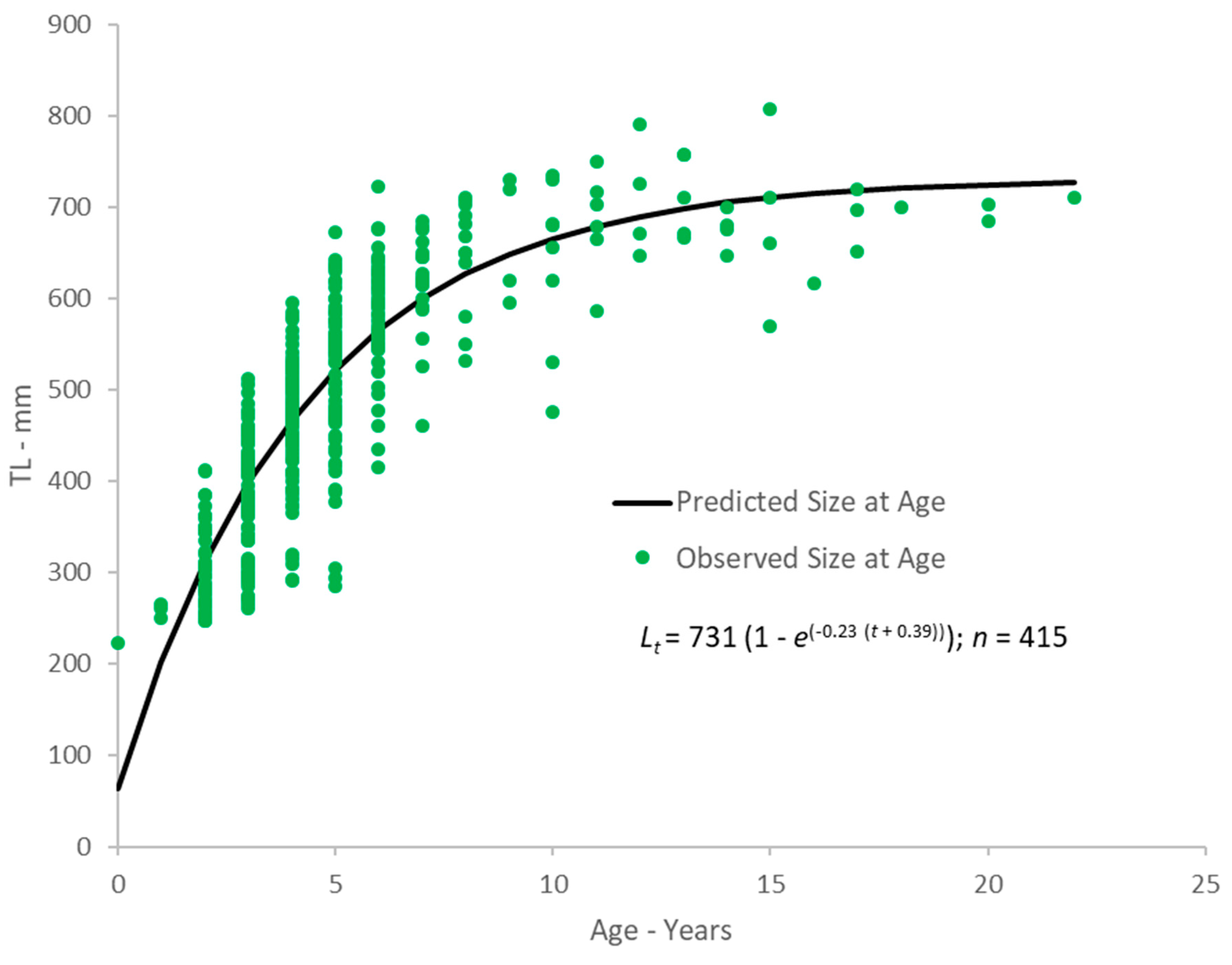
2.2. Growth
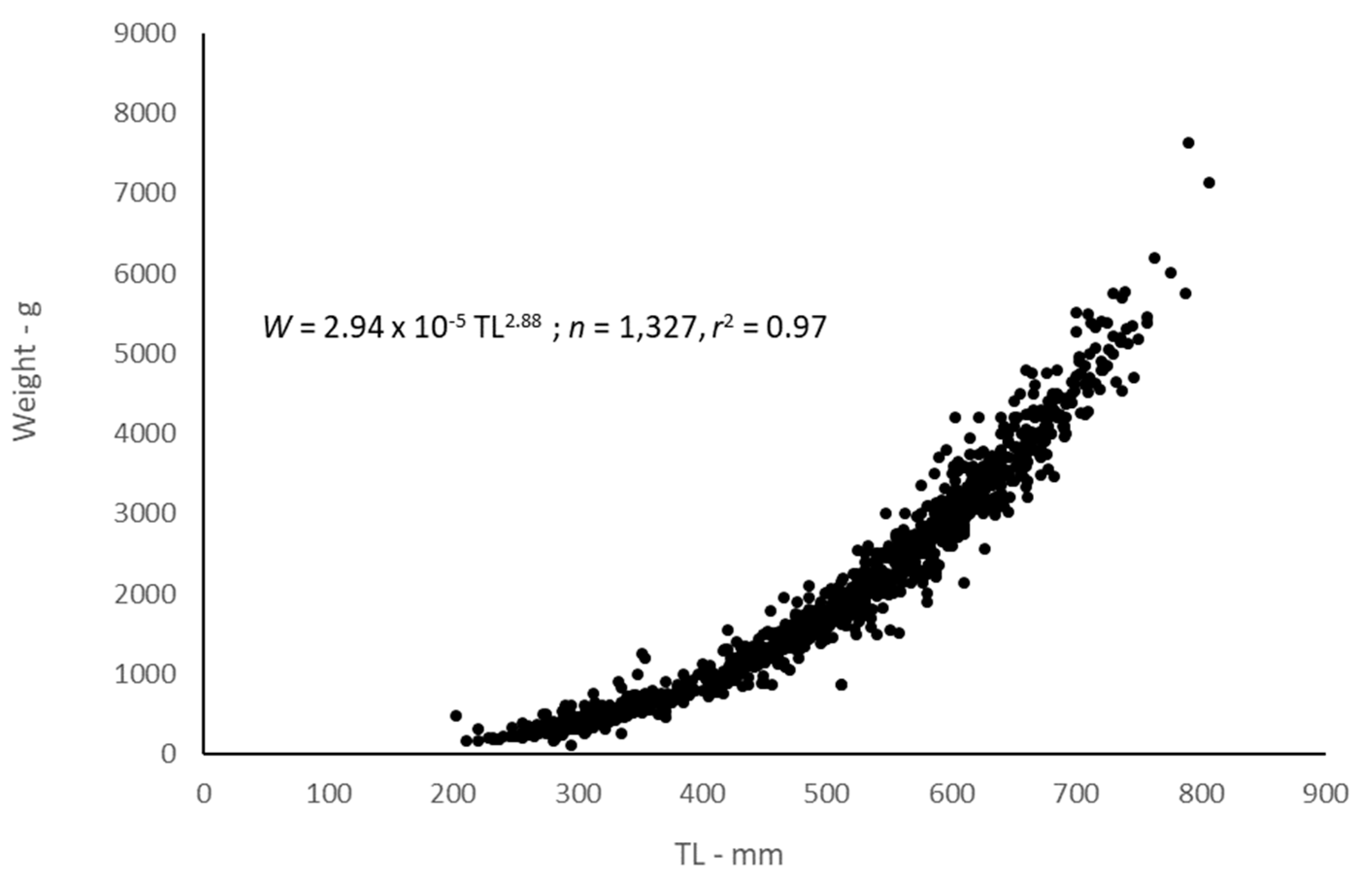
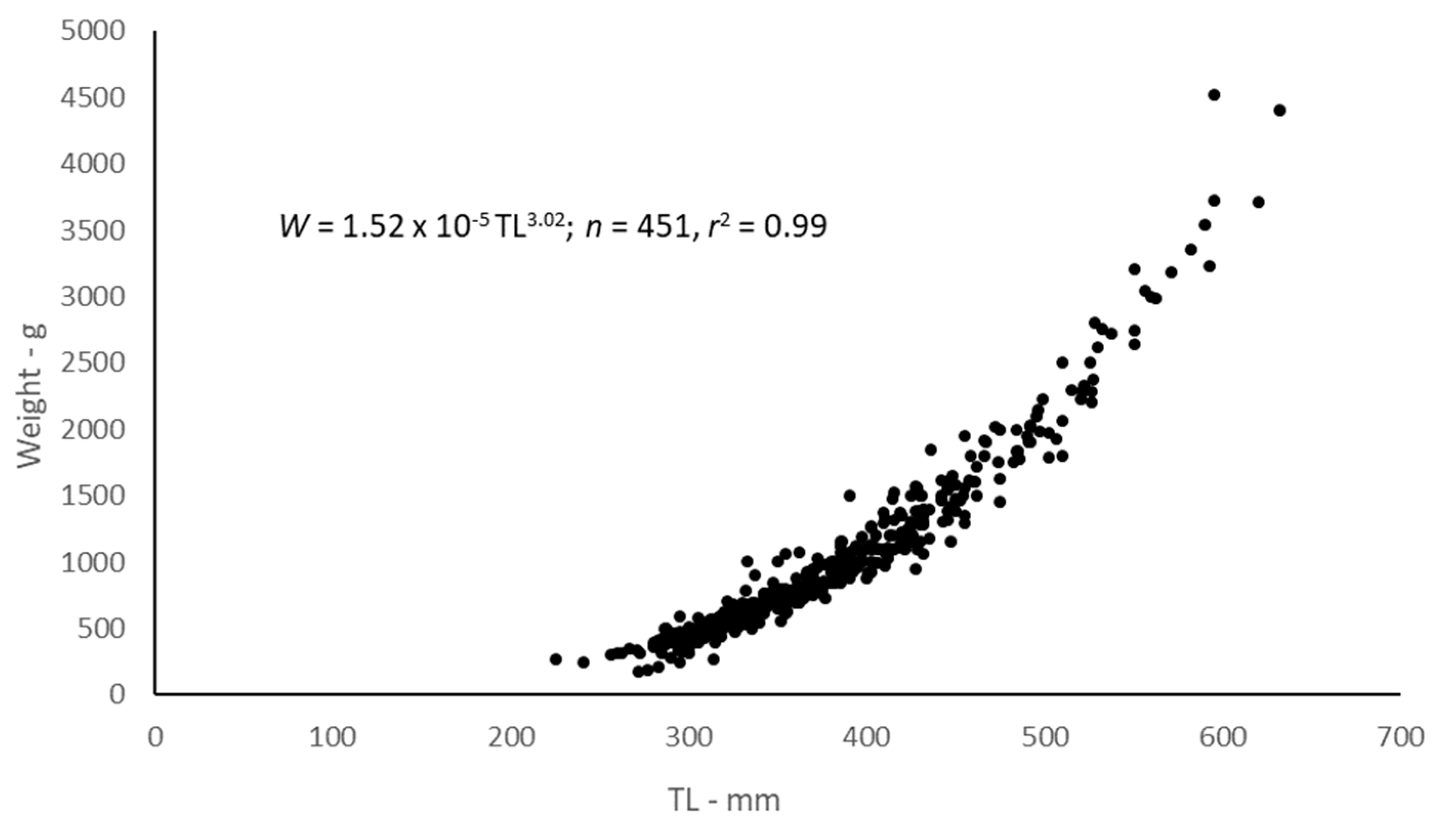
2.3. Body-Size Relationships
2.4. Natural Mortality
3. Discussion
4. Materials and Methods
4.1. Age Determination and Timing of Opaque Zone Formation
4.2. Growth
4.3. Body-Size Relationships
4.4. Natural Mortality
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- McEachran, J.D.; Fechhelm, J.D. Fishes of the Gulf of Mexico, Volume 2: Scorpaeniformes to Tetraodontiformes; University of Texas Press: Austin, TX, USA, 2005; p. 1004. [Google Scholar]
- Spach, H.L.; Godefroid, R.S.; Santos, C.; Schwartz, R.J.R.; de Queiroz, G.M.L. Temporal variation in fish assemblage composition on a tidal flat. Braz. J. Oceanogr. 2004, 52, 47–58. [Google Scholar] [CrossRef] [Green Version]
- Lindeman, K.C.; Snyder, D.B. Nearshore hard bottom fishes of southeast Florida and effects of habitat burial caused by dredging. Fish. Bull. 1999, 97, 508–525. [Google Scholar]
- Xavier, J.H.A.; Cordeiro, C.A.M.M.; Tenorio, G.D.; Diniz, A.F.; Paulo, E.P.N.; Rosa, R.S.; Rosa, I.L. Fish assemblage of the mamanguape environmental protection area, NE Brazil: Abundance, composition and microhabitat availability along the mangrove-reef gradient. Neotrop. Ichthyol. 2012, 10, 109–122. [Google Scholar] [CrossRef]
- Tupper, M.; Rudd, M.A. Species-specific impacts of a small marine reserve on reef fish production and fishing productivity in the Turks and Caicos Islands. Environ. Cons. 2002, 29, 484–492. [Google Scholar] [CrossRef] [Green Version]
- Feitoza, B.M.; Rosa, R.S.; Rocha, L.A. Ecology and zoogeography of deep-reef fishes in northeastern Brazil. Bull. Mar. Sci. 2005, 76, 725–742. [Google Scholar]
- Cervigón, F. Los Peces Marinos de Venezuela, Volume 2; Fundación Científica Los Roques: Caracas, Venezuela, 1993. [Google Scholar]
- Recreational Fisheries Statistics Queries. Available online: https://www.st.nmfs.noaa.gov/st1/recreational/queries/ (accessed on 14 June 2019).
- Brennan, K. Annual Landings from the Southeast Region Headboat Survey; NOAA National Marine Fisheries Service, Southeast Fisheries Science Center: Beaufort, NC, USA, 2019.
- Commercial Fisheries Statistics. Available online: https://www.st.nmfs.noaa.gov/commercial-fisheries/commercial-landings/annual-landings/index (accessed on 14 June 2019).
- SAFMC. Margate: Fish ID and Regs: Regulations by Species. Available online: http://safmc.net/regulations/regulations-by-species/margate/ (accessed on 9 May 2019).
- Florida Saltwater Fishing Regulations. Available online: http://www.eregulations.com/florida/fishing/saltwater/coastal-species/ (accessed on 14 June 2019).
- Billings, V.C.; Munro, J. The biology, ecology, exploitation and managemenr of Caribbean reef fishes. Scientific report of the ODA/UWI Fisheries Ecology Research Project, Port Royal Marine Laboratory, Jamaica, 1969–1973. Part 5e. The biology, ecology and bionomics of Caribbean reef fishes: Pomadasyidae (grunts). Res. Rep. Zool. Dep. Univ. West Indies 1974, 3:pt. 5e. [Google Scholar]
- García-Arteaga, J.P. Edad y crecimiento del jallao, Haemulon album, en la plataforma suroccidental de Cuba. Rep. Invest. Inst. Oceanol. Acad. Cienc. Cuba 1983, 14, 25. [Google Scholar]
- Campana, S.E. Accuracy, precision and quality control in age determination, including a review of the use and abuse of age validation methods. J. Fish Biol. 2001, 59, 197–242. [Google Scholar] [CrossRef]
- Harris, P.J.; Wyanski, D.M.; White, D.B.; Mikell, P.P.; Eyo, P.B. Age, growth, and reproduction of greater amberjack off the southeastern U.S. Atlantic coast. Trans. Am. Fish. Soc. 2007, 136, 1534–1545. [Google Scholar] [CrossRef]
- Von Bertalanffy, L. A quantitative theory of organic growth. Hum. Biol. 1938, 10, 181–243. [Google Scholar]
- Charnov, E.L.; Gislason, H.; Pope, J.G. Evolutionary assembly rules for fish life histories. Fish Fish. 2013, 14, 212–224. [Google Scholar] [CrossRef]
- McGarvey, R.; Fowler, A.J. Seasonal growth of King George whiting (Sillaginodes punctata) from length-at-age samples truncated by legal minimum size. Fish. Bull. 2002, 100, 545–558. [Google Scholar]
- Beauchamp, J.J.; Olson, J.S. Corrections for bias in regression estimates after logarithmic transformations. Ecology 1973, 54, 1403–1407. [Google Scholar] [CrossRef]
- Hewitt, D.A.; Hoenig, J.M. Comparison of two approaches for estimating natural mortality based on longevity. Fish. Bull. 2005, 103, 433–437. [Google Scholar]
- Potts, J.C.; Manooch, C.S., III. Differences in the age and growth of white grunt (Haemulon plumieri) from North Carolina and South Carolina compared with southeast Florida. Bull. Mar. Sci. 2001, 68, 1–12. [Google Scholar]
- Murie, D.J.; Parkyn, D.C. Age and growth of white grunt (Haemulon plumieri): A comparison of two populations along the west coast of Florida. Bull. Mar. Sci. 2005, 76, 73–93. [Google Scholar]
- Potts, J.C.; Manooch, C.S., III. Age and growth of red hind and rock hind collected from North Carolina through the Dry Tortugas, Florida. Bull. Mar. Sci. 1995, 56, 784–794. [Google Scholar]
- Burton, M.L. Age, growth, and mortality of gray snapper from the east coast of Florida. Fish. Bull. 2001, 99, 245–256. [Google Scholar]
- Burton, M.L.; Potts, J.C.; Poholek, A.J.; Ostrowski, A.; Page, J. Age, growth, natural mortality, and reproductive seasonality of knobbed porgy from the southeastern United States. Mar. Coast. Fish. 2019, 11, 231–245. [Google Scholar] [CrossRef]
- Beamish, R.J.; Fournier, D.A. A method for comparing the precision of a set of age determinations. Can. J. Fish. Aquat. Sci. 1981, 38, 982–983. [Google Scholar] [CrossRef]
- SAS Institute, Inc. SAS/STAT Guide for Personal Computers, Version 6 Edition; SAS Institute: Cary, NC, USA, 1987. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Age | n | Mean Observed TL (±SE) | TL Range | Predicted TL | M-y−1 |
|---|---|---|---|---|---|
| 0 | 1 | 222 | – | 64 | 2.93 |
| 1 | 3 | 258 (4) | 250–265 | 202 | 1.11 |
| 2 | 42 | 313 (7) | 246–411 | 311 | 0.68 |
| 3 | 81 | 384 (7) | 260–512 | 398 | 0.51 |
| 4 | 97 | 465 (6) | 290–595 | 467 | 0.42 |
| 5 | 66 | 520 (10) | 285–672 | 522 | 0.36 |
| 6 | 49 | 582 (9) | 415–722 | 565 | 0.32 |
| 7 | 21 | 615 (11) | 460–684 | 599 | 0.30 |
| 8 | 12 | 647 (18) | 532–710 | 626 | 0.28 |
| 9 | 4 | 666 (34) | 595–730 | 648 | 0.27 |
| 10 | 8 | 639 (33) | 476–735 | 665 | 0.26 |
| 11 | 6 | 683 (23) | 586–750 | 679 | 0.25 |
| 12 | 4 | 708 (32) | 647–790 | 690 | 0.25 |
| 13 | 5 | 712 (20) | 666–757 | 698 | 0.24 |
| 14 | 4 | 676 (11) | 647–700 | 705 | 0.24 |
| 15 | 4 | 687 (49) | 570–807 | 710 | 0.24 |
| 16 | 1 | 616 | – | 715 | 0.24 |
| 17 | 3 | 689 (20) | 651–719 | 718 | 0.24 |
| 18 | 1 | 699 | – | 721 | 0.23 |
| 19 | – | – | 723 | 0.23 | |
| 20 | 2 | 694 (9) | 685–703 | 725 | 0.23 |
| 21 | – | – | 726 | 0.23 | |
| 22 | 1 | 710 | – | 727 | 0.23 |
| Age | n | Mean Observed TL (±SE) | TL Range | Predicted TL | M-y−1 |
|---|---|---|---|---|---|
| 0 | – | – | – | 0 | 7.20 |
| 1 | – | – | – | 84 | 1.57 |
| 2 | – | – | – | 155 | 0.83 |
| 3 | 1 | 342 | – | 215 | 0.56 |
| 4 | 6 | 295 (19) | 241–367 | 264 | 0.43 |
| 5 | 11 | 336 (14) | 291–451 | 306 | 0.36 |
| 6 | 10 | 353 (11) | 295–397 | 341 | 0.31 |
| 7 | 24 | 391 (16) | 311–628 | 370 | 0.28 |
| 8 | 20 | 409 (17) | 309–562 | 395 | 0.26 |
| 9 | 18 | 408 (19) | 292–595 | 416 | 0.24 |
| 10 | 15 | 411 (19) | 320–570 | 433 | 0.23 |
| 11 | 7 | 502 (29) | 377–592 | 448 | 0.22 |
| 12 | 3 | 429 (65) | 343–556 | 460 | 0.21 |
| 13 | 6 | 494 (50) | 355–615 | 470 | 0.20 |
| 14 | 1 | 354 | – | 478 | 0.20 |
| 15 | 2 | 576 (66) | 510–641 | 485 | 0.19 |
| 16 | 2 | 462 (41) | 421–502 | 492 | 0.19 |
| 17 | 2 | 485 (65) | 420–550 | 497 | 0.19 |
| 18 | 2 | 479 (34) | 445–512 | 501 | 0.19 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Burton, M.L.; Potts, J.C.; Ostrowski, A.D. Preliminary Estimates of Age, Growth and Natural Mortality of Margate, Haemulon album, and Black Margate, Anisotremus surinamensis, from the Southeastern United States. Fishes 2019, 4, 44. https://0-doi-org.brum.beds.ac.uk/10.3390/fishes4030044
Burton ML, Potts JC, Ostrowski AD. Preliminary Estimates of Age, Growth and Natural Mortality of Margate, Haemulon album, and Black Margate, Anisotremus surinamensis, from the Southeastern United States. Fishes. 2019; 4(3):44. https://0-doi-org.brum.beds.ac.uk/10.3390/fishes4030044
Chicago/Turabian StyleBurton, Michael L., Jennifer C. Potts, and Andrew D. Ostrowski. 2019. "Preliminary Estimates of Age, Growth and Natural Mortality of Margate, Haemulon album, and Black Margate, Anisotremus surinamensis, from the Southeastern United States" Fishes 4, no. 3: 44. https://0-doi-org.brum.beds.ac.uk/10.3390/fishes4030044