Modulation of Pituitary Response by Dietary Lipids and Throughout a Temperature Fluctuation Challenge in Gilthead Sea Bream

 ,
,
Abstract
:1. Introduction
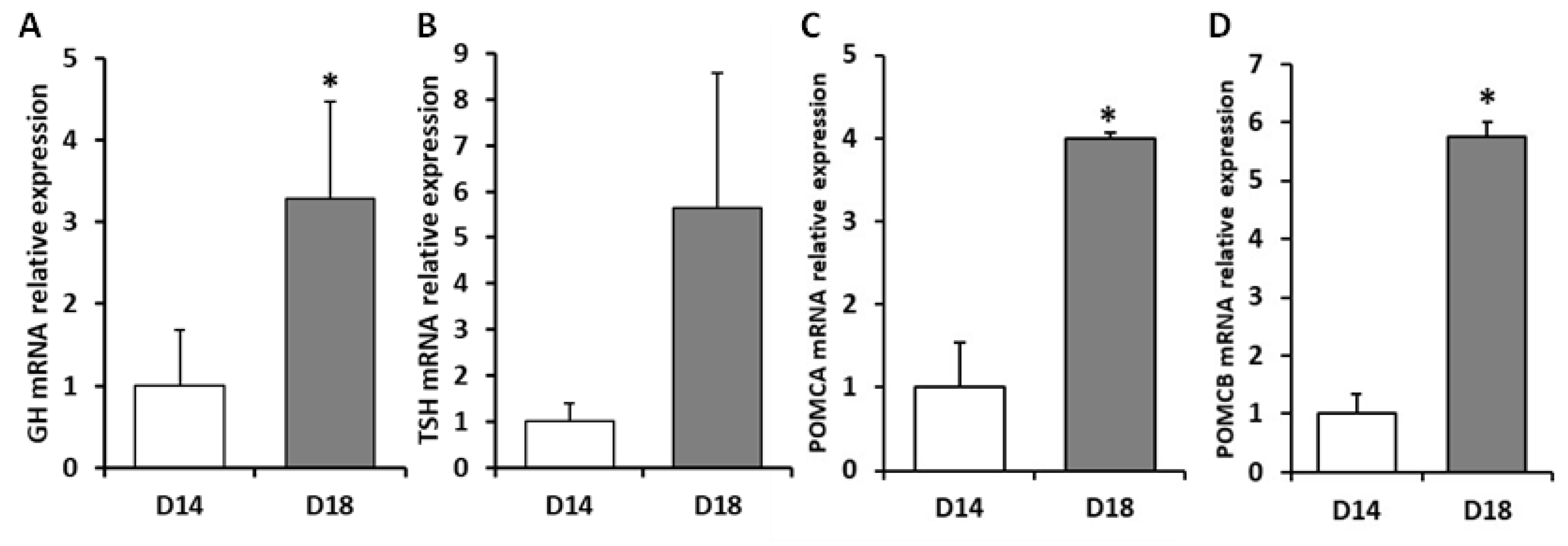
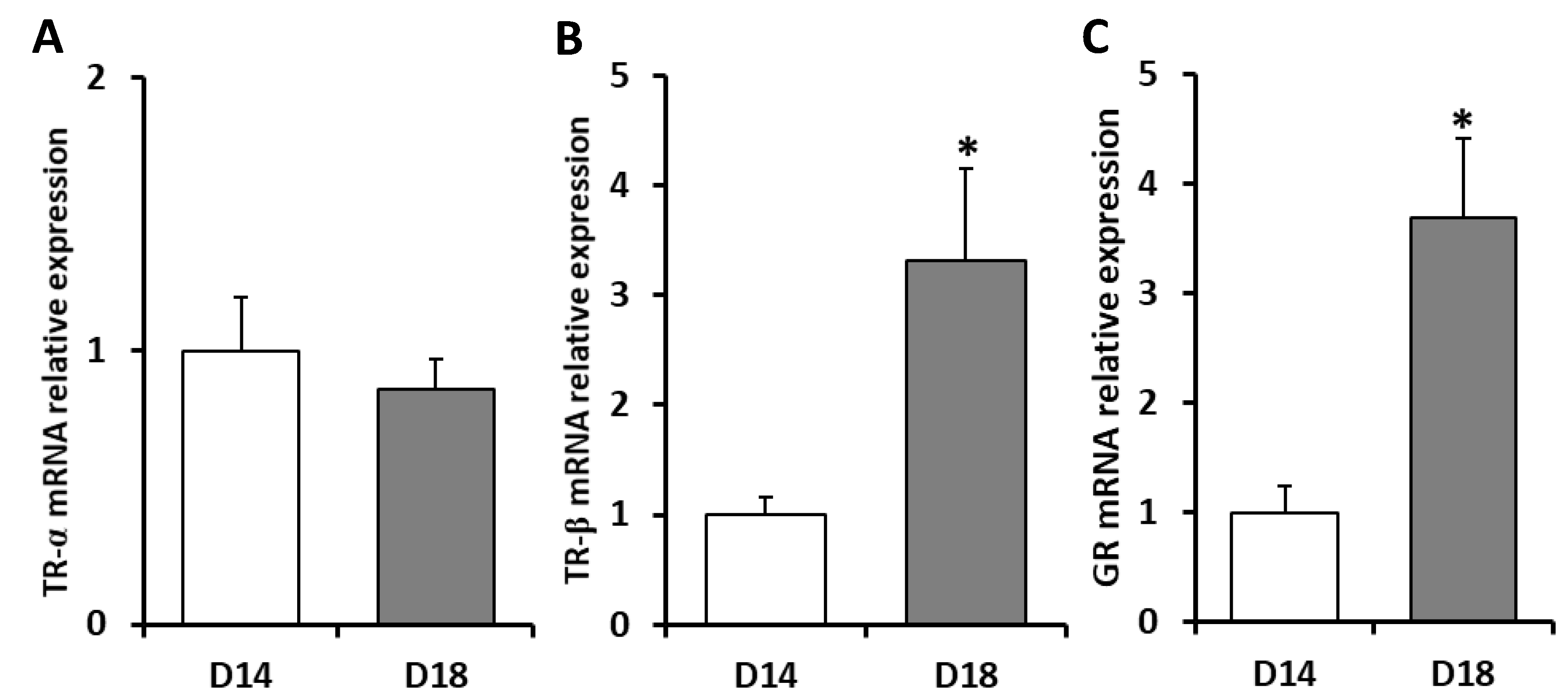
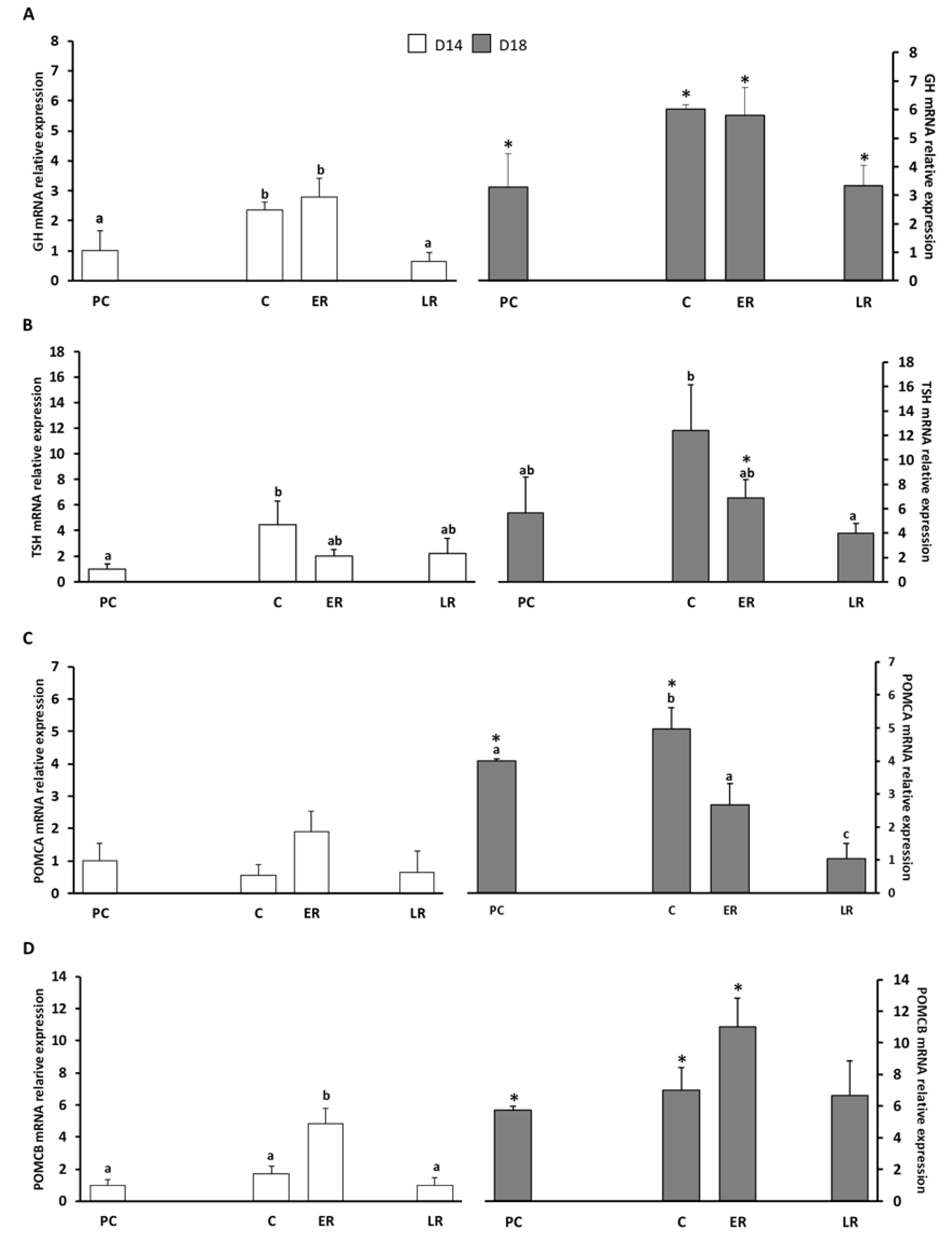
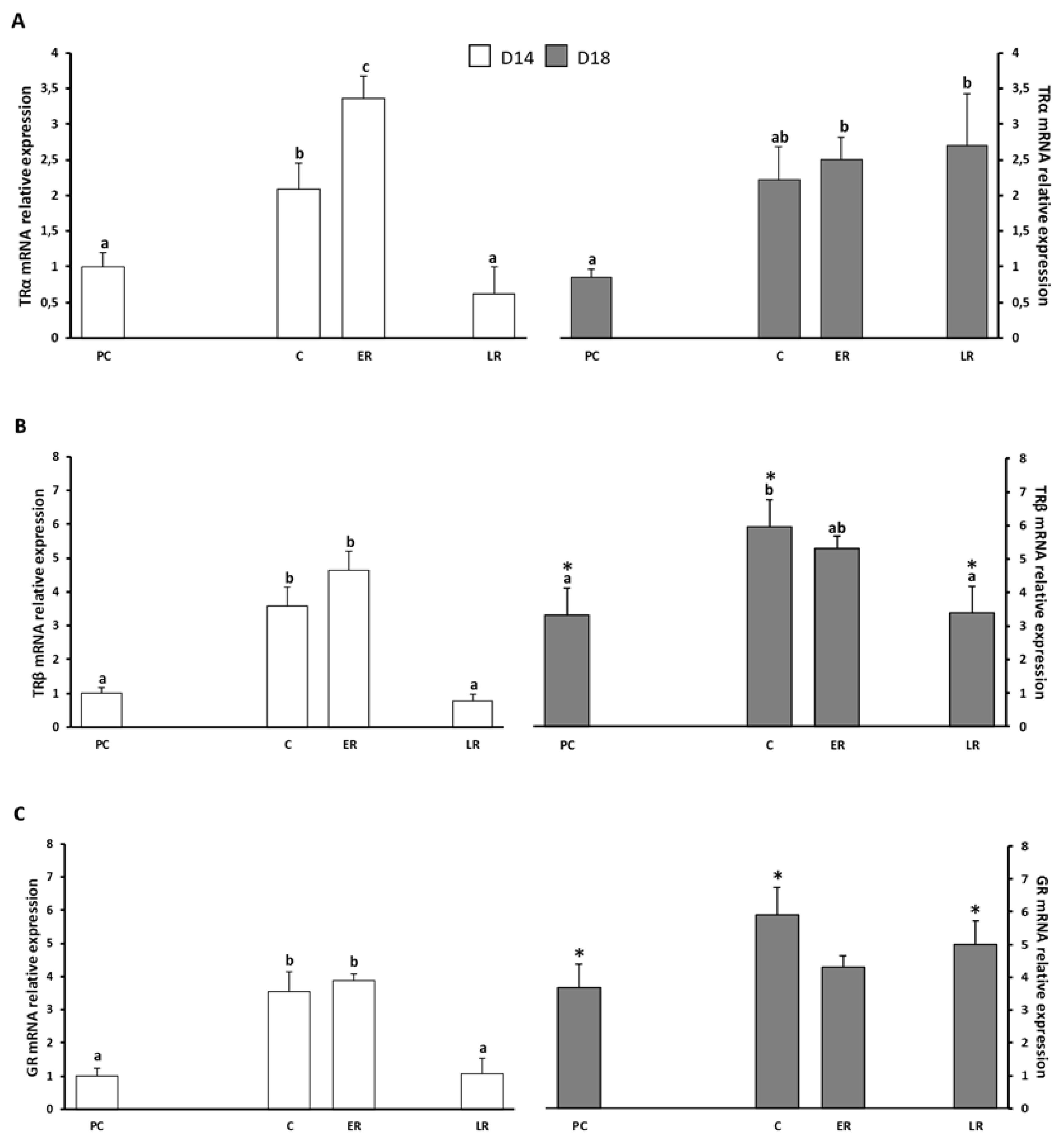
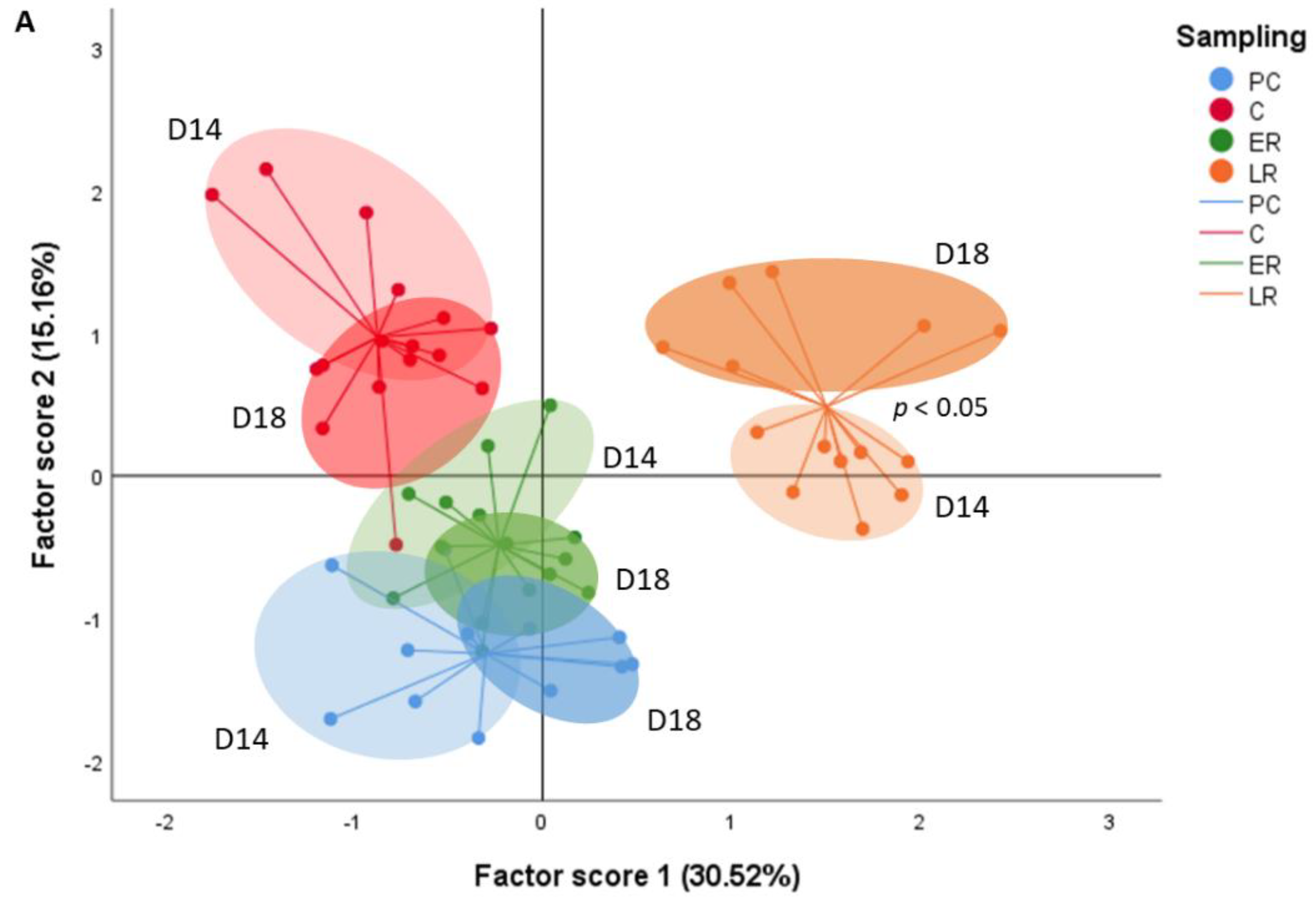
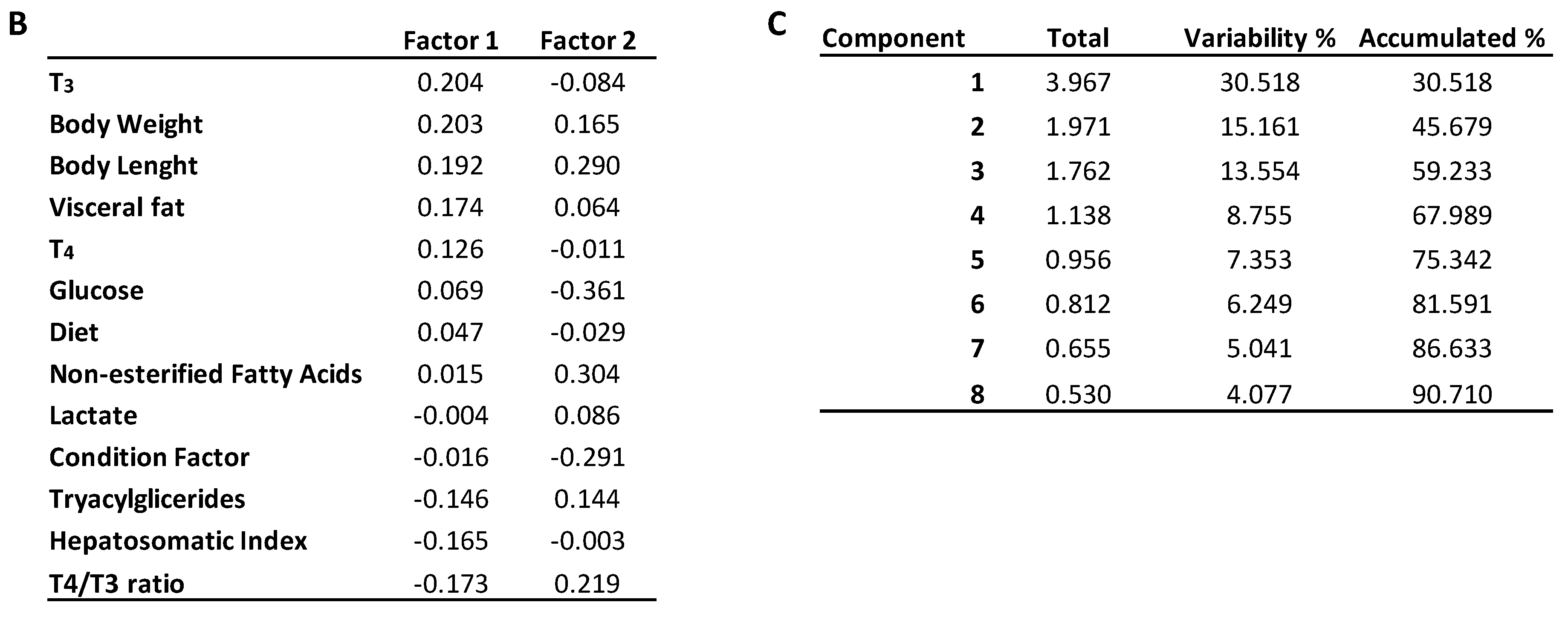
2. Results
2.1. Effects of Dietary Lipid on Pituitary Expression
2.2. Effects of Thermal Fluctuation on Pituitary Expression
2.3. Effects of Dietary Lipids and Thermal Fluctuation on Plasma Thyroid Hormones
3. Discussion
3.1. Dietary Modulation of Pituitary Response at Warm Temperature
3.2. Cold Effects
3.3. Temperature Recovery
4. Materials and Methods
4.1. Animal Condition and Sampling
4.2. Plasma Thyroid Hormones
4.3. Transcript Expression Quantification
4.4. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hurst, T.P. Causes and consequences of winter mortality in fishes. J. Fish Biol. 2007, 71, 315–345. [Google Scholar] [CrossRef]
- Donaldson, M.R.; Cooke, S.J.; Patterson, D.A.; Macdonald, J.S. Cold shock and fish. J. Fish Biol. 2008, 73, 1491–1530. [Google Scholar] [CrossRef]
- Davis, P.S. Two occurrences of the gilthead, Sparus aurata Linnaeus 1758, on the coast of Northumberland, England. J. Fish Biol. 1988, 33, 951. [Google Scholar] [CrossRef]
- Ibarz, A.; Padrós, F.; Gallardo, M.Á.; Fernández-Borràs, J.; Blasco, J.; Tort, L. Low-temperature challenges to gilthead sea bream culture: Review of cold-induced alterations and “Winter Syndrome”. Rev. Fish Biol. Fish. 2010, 20, 539–556. [Google Scholar] [CrossRef]
- Ibarz, A.; Fernández-Borràs, J.; Blasco, J.; Gallardo, M.A.; Sánchez, J. Oxygen consumption and feeding rates of gilthead sea bream (Sparus aurata) reveal lack of acclimation to cold. Fish Physiol. Biochem. 2003, 29, 313–321. [Google Scholar] [CrossRef]
- Tort, L.; Padrós, F.; Rotllant, J.; Crespo, S. Winter syndrome in the gilthead sea breamSparus aurata. Immunological and histopathological features. Fish Shellfish Immunol. 1998, 8, 37–47. [Google Scholar] [CrossRef]
- Sarusic, G. Clinical signs of the winter disease phenomenon in sea bream (Sparus aurata, L.). Bull. Eur. Assoc. Fish Pathol. 1999, 19, 113. [Google Scholar]
- Sánchez-Nuño, S.; Sanahuja, I.; Ferández-Alacid, L.; Ordóñez-Grande, B.; Fontanillas, R.; Fernández-Borràs, J.; Blasco, J.; Carbonell, T.; Ibarz, A. Redox challenge in a cultured temperate marine species during low temperature and temperature recovery. Front. Physiol. 2018, 9, 923. [Google Scholar] [CrossRef]
- Elliot, J. Tolerance and resistence to thermal stress in juvenile Atlantic salmon, Salmo salar. Freshw Biol 1991, 25, 61–70. [Google Scholar] [CrossRef]
- Ibarz, A.; Beltrán, M.; Fernández-Borràs, J.; Gallardo, M.A.; Sánchez, J.; Blasco, J. Alterations in lipid metabolism and use of energy depots of gilthead sea bream (Sparus aurata) at low temperatures. Aquaculture 2007, 262, 470–480. [Google Scholar] [CrossRef]
- Ibarz, A.; Blasco, J.; Sala-Rabanal, M.; Gallardo, Á.; Redondo, A.; Fernández-Borràs, J. Metabolic rate and tissue reserves in gilthead sea bream (Sparus aurata) under thermal fluctuations and fasting and their capacity for recovery. Can. J. Fish. Aquat. Sci. 2007, 64, 1034–1042. [Google Scholar] [CrossRef]
- Ibarz, A.; Blasco, J.; Beltrán, M.; Gallardo, M.A.; Sánchez, J.; Sala, R.; Fernández-Borràs, J. Cold-induced alterations on proximate composition and fatty acid profiles of several tissues in gilthead sea bream (Sparus aurata). Aquaculture 2005, 249, 477–486. [Google Scholar] [CrossRef]
- Blasco, J.; Moya, A.; Millán-Cubillo, A.; Vélez, E.J.; Capilla, E.; Pérez-Sánchez, J.; Gutiérrez, J.; Fernández- Borrás, J. Growth-promoting effects of sustained swimming in fingerlings of gilthead sea bream (Sparus aurata L.). J. Comp. Physiol. B 2015, 185, 859–868. [Google Scholar] [CrossRef] [PubMed]
- Sangiao-Alvarellos, S.; Laiz-Carrión, R.; Guzmán, J.M.; Martín del Río, M.P.; Miguez, J.M.; Mancera, J.M.; Soengas, J.L. Acclimation of S. aurata to various salinities alters energy metabolism of osmoregulatory and nonosmoregulatory organs. Am. J. Physiol. Integr. Comp. Physiol. 2003, 285, R897–R907. [Google Scholar] [CrossRef] [PubMed]
- Polakof, S.; Arjona, F.J.; Sangiao-Alvarellos, S.; Martín del Río, M.P.; Mancera, J.M.; Soengas, J.L. Food deprivation alters osmoregulatory and metabolic responses to salinity acclimation in gilthead sea bream Sparus auratus. J. Comp. Physiol. B 2006, 176, 441–452. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Nuño, S.; Eroldogan, O.; Sanahuja, I.; Özsahinoglu, I.; Blasco, J.; Fernández-Borràs, J.; Fontanillas, R.; Guerreiro, P.; Ibarz, I. Cold-induced growth arrest in gilthead sea bream Sparus aurata: Metabolic reorganisation and recovery. Aquac. Environ. Interact. 2018, 10, 511–528. [Google Scholar] [CrossRef]
- Mingarro, M.; Vega-Rubén De Celis, S.; Astola, A.; Pendón, C.; Valdivia, M.M.; Pérez-Sánchez, J. Endocrine mediators of seasonal growth in gilthead sea bream (Sparus aurata): The growth hormone and somatolactin paradigm. Gen. Comp. Endocrinol. 2002, 128, 102–111. [Google Scholar] [CrossRef]
- Power, D.M.; Melo, J.; Santos, C.R.A. The effect of food deprivation and refeeding on the liver, thyroid hormones and transthyretin in sea bream. J. Fish Biol. 2000, 56, 374–387. [Google Scholar] [CrossRef]
- Rotllant, J.; Balm, P.H.M.; Wendelaar-Bonga, S.E.; Pérez-Sánchez, J.; Tort, L. A drop in ambient temperature results in a transient reduction of interrenal ACTH responsiveness in the gilthead sea bream (Sparus aurata, L.). Fish Physiol. Biochem. 2000, 23, 265–273. [Google Scholar] [CrossRef]
- Sánchez-Nuño, S.; Sanahuja, I.; Fernández-Alacid, L.; Ordóñez-Grande, B.; Carbonell, T.; Ibarz, A. Oxidative attack during temperature fluctuation challenge compromises liver protein homeostasis of a temperate fish model. Comp. Biochem. Physiol. Part-B Biochem. Mol. Biol. 2019, 236, 110311. [Google Scholar] [CrossRef]
- Bertucci, J.I.; Blanco, A.M.; Sundarrajan, L.; Rajeswari, J.J.; Velasco, C.; Unniappan, S. Nutrient Regulation of Endocrine Factors Influencing Feeding and Growth in Fish. Front. Endocrinol. (Lausanne) 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Ganga, R.; Tort, L.; Acerete, L.; Montero, D.; Izquierdo, M.S. Modulation of ACTH-induced cortisol release by polyunsaturated fatty acids in interrenal cells from gilthead seabream, Sparus aurata. J. Endocrinol. 2006, 190, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Requeni, P.; Kraemer, M.N.; Canosa, L.F. Regulation of somatic growth and gene expression of the GH-IGF system and PRP-PACAP by dietary lipid level in early juveniles of a teleost fish, the pejerrey (Odontesthes bonariensis). J. Comp. Physiol. B Biochem. Syst. Environ. Physiol. 2012, 182, 517–530. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, J.C.R.; Laiz-Carrion, R.; Louro, B.; Silva, N.; Canario, A.V.M.; Mancera, J.M.; Power, D.M. Divergence of duplicate POMC genes in gilthead sea bream Sparus auratus. Gen. Comp. Endocrinol. 2011, 173, 396–404. [Google Scholar] [CrossRef]
- Suárez, M.D.; Trenzado, C.E.; García-Gallego, M.; Furné, M.; García-Mesa, S.; Domezain, A.; Alba, I.; Sanz, A. Interaction of dietary energy levels and culture density on growth performance and metabolic and oxidative status of rainbow trout (Oncorhynchus mykiss). Aquac. Eng. 2015, 67, 59–66. [Google Scholar] [CrossRef]
- Bonaldo, A.; Isani, G.; Fontanillas, R.; Parma, L.; Grilli, E.; Gatta, P.P. Growth and feed utilization of gilthead sea bream (Sparus aurata, L.) fed to satiation and restrictively at increasing dietary energy levels. Aquac. Int. 2010, 18, 909–919. [Google Scholar] [CrossRef]
- Mongile, F.; Bonaldo, A.; Fontanillas, R.; Mariani, L.; Badiani, A.; Bonvini, E.; Parma, L. Effects of dietary lipid level on growth and feed utilisation of gilthead seabream (Sparus aurata L.) reared at Mediterranean summer temperature. Ital. J. Anim. Sci. 2014, 13, 30–34. [Google Scholar] [CrossRef]
- Velázquez, M.; Zamora, S.; Martínez, F.J. Effect of dietary energy content on gilthead sea bream (Sparus aurata) feeding behaviour and nutritional use of the diet. Aquac. Nutr. 2006, 12, 127–133. [Google Scholar] [CrossRef]
- Ibarz, A.; Costa, R.; Harrison, A.P.; Power, D.M. Dietary keto-acid feed-back on pituitary activity in gilthead sea bream: Effects of oral doses of AKG. A proteomic approach. Gen. Comp. Endocrinol. 2010, 169, 284–292. [Google Scholar] [CrossRef]
- MacKenzie, D.S.; Vanputte, C.M.; Leiner, K.A. Nutrient regulation of endocrine function in fish. Aquaculture 1998, 161, 3–25. [Google Scholar] [CrossRef]
- Srijila, C.K.; Rani, A.M.B.; Babu, P.G.; Tiwari, V.K. Ration restriction, compensatory growth and pituitary growth hormone gene expression in Labeo rohita. Aquac. Int. 2014, 22, 1703–1710. [Google Scholar] [CrossRef]
- Company, R.; Calduch-Giner, J.; Kaushik, S.; Pérez-Sánchez, J. Growth performance and adiposity in gilthead sea bream (Sparus aurata): Risks and benefits of high energy diets. Aquaculture 1999, 171, 279–292. [Google Scholar] [CrossRef]
- Deane, E.E.; Woo, N.Y.S. Expression studies on glucose-6-phosphate dehydrogenase in sea bream: Effects of growth hormone, somatostatin, salinity and temperature. J. Exp. Zool. Part A Comp. Exp. Biol. 2005, 303, 676–688. [Google Scholar] [CrossRef] [PubMed]
- Leung, L.Y.; Woo, N.Y.S. Effects of growth hormone, insulin-like growth factor I, triiodothyronine, thyroxine, and cortisol on gene expression of carbohydrate metabolic enzymes in sea bream hepatocytes. Comp. Biochem. Physiol.-A Mol. Integr. Physiol. 2010, 157, 272–282. [Google Scholar] [CrossRef] [PubMed]
- Ryg, M.; Jacobsen, E. Seasonal changes in growth rate, feed intake, growth hormone, and thyroid hormones in young male reindeer (Rangifer tarandus tarandus). Can. J. Zool. 1982, 60, 15–23. [Google Scholar] [CrossRef]
- McGuire, M.A.; Beede, D.K.; Collier, R.J.; Buonomo, F.C.; DeLorenzo, M.A.; Wilcox, C.J.; Huntington, G.B.; Reynolds, C.K. Effects of acute thermal stress and amount of feed intake on concentrations of somatotropin, insulin-like growth factor (IGF)-I and IGF-II, and thyroid hormones in plasma of lactating Holstein cows2. J. Anim. Sci. 1991, 69, 2050–2056. [Google Scholar] [CrossRef]
- Cohn, W.B.; Jones, R.A.; Valverde, R.A.; Leiner, K.A.; MacKenzie, D.S. Molecular cloning and regulation of mRNA expression of the thyrotropin β and glycoprotein hormone α subunits in red drum, Sciaenops ocellatus. Fish Physiol. Biochem. 2010, 36, 1277–1290. [Google Scholar] [CrossRef]
- Montoya, A.; López-Olmeda, J.F.; Garayzar, A.B.S.; Sánchez-Vázquez, F.J. Synchronization of daily rhythms of locomotor activity and plasma glucose, cortisol and thyroid hormones to feeding in Gilthead seabream (Sparus aurata) under a light-dark cycle. Physiol. Behav. 2010, 101, 101–107. [Google Scholar] [CrossRef]
- Lin, X.; Volkoff, H.; Narnaware, Y.; Bernier, N.J.; Peyon, P.; Peter, R.E. Brain regulation of feeding behavior and food intake in fish. Comp. Biochem. Physiol.-A Mol. Integr. Physiol. 2000, 126, 415–434. [Google Scholar] [CrossRef]
- Conde-Sieira, M.; Chivite, M.; Míguez, J.M.; Soengas, J.L. Stress effects on the mechanisms regulating appetite in teleost fish. Front. Endocrinol. (Lausanne) 2018, 9, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Millington, G.W. The role of proopiomelanocortin (POMC) neurones in feeding behaviour. Nutr. Metab. (Lond.) 2007, 4, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Czeisler, C.A.; Klerman, E.B. Circadian and sleep-dependent regulation of hormone release in humans. Recent Prog. Horm. Res. 1999, 54, 97–130. [Google Scholar] [PubMed]
- Chiamolera, M.I.; Wondisford, F.E. Thyrotropin-Releasing Hormone and the Thyroid Hormone Feedback Mechanism. Endocrinology 2009, 150, 1091–1096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nelson, E.R.; Habibi, H.R. Thyroid receptor subtypes: Structure and function in fish. Gen. Comp. Endocrinol. 2009, 161, 90–96. [Google Scholar] [CrossRef] [PubMed]
- Bradley, D.J.; Young, W.S.; Weinberger, C. Differential expression of alpha and beta thyroid hormone receptor genes in rat brain and pituitary. Proc. Natl. Acad. Sci. USA 1989, 86, 7250–7254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Acerete, L.; Balasch, J.C.; Castellana, B.; Redruello, B.; Roher, N.; Canario, A.V.; Planas, J.V.; MacKenzie, S.; Tort, L. Cloning of the glucocorticoid receptor (GR) in gilthead seabream (Sparus aurata). Differential expression of GR and immune genes in gilthead seabream after an immune challenge. Comp. Biochem. Physiol.-B Biochem. Mol. Biol. 2007, 148, 32–43. [Google Scholar] [CrossRef] [PubMed]
- Deane, E.E.; Woo, N.Y.S. Molecular cloning of growth hormone from silver sea bream: Effects of abiotic and biotic stress on transcriptional and translational expression. Biochem. Biophys. Res. Commun. 2006, 342, 1077–1082. [Google Scholar] [CrossRef]
- Ayson, F.G.; de Jesus-Ayson, E.G.T.; Takemura, A. mRNA expression patterns for GH, PRL, SL, IGF-I and IGF-II during altered feeding status in rabbitfish, Siganus guttatus. Gen. Comp. Endocrinol. 2007, 150, 196–204. [Google Scholar] [CrossRef] [PubMed]
- Arslan, G.; Sahin, T.; Hisar, O.; Hisar, S.A. Effects of low temperature and starvation on plasma cortisol, triiodothyronine, thyroxine, thyroid-stimulating hormone and prolactin levels of juvenile common carp (Cyprinus carpio). Mar. Sci. Technol. Bull. 2016, 4, 5–9. [Google Scholar]
- Gaylord, T.G.; MacKenzie, D.S.; Gatlin, D.M., III. Growth performance, body composition and plasma thyroid hormone status of channel catfish (Ictalurus punctatus) in response to short-term feed deprivation and refeeding. Fish Physiol. Biochem. 2001, 24, 73–79. [Google Scholar] [CrossRef]
- De Pedro, N.; Delgado, M.J.; Gancedo, B.; Alonso-Bedate, M. Changes in glucose, glycogen, thyroid activity and hypothalamic catecholamines in tench by starvation and refeeding. J. Comp. Physiol. B Biochem. Syst. Environ. Physiol. 2003, 173, 475–481. [Google Scholar] [CrossRef] [PubMed]
- Blanton, M.L.; Specker, J.L. The hypothalamic-pituitary-thyroid (HPT) axis in fish and its role in fish development and reproduction. Crit. Rev. Toxicol. 2007, 37, 97–115. [Google Scholar] [CrossRef] [PubMed]
- Barton, B.A.; Schreck, C.B. Influence of acclimation temperature on interrenal and carbohydrate stress responses in juvenile chinook salmon (Oncorhynchus tshawytscha). Aquaculture 1987, 62, 299–310. [Google Scholar] [CrossRef]
- Sumpter, J.P.; Dye, H.M.; Benfey, T.J. The effects of stress on plasma ACTH, α-MSH, and cortisol levels in salmonid fishes. Gen. Comp. Endocrinol. 1986, 62, 377–385. [Google Scholar] [CrossRef]
- Staurnes, M.; Sigholt, T.; Åsgård, T.; Baeverfjord, G. Effects of a temperature shift on seawater challenge test performance in Atlantic salmon (Salmo salar) smolt. Aquaculture 2001, 201, 153–159. [Google Scholar] [CrossRef]
- Mateus, A.P.; Costa, R.; Gisbert, E.; Pinto, P.I.S.; Andree, K.B.; Estévez, A.; Power, D.M. Thermal imprinting modifies bone homeostasis in cold-challenged sea bream (Sparus aurata). J. Exp. Biol. 2017, 220, 3442–3454. [Google Scholar] [CrossRef] [Green Version]
- Mateus, A.P.; Costa, R.A.; Cardoso, J.C.R.; Andree, K.B.; Estévez, A.; Gisbert, E.; Power, D.M. Thermal imprinting modifies adult stress and innate immune responsiveness in the teleost sea bream. J. Endocrinol. 2017, 233, 381–394. [Google Scholar] [CrossRef]
- Martyniuk, C.J.; Kroll, K.J.; Porak, W.F.; Steward, C.; Grier, H.J.; Denslow, N.D. Seasonal relationship between gonadotropin, growth hormone, and estrogen receptor mRNA expression in the pituitary gland of largemouth bass. Gen. Comp. Endocrinol. 2009, 163, 306–317. [Google Scholar] [CrossRef] [Green Version]
- Pfaffl, M.W.; Tichopad, A.; Prgomet, C.; Neuvians, T.P. Determination of stable housekeeping genes, differentially regulated target genes and sample integrity: BestKeeper--Excel-based tool using pair-wise correlations. Biotechnol. Lett. 2004, 26, 509–515. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Thyroid Hormones | Pre-Cold (Day 30) | Cold (Day 80) | Early Recovery (Day 87) | Late Recovery (Day 115) |
|---|---|---|---|---|
| T3 (ng/mL) | ||||
| D14 | 4.4 ± 0.9 a | 2.2 ± 0.4 a | 3.7 ± 0.6 a | 6.9 ± 1.1 b |
| D18 | 5.1 ± 0.8 ab | 2.5 ± 0.4 a | 4.1 ± 0.7 ab | 7.2 ± 2.0 b |
| T4 (ng/mL) | ||||
| D14 | 19.2 ± 0.5 a | 19.9 ± 0.2 a | 21.1 ± 0.4 b | 22.5 ± 0.4 c |
| D18 | 20.8 ± 0.5 | 20.9 * ± 0.2 | 20.7 ± 0.5 | 20.5 * ± 0.3 |
| Ratio T4/T3 | ||||
| D14 | 4.5 ± 0.7 ab | 8.9 ± 0.3 c | 6.4 ± 0.9 b | 3.3 ± 0.4 a |
| D18 | 4.0 ± 0.1 a | 7.7 ± 0.8 b | 6.0 ± 1.2 ab | 3.9 ± 1.3 a |
| Pituitary Markers | Gene Name and Abbreviation | Fw/Rv a | Primer Sequence (5’-3’) | Ta b | Bp c |
|---|---|---|---|---|---|
| Hormones | Growth Hormone | Fw | CCGAGGAACCAGATTTCACCCAA | 62 | 110 |
| (gh) | Rv | TGGAGGGCGGAGCTATCAGGGA | |||
| Thyroid stimulating hormone | Fw | GTGTTCCCTTTCTGGCTCTTTTTTC | 52 | 100 | |
| (tsh) | Rv | ACTCACACTCTGGTCTCTCCACGTA | |||
| Proopiomelanocortin A | Fw | CTTGAAGAAACCAAATGAACATC | 60 | 162 | |
| (pomca) | Rv | GAAACAGCCAATGAAGACCTAA | |||
| Proopiomelanocortin B | Fw | GCTCGTTAGCAGACCAAT | 62 | 76 | |
| (pomcb) | Rv | CAAAACACTCTCTCTTCATCTCT | |||
| Receptors | Thyroid receptor alfa | Fw | GAGGCCGGAGCCAAACAC | 60 | 124 |
| (trα) | Rv | GCCGATATCATCCGACAGG | |||
| Thyroid receptor beta | Fw | ACCGACTGGAGCCCACACAG | 60 | 129 | |
| (trβ) | Rv | CCTTCACCCACGCTGCACT | |||
| Glucocorticoid receptor | Rv | CCATCACCTCTGCCGCATCTG | 64 | 195 | |
| (gr) | Fw | TCTGGAGGAACTGCTGCTGAACC | |||
| Reference | 18S ribosomal RNA | Fw | TGACGGAAGGGCACCACCAG | 60 | 158 |
| (18s) | Rv | AATCGCTCCACCAACTAAGAACGG |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sánchez-Nuño, S.; Silva, S.C.; Guerreiro, P.M.; Ordóñez-Grande, B.; Sanahuja, I.; Fernández-Alacid, L.; Ibarz, A. Modulation of Pituitary Response by Dietary Lipids and Throughout a Temperature Fluctuation Challenge in Gilthead Sea Bream. Fishes 2019, 4, 55. https://0-doi-org.brum.beds.ac.uk/10.3390/fishes4040055
Sánchez-Nuño S, Silva SC, Guerreiro PM, Ordóñez-Grande B, Sanahuja I, Fernández-Alacid L, Ibarz A. Modulation of Pituitary Response by Dietary Lipids and Throughout a Temperature Fluctuation Challenge in Gilthead Sea Bream. Fishes. 2019; 4(4):55. https://0-doi-org.brum.beds.ac.uk/10.3390/fishes4040055
Chicago/Turabian StyleSánchez-Nuño, Sergio, Sandra C. Silva, Pedro M. Guerreiro, Borja Ordóñez-Grande, Ignasi Sanahuja, Laura Fernández-Alacid, and Antoni Ibarz. 2019. "Modulation of Pituitary Response by Dietary Lipids and Throughout a Temperature Fluctuation Challenge in Gilthead Sea Bream" Fishes 4, no. 4: 55. https://0-doi-org.brum.beds.ac.uk/10.3390/fishes4040055