
Mulberry Protection through Flowering-Stage Essential Oil of Artemisia annua against the Lesser Mulberry Pyralid, Glyphodes pyloalis Walker


 ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Insects’ Rearing
2.2. Essential Oil
2.2.1. Extraction of the Essential Oil
2.2.2. Determination of Essential Oil Composition
2.3. Insecticidal Activity
2.3.1. Oral Toxicity
2.3.2. Fumigant Activity
2.4. Digestive Enzymes’ Assays
2.4.1. The α-Amylase Activity
2.4.2. Protease Assay
2.4.3. Lipase Estimation
2.4.4. The α- and β-Glucosidase Estimation
2.5. Detoxifying Enzymes’ Assays
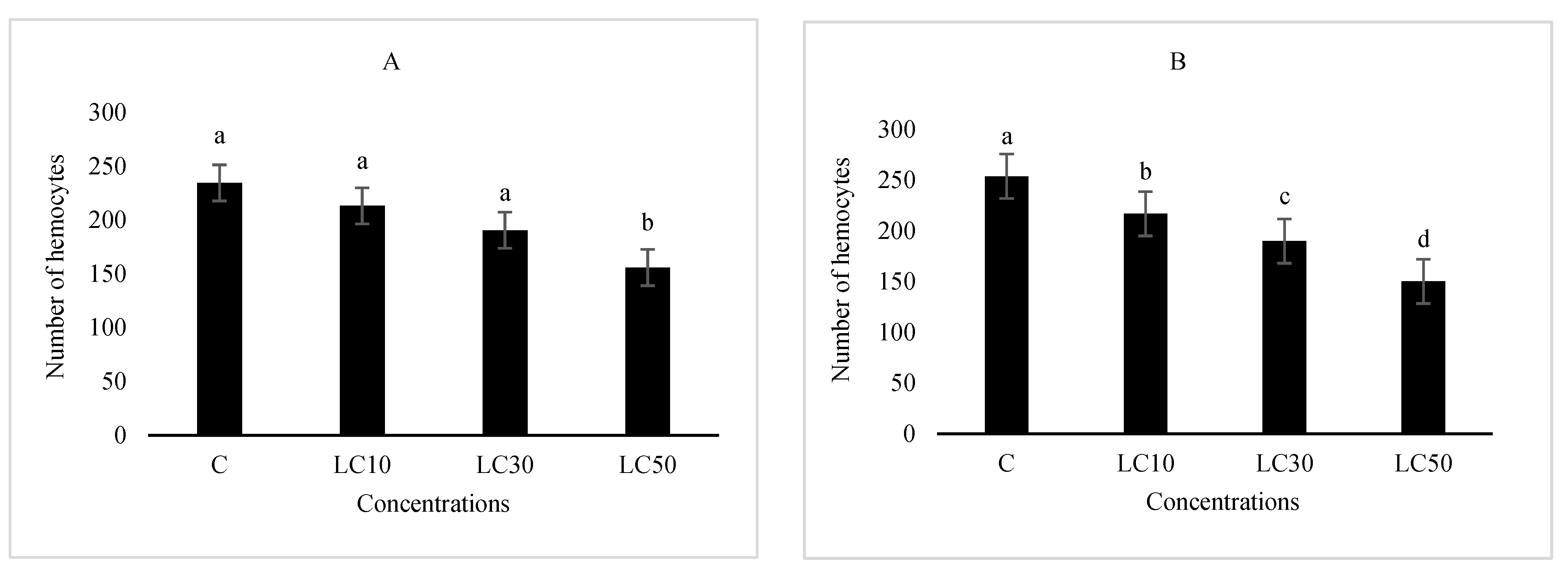
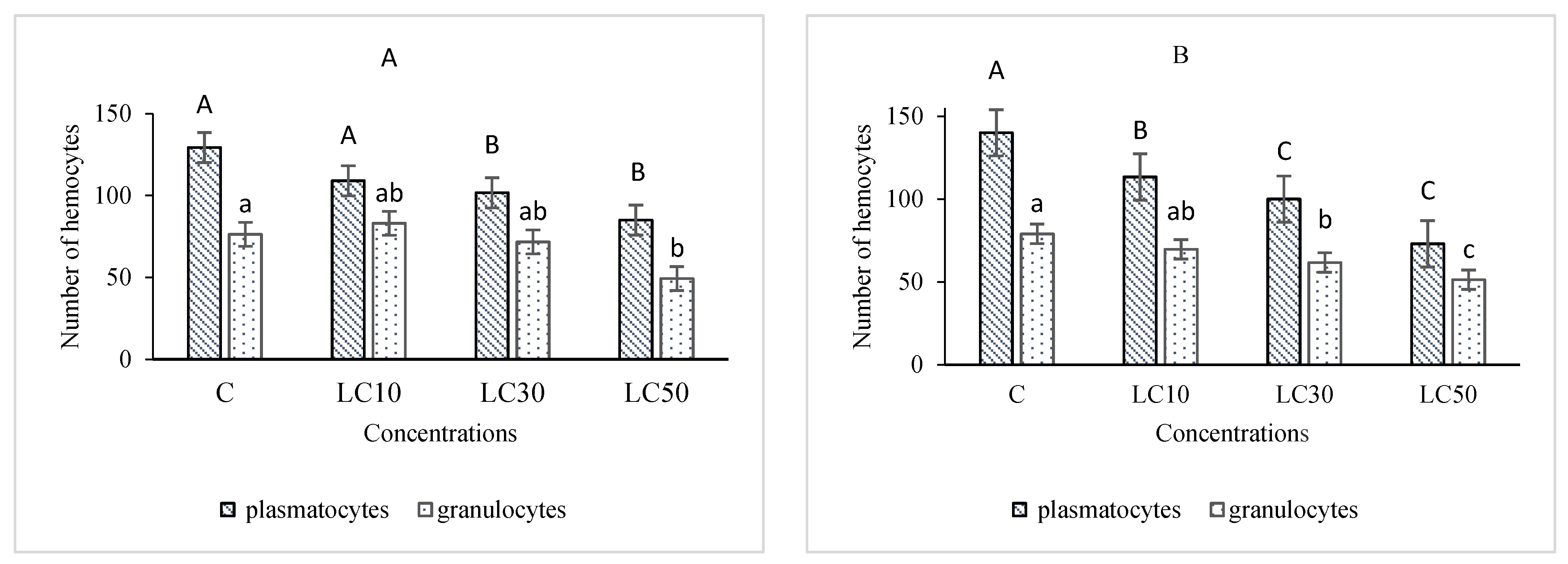
2.6. Hematological Study
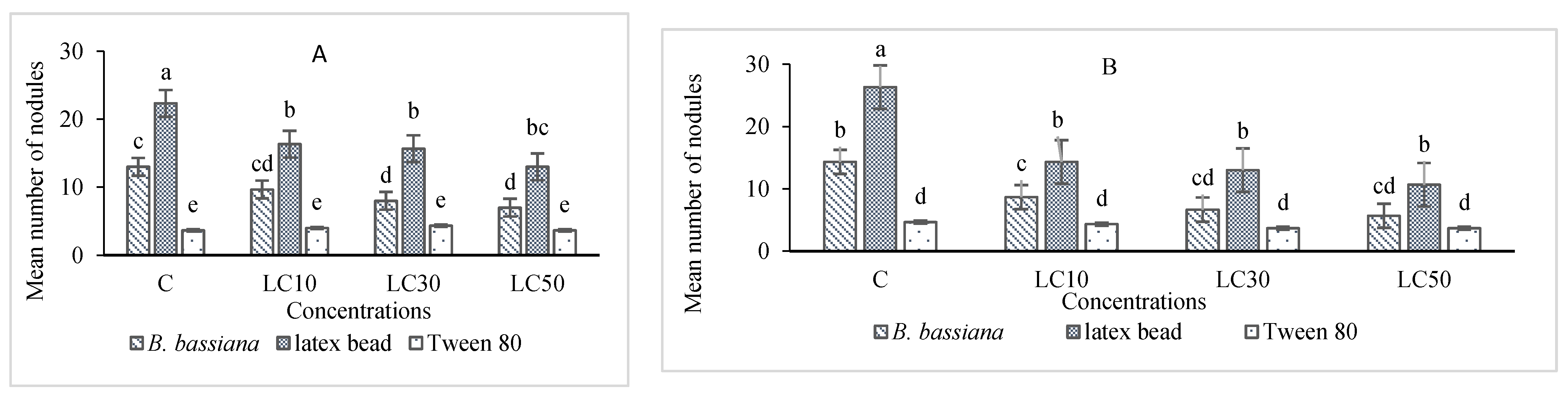
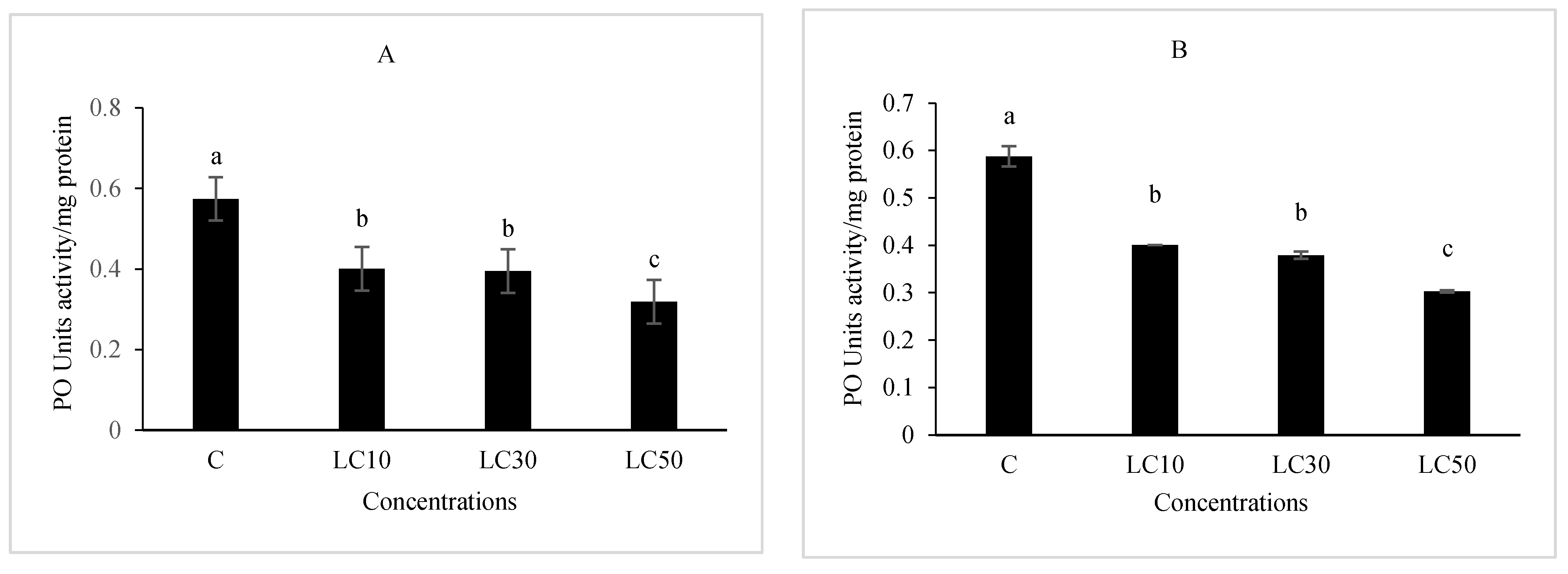
Immunity Responses
2.7. Histological Studies of Larvae Midgut and Adults’ Ovary
2.8. Statistical Analysis
3. Results
3.1. A. annua Essential Oil Analysis
3.2. Insecticidal Activity
3.3. Energy Reserves
3.4. Digestive and Detoxifying Enzymes
3.5. Hematological Study and Immunity Responses
3.6. Histological Studies
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Khosravi, R.; Sendi, J.J. Biology and demography of Glyphodes pyloalis Walker (Lepidoptera: Pyralidae) on mulberry. J. Asia-Pacific Èntomol. 2010, 13, 273–276. [Google Scholar] [CrossRef]
- Isman, M.B. Plant essential oils for pest and disease management. Crop. Prot. 2000, 19, 603–608. [Google Scholar] [CrossRef]
- Lamichhane, J.R.; Dachbrodt-Saaydeh, S.; Kudsk, P.; Messéan, A. Toward a reduced reliance on conventional pesticides in european agriculture. Plant. Dis. 2016, 100, 10–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El Asbahani, A.; Miladi, K.; Badri, W.; Sala, M.; Addi, E.H.A.; Casabianca, H.; El Mousadik, A.; Hartmann, D.; Jilale, A.; Renaud, F.N.R.; et al. Essential oils: From extraction to encapsulation. Int. J. Pharm. 2015, 483, 220–243. [Google Scholar] [CrossRef] [PubMed]
- Isman, M.B.; Grieneisen, M.L. Botanical insecticide research: Many publications, limited useful data. Trends Plant. Sci. 2014, 19, 140–145. [Google Scholar] [CrossRef]
- Ebadollahi, A.; Ziaee, M.; Palla, F. Essential oils extracted from different species of the Lamiaceae plant family as prospective bioagents against several detrimental pests. Molecules 2020, 25, 1556. [Google Scholar] [CrossRef] [Green Version]
- Basaid, K.; Chebli, B.; Mayad, E.H.; Furze, J.N.; Bouharroud, R.; Krier, F.; Barakate, M.; Paulitz, T. Biological activities of essential oils and lipopeptides applied to control plant pests and diseases: A review. Int. J. Pest. Manag. 2020, 1–23. [Google Scholar] [CrossRef]
- Campos, E.V.; Proença, P.L.; Oliveira, J.L.; Bakshi, M.; Abhilash, P.; Fraceto, L.F. Use of botanical insecticides for sustainable agriculture: Future perspectives. Ecol. Indic. 2019, 105, 483–495. [Google Scholar] [CrossRef] [Green Version]
- Nathan, S.S.; Choi, M.-Y.; Paik, C.-H.; Seo, H.-Y. Food consumption, utilization, and detoxification enzyme activity of the rice leaffolder larvae after treatment with Dysoxylum triterpenes. Pestic. Biochem. Physiol. 2007, 88, 260–267. [Google Scholar] [CrossRef]
- Lazarevic, J.; Jevremović, S.; Kostić, I.; Kostić, M.; Vuleta, A.; Jovanović, S.M.; Jovanović, D. Šešlija Toxic, oviposition deterrent and oxidative stress effects of Thymus vulgaris essential oil against Acanthoscelides obtectus. Insects 2020, 11, 563. [Google Scholar] [CrossRef]
- Afraze, Z.; Sendi, J.J.; Karimi-Malati, A.; Zibaee, A. Methanolic extract of winter cherry causes morpho-histological and immunological ailments in mulberry pyralid Glyphodes pyloalis. Front. Physiol. 2020, 11, 908. [Google Scholar] [CrossRef] [PubMed]
- Konovalov, D.; Khamilonov, A.A. Biologically active compounds of Artemisia annua essential oil. Pharm. Pharmacol. 2016, 4, 4–33. [Google Scholar] [CrossRef] [Green Version]
- Xiao, L.; Tan, H.; Zhang, L. Artemisia annua glandular secretory trichomes: The biofactory of antimalarial agent artemisinin. Sci. Bull. 2016, 61, 26–36. [Google Scholar] [CrossRef]
- Shekari, M.; Sendi, J.J.; Etebari, K.; Zibaee, A.; Shadparvar, A. Effects of Artemisia annua L. (Asteracea) on nutritional physiology and enzyme activities of elm leaf beetle, Xanthogaleruca luteola Mull. (Coleoptera: Chrysomellidae). Pestic. Biochem. Physiol. 2008, 91, 66–74. [Google Scholar] [CrossRef]
- Hasheminia, S.M.; Sendi, J.J.; Jahromi, K.T.; Moharramipour, S. The effects of Artemisia annua L. and Achillea millefolium L. crude leaf extracts on the toxicity, development, feeding efficiency and chemical activities of small cabbage Pieris rapae L. (Lepidoptera: Pieridae). Pestic. Biochem. Physiol. 2011, 99, 244–249. [Google Scholar] [CrossRef]
- Zibaee, A. Botanical Insecticides and Their Effects on Insect Biochemistry and Immunity. In Pesticides in the Modern World—Pests Control and Pesticides Exposure and Toxicity Assessment; IntechOpen: London, UK, 2011. [Google Scholar]
- Mojarab-Mahboubkar, M.; Sendi, J.J.; Aliakbar, A. Effect of Artemisia annua L. essential oil on toxicity, enzyme activities, and energy reserves of cotton bollworm Helicoverpa armigera (Hübner) (Lepidoptera: Noctuidae). J. Plant. Prot. Res. 2015, 55, 371–377. [Google Scholar] [CrossRef]
- Oftadeh, M.; Sendi, J.J.; Ebadollahi, A. Toxicity and deleterious effects of Artemisia annua essential oil extracts on mulberry pyralid (Glyphodes pyloalis). Pestic. Biochem. Physiol. 2020, 170, 104702. [Google Scholar] [CrossRef]
- Adams, R.P. Identification of Essential Oil Components by Gas. Chromatography/Mass Spectrometry, 4th ed.; Allured Publishing Corporation: Carol Stream, IL, USA, 2007. [Google Scholar]
- NIST. NIST17; National Institute of Standards and Technology: Gaithersburg, MD, USA, 2017. [Google Scholar]
- Bernfeld, P. Amylases alpha and beta. Methods Enzymol. 1955, 1, 140–146. [Google Scholar]
- García-Carreño, F.L.; Haard, N.F. Characterization of proteinase classes in Langostilla (Pleuroncodes planipes) and Crayfish (Pacifastacus astacus) extracts. J. Food Biochem. 1993, 17, 97–113. [Google Scholar] [CrossRef]
- Tsujita, T.; Ninomiya, H.; Okuda, H. P-nitrophenyl 865. buyrate hydrolyzing activity of Hormone-senditive lipase 866 from bovine adipose tissue. J. Lipid. Res. 1989, 30, 867–997. [Google Scholar] [CrossRef]
- Ferreira, C.; Terra, W.R. Physical and kinetic properties of a plasma-membrane-bound β-d-glucosidase (cellobiase) from midgut cells of an insect (Rhynchosciara americana larva). Biochem. J. 1983, 213, 43–51. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-Dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Siegert, K.J. Carbohydrate metabolism in Manduca sexta during late larval development. J. Insect Physiol. 1987, 33, 421–427. [Google Scholar] [CrossRef]
- Van Asperen, K. A study of housefly esterases by means of a sensitive colorimetric method. J. Insect Physiol. 1962, 8, 401–416. [Google Scholar] [CrossRef]
- Habing, W.H.; Pabst, M.J.; Jakoby, W.B. Glutathione S-transferases. The first step in mercapturic acid formation. J. Biol. Chem. 1974, 249, 7130–7139. [Google Scholar]
- Parkinson, N.M.; Weaver, R.J. Noxious Components of Venom from the Pupa-Specific Parasitoid Pimpla hypochondriaca. J. Invertebr. Pathol. 1999, 73, 74–83. [Google Scholar] [CrossRef]
- El-Aziz, N.M.A.; Awad, H.H. Changes in the haemocytes of Agrotis ipsilon larvae (Lepidoptera: Noctuidae) in relation to dimilin and Bacillus thuringiensis infections. Micron 2010, 41, 203–209. [Google Scholar] [CrossRef]
- Jones, J.C. Current concepts concerning insect hemocytes. Am. Zoöl. 1962, 2, 209–246. [Google Scholar] [CrossRef] [Green Version]
- Arnold, J.W.; Hinks, C.F. Haemopoiesis in Lepidoptera. I. The multiplication of circulating haemocytes. Can. J. Zool. 1976, 54, 1003–1012. [Google Scholar] [CrossRef]
- Seyedtalebi, F.S.; Safavi, S.; Talaei-Hasanloui, A.R.; Bandani, A.R. Quantitative comparison for some immune responses among Eurygaster integriceps, Ephestia kuehniella and Zophobas morio against the entomopathogenic fungus Beuveria bassiana. Invert. Surviv. J. 2017, 14, 174–181. [Google Scholar] [CrossRef]
- LeOra Software. Polo Plus, a User’s Guide to Probit or Logit Analysis; LeOra Software: Berkeley, CA, USA, 2002. [Google Scholar]
- SAS Institute. SAS/STAT User’s Guide for Personal Computers; SAS Institute: Cary, NC, USA, 1997. [Google Scholar]
- Mojarab-Mahboubkar, M.; Sendi, J.J. Chemical composition, insecticidal and physiological effect of methanol extract of sweet wormwood (Artemisia annua L.) on Helicoverpa armigera (Hübner) (Lepidoptera: Noctuidae). Toxin Rev. 2016, 35, 106–115. [Google Scholar] [CrossRef]
- Bedini, S.; Flamini, G.; Cosci, F.; Ascrizzi, R.; Echeverria, M.C.; Guidi, L.; Landi, M.; Benelli, G.; Conti, B. Artemisia spp. essential oils against the disease-carrying blowfly Calliphora vomitoria. Parasit. Vectors 2017, 10, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pandey, A.K.; Singh, P. The Genus Artemisia: A 2012–2017 Literature review on chemical composition, antimicrobial, insecticidal and antioxidant activities of essential oils. Medicines 2017, 4, 68. [Google Scholar] [CrossRef] [Green Version]
- Nigam, M.; Atanassova, M.; Mishra, A.P.; Pezzani, R.; Devkota, H.P.; Plygun, S.; Salehi, B.; Setzer, W.N.; Sharifi-Rad, J. Bio-active compounds and health benefits of Artemisia species. Nat Prod. Commun 2019. [Google Scholar] [CrossRef] [Green Version]
- Isman, M.B. Commercial development of plant essential oils and their constituents as active ingredients in bioinsecticides. Phytochem. Rev. 2020, 19, 235–241. [Google Scholar] [CrossRef]
- Kumar, P.; Mishra, S.; Malik, A.; Satya, S. Repellent, larvicidal and pupicidal properties of essential oils and their formulations against the housefly, Musca domestica. Med. Veter Èntomol. 2011, 25, 302–310. [Google Scholar] [CrossRef] [PubMed]
- Filomeno, C.A.; Barbosa, L.C.; Teixeira, R.R.; Pinheiro, A.L.; Farias, E.D.S.; Ferreira, J.S.; Picanço, M.C. Chemical diversity of essential oils of Myrtaceae species and their insecticidal activity against Rhyzopertha dominica. Crop. Prot. 2020, 137, 105309. [Google Scholar] [CrossRef]
- Shannag, H.K.; Capinera, J.L.; Freihat, N.M. Effects of neem-based insecticides on consumption and utilization of food in larvae of Spodoptera eridania (Lepidoptera: Noctuidae). J. Insect Sci. 2015, 15, 152. [Google Scholar] [CrossRef] [Green Version]
- Zou, C.; Wang, Y.; Zou, H.; Ding, N.; Geng, N.; Cao, C.; Zhang, G. Sanguinarine in Chelidonium majus induced antifeeding and larval lethality by suppressing food intake and digestive enzymes in Lymantria dispar. Pestic. Biochem. Physiol. 2019, 153, 9–16. [Google Scholar] [CrossRef]
- Zibaee, A.; Bandani, A.R. A study on the toxicity of the medicinal plant, Artemisia annua L. (Astracea) extracts the Sunn pest, Eurygaster integriceps Puton (Heteroptera: Scutelleridae). J. Plant. Prot. Res. 2010, 50, 48–54. [Google Scholar] [CrossRef]
- Bezzar-Bendjazia, R.; Kilani-Morakchi, S.; Maroua, F.; Aribi, N. Azadirachtin induced larval avoidance and antifeeding by disruption of food intake and digestive enzymes in Drosophila melanogaster (Diptera: Drosophilidae). Pestic. Biochem. Physiol. 2017, 143, 135–140. [Google Scholar] [CrossRef] [PubMed]
- Magierowicz, K.; Górska-Drabik, E.; Sempruch, C. The effect of Tanacetum vulgare essential oil and its main components on some ecological and physiological parameters of Acrobasis advenella (Zinck.) (Lepidoptera: Pyralidae). Pestic. Biochem. Physiol. 2020, 162, 105–112. [Google Scholar] [CrossRef] [PubMed]
- Mishra, M.; Sharma, A.; Dagar, V.S.; Kumar, S. Effects of β-sitosterol on growth, development and midgut enzymes of Helicoverpa armigera Hübner. Arch. Biol. Sci. 2020, 72, 271–278. [Google Scholar] [CrossRef]
- Klowden, M.J. Physiological Systems in Insects, 2nd ed.; Academic Press: Cambridge, MA, USA, 2007. [Google Scholar]
- Wu, M.-Y.; Ying, Y.-Y.; Zhang, S.-S.; Li, X.-G.; Yan, W.-H.; Yao, Y.-C.; Shah, S.; Wu, G.; Yang, F.-L. Effects of diallyl trisulfide, an active substance from garlic essential oil, on energy metabolism in male moth Sitotroga cerealella (Olivier). Insects 2020, 11, 270. [Google Scholar] [CrossRef]
- Nattudurai, G.; Baskar, K.; Paulraj, M.G.; Islam, V.I.H.; Ignacimuthu, S.; Duraipandiyan, V. Toxic effect of Atalantia monophylla essential oil on Callosobruchus maculatus and Sitophilus oryzae. Environ. Sci. Pollut. Res. 2016, 24, 1619–1629. [Google Scholar] [CrossRef]
- Perazzolo, L.M.; Gargioni, R.; Ogliari, P.; Barracco, M.A. Evaluation of some hemato-immunological parameters in the shrimp Farfantepenaeus paulensis submitted to environmental and physiological stress. Aquac. 2002, 214, 19–33. [Google Scholar] [CrossRef]
- Kraaijeveld, A.R.; Limentani, E.C.; Godfray, H.C.J. Basis of the trade-off between parasitoid resistance and larval competitive ability in Drosophila melanogaster. In Proceedings of the Royal Society B: Biological Sciences; The Royal Society: London, UK, 2001; Volume 268, pp. 259–261. [Google Scholar]
- Ghasemi, V.; Yazdi, A.K.; Tavallaie, F.Z.; Sendi, J.J. Effect of essential oils from Callistemon viminalis and Ferula gummosaon toxicity and on the hemocyte profile of Ephestia kuehniella (Lep.: Pyralidae). Arch. Phytopathol. Plant. Prot. 2014, 47, 268–278. [Google Scholar] [CrossRef]
- Sadeghi, R.; Eshrati, M.R.; Mortazavian, S.M.M.; Jamshidnia, A. The Effects of the essential oils isolated from four ecotypes of cumin (Cuminum cyminum L.) on the blood cells of the pink stem borer, Sesamia cretica Ledere (Lepidoptera: Noctuidae). J. Kans. Èntomol. Soc. 2019, 92, 390–399. [Google Scholar] [CrossRef]
- Clark, K.D.; Strand, M.R. Hemolymph melanization in the silkmoth bombyx mori involves formation of a high molecular mass complex that metabolizes tyrosine. J. Biol. Chem. 2013, 288, 14476–14487. [Google Scholar] [CrossRef] [Green Version]
- Khanikor, B.; Bora, D. Effect of plant based essential oil on immune response of silkworm, Antheraea assama Westwood (Lepidoptera: Saturniidae). Int. J. Ind. Èntomol. 2012, 25, 139–146. [Google Scholar] [CrossRef] [Green Version]
- Ghoneim, K. Disturbed hematological and immunological parameters of insects by botanicals as an effective approach of pest control: A review of recent progress. South Asian. J. Exp. Biol. 2018, 1, 112–144. [Google Scholar]
- Rahimi, V.; Hajizadeh, J.; Zibaee, A.; Sendi, J.J. Changes in immune responses of Helicoverpa armigera Hübner followed by feeding on Knotgrass, Polygonum persicaria agglutinin. Arch. Insect Biochem. Physiol. 2019, 101, e21543. [Google Scholar] [CrossRef] [PubMed]
- Cerenius, L.; Lee, B.L.; Söderhäll, K. The proPO-system: Pros and cons for its role in invertebrate immunity. Trends Immunol. 2008, 29, 263–271. [Google Scholar] [CrossRef] [PubMed]
- Murfadunnisa, S.; Vasantha-Srinivasan, P.; Ganesan, R.; Senthil-Nathan, S.; Kim, T.-J.; Ponsankar, A.; Kumar, S.D.; Chandramohan, D.; Krutmuang, P. Larvicidal and enzyme inhibition of essential oil from Spheranthus amaranthroids (Burm.) against lepidopteran pest Spodoptera litura (Fab.) and their impact on non-target earthworms. Biocatal. Agric. Biotechnol. 2019, 21. [Google Scholar] [CrossRef]
- Riddiford, L.M. How does juvenile hormone control insect metamorphosis and reproduction? Gen. Comp. Endocrinol. 2012, 179, 477–484. [Google Scholar] [CrossRef]
- Reis, T.C.; Soares, M.A.; Dos Santos, J.B.; Dos Santos, C.A.; Serrão, J.E.; Zanuncio, J.C.; Ferreira, E.A. Atrazine and nicosulfuron affect the reproductive fitness of the predator Podisus nigrispinus (Hemiptera: Pentatomidae). Anais da Academia Brasileira de Ciências 2018, 90, 3625–3633. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
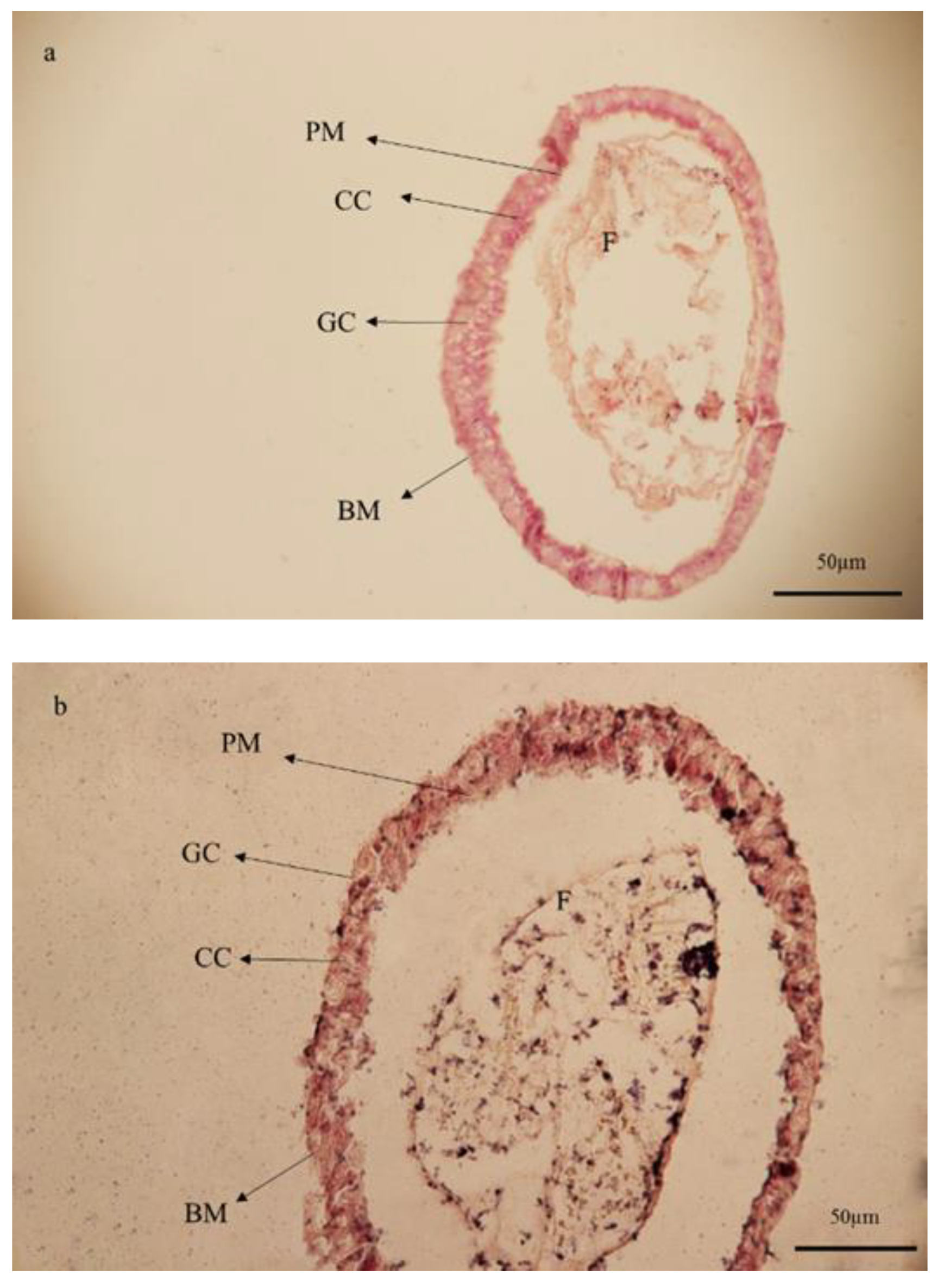{kind=link}
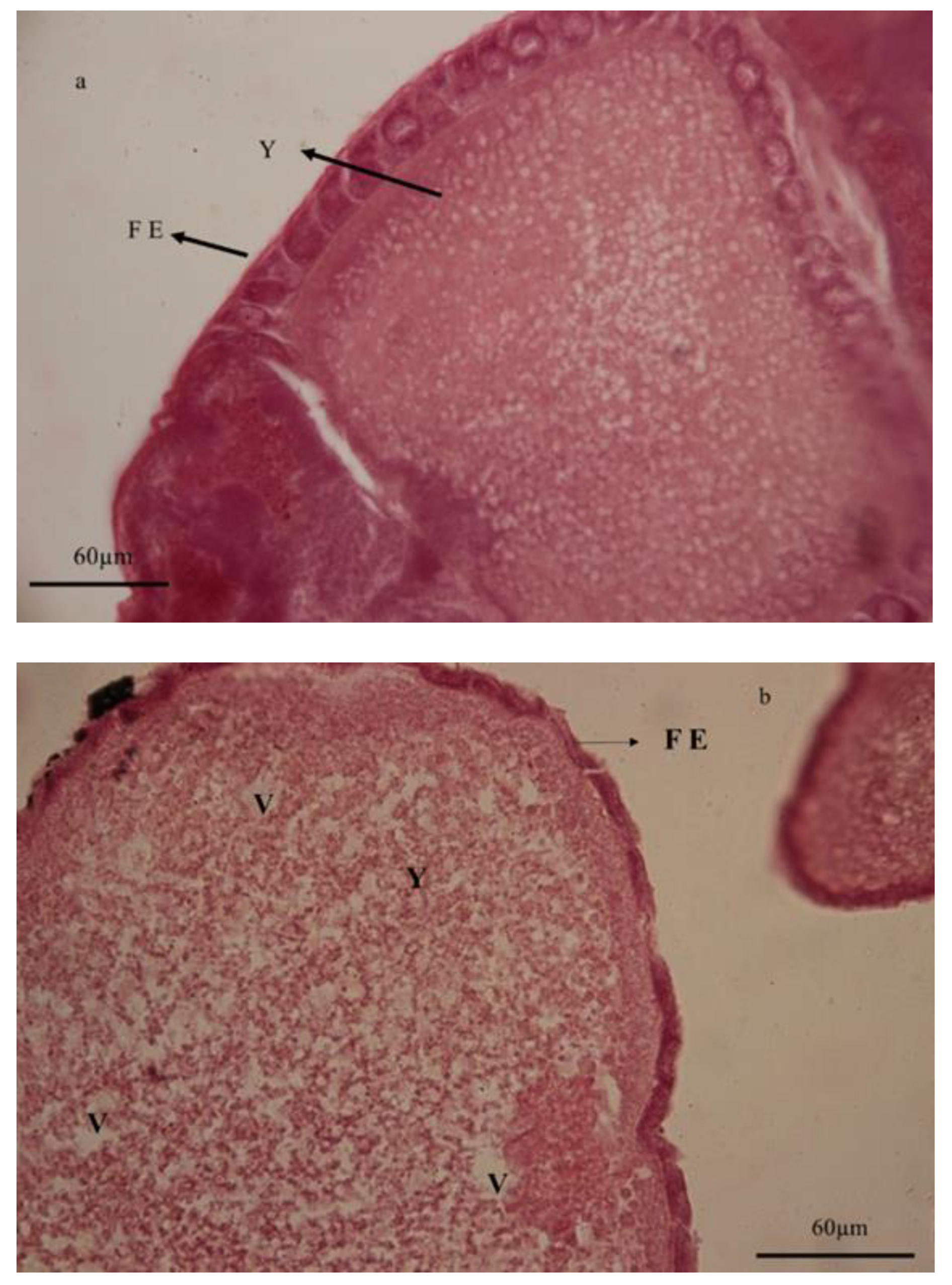{kind=link}
| RIcalc | RIdb | Compound | % | RIcalc | RIdb | Compound | % |
|---|---|---|---|---|---|---|---|
| 923 | 926 | Tricyclene MH | 0.2 | 1258 | 1259 | Lepalone OM | 0.1 |
| 938 | 939 | α-Pinene MH | 5.9 | 1281 | 1278 | Lepalol OM | 0.3 |
| 978 | 975 | Sabinene MH | 0.3 | 1299 | 1290 | p-Cymen-7-ol OM | 0.2 |
| 982 | 979 | β-Pinene MH | 0.1 | 1337 | 1327 | p-Mentha-1,4-dien-7-ol OM | 0.2 |
| 992 | 990 | Myrcene MH | 0.4 | 1361 | 1359 | Eugenol PP | 0.6 |
| 1013 | 999 | Yomogi alcohol OM | 1.2 | 1374 | 1376 | α-Copaene SH | 1.0 |
| 1021 | 1024 | p-Cymene MH | 0.8 | 1391 | 1392 | Benzyl 2-methylbutanoate E | 0.3 |
| 1026 | 1026 | o-Cymene MH | 0.8 | 1402 | 1392 | (Z)-Jasmone OC | 0.1 |
| 1030 | 1031 | 1,8-Cineole OM | 6.8 | 1420 | 1419 | (E)-β-Caryophyllene SH | 3.1 |
| 1061 | 1062 | Artemisia ketone OM | 11.8 | 1426 | 1432 | β-Copaene SH | 0.2 |
| 1074 | 1070 | cis-Sabinene hydrate OM | 0.5 | 1448 | 1454 | α-Humulene SH | 0.3 |
| 1082 | 1083 | Artemisia alcohol OM | 1.4 | 1455 | 1456 | (E)-β-Farnesene SH | 1.0 |
| 1104 | 1114 | 3-Methyl-3-butenyl 3-methylbutanoate E | 0.8 | 1471 | 1477 | β-Chamigrene SH | 0.2 |
| 1119 | 1126 | α-Campholenal OM | 0.7 | 1478 | 1485 | Germacrene D SH | 0.7 |
| 1131 | 1144 | trans-Pinocarveol OM | 0.4 | 1489 | 1490 | β-Selinene SH | 10.7 |
| 1144 | 1146 | Camphor OM | 13.1 | 1510 | 1516 | Isobornyl isovalerate OM | 0.1 |
| 1161 | 1164 | Pinocarvone OM | 7.4 | 1517 | 1523 | δ-Cadinene SH | 0.1 |
| 1169 | 1169 | Borneol OM | 1.5 | 1547 | 1555 | iso-Caryophyllene oxide OS | 0.3 |
| 1179 | 1177 | Terpinene-4-ol OM | 2.2 | 1585 | 1583 | Caryophyllene oxide OS | 5.4 |
| 1192 | 1188 | α-Terpineol OM | 0.9 | 1588 | 1590 | β-Copaene-4α-ol OS | 0.2 |
| 1199 | 1195 | Myrtenol OM | 2.6 | 1594 | 1594 | Salvial-4(14)-en-1-one OS | 0.2 |
| 1211 | 1205 | Verbenone OM | 0.3 | 1643 | 1640 | Caryophylla-4(12),8(13)-dien-5β-ol OS | 1.3 |
| 1219 | 1216 | trans-Carveol OM | 0.6 | 1700 | 1695 | Germacra-4(15),5,10(14)-trien-1β-ol OS | 1.7 |
| 1227 | 1230 | cis-p-Mentha-1(7),8-dien-2-ol OM | 0.2 | 1765 | 1767 | β-Costol OS | 1.3 |
| 1229 | 1235 | (3Z)-Hexenyl 3-methylbutanoate E | 0.2 | 1854 | 1847 | Phytone OC | 0.4 |
| 1234 | 1236 | n-Hexyl 2-methylbutanoate E | 0.1 | 1984 | 1960 | Palmitic acid OC | 1.2 |
| 1240 | 1241 | Cuminaldehyde OM | 0.2 | 2087 | 2106 | Phytol DT | 0.3 |
| 1244 | 1243 | Carvone OM | 0.1 | Total identified | 93.0 | ||
| Bioassay | LC10 (95% CL) | LC30 (95% CL) | LC50 (95% CL) | LC90 (95% CL) | Slope ± SE | X2 (df = 3) |
|---|---|---|---|---|---|---|
| Oral toxicity | 0.593 (0.395–0.735) | 0.901 (0.725–1.058) | 1.204 (1.024–1.466) | 2.445 (1.882–4.128) | 4.165 ± 0.631 | 3.2567 |
| Fumigant toxicity | 1.945 (1.568–2.240) | 2.678 (2.347–2.948) | 3.343 (3.048–3.632) | 5.745 (5.112–6.825) | 5.449 ± 0.788 | 2.976 |
| Bio-assay | Concentrations | Protein (mg/dL) | Glucose (mg/dL) | Triglyceride (mg/dL) |
|---|---|---|---|---|
| Oral toxicity (% W/V) | Control | 1.0200 ± 0.0360 a | 1.7733 ± 0.0247 a | 1.8800 ± 0.0145 a |
| LC10 | 0.9833 ± 0.0088 a | 1.6666 ± 0.0033 a | 1.8033 ± 0.0617 a | |
| LC30 | 0.9700 ± 0.0057 a | 1.6533 ± 0.0290 a | 1.6557 ± 0.0531 a | |
| LC50 | 0.9533 ± 0.0088 a | 1.1733 ± 0.0783 b | 1.1700 ± 0.0577 b | |
| F-Value | 2.16 | 29.51 | 19.65 | |
| Pr | 0.0170 | 0.0001 | 0.0005 | |
| Fumigant toxicity (μL/L) | Control | 1.0400 ± 0.0208 a | 1.8100 ± 0.0655 a | 1.9200 ± 0.0964 a |
| LC10 | 0.9900 ± 0.0057 ab | 1.7266 ± 0.0384 a | 1.7533 ± 0.0635 ab | |
| LC30 | 0.9700 ± 0.0032 b | 1.6900 ± 0.0208 a | 1.433 ± 0.2185 ab | |
| LC50 | 0.9366 ± 0.0088 b | 1.1633 ± 0.0317 b | 1.3000 ± 0.0765 b | |
| F-Value | 12.94 | 47.80 | 5.04 | |
| Pr | 0.0019 | 0.0001 | 0.0300 |
| Bio-assay | Digestive Enzymes (U/mg Protein) | Control | LC10 | LC30 | LC50 | F-Value | Pr |
|---|---|---|---|---|---|---|---|
| Oral toxicity (% W/V) | Protease | 1.9467 ± 0.3525 a | 1.7833 ± 0.1201 ab | 1.5433 ± 0.0876 ab | 1.0667 ± 0.0437 b | 3.96 | 0.0531 |
| α-glucosidase | 1.374 ± 0.192 a | 1.046 ± 0.0825 ab | 0.7119 ± 0.0333 b | 0.5640 ± 0.0360 b | 9.31 | 0.0055 | |
| β-glucosidase | 1.4451 ± 0.1165 a | 1.1635 ± 0.0955 a | 0.8757 ± 0.05365 b | 0.6873 ± 0.0515 b | 15.61 | 0.0010 | |
| α-amylase | 0.3066 ± 1.732 a | 0.2633 ± 0.0202 ab | 0.2333 ± 0.01763 b | 0.0833 ± 0.0120 c | 41.32 | 0.0001 | |
| Lipase | 0.0571 ± 0.032 a | 0.0387 ± 0.064 ab | 0.03806 ± 0.089 b | 0.03700 ± 0.059 b | 22.75 | 0.0003 | |
| Fumigant toxicity (μL/L) | Protease | 1.8333 ± 0.1244 a | 0.8967 ± 0.1197 b | 0.7167 ± 0.1591 b | 0.4067 ± 0.1591 b | 15.83 | 0.0010 |
| α-glucosidase | 1.2034 ± 0.039 a | 1.1083 ± 0.266 a | 0.8870 ± 0.064 b | 0.6921 ± 0.038 b | 20.80 | 0.0004 | |
| β-glucosidase | 1.3451 ± 0.0330 a | 1.3183 ± 0.1830 a | 0.9537 ± 0.0282 ab | 0.7591 ± 0.0717 b | 8.07 | 0.0084 | |
| α-amylase | 0.2800 ± 0.0057 a | 0.2700 ± 0.01731 ab | 0.2300 ± 0.11541 ab | 0.1333 ± 0.0145 b | 37.49 | 0.0001 | |
| Lipase | 0.0559 ± 0.0010 a | 0.0436 ± 0.0012 b | 0.0378 ± 0.0027 b | 0.02620 ± 0.0025 c | 37.68 | 0.0001 |
| Bio-assay | Concentrations | GST (U/mg Protein) | Esterase (U/mg Protein) | |
|---|---|---|---|---|
| Oral toxicity (% W/V) | Control | 0.02300 ± 0.001 a | 0.0953 ± 0.004 a | |
| LC 10 | 0.01733 ± 0.0032 a | 0.08266 ± 0.007 ab | ||
| LC 30 | 0.0065 ± 0.0025 b | 0.07366 ± 0.002 ab | ||
| LC 50 | 0.0001 ± 0.00001 b | 0.06700 ± 0.001 b | ||
| F-Value | 23.46 | 14.13 | ||
| Pr | 0.0003 | 0.0483 | ||
| Fumigant toxicity (μL/L) | Control | 0.02266 ± 0.0008 a | 0.09566 ± 0.004 a | |
| LC 10 | 0.01533 ± 0.0006 a | 0.07966 ± 0.0005 ab | ||
| LC 30 | 0.0010 ± 0.0001 b | 0.06066 ± 0.0063 ab | ||
| LC 50 | 0.0001 ± 0.0000 b | 0.04600 ± 0.0024 b | ||
| F-Value | 30.13 | 22.27 | ||
| Pr | 0.0001 | 0.0003 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oftadeh, M.; Sendi, J.J.; Ebadollahi, A.; Setzer, W.N.; Krutmuang, P. Mulberry Protection through Flowering-Stage Essential Oil of Artemisia annua against the Lesser Mulberry Pyralid, Glyphodes pyloalis Walker. Foods 2021, 10, 210. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10020210
Oftadeh M, Sendi JJ, Ebadollahi A, Setzer WN, Krutmuang P. Mulberry Protection through Flowering-Stage Essential Oil of Artemisia annua against the Lesser Mulberry Pyralid, Glyphodes pyloalis Walker. Foods. 2021; 10(2):210. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10020210
Chicago/Turabian StyleOftadeh, Marziyeh, Jalal Jalali Sendi, Asgar Ebadollahi, William N. Setzer, and Patcharin Krutmuang. 2021. "Mulberry Protection through Flowering-Stage Essential Oil of Artemisia annua against the Lesser Mulberry Pyralid, Glyphodes pyloalis Walker" Foods 10, no. 2: 210. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10020210