
Evaluation of Recombinant Kpkt Cytotoxicity on HaCaT Cells: Further Steps towards the Biotechnological Exploitation Yeast Killer Toxins


 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Microorganisms, Cloning Vectors, and Growth Media
2.2. Production of Native and Recombinant Kpkt in Flask and Bioreactor
2.3. Cell-Free Supernatant Ultrafiltration and Lyophilization
2.4. Well Plate Assay for the Evaluation of Killer Activity and the Determination of Killer Toxin Concentration
2.5. Evaluation of β-Glucanase Activity
2.6. Human Cell Line Toxicity Test
2.7. Data Analysis
3. Results and Discussion
3.1. Scale up of Recombinant Kpkt Production
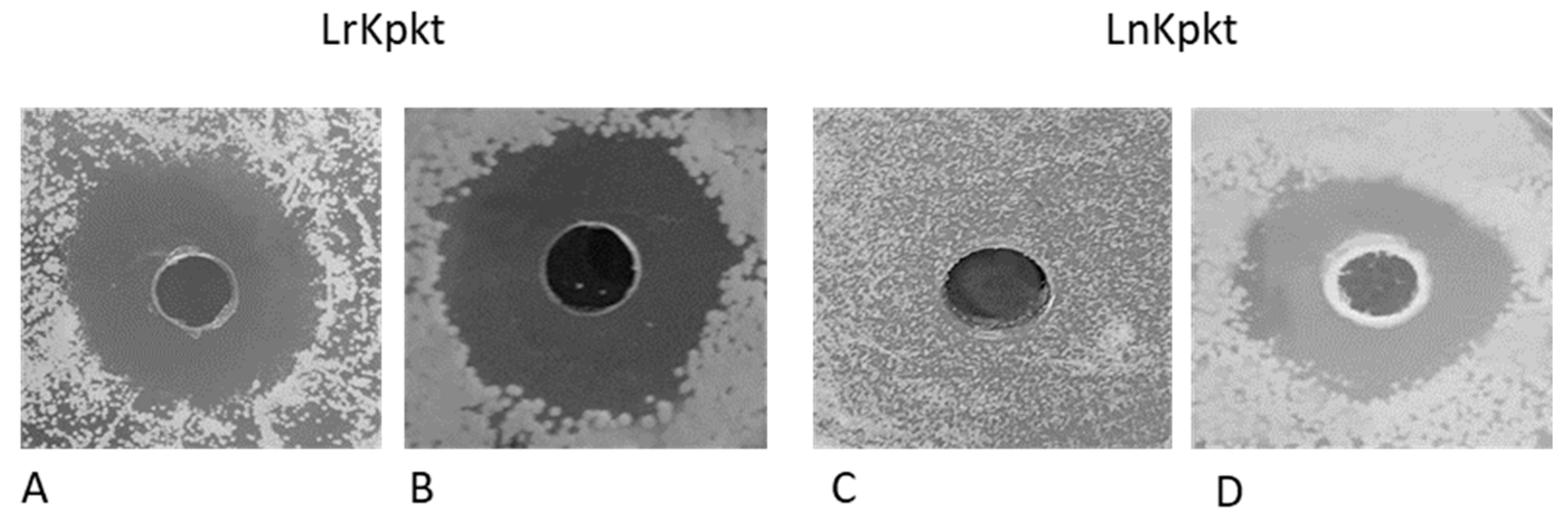
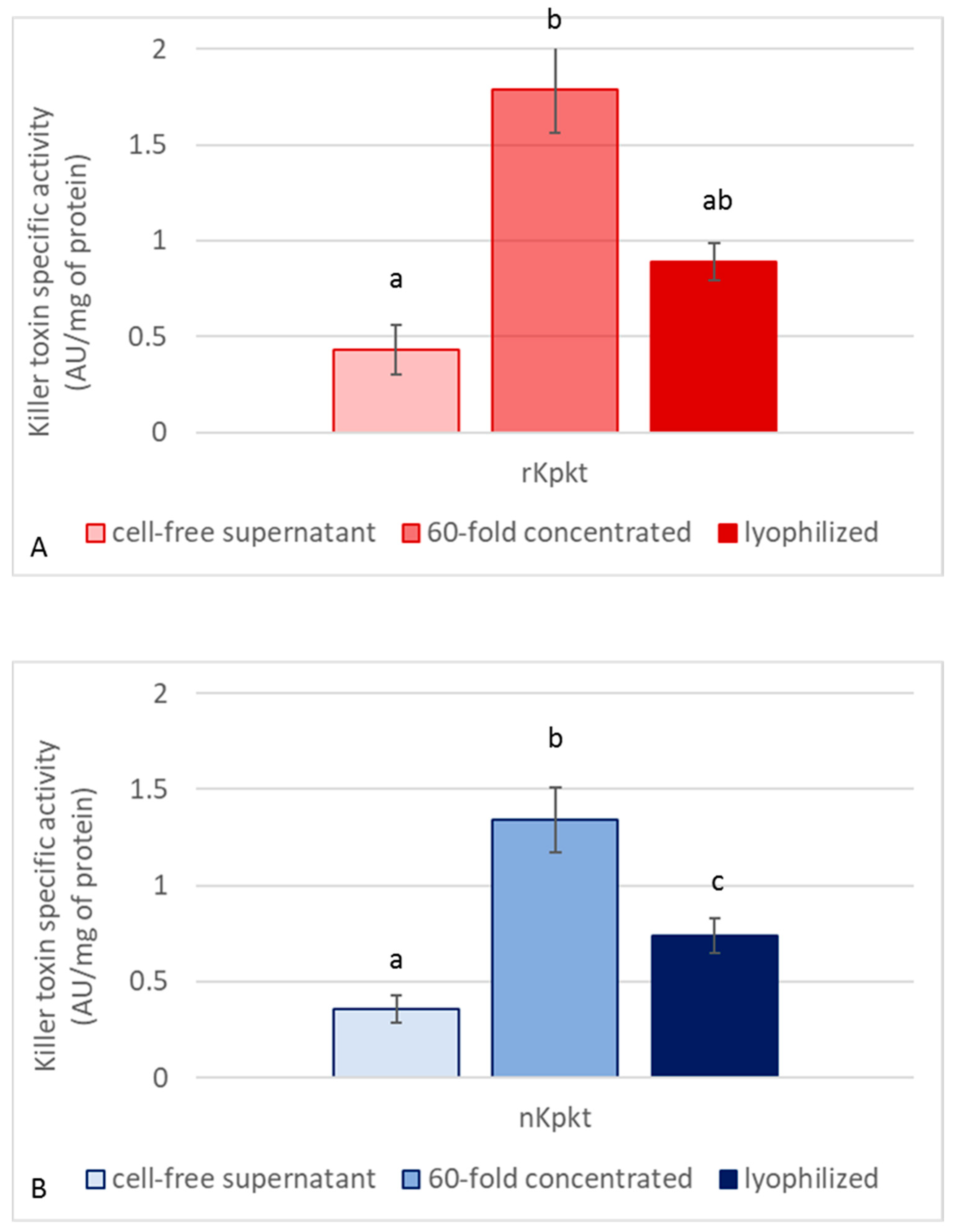
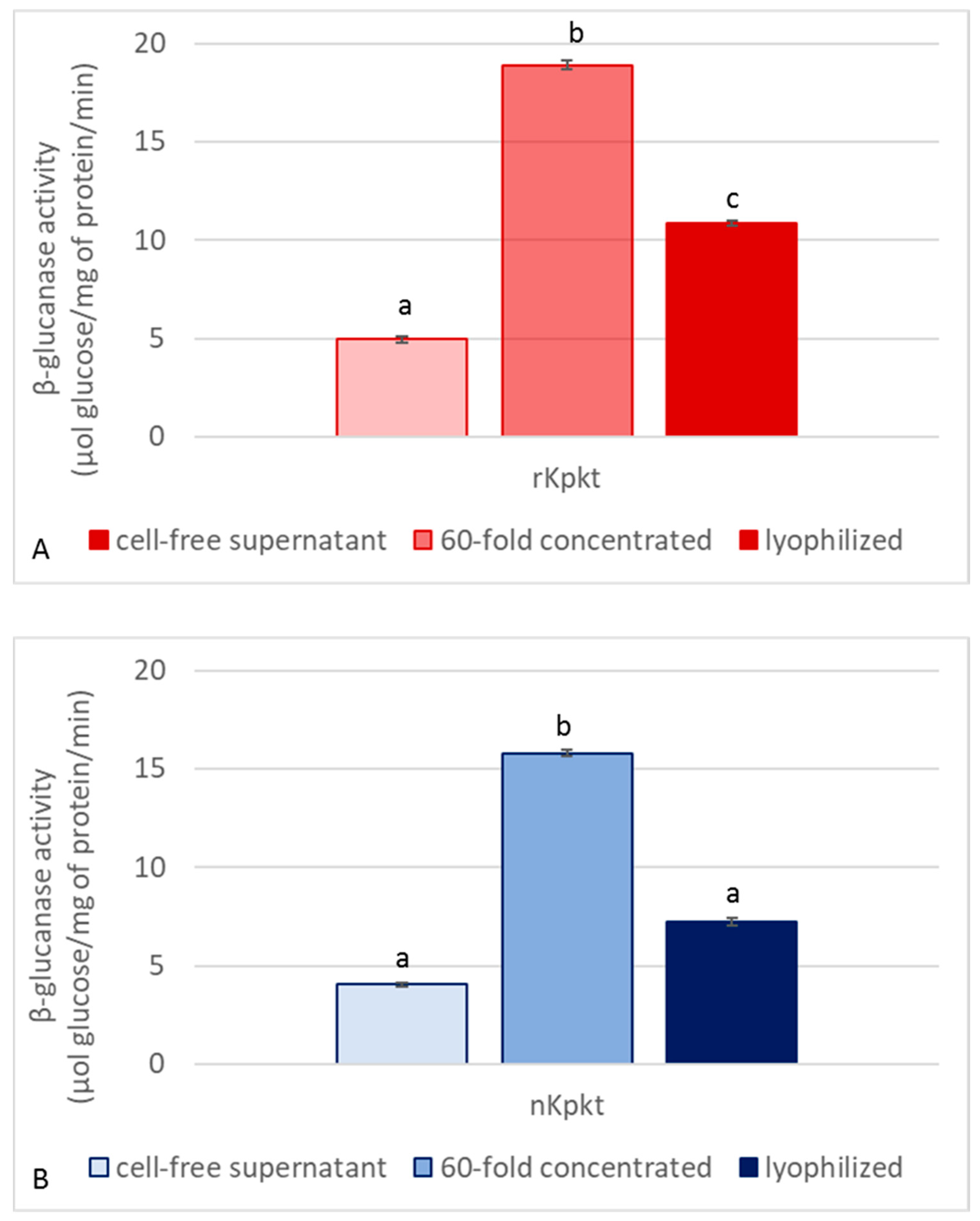
3.2. Production of Lyophilized Preparations of Kpkt and Evaluation of Killer and β-Glucanase Activities
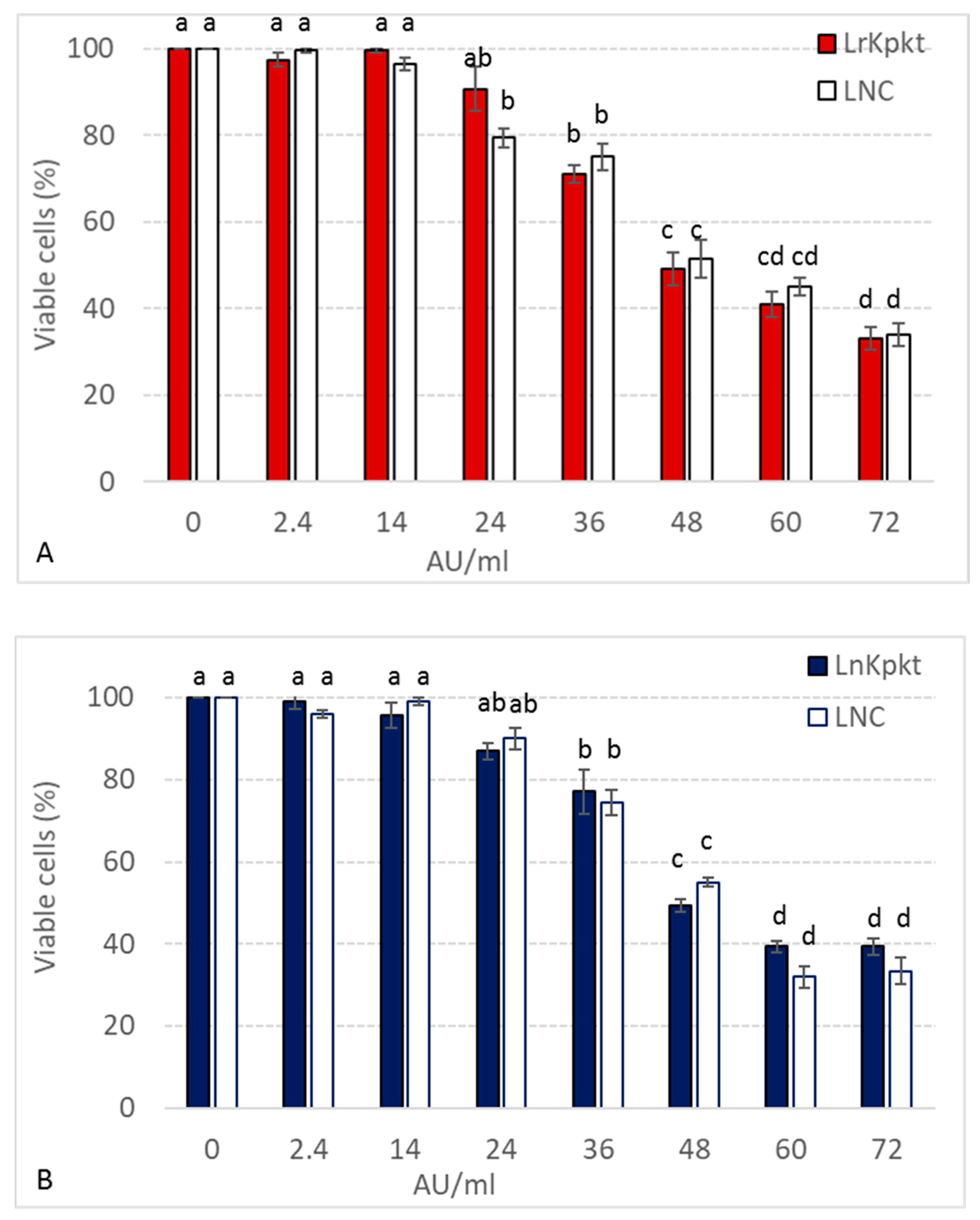
3.3. Cytotoxicity of rKpkt and nKpkt on HaCaT Cell Lines
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
Ethical Statement
References
- Klassen, R.; Schaffrath, R.; Buzzini, P.; Ganter, P.F. Antagonistic interactions and killer yeasts. In Yeasts in Natural Ecosystems: Ecology; Buzzini, P., Lachance, M.A., Yurkov, A., Eds.; Springer: Cham, Switzerland, 2017; pp. 229–275. [Google Scholar]
- Van Vuuren, H.J.J.; Jacobs, C.J. Killer yeasts in the wine industry: A review. Am. J. Enol. Vitic. 1992, 43, 119–128. [Google Scholar]
- Todd, B.E.N.; Fleet, G.H.; Henscke, P.A. Promotion of autolysis through the interaction of killer and sensitive yeasts: Potential application in sparkling wine production. Am. J. Enol. Vitic. 2000, 51, 65–72. [Google Scholar]
- Ciani, M.; Fatichenti, F. Killer toxin of Kluyveromyces phaffii DBVPG 6076 as a biopreservative agent to control apiculate wine yeasts. Appl. Environ. Microbiol. 2001, 67, 3058–3063. [Google Scholar] [CrossRef] [Green Version]
- Pérez, F.; Ramírez, M.; Regodón, J.A. Influence of killer strains of Saccharomyces cerevisiae on wine fermentation. Antonie van Leewen 2001, 79, 393–399. [Google Scholar] [CrossRef]
- Comitini, F.; De Ingeniis, J.; Pepe, L.; Mannazzu, I.; Ciani, M. Pichia anomala and Kluyveromyces wickeramii killer toxins as new tools against Dekkera/Brettanomyces spoilage yeasts. FEMS Microbiol. Lett. 2004, 238, 235–240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Comitini, F.; Di Pietro, N.; Zacchi, L.; Mannazzu, I.; Ciani, M. Kluyveromyces phaffii killer toxin active against wine spoilage yeasts: Purification and characterization. Microbiology 2004, 150, 2535–2541. [Google Scholar] [CrossRef] [Green Version]
- Velázquez, R.; Zamora, E.; Álvarez, M.L.; Hernández, L.M.; Ramírez, M. Effects of new Torulaspora delbrueckii killer yeasts on the must fermentation kinetics and aroma compounds of white table wine. Front. Microbiol. 2015, 6, 1222. [Google Scholar] [CrossRef] [Green Version]
- Ramírez, M.; Velázquez, R.; Maqueda, M.; Zamora, E.; López-Piñeiro, A.; Hernández, L.M. Influence of the dominance of must fermentation by Torulaspora delbrueckii on the malolactic fermentation and organoleptic quality of red table wine. Int. J. Food Microbiol. 2016, 238, 311–319. [Google Scholar] [CrossRef]
- Villalba, M.L.; Sàez, J.S.; del Monaco, S.; Lopes, C.A.; Sangorrín, M.P. TdKT, a new killer toxin produced by Torulaspora delbrueckii effective against wine spoilage yeasts. Int. J. Food Microbiol. 2016, 217, 94–100. [Google Scholar] [CrossRef] [PubMed]
- Velázquez, R.; Zamora, E.; Álvarez, M.L.; Ramírez, M. Using killer yeasts in the Torulaspora delbrueckii elaboration of base wine and traditional sparkling wine. Int. J. Food Microbiol. 2019, 289, 134–144. [Google Scholar] [CrossRef]
- Villalba, M.L.; Mazzucco, M.B.; Lopes, C.A.; Ganga, M.A.; Sangorrín, M.P. Purification and characterization of Saccharomyces eubayanus killer toxin: Biocontrol effectiveness against wine spoilage yeasts. Int. J. Food Microbiol. 2020, 108714. [Google Scholar] [CrossRef]
- Palpacelli, V.; Ciani, M.; Rosini, G. Activity of different ‘killer’ yeasts on strains of yeast species undesirable in the food industry. FEMS Microbiol. Lett. 1991, 68, 75–78. [Google Scholar] [CrossRef]
- Lowes, K.F.; Shearman, C.A.; Payne, J.; MacKenzie, D.; Archer, D.B.; Merry, R.J.; Gasson, M.J. Prevention of yeast spoilage in feed and food by the yeast mycocin HMK. Appl. Environ. Microbiol. 2000, 66, 1066–1076. [Google Scholar] [CrossRef] [Green Version]
- Seguy, N.; Cailliez, J.; Polonelli, L.; Dei-Cas, E.; Camus, D. Inhibitory effect of a Pichia anomala killer toxin on Pneumocystis carinii infectivity to the SCID mouse. Parasitol. Res. 1996, 82, 114–116. [Google Scholar] [CrossRef]
- Izgü, F.; Altinby, D. Killer toxins of certain yeast strains have potential growth inhibitory activity on gram-positive pathogenic bacteria. Microbios 1997, 89, 15–22. [Google Scholar] [PubMed]
- Theisen, S.; Molkenau, E.; Schmitt, M.J. Wicaltin, a new protein toxin secreted by the yeast Williopsis californica and its broad-spectrum antimycotic potential. J. Microbiol. Biotechnol. 2000, 10, 547–550. [Google Scholar]
- Carboni, G.; Fancello, F.; Zara, G.; Zara, S.; Ruiu, L.; Marova, I.; Pinna, G.; Budroni, M.; Mannazzu, I. Production of a lyophilized ready-to-use yeast killer toxin with possible applications in the wine and food industries. Int. J. Food Microbiol. 2020, 335, 108883. [Google Scholar] [CrossRef]
- Weiler, F.; Schmitt, M.J. Zygocin a secreted antifungal toxin of the yeast Zygosaccharomyces bailii and its effect on sensitive fungal cells. FEMS Yeast Res. 2003, 3, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Santos, A.; Sanchez, A.; Marquina, D. Yeasts as biological agents to control Botrytis cinerea. Microbiol. Res. 2004, 159, 331–338. [Google Scholar] [CrossRef]
- Pérez, M.F.; Contreras, L.; Garnica, N.M.; Fernandez-Zenoff, F.V.; Farías, M.E.; Sepulveda, M.; Ramallo, J.; Dib, J.R. Native killer yeasts as biocontrol agents of postharvest fungal diseases in lemons. PLoS ONE 2016, 11, e0165590. [Google Scholar] [CrossRef] [Green Version]
- Rosa-Magri, M.M.; Tauk-Tornisielo, S.M.; Ceccato-Antonini, S.R. Bioprospection of yeasts as biocontrol agents against phytopathogenic molds. Braz. Arch. Biol. Technol. 2011, 54, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Shaffrath, R.; Meinhardt, F.; Klassen, R. Yeast Killer toxins: Fundamentals and Applications. In Physiology and Genetics. The Mycota: A Comprehensive Treatise on Fungi as Experimental Systems for Basic and Applied Research; Anke, T., Schüffler, A., Eds.; Springer: Cham, Switzerland, 2018; pp. 87–118. [Google Scholar]
- Mannazzu, I.; Domizio, P.; Carboni, G.; Zara, S.; Zara, G.; Comitini, F.; Budroni, M.; Ciani, M. Yeast killer toxins: From ecological significance to application. Crit. Rev. Biotechnol. 2019, 39, 603–617. [Google Scholar] [CrossRef] [PubMed]
- Mehlomakulu, N.N.; Setati, M.E.; Divol, B. Non-Saccharomyces killer toxins: Possible biocontrol agents against Brettanomyces in wine? S. Afr. J. Enol. Vitic. 2015, 36, 94–104. [Google Scholar] [CrossRef] [Green Version]
- Comitini, F.; Mannazzu, I.; Ciani, M. Tetrapisispora phaffii killer toxin is a highly specific β-glucanase that disrupts the integrity of the yeast cell wall. Microb. Cell Factories 2009, 8, 55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oro, L.; Zara, S.; Fancellu, F.; Mannazzu, I.; Budroni, M.; Ciani, M.; Comitini, F. TpBGL2 codes for a Tetrapisispora phaffii killer toxin active against wine spoilage yeasts. FEMS Yeast Res. 2014, 14, 464–471. [Google Scholar] [CrossRef] [Green Version]
- Comitini, F.; Ciani, M. The zymocidial activity of Tetrapisispora phaffii in the control of Hanseniaspora uvarum during the early stages of winemaking. Lett. Appl. Microbiol. 2010, 50, 50–56. [Google Scholar] [CrossRef] [PubMed]
- Chessa, R.; Landolfo, S.; Ciani, M.; Budroni, M.; Zara, S.; Ustun, M.; Çakar, Z.P.; Mannazzu, I. Biotechnological exploitation of Tetrapisispora phaffii killer toxin: Heterologous production in Komagataella phaffii (Pichia pastoris). Appl. Microbiol. Biotechnol. 2017, 101, 2931–2942. [Google Scholar] [CrossRef]
- Ahmad, M.; Hirz, M.; Pichler, H.; Schwab, H. Protein expression in Pichia pastoris: Recent achievements and perspectives for heterologous protein production. Appl. Microbiol. Biotechnol. 2014, 98, 5301–5317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfeiffer, P.F.; Radler, G.; Caspritz, H.H. Effect of a killer toxin of yeast on eukaryotic systems. Appl. Environ. Microbiol. 1988, 54, 1068–1069. [Google Scholar] [CrossRef] [Green Version]
- Pettoello-Mantovani, M.; Nocerino, A.; Polonelli, L.; Morace, G.; Conti, S.; Di Martino, L.; De Ritis, G.; Iafusco, M.; Guandalini, S. Hansenula anomala killer toxin induces secretion and severe acute injury in the rat intestine. Gastroenterology 1995, 109, 1900–1906. [Google Scholar] [CrossRef]
- Ross, M.H.; Pawlina, W. Histology: A Test and Atlas with Correlated. Cell and Molecular Biology, 7th ed.; Lippincott Williams & Wilkins: Baltimore, MD, USA, 2015. [Google Scholar]
- Kitagawa, N.; Otani, T.; Inai, T. Nisin, a food preservative produced by Lactococcus lactis, affects the localization pattern of intermediate filament protein in HaCaT cells. Anat. Sci. Int. 2019, 94, 163–171. [Google Scholar] [CrossRef] [Green Version]
- López-García, J.; Lehocký, M.; Humpolíček, P.; Sáha, P. HaCaT keratinocytes response on antimicrobial atelocollagen substrates: Extent of cytotoxicity, cell viability and proliferation. J. Funct. Biomater. 2014, 5, 43–57. [Google Scholar] [CrossRef] [Green Version]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein–dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Notario, V. β-glucanases from Candida albicans: Purification, characterization and the nature of their attachment to cell wall components. Microbiology 1982, 128, 747–759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, W.; Inan, M.; Meagher, M.M. Fermentation strategies for recombinant protein expression in the methylotrophic yeast Pichia pastoris. Biotechnol. Bioprocess Eng. 2000, 5, 275–287. [Google Scholar] [CrossRef] [Green Version]
- Inan, M.; Meagher, M.M. Non-repressing carbon sources for alcohol oxidase (AOX1) promoter of Pichia pastoris. J. Biosci. Bioeng. 2001, 92, 585–589. [Google Scholar] [CrossRef]
- Juturu, V.; Wu, J.C. Production of high concentration of l-lactic acid from oil palm empty fruit bunch by thermophilic Bacillus coagulans JI12. Biotechnol. Appl. Biochem. 2018, 65, 145–149. [Google Scholar] [CrossRef]
- Liu, W.-C.; Inwood, S.; Gong, T.; Sharma, A.; Yu, L.-Y.; Zhu, P. Fed-batch high-cell-density fermentation strategies for Pichia pastoris growth and production. Crit. Rev. Biotechnol. 2019, 39, 258–271. [Google Scholar] [CrossRef]
- Sun, Q.; Chen, F.; Geng, F.; Luo, Y.; Gong, S.; Jiang, Z. A novel aspartic protease from Rhizomucor miehei expressed in Pichia pastoris and its application on meat tenderization and preparation of turtle peptides. Food Chem. 2018, 245, 570–577. [Google Scholar] [CrossRef]
- Vina-Gonzalez, J.; Elbl, K.; Ponte, X.; Valero, F.; Alcalde, M. Functional expression of aryl-alcohol oxidase in Saccharomyces cerevisiae and Pichia pastoris by directed evolution. Biotechnol. Bioeng. 2018, 115, 1666–1674. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Source | Characteristic |
|---|---|---|
| Tetrapisispora phaffii 6076 | DBVPG | Native producer of Kpkt |
| Komagataella phaffii GS115 | Invitrogen | Host for heterologous expression, his4 |
| Saccharomyces cerevisiae 6500 | DBVPG | Sensitive to Kpkt and rKpkt |
| Dekkera bruxellensis 692 | DiSVA | Sensitive to rKpkt |
| Komagataella phaffii rc#24 | UNISS | negative control of killer activity [29] |
| Komagataella phaffii rc#17 | This study | pPIC9TpIMHisTag in GS115 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carboni, G.; Marova, I.; Zara, G.; Zara, S.; Budroni, M.; Mannazzu, I. Evaluation of Recombinant Kpkt Cytotoxicity on HaCaT Cells: Further Steps towards the Biotechnological Exploitation Yeast Killer Toxins. Foods 2021, 10, 556. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10030556
Carboni G, Marova I, Zara G, Zara S, Budroni M, Mannazzu I. Evaluation of Recombinant Kpkt Cytotoxicity on HaCaT Cells: Further Steps towards the Biotechnological Exploitation Yeast Killer Toxins. Foods. 2021; 10(3):556. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10030556
Chicago/Turabian StyleCarboni, Gavino, Ivana Marova, Giacomo Zara, Severino Zara, Marilena Budroni, and Ilaria Mannazzu. 2021. "Evaluation of Recombinant Kpkt Cytotoxicity on HaCaT Cells: Further Steps towards the Biotechnological Exploitation Yeast Killer Toxins" Foods 10, no. 3: 556. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10030556