
Biofilm-Forming Ability of Microbacterium lacticum and Staphylococcus capitis Considering Physicochemical and Topographical Surface Properties

 ,
,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Selected Bacterial Isolates and Culture Conditions
2.2. Surface Materials and Cleaning Procedure
2.3. Biofilm Formation on Model Surface Materials
2.4. Culture-Based Analysis of Biofilms
2.5. Surface Coverage and Cell Viability with Epifluorescence Microscopy (EFM)
2.6. Descriptive and Structural Evaluation of the Biofilm Matrix by Scanning Electron Microscopy (SEM)
2.7. Surface Topography Measurement
2.8. Physicochemical Surface Properties
2.9. Statistical Analysis
3. Results and Discussion
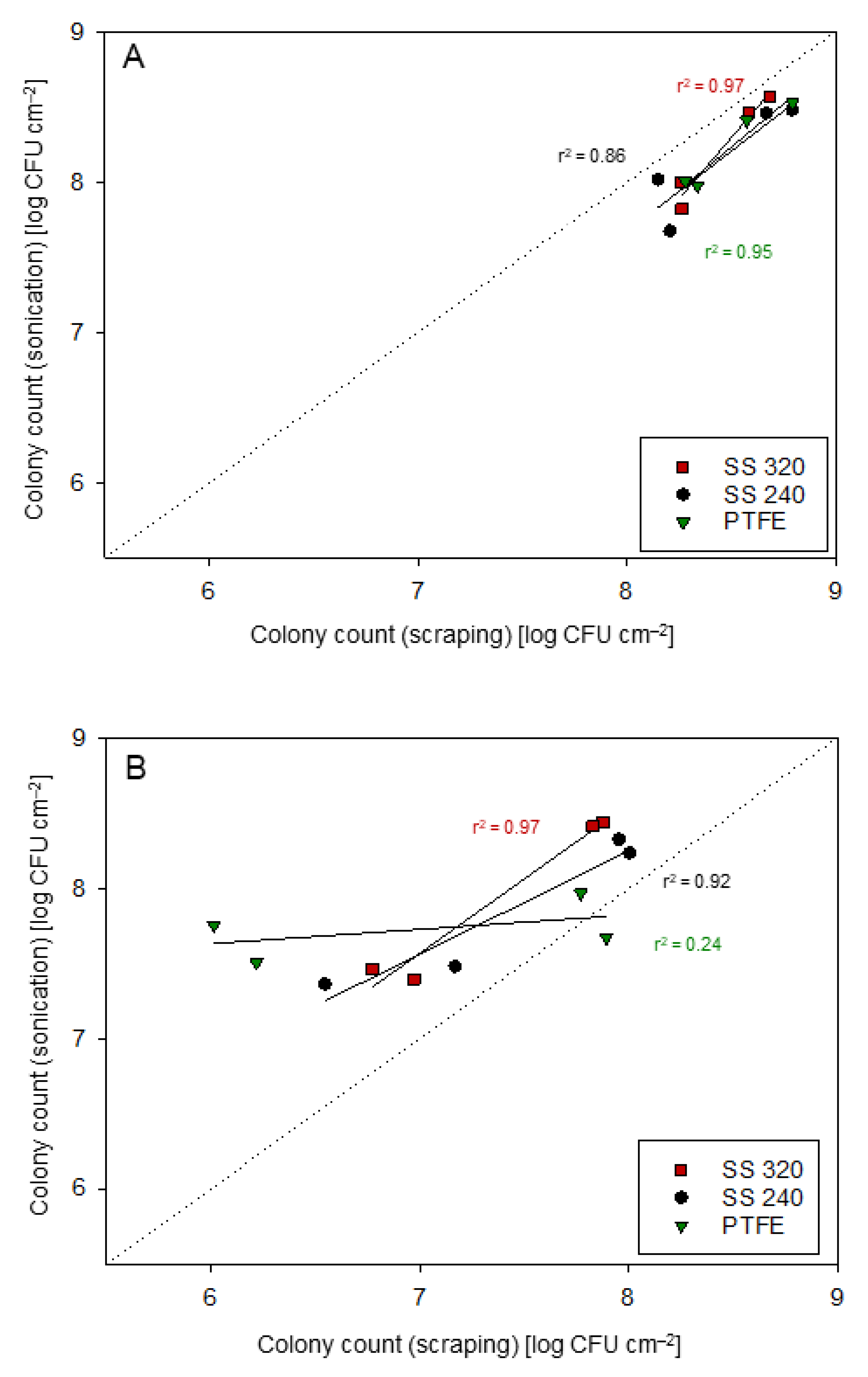
3.1. Comparison of the Bacterial Cell Detachment Approaches
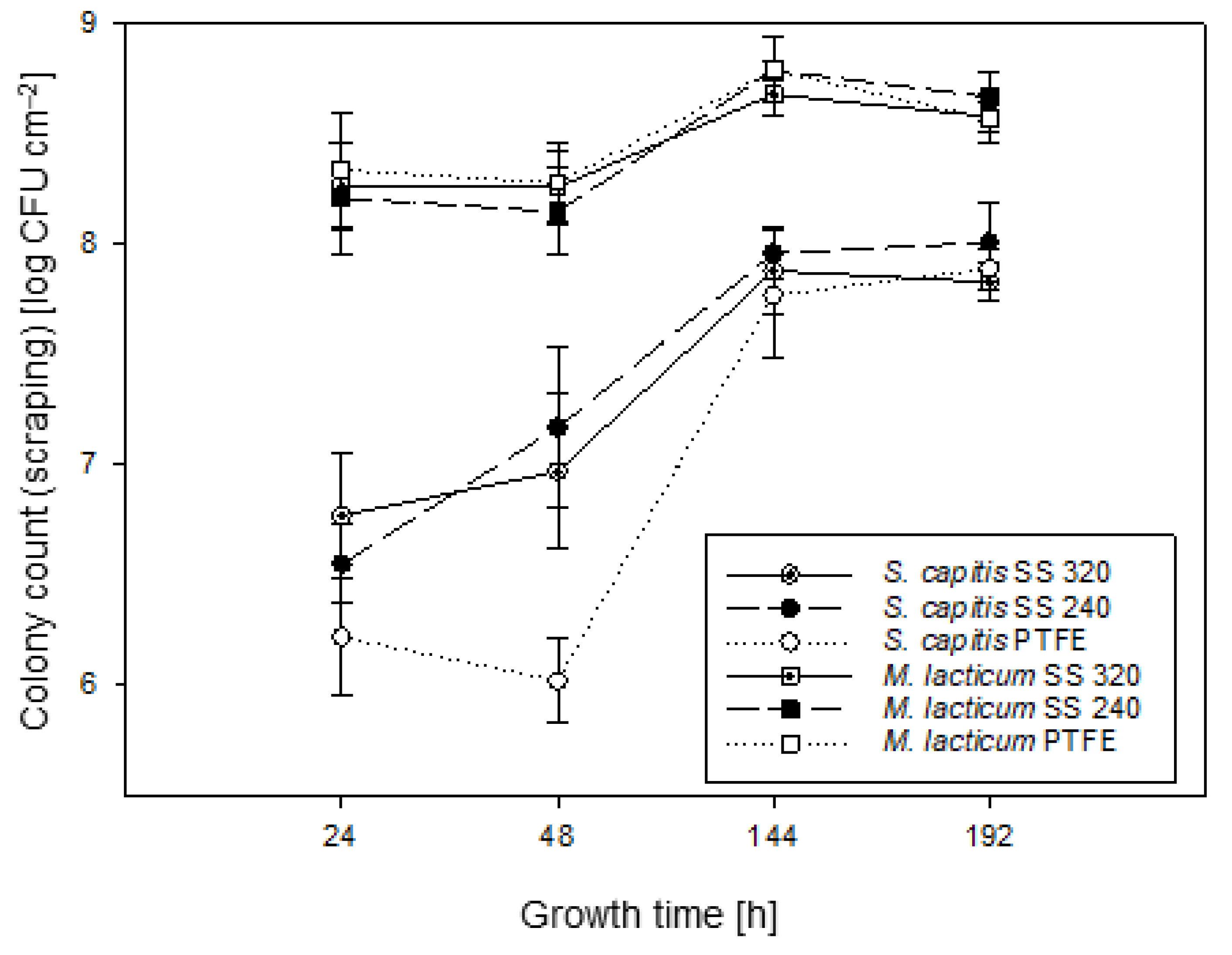
3.2. Culture-Based Enumeration of Biofilm Development Using the Scraping Technique
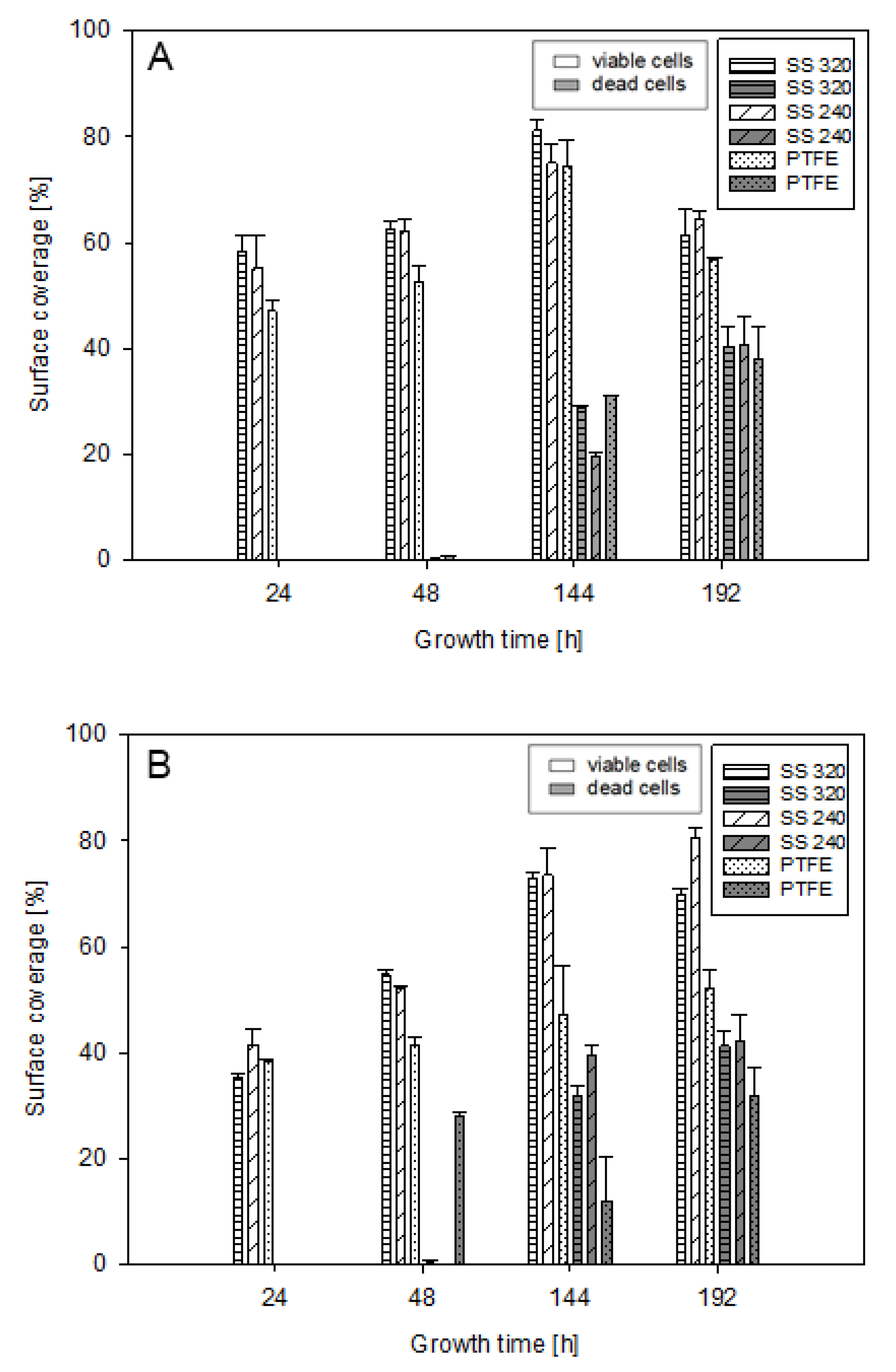
3.3. Surface Coverage and Biofilm Viability
3.4. Qualitative Structural and Morphological Biofilm Analysis
3.5. Effect of Surface Roughness on Biofilm Formation
3.6. Effect of Physicochemical and Interfacial Properties on Biofilm Formation
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Srey, S.; Jahid, I.K.; Ha, S.-D. Biofilm formation in food industries: A food safety concern. Food Control 2013, 31, 572–585. [Google Scholar] [CrossRef]
- Giaouris, E.; Heir, E.; Hébraud, M.; Chorianopoulos, N.; Langsrud, S.; Møretrø, T.; Habimana, O.; Desvaux, M.; Renier, S.; Nychas, G.-J. Attachment and biofilm formation by foodborne bacteria in meat processing environments: Causes, implications, role of bacterial interactions and control by alternative novel methods. Meat Sci. 2014, 97, 298–309. [Google Scholar] [CrossRef]
- Wang, R. Biofilms and Meat Safety: A Mini-Review. J. Food Prot. 2019, 82, 120–127. [Google Scholar] [CrossRef]
- Galiè, S.; García-Gutiérrez, C.; Miguélez, E.M.; Villar, C.J.; Lombó, F. Biofilms in the food industry: Health aspects and control methods. Front. Microbiol. 2018, 9, 898. [Google Scholar] [CrossRef] [PubMed]
- Alonso-Calleja, C.; Gómez-Fernández, S.; Carballo, J.; Capita, R. Prevalence, Molecular Typing, and Determination of the Biofilm-Forming Ability of Listeria monocytogenes Serotypes from Poultry Meat and Poultry Preparations in Spain. Microorganisms 2019, 7, 529. [Google Scholar] [CrossRef] [Green Version]
- Jamal, M.; Ahmad, W.; Andleeb, S.; Jalil, F.; Imran, M.; Nawaz, M.A.; Hussain, T.; Ali, M.; Rafiq, M.; Kamil, M.A. Bacterial biofilm and associated infections. J. Chin. Med. Assoc. 2018, 81, 7–11. [Google Scholar] [CrossRef]
- Bridier, A.; Briandet, R.; Thomas, V.; Dubois-Brissonnet, F. Resistance of bacterial biofilms to disinfectants: A review. Biofouling 2011, 27, 1017–1032. [Google Scholar] [CrossRef] [PubMed]
- Gazula, H.; Scherm, H.; Li, C.; Takeda, F.; Wang, P.; Chen, J. Ease of biofilm accumulation, and efficacy of sanitizing treatments in removing the biofilms formed, on coupons made of materials commonly used in blueberry packing environment. Food Control 2019, 104, 167–173. [Google Scholar] [CrossRef]
- Weber, M.; Liedtke, J.; Plattes, S.; Lipski, A. Bacterial community composition of biofilms in milking machines of two dairy farms assessed by a combination of culture-dependent and –independent methods. PLoS ONE 2019, 14, e0222238. [Google Scholar] [CrossRef]
- Karunakaran, E.; Mukherjee, J.; Ramalingam, B.; Biggs, C.A. “Biofilmology”: A multidisciplinary review of the study of microbial biofilms. Appl. Microbiol. Biotechnol. 2011, 90, 1869–1881. [Google Scholar] [CrossRef]
- Lianou, A.; Koutsoumanis, K.P. Strain variability of the biofilm-forming ability of Salmonella enterica under various environmental conditions. Int. J. Food Microbiol. 2012, 160, 171–178. [Google Scholar] [CrossRef]
- Zou, M.; Liu, D. A systematic characterization of the distribution, biofilm-forming potential and the resistance of the biofilms to the CIP processes of the bacteria in a milk powder processing factory. Food Res. Int. 2018, 113, 316–326. [Google Scholar] [CrossRef]
- Yong, Y.Y.; Dykes, G.A.; Choo, W.S. Biofilm formation by staphylococci in health-related environments and recent reports on their control using natural compounds. Crit. Rev. Microbiol. 2019, 45, 201–222. [Google Scholar] [CrossRef]
- Møretrø, T.; Hermansen, L.; Holck, A.L.; Sidhu, M.S.; Rudi, K.; Langsrud, S. Biofilm Formation and the Presence of the Intercellular Adhesion Locus ica among Staphylococci from Food and Food Processing Environments. Appl. Environ. Microbiol. 2003, 69, 5648. [Google Scholar] [CrossRef] [Green Version]
- Cherif-Antar, A.; Moussa–Boudjemâa, B.; Didouh, N.; Medjahdi, K.; Mayo, B.; Flórez, A.B. Diversity and biofilm-forming capability of bacteria recovered from stainless steel pipes of a milk-processing dairy plant. Dairy Sci. Technol. 2016, 96, 27–38. [Google Scholar] [CrossRef]
- Marchand, S.; De Block, J.; De Jonghe, V.; Coorevits, A.; Heyndrickx, M.; Herman, L. Biofilm Formation in Milk Production and Processing Environments; Influence on Milk Quality and Safety. Compr. Rev. Food Sci. Food Saf. 2012, 11, 133–147. [Google Scholar] [CrossRef]
- Otto, M. Staphylococcal biofilms. In Gram-Positive Pathogens; Fischetti, V.A., Novick, R.P., Ferretti, J.J., Portnoy, D.A., Braunstein, M., Rood, J.I., Eds.; ASM Press: Washington, DC, USA, 2019; pp. 699–711. [Google Scholar] [CrossRef]
- Fagerlund, A.; Langsrud, S.; Heir, E.; Mikkelsen, M.I.; Møretrø, T. Biofilm Matrix Composition Affects the Susceptibility of Food Associated Staphylococci to Cleaning and Disinfection Agents. Front. Microbiol. 2016, 7, 856. [Google Scholar] [CrossRef] [Green Version]
- Maes, S.; Heyndrickx, M.; Vackier, T.; Steenackers, H.; Verplaetse, A.; Reu, K.D. Identification and Spoilage Potential of the Remaining Dominant Microbiota on Food Contact Surfaces after Cleaning and Disinfection in Different Food Industries. J. Food Prot. 2019, 82, 262–275. [Google Scholar] [CrossRef]
- Techer, C.; Jan, S.; Gonnet, F.; Grosset, N.; Gautier, M.; Baron, F. Bacterial diversity on stainless steel surfaces of egg processing companies and potential of selected isolates to spoil liquid whole egg products. J. Appl. Microbiol. 2019, 127, 1501–1510. [Google Scholar] [CrossRef] [PubMed]
- Jahid, I.K.; Ha, S.-D. A review of microbial biofilms of produce: Future challenge to food safety. Food Sci. Biotechnol. 2012, 21, 299–316. [Google Scholar] [CrossRef]
- Flemming, H.-C.; Wuertz, S. Bacteria and archaea on Earth and their abundance in biofilms. Nat. Rev. Microbiol. 2019, 17, 247–260. [Google Scholar] [CrossRef] [PubMed]
- Sauer, K. The genomics and proteomics of biofilm formation. Genome Biol. 2003, 4, 219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kishen, A.; Haapasalo, M. Biofilm models and methods of biofilm assessment. Endod. Top. 2010, 22, 58–78. [Google Scholar] [CrossRef]
- Gobbetti, M.; De Angelis, M.; Di Cagno, R.; Minervini, F.; Limitone, A. Cell–cell communication in food related bacteria. Int. J. Food Microbiol. 2007, 120, 34–45. [Google Scholar] [CrossRef]
- Chia, T.W.R.; Goulter, R.M.; McMeekin, T.; Dykes, G.A.; Fegan, N. Attachment of different Salmonella serovars to materials commonly used in a poultry processing plant. Food Microbiol. 2009, 26, 853–859. [Google Scholar] [CrossRef]
- Chmielewski, R.A.N.; Frank, J.F. Biofilm Formation and Control in Food Processing Facilities. Compr. Rev. Food Sci. Food Saf. 2003, 2, 22–32. [Google Scholar] [CrossRef]
- EHEDG Document No. 43. Hygienic Design of Belt Conveyors for the Food Industry; European Hygienic Enginnering & Design Group: Frankfurt, Germany, 2016.
- Fink, R. Good Hygiene Practices and Their Prevention of Biofilms in the Food Industry; Cambridge Scholars Publishing: Newcastle, UK, 2019. [Google Scholar]
- Partington, E. EHEDG Document No. 32. Materials of Construction for Equipment in Contact with Food; European Hygienic Enginnering & Design Group: Frankfurt, Germany, 2005. [Google Scholar] [CrossRef]
- Cheng, Y.; Feng, G.; Moraru, C.I. Micro- and Nanotopography Sensitive Bacterial Attachment Mechanisms: A Review. Front. Microbiol. 2019, 10, 191. [Google Scholar] [CrossRef] [Green Version]
- Hauser, G.; Curiel, G.J.; Bellin, H.W.; Cnossen, H.J.; Hofmann, J.; Kastelein, J.; Partington, E.; Peltier, Y.; Timperley, A.W. EHEDG Document No. 13. Hygienic Design of Equipment for Open Processing, 2nd ed.; European Hygienic Enginnering & Design Group: Frankfurt, Germany, 2005. [Google Scholar]
- Pérez Ibarreche, M.; Castellano, P.; Vignolo, G. Evaluation of anti-Listeria meat borne Lactobacillus for biofilm formation on selected abiotic surfaces. Meat Sci. 2014, 96, 295–303. [Google Scholar] [CrossRef]
- Schulte, S. Efficiency of Hydrogen Peroxide against Biofilms; University of Duisburg-Essen: Duisburg, Germany, 2004. [Google Scholar]
- Rodríguez-Melcón, C.; Capita, R.; Rodríguez-Jerez, J.J.; Martínez-Suárez, J.V.; Alonso-Calleja, C. Effect of Low Doses of Disinfectants on the Biofilm-Forming Ability of Listeria monocytogenes. Foodborne Pathog. Dis. 2019, 16, 262–268. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Murtey, M.D.; Ramasamy, P. Sample Preparations for Scanning Electron Microscopy–Life. Sciences 2016, 23, 45. [Google Scholar] [CrossRef] [Green Version]
- ISO 25178-2:2012. Geometrical Product Specifications (GPS)—Surface Texture: Areal—Part 2: Terms, Definitions and Surface Texture Parameters; ISO: Geneva, Switzerland, 2021. [Google Scholar]
- ISO 4288:1998. Geometrical Product Specifications (GPS)—Surface Texture: Profile Method—Rules and Procedures for the Assessment of Surface Texture; ISO: Geneva, Switzerland, 1998. [Google Scholar]
- Eifler, M.; Garretson, I.C.; Linke, B.S.; Das, J.; Torner, F.; Seewig, J. Effects of vibratory finishing of 304 stainless steel samples on areal roughness parameters: A correlational analysis for anisotropy parameters. J. Mater. Process Technol. 2019, 273, 116256. [Google Scholar] [CrossRef]
- Van Oss, C.J.; Good, R.J.; Chaudhury, M.K. Additive and nonadditive surface tension components and the interpretation of contact angles. Langmuir 1988, 4, 884–891. [Google Scholar] [CrossRef]
- Gindl, M.; Sinn, G.; Gindl, W.; Reiterer, A.; Tschegg, S. A comparison of different methods to calculate the surface free energy of wood using contact angle measurements. Colloids Surf. A Physicochem. Eng. Asp. 2001, 181, 279–287. [Google Scholar] [CrossRef]
- Bargir, S.; Dunn, S.; Jefferson, B.; Macadam, J.; Parsons, S. The use of contact angle measurements to estimate the adhesion propensity of calcium carbonate to solid substrates in water. Appl. Surf. Sci. 2009, 255, 4873–4879. [Google Scholar] [CrossRef]
- Makovcova, J.; Babak, V.; Kulich, P.; Masek, J.; Slany, M.; Cincarova, L. Dynamics of mono- and dual-species biofilm formation and interactions between Staphylococcus aureus and Gram-negative bacteria. Microb. Biotechnol. 2017, 10, 819–832. [Google Scholar] [CrossRef]
- Nikinmaa, S.; Alapulli, H.; Auvinen, P.; Vaara, M.; Rantala, J.; Kankuri, E.; Sorsa, T.; Meurman, J.; Pätilä, T. Dual-light photodynamic therapy administered daily provides a sustained antibacterial effect on biofilm and prevents Streptococcus mutans adaptation. PLoS ONE 2020, 14, e0232775. [Google Scholar] [CrossRef]
- Galdiero, E.; Di Onofrio, V.; Maione, A.; Gambino, E.; Gesuele, R.; Menale, B.; Ciaravolo, M.; Carraturo, F.; Guida, M. Allium ursinum and Allium oschaninii against Klebsiella pneumoniae and Candida albicans Mono- and Polymicrobic Biofilms in In Vitro Static and Dynamic Models. Microorganisms 2020, 8, 336. [Google Scholar] [CrossRef] [Green Version]
- Granick, M.S.; Paribathan, C.; Shanmugam, M.; Ramasubbu, N. Direct-contact low-frequency ultrasound clearance of biofilm from metallic implant materials. Eplasty 2017, 17, 17. [Google Scholar]
- Zips, A.; Schaule, G.; Flemming, H. Ultrasound as a means of detaching biofilms. Biofouling 1990, 2, 323–333. [Google Scholar] [CrossRef]
- Baumann, A.R.; Martin, S.E.; Feng, H. Removal of Listeria monocytogenes Biofilms from Stainless Steel by Use of Ultrasound and Ozone. J. Food Prot. 2009, 72, 1306–1309. [Google Scholar] [CrossRef]
- Badalyan, G.; Díaz, C.; Bücking, M.; Lipski, A. Novel sensor platform for rapid detection and quantification of coliforms on food contact surfaces. J. Microbiol. Methods 2018, 153, 74–83. [Google Scholar] [CrossRef]
- Wang, H.; Wu, N.; Jiang, Y.; Ye, K.; Xu, X.; Zhou, G. Response of long-term acid stress to biofilm formation of meat-related Salmonella Enteritidis. Food Control 2016, 69, 214–220. [Google Scholar] [CrossRef]
- Bjerkan, G.; Witsø, E.; Bergh, K. Sonication is superior to scraping for retrieval of bacteria in biofilm on titanium and steel surfaces in vitro. Acta Orthop. 2009, 80, 245–250. [Google Scholar] [CrossRef] [Green Version]
- Kragh, K.N.; Alhede, M.; Kvich, L.; Bjarnsholt, T. Into the well—A close look at the complex structures of a microtiter biofilm and the crystal violet assay. Biofilm 2019, 1, 100006. [Google Scholar] [CrossRef]
- Planchon, S.; Gaillardmartinie, B.; Leroy, S.; Bellonfontaine, M.; Fadda, S.; Talon, R. Surface properties and behaviour on abiotic surfaces of Staphylococcus carnosus, a genetically homogeneous species. Food Microbiol. 2007, 24, 44–51. [Google Scholar] [CrossRef] [PubMed]
- Iñiguez-Moreno, M.; Avila-Novoa, M.G.; Gutiérrez-Lomelí, M. Resistance of pathogenic and spoilage microorganisms to disinfectants in the presence of organic matter and their residual effect on stainless steel and polypropylene. J. Glob. Antimicrob. Resist. 2018, 14, 197–201. [Google Scholar] [CrossRef] [PubMed]
- Iñiguez-Moreno, M.; Gutiérrez-Lomelí, M.; Avila-Novoa, M.G. Kinetics of biofilm formation by pathogenic and spoilage microorganisms under conditions that mimic the poultry, meat, and egg processing industries. Int. J. Food Microbiol. 2019, 303, 32–41. [Google Scholar] [CrossRef]
- Heilmann, C.; Hussain, M.; Peters, G.; Götz, F. Evidence for autolysin-mediated primary attachment of Staphylococcus epidermidis to a polystyrene surface. Mol. Microbiol. 1997, 24, 1013–1024. [Google Scholar] [CrossRef]
- Cui, B.; Deighton, M.A.; Smooker, P.M.; Rouch, D.A. Effects of erythromycin on the phenotypic and genotypic biofilm expression in two clinical Staphylococcus capitis subspecies and a functional analysis of Ica proteins in S. capitis. J. Med. Microbiol. 2015, 65, 257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heilmann, C. Identification and characterization of a novel autolysin (Aae) with adhesive properties from Staphylococcus epidermidis. Microbiology 2003, 149, 2769–2778. [Google Scholar] [CrossRef] [Green Version]
- Vithanage, N.R.; Dissanayake, M.; Bolge, G.; Palombo, E.A.; Yeager, T.R.; Datta, N. Biodiversity of culturable psychrotrophic microbiota in raw milk attributable to refrigeration conditions, seasonality and their spoilage potential. Int. Dairy J. 2016, 57, 80–90. [Google Scholar] [CrossRef]
- Ripolles-Avila, C.; Hascoët, A.S.; Guerrero-Navarro, A.E.; Rodríguez-Jerez, J.J. Establishment of incubation conditions to optimize the in vitro formation of mature Listeria monocytogenes biofilms on food-contact surfaces. Food Control 2018, 92, 240–248. [Google Scholar] [CrossRef]
- Rice, K.C.; Bayles, K.W. Death’s toolbox: Examining the molecular components of bacterial programmed cell death. Mol. Microbiol. 2003, 50, 729–738. [Google Scholar] [CrossRef]
- Ripolles-Avila, C.; García-Hernández, N.; Cervantes-Huamán, B.H.; Mazaheri, T.; Rodríguez-Jerez, J.J. Quantitative and Compositional Study of Monospecies Biofilms of Spoilage Microorganisms in the Meat Industry and Their Interaction in the Development of Multispecies Biofilms. Microorganisms 2019, 7, 655. [Google Scholar] [CrossRef] [Green Version]
- Kerstens, M.; Boulet, G.; Van Kerckhoven, M.; Clais, S.; Lanckacker, E.; Delputte, P.; Maes, L.; Cos, P. A flow cytometric approach to quantify biofilms. Folia Microbiol. 2015, 60, 335–342. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Melcón, C.; Riesco-Peláez, F.; Carballo, J.; García-Fernández, C.; Capita, R.; Alonso-Calleja, C. Structure and viability of 24- and 72-h-old biofilms formed by four pathogenic bacteria on polystyrene and glass contact surfaces. Food Microbiol. 2018, 76, 513–517. [Google Scholar] [CrossRef]
- Stiefel, P.; Schmidt-Emrich, S.; Maniura-Weber, K.; Ren, Q. Critical aspects of using bacterial cell viability assays with the fluorophores SYTO9 and propidium iodide. BMC Microbiol. 2015, 15, 36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.; Hoe, Y.W.; Zheng, Q.; Chung, H.-J.; Yuk, H.-G. Biofilm formation by Salmonella Enteritidis in a simulated liquid egg processing environment and its sensitivity to chlorine and hot water treatment. Food Control 2016, 73, 595–600. [Google Scholar] [CrossRef]
- Jun, W.; Kim, M.S.; Cho, B.-K.; Millner, P.D.; Chao, K.; Chan, D.E. Microbial biofilm detection on food contact surfaces by macro-scale fluorescence imaging. J. Food Eng. 2010, 99, 314–322. [Google Scholar] [CrossRef]
- Haider, S.A.; Cameron, A.; Siva, P.; Lui, D.; Shafiee, M.J.; Boroomand, A.; Haider, N.; Wong, A. Fluorescence microscopy image noise reduction using a stochastically-connected random field model. Sci. Rep. 2016, 6, 20640. [Google Scholar] [CrossRef] [Green Version]
- El Abed, S.; Ibnsouda, S.K.; Latrache, H.; Hamadi, F. Scanning electron microscopy (SEM) and environmental SEM: Suitable tools for study of adhesion stage and biofilm formation. In Scanning Electron Microscopy; Kazmiruk, V., Ed.; Intechopen: London, UK, 2012. [Google Scholar] [CrossRef] [Green Version]
- Liu, F.; Jin, P.; Sun, Z.; Du, L.; Wang, D.; Zhao, T.; Doyle, M.P. Carvacrol oil inhibits biofilm formation and exopolysaccharide production of Enterobacter cloacae. Food Control 2021, 119, 107473. [Google Scholar] [CrossRef]
- Sadekuzzaman, M.; Yang, S.; Rahaman Mizan, M.F.; Kim, H.-S.; Ha, S.-D. Effectiveness of a phage cocktail as a biocontrol agent against L. monocytogens Biofilms. Food Control 2016, 16, 56. [Google Scholar] [CrossRef]
- Zhou, W.; Apkarian, R.; Wang, Z.L.; Joy, D. Fundamentals of Scanning Electron Microscopy (SEM). In Scanning Microscopy for Nanotechnology: Techniques and Applications; Zhou, W., Wang, Z.L., Eds.; Springer: New York, NY, USA, 2007; pp. 1–40. [Google Scholar] [CrossRef]
- Hofmann, J.; Akesson, S.; Curiel, G.; Wouters, P.; Timperley, A. EHEDG Document No. 8. Hygienic Design Principles, 3rd ed.; European Hygienic Enginnering & Design Group: Frankfurt, Germany, 2018. [Google Scholar]
- Jones, C.R.; Adams, M.R.; Zhdan, P.A.; Chamberlain, A.H.L. The role of surface physicochemical properties in determining the distribution of the autochthonous microflora in mineral water bottles. J. Appl. Microbiol. 1999, 86, 917–927. [Google Scholar] [CrossRef]
- Gadelmawla, E.S.; Koura, M.M.; Maksoud, T.M.A.; Elewa, I.M.; Soliman, H.H. Roughness parameters. J. Mater. Process Technol. 2002, 123, 133–145. [Google Scholar] [CrossRef]
- Leach, R. Characterisation of Areal Surface Texture; Springer Sience & Business Media: Berlin/Heidelberg, Germany, 2013. [Google Scholar]
- Hsu, L.C.; Fang, J.; Borca-Tasciuc, D.A.; Worobo, R.W.; Moraru, C.I. Effect of Micro- and Nanoscale Topography on the Adhesion of Bacterial Cells to Solid Surfaces. Appl. Environ. Microbiol. 2013, 79, 2703–2712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friedlander, R.S.; Vlamakis, H.; Kim, P.; Khan, M.; Kolter, R.; Aizenberg, J. Bacterial flagella explore microscale hummocks and hollows to increase adhesion. Proc. Natl. Acad. Sci. USA 2013, 110, 5624–5629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hochbaum, A.I.; Aizenberg, J. Bacteria Pattern Spontaneously on Periodic Nanostructure Arrays. Nano Lett. 2010, 10, 3717–3721. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, P.; Lima, J.; Azeredo, J.; Oliveira, R. Adhesion of Listeria monocytogenes to materials commonly found in domestic kitchens. Int. J. Food Sci. Technol. 2008, 43, 1239–1244. [Google Scholar] [CrossRef] [Green Version]
- Bezek, K.; Nipič, D.; Torkar, K.G.; Oder, M.; Dražić, G.; Abram, A.; Žibert, J.; Raspor, P.; Bohinc, K. Biofouling of stainless steel surfaces by four common pathogens: The effects of glucose concentration, temperature and surface roughness. Biofouling 2019, 35, 273–283. [Google Scholar] [CrossRef] [PubMed]
- Ammar, Y.; Swailes, D.; Bridgens, B.; Chen, J. Influence of surface roughness on the initial formation of biofilm. Surf. Coat. Technol. 2015, 284, 410–416. [Google Scholar] [CrossRef] [Green Version]
- Dürr, H. Influence of Surface Roughness and Wettability of Stainless Steel on Soil Adhesion, Cleanability and Microbial Inactivation. Food Bioprod. Process. 2007, 85, 49–56. [Google Scholar] [CrossRef]
- Awad, T.S.; Asker, D.; Hatton, B.D. Food-Safe Modification of Stainless Steel Food-Processing Surfaces to Reduce Bacterial Biofilms. ACS Appl. Mater. Interfaces 2018, 10, 22902–22912. [Google Scholar] [CrossRef] [PubMed]
- Park, S.-J.; Seo, M.-K. Chapter 2—Solid-Gas Interaction. In Interface Science and Technology; Park, S.-J., Seo, M.-K., Eds.; Elsevier: Amsterdam, The Netherlands, 2011; Volume 18, pp. 59–145. [Google Scholar]
- Silva, S.; Teixeira, P.; Oliveira, R.; Azeredo, J. Adhesion to and Viability of Listeria monocytogenes on Food Contact Surfaces. J. Food Prot. 2008, 71, 1379–1385. [Google Scholar] [CrossRef]
- Vogler, E.A. Structure and reactivity of water at biomaterial surfaces. Adv. Colloid Interface Sci. 1998, 74, 69–117. [Google Scholar] [CrossRef]
- Hamadi, F.; Asserne, F.; Elabed, S.; Bensouda, S.; Mabrouki, M.; Latrache, H. Adhesion of Staphylococcus aureus on stainless steel treated with three types of milk. Food Control 2014, 38, 104–108. [Google Scholar] [CrossRef]
- Azelmad, K.; Hamadi, F.; Mimouni, R.; Amzil, K.; Latrache, H.; Mabrouki, M.; El Boulani, A. Adhesion of Staphylococcus aureus and Staphylococcus xylosus to materials commonly found in catering and domestic kitchens. Food Control 2017, 73, 156–163. [Google Scholar] [CrossRef]
- Zhao, Q.; Wang, C.; Liu, Y.; Wang, S. Bacterial adhesion on the metal-polymer composite coatings. Int. J. Adhes. Adhes. 2007, 27, 85–91. [Google Scholar] [CrossRef]
- Pereni, C.I.; Zhao, Q.; Liu, Y.; Abel, E. Surface free energy effect on bacterial retention. Colloids Surf. B Biointerfaces 2006, 48, 143–147. [Google Scholar] [CrossRef]
- Hood, S.K.; Zottola, E.A. Adherence to stainless steel by foodborne microorganisms during growth in model food systems. Int. J. Food Microbiol. 1997, 37, 145–153. [Google Scholar] [CrossRef]
- Giaouris, E.; Chapot-Chartier, M.-P.; Briandet, R. Surface physicochemical analysis of natural Lactococcus lactis strains reveals the existence of hydrophobic and low charged strains with altered adhesive properties. Int. J. Food Microbiol. 2009, 131, 2–9. [Google Scholar] [CrossRef] [PubMed]
- Da Silva Meira, Q.G.; De Medeiros Barbosa, I.; Alves Aguiar Athayde, A.J.; De Siqueira-Júnior, J.P.; De Souza, E.L. Influence of temperature and surface kind on biofilm formation by Staphylococcus aureus from food-contact surfaces and sensitivity to sanitizers. Food Control 2012, 25, 469–475. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Material | Line Roughness [µm] b | Surface Roughness [µm] c | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Ra | Rz | Rq | Rv | Rp | RSm | Sa | Sz | Sq | Sv | Sp | |
| SS 320 | 0.5 ± 0.0 | 2.5 ± 0.1 | 0.7 ± 0.0 | 1.3 ± 0.1 | 1.5 ± 0.1 | 10.2 ± 0.5 | 0.7 | 7.5 | 0.6 | 2.7 | 2.9 |
| SS 240 | 1.0 ± 0.0 | 4.2 ± 0.1 | 1.0 ± 0.1 | 1.7 ± 0.1 | 2.1 ± 0.1 | 10.7 ± 1.3 | 1.1 | 17.6 | 1.0 | 9.7 | 5.1 |
| PTFE | 0.2 ± 0.0 | 0.9 ± 0.2 | 0.3 ± 0.1 | 0.5 ± 0.1 | 0.4 ± 0.1 | 9.7 ± 2.2 | 0.5 | 15.1 | 0.5 | 6.9 | 3.2 |
| Material | Contact Angle (°) b | Surface Free Energy (mJ m−2) c | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| θTSB | θMB | θW | θD | θF | ||||||
| SS 320 | 54.6 ± 1.3 | 61.4 ± 3.1 | 65.9 ± 11.7 | 36.9 ± 5.5 | 52.7 ± 6.3 | 41.3 ± 0.4 | 41.0 ± 0.2 | 0.3 ± 0.2 | 0.0 ± 0.0 | 17.1 ± 0.3 |
| SS 240 | 58.9 ± 3.2 | 60.1 ± 4.7 | 85.6 ± 7.9 | 41.4 ± 1.1 | 55.7 ± 8.2 | 40.7 ± 0.2 | 39.0 ± 0.1 | 1.8 ± 0.1 | 0.5 ± 0.0 | 1.7 ± 0.1 |
| PTFE | 76.3 ± 7.9 | 94.5 ± 4.5 | 94.5 ± 8.0 | 72.8 ± 10.8 | 66.5 ± 5.8 | 23.5 ± 0.3 | 21.2 ± 0.1 | 2.4 ± 0.1 | 2.9 ± 0.1 | 0.5 ± 0.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zand, E.; Pfanner, H.; Domig, K.J.; Sinn, G.; Zunabovic-Pichler, M.; Jaeger, H. Biofilm-Forming Ability of Microbacterium lacticum and Staphylococcus capitis Considering Physicochemical and Topographical Surface Properties. Foods 2021, 10, 611. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10030611
Zand E, Pfanner H, Domig KJ, Sinn G, Zunabovic-Pichler M, Jaeger H. Biofilm-Forming Ability of Microbacterium lacticum and Staphylococcus capitis Considering Physicochemical and Topographical Surface Properties. Foods. 2021; 10(3):611. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10030611
Chicago/Turabian StyleZand, Elena, Hedwig Pfanner, Konrad J. Domig, Gerhard Sinn, Marija Zunabovic-Pichler, and Henry Jaeger. 2021. "Biofilm-Forming Ability of Microbacterium lacticum and Staphylococcus capitis Considering Physicochemical and Topographical Surface Properties" Foods 10, no. 3: 611. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10030611