
Prevalence of Salmonella in Free-Range Pigs: Risk Factors and Intestinal Microbiota Composition

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design
2.2. Isolation of Salmonella
2.3. Antimicrobial Susceptibility Testing
2.4. Epidemiological Study and Risk Factors
2.5. Microbiota Analyses of Cecum Content
2.6. Intestinal Content DNA Extraction, PCR Amplification and Microbiota Analyses
2.7. Quality Control, Operational Taxonomic Unit (OTUs), Diversity and Composition Analyses
3. Results
3.1. Prevalence of Salmonella in the Extensive System
3.2. Serotyping and Antimicrobial Susceptibility
3.3. Risk Factors Associated with the Presence of Salmonella in IC Samples
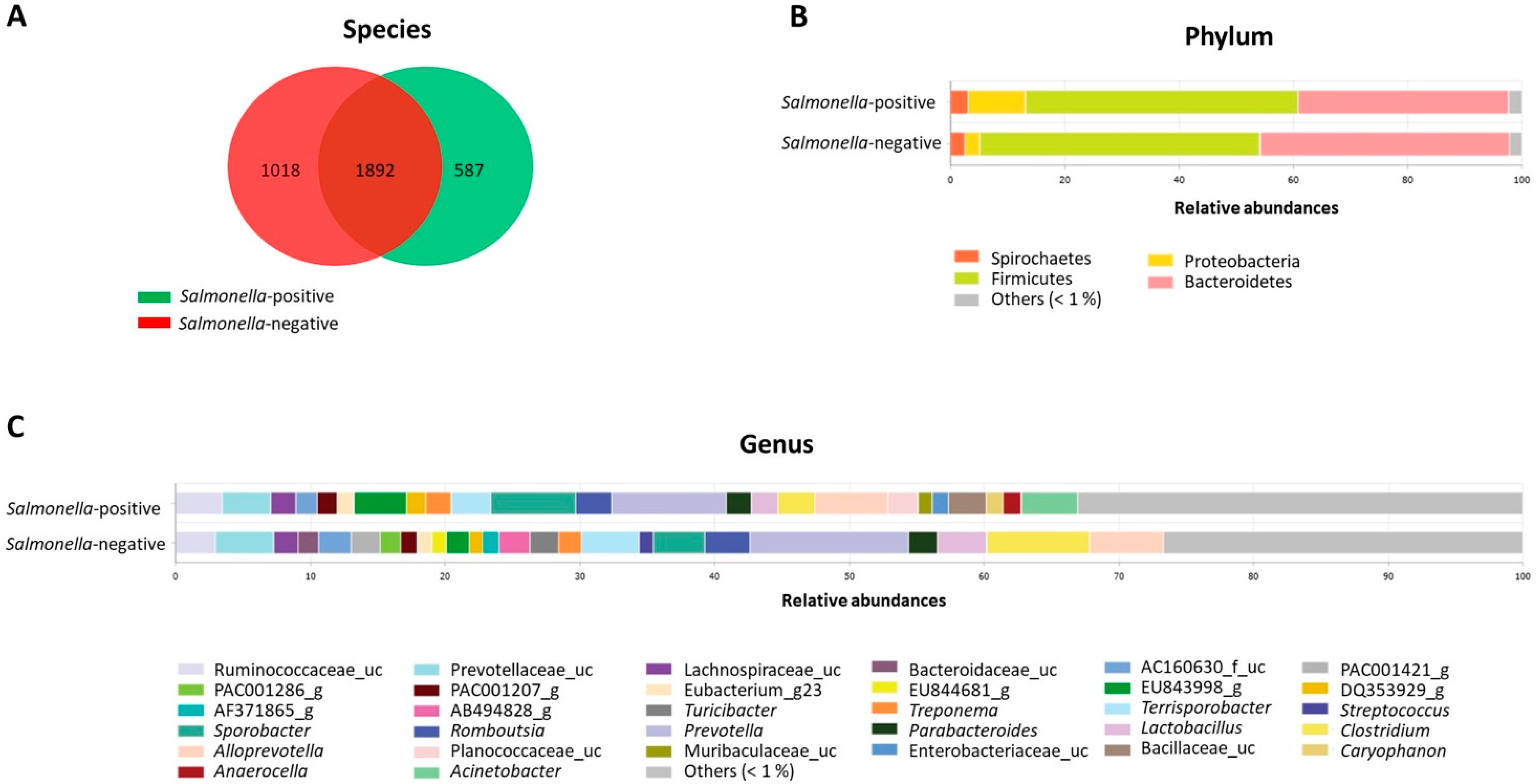
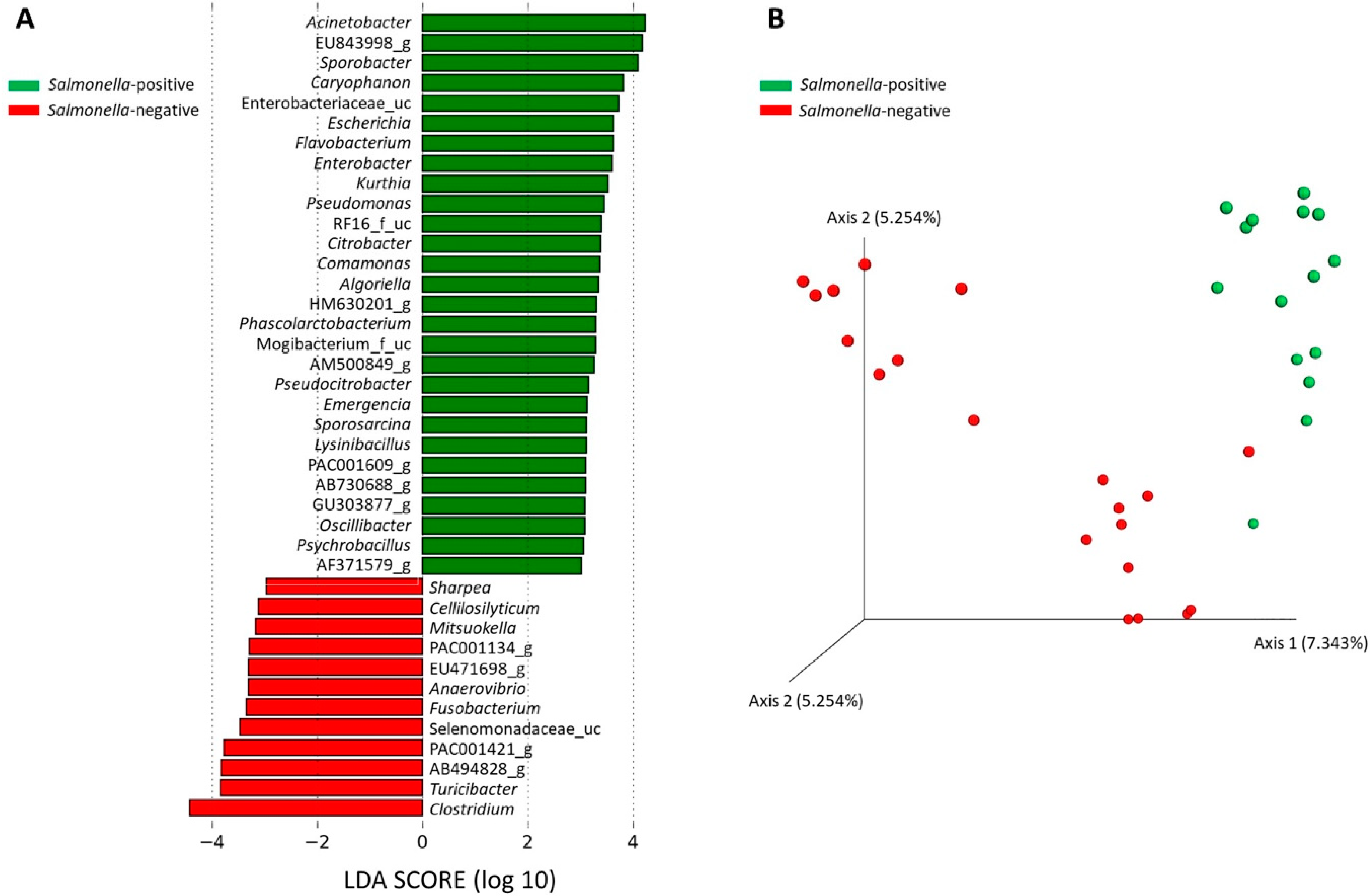
3.4. Microbiota Analysis in Two Pig Populations
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- European Food Safety Authority; European Centre for Disease Prevention and Control. The European Union One Health 2019 Zoonoses Report. EFSA J. 2021, 19, e06406. [Google Scholar] [CrossRef]
- Pires, S.M.; de Knegt, L.; Hald, T. Estimation of the relative contribution of different food and animal sources to human Salmonella infections in the European Union. In Scientific/Technical Report Submitted to EFSA; National Food Institute, Technical University of Denmark: Søborg, Denmark, 2011. [Google Scholar]
- EFSA-ECDC. The European Union One Health 2018 Zoonoses Report. EFSA J. 2019, 17, 5926. [Google Scholar] [CrossRef] [Green Version]
- MAPAMA. Caracterización del Sector Porcino Español Año 2019. 2020. Available online: https://www.mapa.gob.es/es/ganaderia/temas/produccion-y-mercados-ganaderos/indicadoreseconomicoscarnedecerdo2019_tcm30-379728.pdf (accessed on 17 June 2021).
- Thorslund, C.A.; Aaslyng, M.D.; Lassen, J. Perceived importance and responsibility for market-driven pig welfare: Literature review. Meat Sci. 2017, 125, 37–45. [Google Scholar] [CrossRef]
- Backhed, F.; Ley, R.E.; Sonnenburg, J.L.; Peterson, D.A.; Gordon, J.I. Host-bacterial mutualism in the human intestine. Science 2005, 307, 1915–1920. [Google Scholar] [CrossRef] [Green Version]
- Isaacson, R.; Kim, H.B. The intestinal microbiome of the pig. Anim. Health Res. Rev. 2012, 13, 100–109. [Google Scholar] [CrossRef]
- Bearson, S.M.; Allen, H.K.; Bearson, B.L.; Looft, T.; Brunelle, B.W.; Kich, J.D.; Tuggle, C.K.; Bayles, D.O.; Alt, D.; Levine, U.Y.; et al. Profiling the gastrointestinal microbiota in response to Salmonella: Low versus high Salmonella shedding in the natural porcine host. Infect. Genet. Evol. 2013, 16, 330–340. [Google Scholar] [CrossRef] [PubMed]
- Borewicz, K.A.; Kim, H.B.; Singer, R.S.; Gebhart, C.J.; Sreevatsan, S.; Johnson, T.; Isaacson, R.E. Changes in the porcine intestinal microbiome in response to infection with Salmonella enterica and Lawsonia intracellularis. PLoS ONE 2015, 10, e0139106. [Google Scholar] [CrossRef]
- ISO. International Organisation for Standardisation. ISO 6579:2002/Amd. 1:2007. Microbiology of food and animal feeding stuffs. Horizontal method for the detection of Salmonella spp. Amendment 1 Annex D: Detection of Salmonella spp. In Animal Faeces and in Samples from the Primary Production Stage; ISO: Geneve, Switzerland, 2007. [Google Scholar]
- Garrido, V.; Sanchez, S.; San Roman, B.; Fraile, L.; Migura-Garcia, L.; Grillo, M.J. Salmonella Infection in Mesenteric Lymph Nodes of Breeding Sows. Foodborne Pathog. Dis. 2020, 17, 411–417. [Google Scholar] [CrossRef] [PubMed]
- San Roman, B.; Garrido, V.; Sanchez, S.; Martinez-Ballesteros, I.; Garaizar, J.; Mainar-Jaime, R.C.; Migura-Garcia, L.; Grillo, M.J. Relationship between Salmonella infection, shedding and serology in fattening pigs in low-moderate prevalence areas. Zoonoses Public Health 2018, 65, 481–489. [Google Scholar] [CrossRef] [PubMed]
- Garrido, V.; Sanchez, S.; San Roman, B.; Zabalza-Barangua, A.; Diaz-Tendero, Y.; de Frutos, C.; Mainar-Jaime, R.C.; Grillo, M.J. Simultaneous infections by different Salmonella strains in mesenteric lymph nodes of finishing pigs. BMC Vet. Res. 2014, 10, 59. [Google Scholar] [CrossRef] [Green Version]
- Grimont, P.A.; Weill, F.X. Antigenic formulae of the Salmonella Serovars, 9th ed.; World Health Organization (WHO) Collaborating Centre for Reference and Research on Salmonella; Institut Pasteur: Paris, France, 2007. [Google Scholar]
- Mainar-Jaime, R.C.; Atashparvar, N.; Chirino-Trejo, M. Estimation of the diagnostic accuracy of the invA-gene-based PCR technique and a bacteriological culture for the detection of Salmonella spp. in caecal content from slaughtered pigs using Bayesian analysis. Zoonoses Public Health 2008, 55, 112–118. [Google Scholar] [CrossRef] [PubMed]
- Landis, J.R.; Koch, G.G. The measurement of observer agreement for categorical data. Biometrics 1977, 33, 159–174. [Google Scholar] [CrossRef] [Green Version]
- EU. Commission Decision of 12 June 2007 on a Harmonised monitoring of antimicrobial resistance in Salmonella in poultry and pigs. 2007/407/EC. Official Journal of the European Union. Off. J. Eur. Union 2007, 50, L153. [Google Scholar]
- CLSI. Performance Standards for Antimicrobial Susceptibility Testing. Standard M100S; CLSI: Wayne, PA, USA, 2016. [Google Scholar]
- Yoon, S.H.; Ha, S.M.; Kwon, S.; Lim, J.; Kim, Y.; Seo, H.; Chun, J. Introducing EzBioCloud: A taxonomically united database of 16S rRNA gene sequences and whole-genome assemblies. Int. J. Syst. Evol. Microbiol. 2017, 67, 1613–1617. [Google Scholar] [CrossRef]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef] [Green Version]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahe, F. VSEARCH: A versatile open source tool for metagenomics. PeerJ 2016, 4, e2584. [Google Scholar] [CrossRef] [PubMed]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Pena, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [Green Version]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef] [Green Version]
- Thakur, S.; Tadesse, D.A.; Morrow, M.; Gebreyes, W.A. Occurrence of multidrug resistant Salmonella in antimicrobial-free (ABF) swine production systems. Vet. Microbiol. 2007, 125, 362–367. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Laguna, J.; Hernandez, M.; Creus, E.; Echeita, A.; Otal, J.; Herrera-Leon, S.; Astorga, R.J. Prevalence and antimicrobial susceptibility of Salmonella infections in free-range pigs. Vet. J. 2011, 190, 176–178. [Google Scholar] [CrossRef] [PubMed]
- Jensen, A.N.; Dalsgaard, A.; Stockmarr, A.; Nielsen, E.M.; Baggesen, D.L. Survival and transmission of Salmonella enterica serovar typhimurium in an outdoor organic pig farming environment. Appl. Environ. Microbiol. 2006, 72, 1833–1842. [Google Scholar] [CrossRef] [Green Version]
- Smith, R.P.; Andres, V.; Dormer, L.; Gosling, R.; Oastler, C.; Davies, R.H. Study of the impact on Salmonella of moving outdoor pigs to fresh land. Epidemiol. Infect. 2017, 145, 1983–1992. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- EFSA. Report of the task force on zoonoses data collection on the analysis of the baseline survey on the prevalence of Salmonella in slaughter pigs. Part A. EFSA J. 2008, 135, 1–111. [Google Scholar]
- Vico, J.P.; Rol, I.; Garrido, V.; San Román, B.; Grilló, M.J.; Mainar-Jaime, R.C. Salmonellosis in finishing pigs in Spain: Prevalence, antimicrobial agent susceptibilities, and risk factor analysis. J. Food Prot. 2011, 74, 1070–1078. [Google Scholar] [CrossRef] [Green Version]
- Echeita, M.A.; Herrera, S.; Usera, M.A. Atypical, fljB-negative Salmonella enterica subsp. enterica strain of serovar 4,5,12:i:- appears to be a monophasic variant of serovar Typhimurium. J. Clin. Microbiol. 2001, 39, 2981–2983. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biswas, S.; Li, Y.; Elbediwi, M.; Yue, M. Emergence and Dissemination of mcr-Carrying Clinically Relevant Salmonella Typhimurium Monophasic Clone ST34. Microorganisms 2019, 7, 298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- EFSA. Scientific Report on the analysis of the baseline survey on the prevalence of Salmonella in slaughter pigs, in the EU, 2006-2007. Part B: Factors associated with Salmonella infection in lymph nodes, Salmonella surface contamination of carcasses, and the distribution of Salmonella serovars. EFSA J. 2008, 206, 1–111. [Google Scholar]
- Petrovska, L.; Mather, A.E.; AbuOun, M.; Branchu, P.; Harris, S.R.; Connor, T.; Hopkins, K.L.; Underwood, A.; Lettini, A.A.; Page, A.; et al. Microevolution of Monophasic Salmonella Typhimurium during Epidemic, United Kingdom, 2005–2010. Emerg. Infect. Dis. 2016, 22, 617–624. [Google Scholar] [CrossRef] [Green Version]
- Andres-Barranco, S.; Vico, J.P.; Garrido, V.; Samper, S.; Herrera-Leon, S.; de Frutos, C.; Mainar-Jaime, R.C. Role of wild bird and rodents in the epidemiology of subclinical salmonellosis in finishing pigs. Foodborne Pathog. Dis. 2014, 11, 689–697. [Google Scholar] [CrossRef]
- Andres, S.; Vico, J.P.; Garrido, V.; Grillo, M.J.; Samper, S.; Gavin, P.; Herrera-Leon, S.; Mainar-Jaime, R.C. Epidemiology of subclinical salmonellosis in wild birds from an area of high prevalence of pig salmonellosis: Phenotypic and genetic profiles of Salmonella isolates. Zoonoses Public Health 2013, 60, 355–365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rivera-Chavez, F.; Baumler, A.J. The Pyromaniac Inside You: Salmonella Metabolism in the Host Gut. Annu. Rev. Microbiol. 2015, 69, 31–48. [Google Scholar] [CrossRef] [Green Version]
- Rivera-Chavez, F.; Zhang, L.F.; Faber, F.; Lopez, C.A.; Byndloss, M.X.; Olsan, E.E.; Xu, G.; Velazquez, E.M.; Lebrilla, C.B.; Winter, S.E.; et al. Depletion of Butyrate-producing clostridia from the gut microbiota drives an aerobic luminal expansion of Salmonella. Cell Host Microbe 2016, 19, 443–454. [Google Scholar] [CrossRef] [Green Version]
- Chu, B.; Zhu, Y.; Su, J.; Xia, B.; Zou, Y.; Nie, J.; Zhang, W.; Wang, J. Butyrate-mediated autophagy inhibition limits cytosolic Salmonella Infantis replication in the colon of pigs treated with a mixture of Lactobacillus and Bacillus. Vet. Res. 2020, 51, 99. [Google Scholar] [CrossRef]
- Lawhon, S.D.; Maurer, R.; Suyemoto, M.; Altier, C. Intestinal short-chain fatty acids alter Salmonella Typhimurium invasion gene expression and virulence through BarA/SirA. Mol. Microbiol. 2002, 46, 1451–1464. [Google Scholar] [CrossRef]
- Nishitsuji, K.; Xiao, J.; Nagatomo, R.; Umemoto, H.; Morimoto, Y.; Akatsu, H.; Inoue, K.; Tsuneyama, K. Analysis of the gut microbiome and plasma short-chain fatty acid profiles in a spontaneous mouse model of metabolic syndrome. Sci. Rep. 2017, 7, 15876. [Google Scholar] [CrossRef]
- Hai, D.; Lu, Z.; Huang, X.; Lv, F.; Bie, X. In Vitro Screening of Chicken-Derived Lactobacillus Strains that Effectively Inhibit Salmonella Colonization and Adhesion. Foods 2021, 10, 569. [Google Scholar] [CrossRef] [PubMed]
- Jones, B.D.; Ghori, N.; Falkow, S. Salmonella Typhimurium initiates murine infection by penetrating and destroying the specialized epithelial M cells of the Peyer’s patches. J. Exp. Med. 1994, 180, 15–23. [Google Scholar] [CrossRef] [Green Version]
- GrechMora, I.; Fardeau, M.L.; Patel, B.K.C.; Ollivier, B.; Rimbault, A.; Prensier, G.; Garcia, J.L.; GarnierSillam, E. Isolation and characterization of Sporobacter termitidis gen nov sp nov, from the digestive tract of the wood-feeding termite Nasutitermes lujae. Int. J. Syst. Bacteriol. 1996, 46, 512–518. [Google Scholar] [CrossRef] [Green Version]
- Shin, N.R.; Whon, T.W.; Bae, J.W. Proteobacteria: Microbial signature of dysbiosis in gut microbiota. Trends Biotechnol. 2015, 33, 496–503. [Google Scholar] [CrossRef]
- Litvak, Y.; Byndloss, M.X.; Tsolis, R.M.; Baumler, A.J. Dysbiotic Proteobacteria expansion: A microbial signature of epithelial dysfunction. Curr. Opin. Microbiol. 2017, 39, 1–6. [Google Scholar] [CrossRef]
- Johnson, J.S.; Spakowicz, D.J.; Hong, B.Y.; Petersen, L.M.; Demkowicz, P.; Chen, L.; Leopold, S.R.; Hanson, B.M.; Agresta, H.O.; Gerstein, M.; et al. Evaluation of 16S rRNA gene sequencing for species and strain-level microbiome analysis. Nat. Commun. 2019, 10, 5029. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Salmonella spp. Isolation | Intestinal Content |
|---|---|
| No. (%; CI95) 1 positive pigs 2/total pigs | 58/180 (32.2%; 25.8–39.3) |
| No. (%; CI95) 1 positive farms 2/total farms | 10/12 (83.3%; 55.2–95.3) |
| No. (%; CI95) 1 positive pigs 2/pigs in positive farms | 58/150 (38.7; 31.2–46.6) |
| No. (%) farms above 20% prevalence/total farms | 8/12 (66.7%; 39.1–86.2) |
| Farm Code | No. (%) Samples Positive/Total | Serovars (No. of Strains) | AR Profiles 1 (No. of Strains) |
|---|---|---|---|
| 1 | 13/15 (86.7%) | 4,5,12:i:- (13) | SSu (5); ASSuT (4); ASSu (2); ACSSuT (2) |
| 2 | 8/15 (53.3) | 4,5,12:i:- (8) | ACS (3); CS (2); ACSSu (1); AS (1); ACSSuT (1) |
| 3 | 7/15 (46.7%) | 4,5,12:i:- (7) | SSu (4); ASSuT (3) |
| 4 | 7/15 (46.7%) | 4,5,12:i:- (7) | ASSu (4); ASSuT (2); SSu (1) |
| 5 | 6/15 (40%) | 4,5,12:i:- (5); diarizonae (1) | ASSuT (3); SSu (1); A (1) Susceptible (1) |
| 6 | 5/15 (33.3%) | Bovismorbificans (4); Altona (1) | ACSSuT (2); CSSuT (2); S (1) |
| 7 | 4/15 (26.7%) | Bovismorbificans (2); Meleagridis (2); | CSSuT (2); Susceptible (2) |
| 8 | 4/15 (26.7%) | 4,5,12:i:- (2); Amsterdam (1); Altona (1) | ASSuT (2); Susceptible (2) |
| 9 | 3/15 (20%) | 4,5,12:i:- (3) | ASSu (2); Susceptible (1) |
| 10 | 1/15 (6.7%) | 4,5,12:i:- (1) | ASSuT (1) |
| 11 | 0/15 (0%) | N.A. | N.A. |
| 12 | 0/15 (0%) | N.A. | N.A. |
| Total | 58/180 (32.2%) | 6 serovars (58) | 11 AR profiles |
| Risk Factors | Statistical Analysis (p Values) | |||
|---|---|---|---|---|
| Univariable | Multivariable | |||
| Prevalence | Low/High | Prevalence | Low/High | |
| (i) Large farms (>100 animals) | 0.02 | 0.06 | 0.11 | 0.19 |
| (ii) Feed with A brand | 0.13 | 0.02 | 0.29 | 0.008 |
| (iii) Extra feed with grass | 0.38 | 0.02 | N.A. | N.A. |
| (iv) Neither cleaning nor disinfection of silos | 0.04 | 0.02 | N.A. | N.A. |
| Parameter | OTU Richness and Alpha Diversity Indexes Median and (IQR) a Values | ||
|---|---|---|---|
| Salmonella-Positive Pigs | Salmonella-Negative Pigs | p Values | |
| OTU number | 3132 (583) | 3047 (714.73) | 0.271 |
| Chao1 | 3156.06 (581.84) | 3085.74 (725.19) | 0.301 |
| ACE b | 3227.75 (580.90) | 3166.53 (725.19) | 0.301 |
| Shannon | 6.44 (0.64) | 6.04 (0.80) | 0.072 |
| Simpson | 0.01 (0) | 0.01 (0.01) | 0.062 |
| Phylogenetic diversity | 1366 (80) | 1288 (119.50) | 0.044 c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Garrido, V.; Migura-García, L.; Gaitán, I.; Arrieta-Gisasola, A.; Martínez-Ballesteros, I.; Fraile, L.; Grilló, M.J. Prevalence of Salmonella in Free-Range Pigs: Risk Factors and Intestinal Microbiota Composition. Foods 2021, 10, 1410. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10061410
Garrido V, Migura-García L, Gaitán I, Arrieta-Gisasola A, Martínez-Ballesteros I, Fraile L, Grilló MJ. Prevalence of Salmonella in Free-Range Pigs: Risk Factors and Intestinal Microbiota Composition. Foods. 2021; 10(6):1410. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10061410
Chicago/Turabian StyleGarrido, Victoria, Lourdes Migura-García, Inés Gaitán, Ainhoa Arrieta-Gisasola, Ilargi Martínez-Ballesteros, Lorenzo Fraile, and María Jesús Grilló. 2021. "Prevalence of Salmonella in Free-Range Pigs: Risk Factors and Intestinal Microbiota Composition" Foods 10, no. 6: 1410. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10061410