
Identification of the Specific Spoilage Organism in Farmed Sturgeon (Acipenser baerii) Fillets and Its Associated Quality and Flavour Change during Ice Storage


Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation and Identification of Strains
2.1.1. Total Bacterial DNA Extraction and 16S rDNA V3 Variable Region Amplification
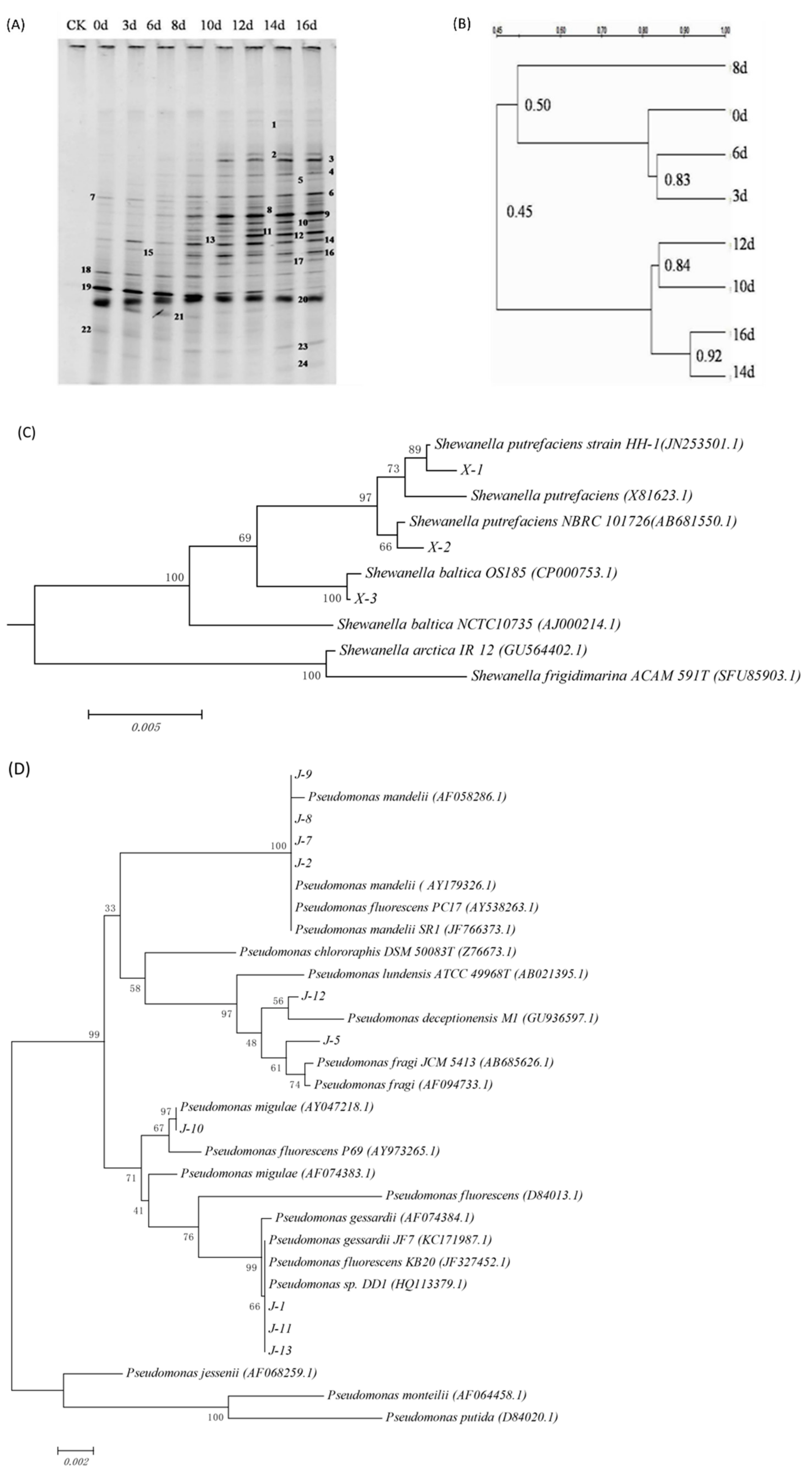
2.1.2. DGGE Analysis
2.2. Sterile Fillet Sample Preparation
2.3. Sensory Analysis
2.4. Microbiological Analysis
2.5. Biochemical Analysis
2.6. Determination of Histamine
2.7. Volatile Compounds Analysis
2.8. Statistical Analysis
3. Results and Discussion
3.1. Bacterial Strains
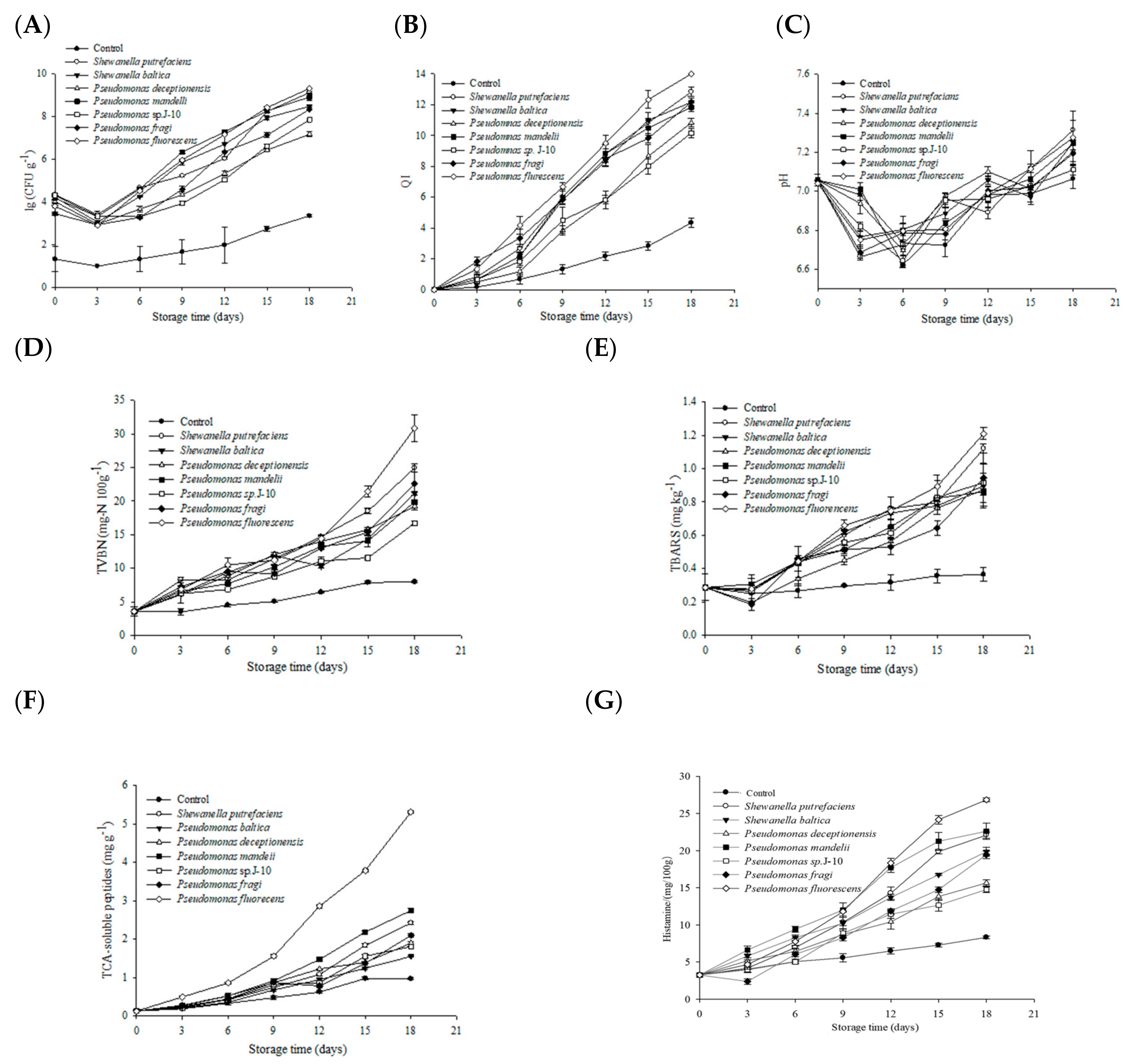
3.2. Microbiological Analysis
3.3. Sensory Analysis
3.4. Biochemical Analysis
3.5. Changes of Histamine
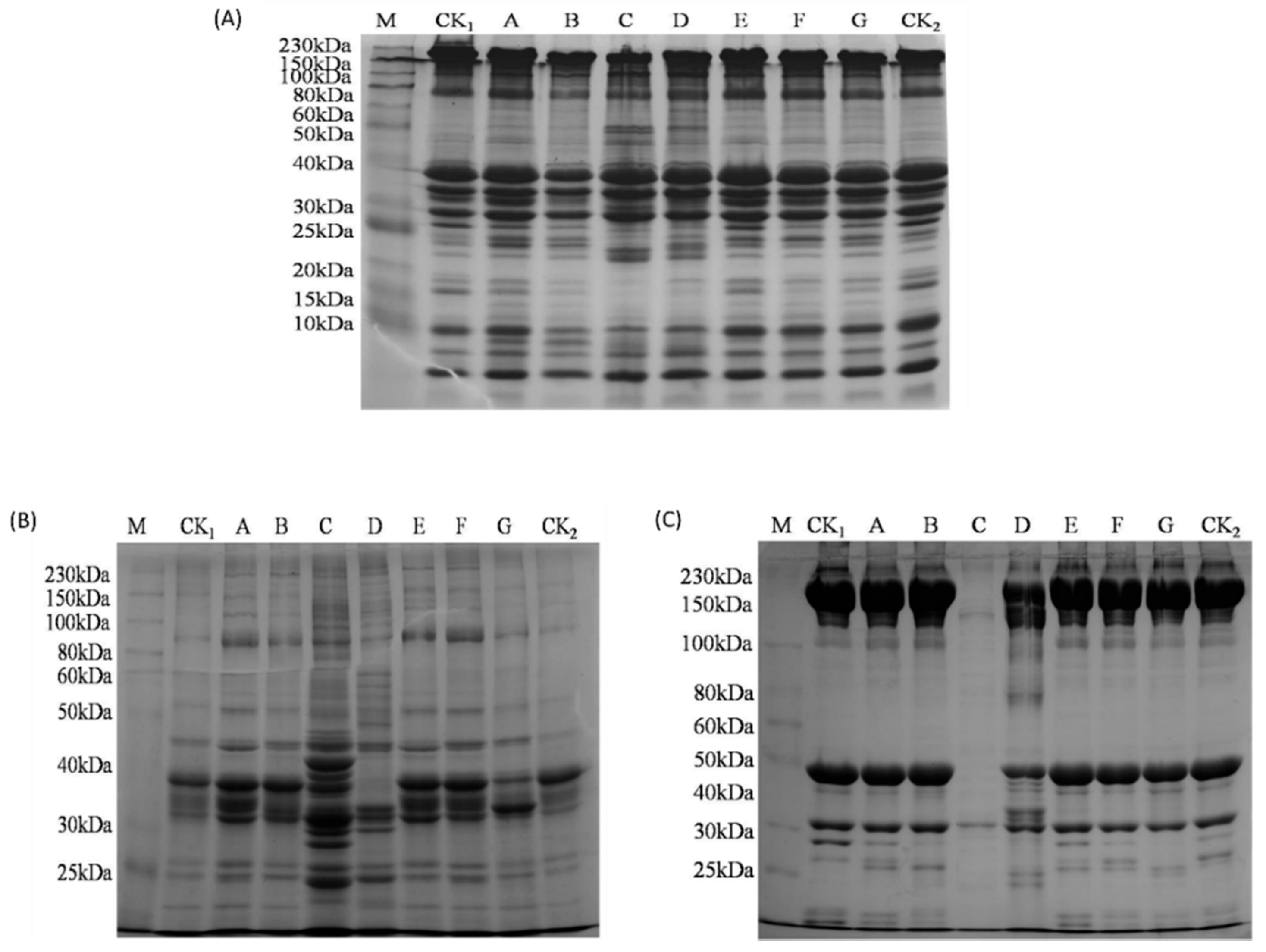
3.6. Proteolytic Degradation Analyzed by SDS-PAGE
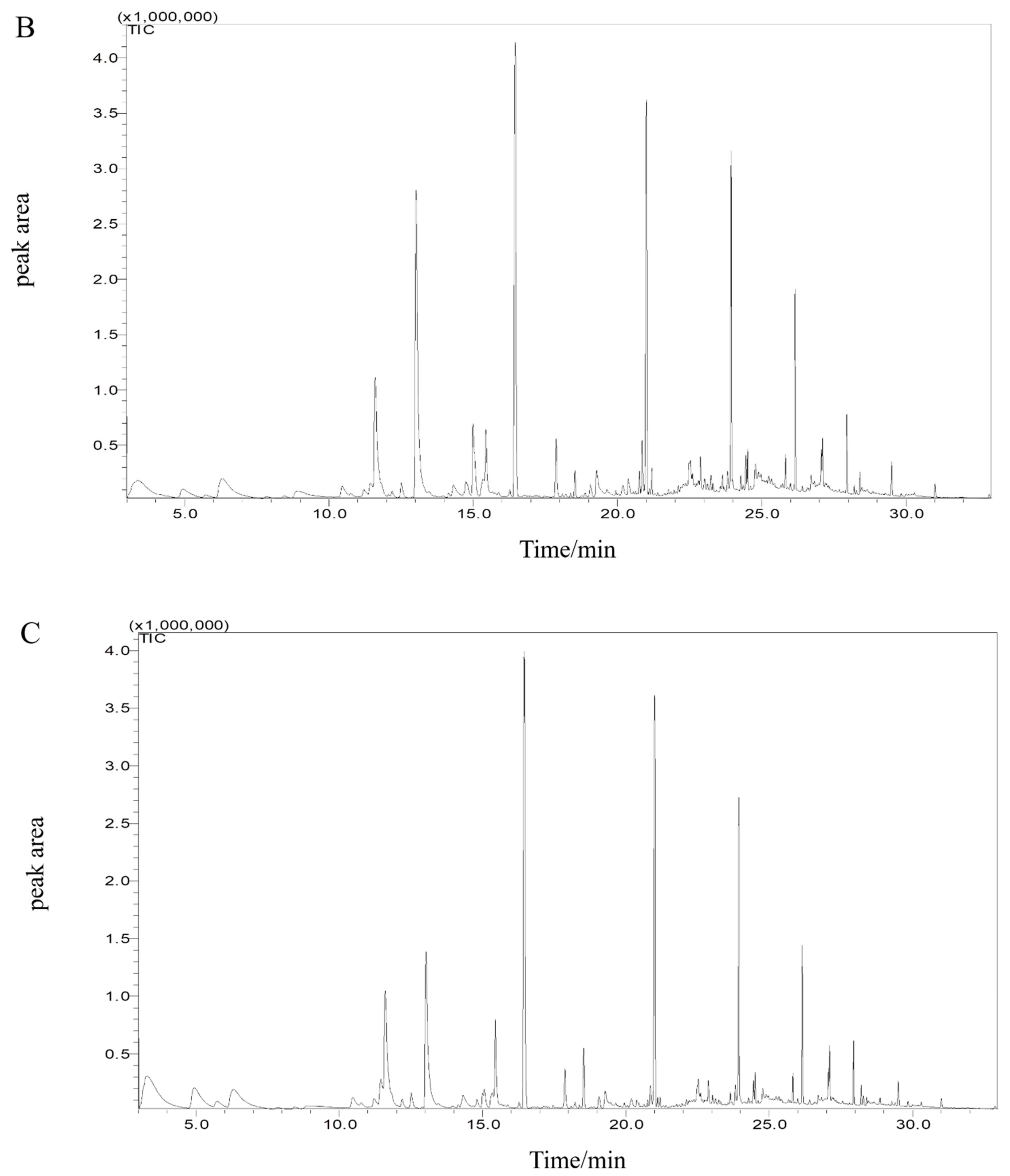
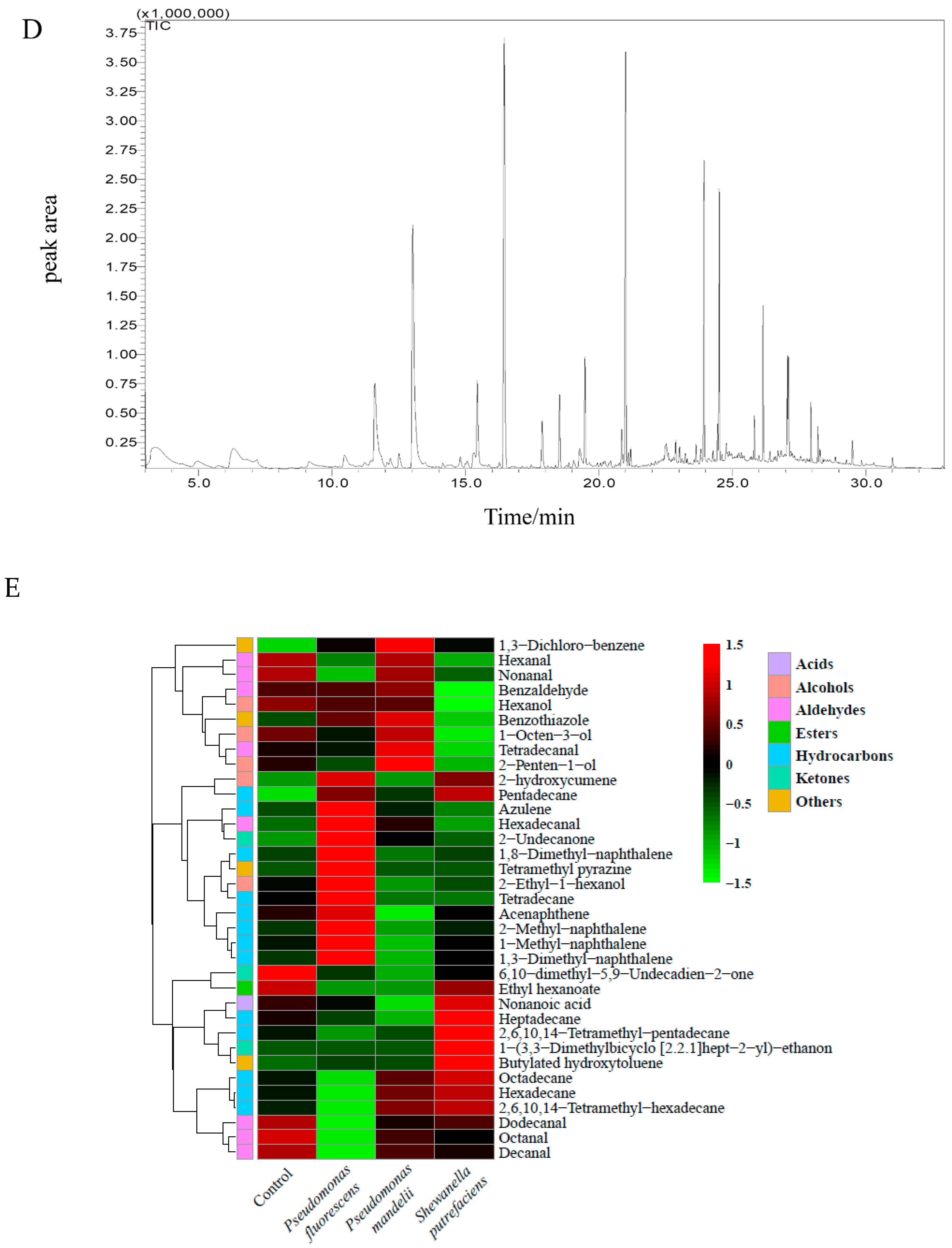
3.7. Effects of Specific Spoilage Bacteria on Volatile Flavor Compounds of Sturgeon
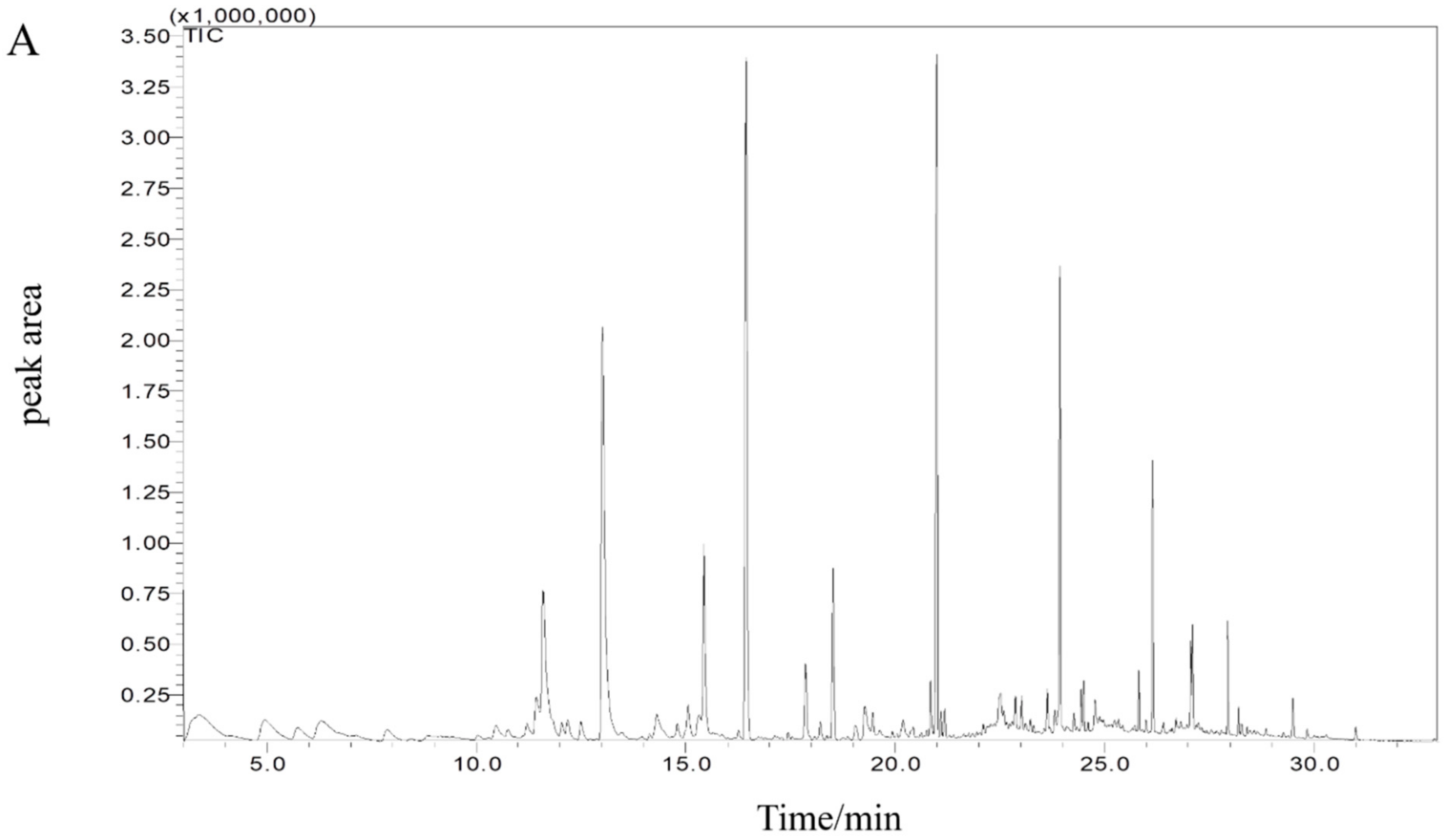
3.7.1. Volatile Compounds Analysis
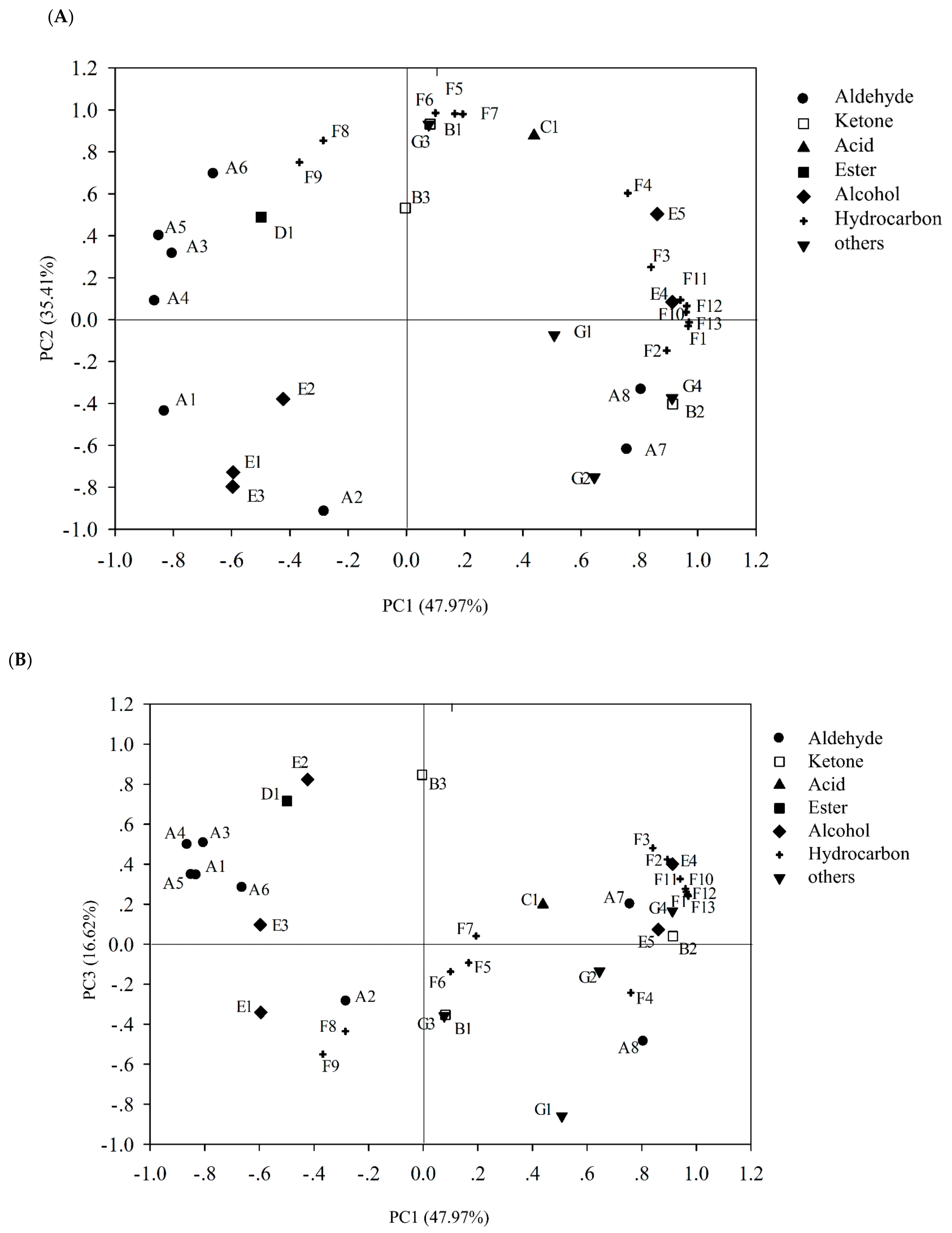
3.7.2. Principal Component Analysis (PCA)
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Food and Agriculture Organization (FAO). The State of World Fisheries and Aquaculture; FAO: Rome, Italy, 2020. [Google Scholar]
- Wei, Q.W.; Zou, Y.; Li, P.; Li, L. Sturgeon aquaculture in China: Progress, strategies and prospects assessed on the basis of nation-wide surveys (2007–2009). J. Appl. Ichthyol. 2011, 27, 162–168. [Google Scholar] [CrossRef]
- Cai, W.; Chen, Y.; Dong, X.; Shi, Y.; Wei, J.; Liu, F. Protein oxidation analysis based on comparative proteomic of Russian sturgeon (Acipenser gueldenstaedti) after sous-vide cooking. Food Control 2021, 121, 107594. [Google Scholar] [CrossRef]
- Islam, M.R.; Yuhi, T.; Meng, D.; Yoshioka, T.; Ogata, Y.; Ura, K.; Takagi, Y. Purity and properties of gelatins extracted from the head tissue of the hybrid kalamtra sturgeon. LWT Food Sci. Technol. 2021, 142, 110944. [Google Scholar] [CrossRef]
- Gui, M.; Wu, R.; Liu, L.; Wang, S.; Zhang, L.; Li, P. Effects of quorum quenching by AHL lactonase on AHLs, protease, motility and proteome patterns in Aeromonas veronii LP-11. Int. J. Food Microbiol. 2017, 252, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Dalgaard, P. Qualitative and quantitative characterization of spoilage bacteria from packed fish. Int. J. Food Microbiol. 1995, 26, 319–333. [Google Scholar] [CrossRef]
- Huang, Z.; Jia, S.; Zhang, L.; Liu, X.; Luo, Y. Inhibitory effects and membrane damage caused to fish spoilage bacteria by cinnamon bark (Cinnamomum tamala) oil. LWT Food Sci. Technol. 2019, 112, 108195. [Google Scholar] [CrossRef]
- Wang, J.; Fang, J.; Wei, L.; Zhang, Y.; Deng, H.; Guo, Y.; Hu, C.; Meng, Y. Decrease of microbial community diversity, biogenic amines formation, and lipid oxidation by phloretin in Atlantic salmon fillets. LWT Food Sci. Technol. 2019, 101, 419–426. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, Y.; Chen, J.; Koseki, S.; Yang, Q.; Yu, H.; Fu, L. Screening and preservation application of quorum sensing inhibitors of Pseudomonas fluorescens and Shewanella baltica in seafood products. LWT Food Sci. Technol. 2021, 149, 111749. [Google Scholar] [CrossRef]
- Cen, S.; Fang, Q.; Tong, L.; Yang, W.; Zhang, J.; Lou, Q.; Huang, T. Effects of chitosan-sodium alginate-nisin preservatives on the quality and spoilage microbiota of Penaeus vannamei shrimp during cold storage. Int. J. Food Microbiol. 2021, 349, 109227. [Google Scholar] [CrossRef]
- Wong, W.W.; Greening, C.; Shelley, G.; Lappan, R.; Leung, P.M.; Kessler, A.; Winfrey, B.; Poh, S.C.; Cook, P. Effects of drift algae accumulation and nitrate loading on nitrogen cycling in a eutrophic coastal sediment. Sci. Total Environ. 2021, 790, 147749. [Google Scholar] [CrossRef]
- Zhao, Y.; Qin, Z.; Huang, Z.; Bao, Z.; Luo, T.; Jin, Y. Effects of polyethylene microplastics on the microbiome and metabolism in larval zebrafish. Environ. Pollut. 2021, 282, 117039. [Google Scholar] [CrossRef]
- Parlapani, F.F.; Michailidou, S.; Anagnostopoulos, D.A.; Koromilas, S.; Kios, K.; Pasentsis, K.; Psomopoulos, F.; Argiriou, A.; Haroutounian, S.A.; Boziaris, I.S. Bacterial communities and potential spoilage markers of whole blue crab (Callinectes sapidus) stored under commercial simulated conditions. Food Microbiol. 2019, 82, 325–333. [Google Scholar] [CrossRef]
- Silbande, A.; Cornet, J.; Cardinal, M.; Chevalier, F.; Rochefort, K.; Smith-Ravin, J.; Adenet, S.; Leroi, F. Characterization of the spoilage potential of pure and mixed cultures of bacterial species isolated from tropical yellowfin tuna (Thunnus albacares). J. Appl. Microbiol. 2018, 124, 559–571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Odeyemi, O.A.; Burke, C.M.; Bolch, C.; Stanley, R. Evaluation of spoilage potential and volatile metabolites production by Shewanella baltica isolated from modified atmosphere packaged live mussels. Food Res. Int. 2018, 103, 415–425. [Google Scholar] [CrossRef] [PubMed]
- Pedros-Garrido, S.; Clemente, I.; Calanche, J.B.; Condon-Abanto, S.; Beltran, J.A.; Lyng, J.G.; Brunton, N.; Bolton, D.; Whyte, P. Antimicrobial activity of natural compounds against listeria spp. and their effects on sensory attributes in salmon (Salmo salar) and cod (Gadus morhua). Food Control 2020, 107, 106768. [Google Scholar] [CrossRef]
- Jia, S.; Huang, Z.; Lei, Y.; Zhang, L.; Li, Y.; Luo, Y. Application of Illumina-MiSeq high throughput sequencing and culture-dependent techniques for the identification of microbiota of silver carp (Hypophthalmichthys molitrix) treated by tea polyphenols. Food Microbiol. 2018, 76, 52–61. [Google Scholar] [CrossRef]
- Joffraud, J.J.; Cardinal, M.; Cornet, J.; Chasles, J.S.; Leon, S.; Gigout, F.; Leroi, F. Effect of bacterial interactions on the spoilage of cold-smoked salmon. Int. J. Food Microbiol. 2006, 112, 51–61. [Google Scholar] [CrossRef]
- Hauschild, P.; Hilgarth, M.; Vogel, R.F. Hydrostatic pressure- and halotolerance of Photobacterium phosphoreum and P. carnosum isolated from spoiled meat and salmon. Food Microbiol. 2021, 99, 103679. [Google Scholar] [CrossRef]
- Leroi, F.; Cornet, J.; Chevalier, F.; Cardinal, M.; Coeuret, G.; Chaillou, S.; Joffraud, J.J. Selection of bioprotective cultures for preventing cold-smoked salmon spoilage. Int. J. Food Microbiol. 2015, 213, 79–87. [Google Scholar] [CrossRef] [Green Version]
- Schaffer, P.A.; Lifland, B.; Van Sommeran, S.; Casper, D.R.; Davis, C.R. Meningoencephalitis associated with Carnobacterium maltaromaticum-like bacteria in stranded juvenile salmon sharks (Lamna ditropis). Vet. Pathol. 2013, 50, 412–417. [Google Scholar] [CrossRef]
- Wagner, E.M.; Fischel, K.; Rammer, N.; Beer, C.; Palmetzhofer, A.L.; Conrady, B.; Roch, F.; Hanson, B.T.; Wagner, M.; Rychli, K. Bacteria of eleven different species isolated from biofilms in a meat processing environment have diverse biofilm forming abilities. Int. J. Food Microbiol. 2021, 349, 109232. [Google Scholar] [CrossRef]
- Turcihan, G.; Turgay, E.; Yardimci, R.E.; Eryalcin, K.M. The effect of feeding with different microalgae on survival, growth, and fatty acid composition of Artemia franciscana metanauplii and on predominant bacterial species of the rearing water. Aquacult. Int. 2021, 1–19. [Google Scholar] [CrossRef]
- Miller, A.; Scanlan, R.; Lee, J.; Libbey, L. Volatile compounds produced in sterile fish muscle (Sebastes melanops) by Pse-domonas putrefaciens, Pseudomonas fluorescens, and an Achromobacter species. Appl. Microbiol. 1973, 26, 18–21. [Google Scholar] [CrossRef]
- Wang, S.; He, J. Two-step denaturing gradient gel electrophoresis (2S-DGGE), a gel-based strategy to capture full-length 16S rRNA gene sequences. Appl. Microbiol. Biot. 2012, 95, 1305–1312. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Liu, X.; Jia, S.; Zhang, L.; Luo, Y. The effect of essential oils on microbial composition and quality of grass carp (Ctenopharyngodon idellus) fillets during chilled storage. Int. J. Food Microbiol. 2018, 266, 52–59. [Google Scholar] [CrossRef] [PubMed]
- Hovda, M.B.; Sivertsvik, M.; Lunestad, B.T.; Lorentzen, G.; Rosnes, J.T. Characterisation of the dominant bacterial population in modified atmosphere packaged farmed halibut (Hippoglossus hippoglossus) based on 16S rDNA-DGGE. Food Microbiol. 2007, 24, 362–371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Botsoglou, N.; Fletouris, D.; Papageorgiou, G.; Vassilopoulos, V.; Mantis, A.; Trakatellis, A. Rapid, sensitive, and specific thi-obarbituric acid method for measuring lipid peroxidation in animal tissue, food, and feedstuff samples. J. Agric. Food Chem. 1994, 42, 1931–1937. [Google Scholar] [CrossRef]
- Broekaert, K.; Heyndrickx, M.; Herman, L.; Devlieghere, F.; Vlaemynck, G. Seafood quality analysis: Molecular identification of dominant microbiota after ice storage on several general growth media. Food Microbiol. 2011, 28, 1162–1169. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Li, J.; Hu, W. Changes in microbiological, physicochemical and muscle proteins of post mortem large yellow croaker (Pseudosciaena crocea). Food Control 2013, 34, 514–520. [Google Scholar] [CrossRef]
- Liu, D.; Liang, L.; Xia, W.; Regenstein, J.M.; Zhou, P. Biochemical and physical changes of grass carp (Ctenopharyngodon idella) fillets stored at −3 and 0 °C. Food Chem. 2013, 140, 105–114. [Google Scholar] [CrossRef]
- Dabade, D.S.; Azokpota, P.; Nout, M.J.R.; Hounhouigan, D.J.; Zwietering, M.H.; den Besten, H.M.W. Prediction of spoilage of tropical shrimp (Penaeus notialis) under dynamic temperature regimes. Int. J. Food Microbiol. 2015, 210, 121–130. [Google Scholar] [CrossRef] [PubMed]
- Reynisson, E.; Lauzon, H.L.; Magnusson, H.; Jonsdottir, R.; Olafsdottir, G.; Marteinsson, V.; Hreggviosson, G.O. Bacterial composition and succession during storage of North-Atlantic cod (Gadus morhua) at superchilled temperatures. BMC Microbiol. 2009, 9, 250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Q.; Zhang, L.; Luo, Y. Changes in microbial communities and quality attributes of white muscle and dark muscle from common carp (Cyprinus carpio) during chilled and freeze-chilled storage. Food Microbiol. 2018, 73, 237–244. [Google Scholar] [CrossRef] [PubMed]
- Mondal, S.; Pramanik, K.; Ghosh, S.K.; Pal, P.; Mondal, T.; Soren, T.; Maiti, T.K. Unraveling the role of plant growth-promoting rhizobacteria in the alleviation of arsenic phytotoxicity: A review. Microbiol. Res. 2021, 250, 126809. [Google Scholar] [CrossRef]
- Liu, X.; Huang, Z.; Jia, S.; Zhang, J.; Li, K.; Luo, Y. The roles of bacteria in the biochemical changes of chill-stored bighead carp (Aristichthys nobilis): Proteins degradation, biogenic amines accumulation, volatiles production, and nucleotides catabolism. Food Chem. 2018, 255, 174–181. [Google Scholar] [CrossRef]
- Hernandez, M.D.; Lopez, M.B.; Alvarez, A.; Ferrandini, E.; Garcia, B.G.; Garrido, M.D. Sensory, physical, chemical and microbiological changes in aquacultured meagre (Argyrosomus regius) fillets during ice storage. Food Chem. 2009, 114, 237–245. [Google Scholar] [CrossRef]
- Zhang, Q.; Chen, X.; Ding, Y.; Ke, Z.; Zhou, X.; Zhang, J. Diversity and succession of the microbial community and its correlation with lipid oxidation in dry-cured black carp (Mylopharyngodon piceus) during storage. Food Microbiol. 2021, 98, 103686. [Google Scholar] [CrossRef]
- Nychas, G.; Dillon, V.; Board, R. Glucose, the key substrate in the microbiological changes occurring in meat and certain meat products. Biotechnol. Appl. Bioc. 1988, 10, 203–231. [Google Scholar]
- Saenz-Garcia, C.E.; Castaneda-Serrano, P.; Silva, E.M.M.; Alvarado, C.Z.; Nava, G.M. Insights into the Identification of the Specific Spoilage Organisms in Chicken Meat. Foods 2020, 9, 225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doulgeraki, A.I.; Ercolini, D.; Villani, F.; Nychas, G.E. Spoilage microbiota associated to the storage of raw meat in different conditions. Int. J. Food Microbiol. 2012, 157, 130–141. [Google Scholar] [CrossRef]
- Liu, C.; Li, W.; Lin, B.; Yi, S.; Ye, B.; Mi, H.; Li, J.; Wang, J.; Li, X. Effects of ozone water rinsing on protein oxidation, color, and aroma characteristics of grass carp (Ctenopharyngodon idellus) surimi. J. Food Process. Pres. 2021, e15811. [Google Scholar] [CrossRef]
- Mace, S.; Joffraud, J.; Cardinal, M.; Malcheva, M.; Cornet, J.; Lalanne, V.; Chevalier, F.; Serot, T.; Pilet, M.; Dousset, X. Evaluation of the spoilage potential of bacteria isolated from spoiled raw salmon (Salmo salar) fillets stored under modified atmosphere packaging. Int. J. Food Microbiol. 2013, 160, 227–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Selli, S.; Cayhan, G.G. Analysis of volatile compounds of wild gilthead sea bream (Sparus aurata) by simultaneous distillation-extraction (SDE) and GC-MS. Microchem. J. 2009, 93, 232–235. [Google Scholar] [CrossRef]
- Iglesias, J.; Medina, I.; Bianchi, F.; Careri, M.; Mangia, A.; Musci, M. Study of the volatile compounds useful for the characterisation of fresh and frozen-thawed cultured gilthead sea bream fish by solid-phase microextraction gas chromatography-mass spectrometry. Food Chem. 2009, 115, 1473–1478. [Google Scholar] [CrossRef]
- Zhang, Z.; Li, G.; Luo, L.; Chen, G. Study on seafood volatile profile characteristics during storage and its potential use for freshness evaluation by headspace solid phase microextraction coupled with gas chromatography-mass spectrometry. Anal. Chim. Acta 2010, 659, 151–158. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number | Most Similar Strain | Login Number | Similarity |
|---|---|---|---|
| 1 | Shewanella sp. B157 | FN295752.1 | 95% |
| 2 | Pseudomonas sp. 01WB03.2-36 | FM161452.1 | 96% |
| 3 | Uncultured bacterium clone C16 | AM411553.1 | 95% |
| 4 | Shewanella baltica strain SS3 | JX032783.1 | 96% |
| 5 | Shewanella baltica strain ST5 | JX032790.1 | 97% |
| 6 | Shewanella baltica OS678 | CP002383.1 | 97% |
| 7 | Pseudomonas fluorescens strain M2 | GU947867.1 | 97% |
| 8 | Pseudomonas sp. MFY222 | AY331375.1 | 99% |
| 9 | Pseudomonas sp. R-45822 | FR775122.1 | 100% |
| 10 | Pseudomonas fluorescens | EF408245.1 | 100% |
| 11 | Pseudomonas sp. CB10 | EU482914.1 | 99% |
| 12 | Pseudomonas sp. MDT2-39-4 | JX949559.1 | 99% |
| 13 | Uncultured bacterium clone 07MIC062 | JF340783.1 | 99% |
| 14 | Pseudomonas sp. 01WB02.2-10 | FM161381.1 | 95% |
| 15 | Staphylococcus warneri strain E3b | AY126243.1 | 98% |
| 16 | Bacterium het-w-28 | KC810293.1 | 99% |
| 17 | Pseudomonas sp. CB10 | EU482914.1 | 98% |
| 18 | Pseudomonas sp. IGS61 | JN680232.1 | 99% |
| 19 | Acinetobacter sp. EAXY14 | KC129073.1 | 99% |
| 20 | Shewanella sp. B157 | FN295752.1 | 96% |
| 21 | Microbulbifer maritimus strain MTM147 | HQ705770.1 | 93% |
| 22 | Uncultured Escherichia sp. N5 | AM712054.1 | 98% |
| 23 | Gamma proteobacterium BAL382 | KC140316.1 | 97% |
| 24 | Alteromonadales bacterium HD-I-03-6 | AM931110.1 | 97% |
| PC1 (47.97%) | PC2 (35.41%) | PC3 (16.62%) | |
|---|---|---|---|
| Control | −3.18 | 0.06 | 3.10 |
| Pseudomonas fluorescens | 5.61 | −2.00 | 0.60 |
| Pseudomonas mandelii | −2.93 | −3.01 | −2.42 |
| Shewanella putrefaciens | 0.50 | 4.92 | −1.28 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Z.; Wu, R.; Gui, M.; Jiang, Z.; Li, P. Identification of the Specific Spoilage Organism in Farmed Sturgeon (Acipenser baerii) Fillets and Its Associated Quality and Flavour Change during Ice Storage. Foods 2021, 10, 2021. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10092021
Zhang Z, Wu R, Gui M, Jiang Z, Li P. Identification of the Specific Spoilage Organism in Farmed Sturgeon (Acipenser baerii) Fillets and Its Associated Quality and Flavour Change during Ice Storage. Foods. 2021; 10(9):2021. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10092021
Chicago/Turabian StyleZhang, Zhichao, Ruiyun Wu, Meng Gui, Zhijie Jiang, and Pinglan Li. 2021. "Identification of the Specific Spoilage Organism in Farmed Sturgeon (Acipenser baerii) Fillets and Its Associated Quality and Flavour Change during Ice Storage" Foods 10, no. 9: 2021. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10092021