
Influence of Cabbage (Brassica oleracea) Accession and Growing Conditions on Myrosinase Activity, Glucosinolates and Their Hydrolysis Products


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Reagents and Chemicals
2.3. Sample Preparation
2.4. Myrosinase Enzyme Extraction and Assay
2.5. Protein Assay
2.6. Glucosinolate Extraction and LC-MS2 Analysis
2.7. Extraction of Glucosinolate Hydrolysis Products
2.8. GC-MS Analysis
2.9. Statistical Analysis
3. Results and Discussion
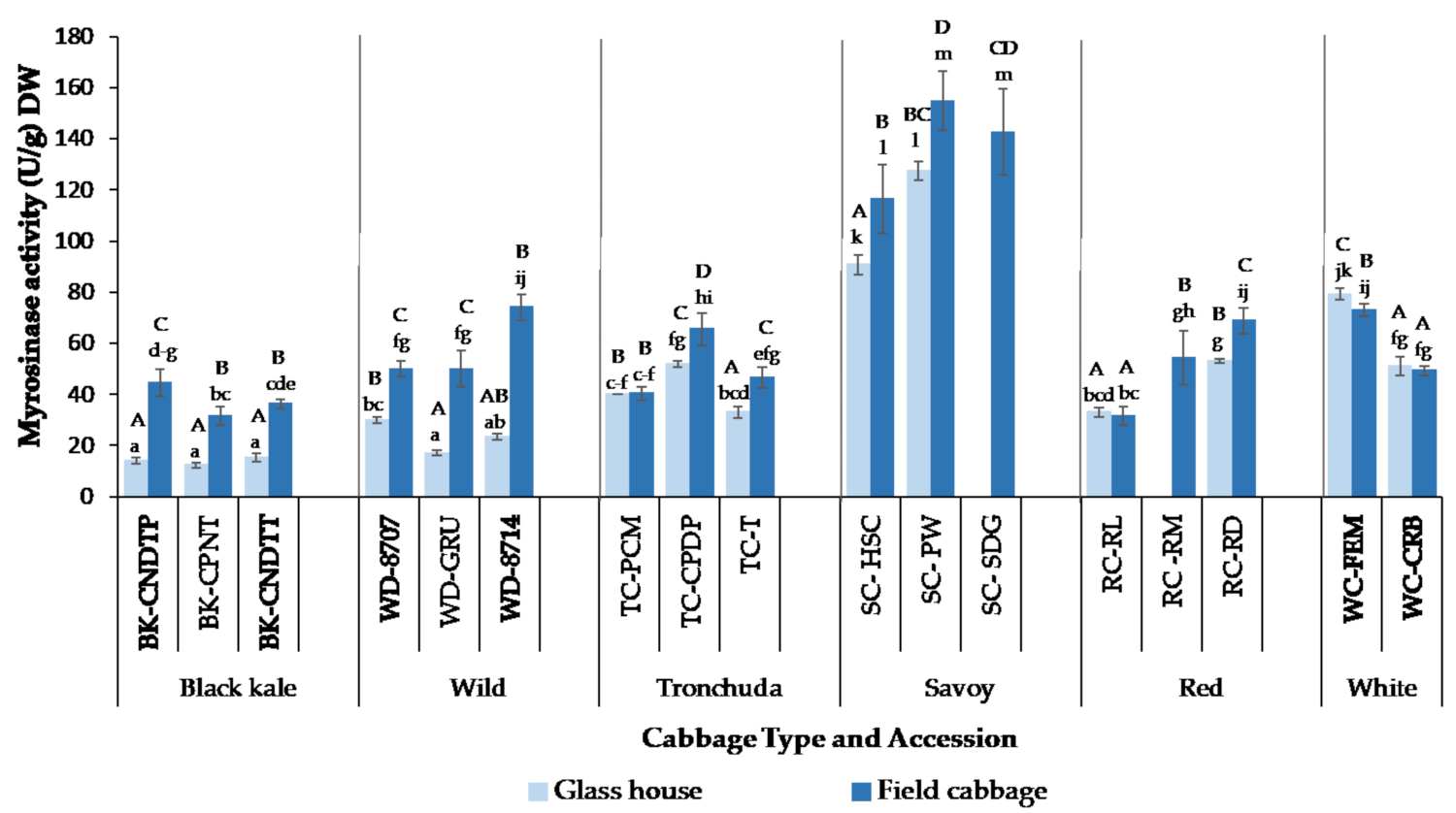
3.1. Effect of Growing Conditions, Cabbage Morphotype and Accession on Myrosinase Activity
3.2. Protein Content and Specific Myrosinase Activity of Glasshouse and Field Grown Cabbages
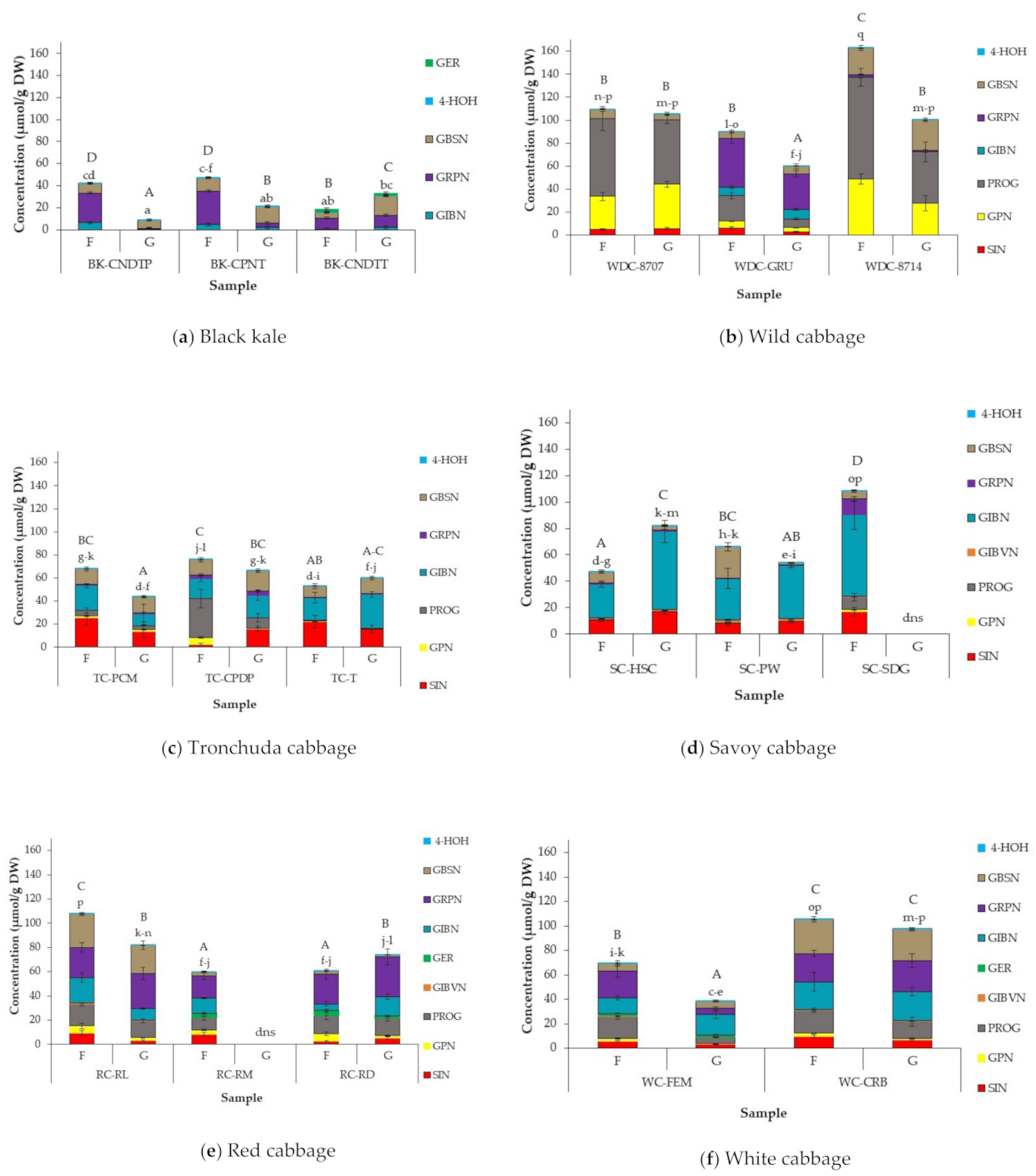
3.3. Effect of Cabbage Morphotype and Accession on GSL Profile and Concentration of Field Grown Cabbages
3.4. Effect of Growing Conditions on GSL Concentrations in Cabbage Accessions
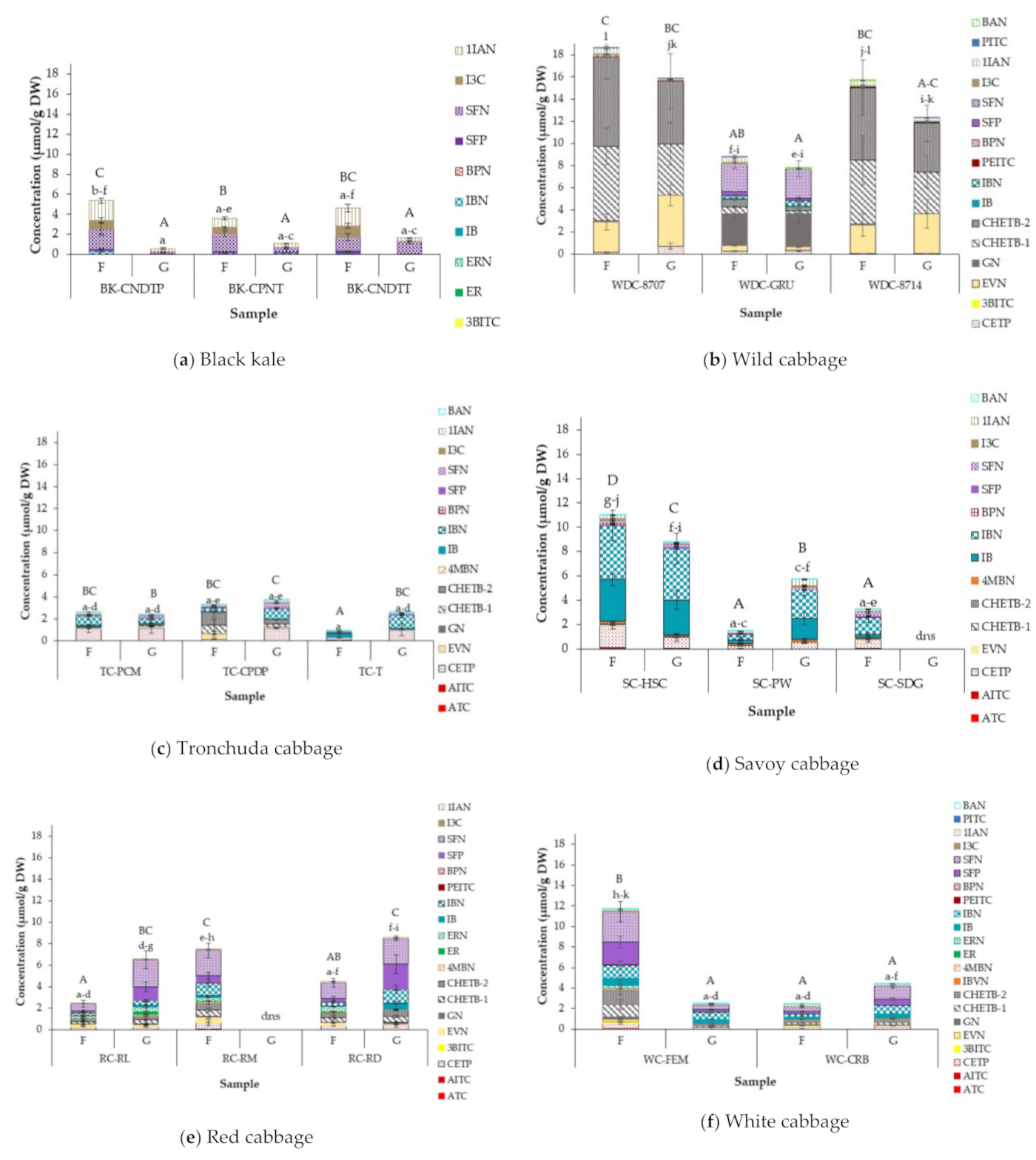
3.5. Effect of of Cabbage Morphotype and Accession on Glucosinolate Hydrolysis Products (GHPs) of Field Grown Cabbages
3.6. Effect of Growing Condition on GHP Concentrations
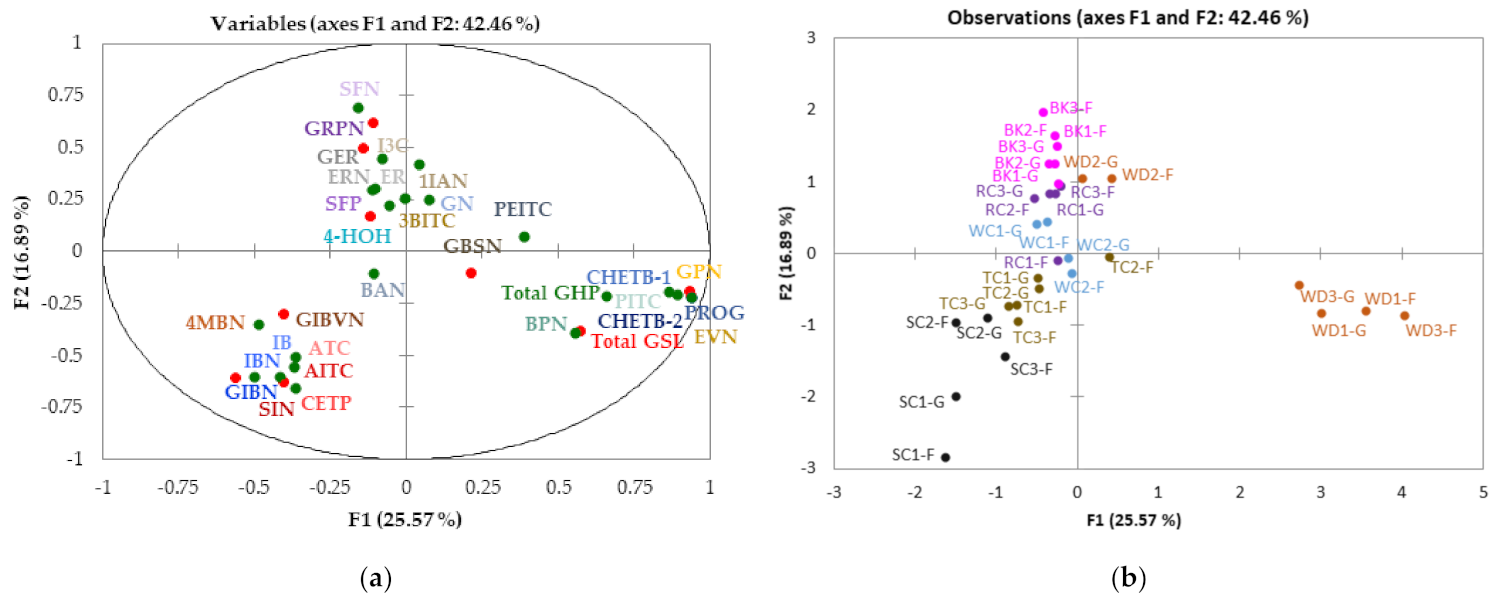
3.7. Multifactor Analysis (MFA) of GSLs and GHPs Identified in Cabbage Accessions Grown under Two Different Conditions
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Plants of the World Online. Available online: http://powo.science.kew.org/taxon/urn:lsid:ipni.org:names:279435-1 (accessed on 24 May 2021).
- Herr, I.; Buchler, M.W. Dietary constituents of broccoli and other cruciferous vegetables: Implications for prevention and therapy of cancer. Cancer Treat. Rev. 2010, 36, 377–383. [Google Scholar] [CrossRef] [PubMed]
- Verkerk, R.; Dekker, M.; Jongen, W.M.F. Post-harvest increase of indolyl glucosinolates in response to chopping and storage of Brassica vegetables. J. Sci. Food Agric. 2001, 81, 953–958. [Google Scholar] [CrossRef]
- Guerrero-Beltran, C.E.; Calderon-Oliver, M.; Pedraza-Chaverri, J.; Chirino, Y.I. Protective effect of sulforaphane against oxidative stress: Recent advances. Exp. Toxicol. Pathol. 2012, 64, 503–508. [Google Scholar] [CrossRef] [PubMed]
- Mithen, R.F.; Dekker, M.; Verkerk, R.; Rabot, S.; Johnson, I.T. The nutritional significance, biosynthesis and bioavailability of glucosinolates in human foods. J. Sci. Food Agric. 2000, 80, 967–984. [Google Scholar] [CrossRef]
- Mithen, R. Glucosinolates—Biochemistry, genetics and biological activity. Plant Growth Regul. 2001, 34, 91–103. [Google Scholar] [CrossRef]
- Redovnikovi, I.R.; Gliveti, T.; Delonga, K.; Vorkapi-Fura, J. Glucosinolates and their potential role in plant. Period. Biol. 2008, 110, 297–309. [Google Scholar]
- Bjorkman, M.; Klingen, I.; Birch, A.N.E.; Bones, A.M.; Bruce, T.J.A.; Johansen, T.J.; Meadow, R.; Molmann, J.; Seljasen, R.; Smart, L.E.; et al. Phytochemicals of Brassicaceae in plant protection and human health—Influences of climate, environment and agronomic practice. Phytochemistry 2011, 72, 538–556. [Google Scholar] [CrossRef] [PubMed]
- Wittstock, U.; Halkier, B.A. Glucosinolate research in the Arabidopsis era. Trends Plant Sci. 2002, 7, 263–270. [Google Scholar] [CrossRef]
- Halkier, B.A.; Gershenzon, J. Biology and Biochemistry of Glucosinolates. Annu. Rev. Plant Biol. 2006, 57, 303–333. [Google Scholar] [CrossRef] [Green Version]
- Blažević, I.; Montaut, S.; Burčul, F.; Olsen, C.; Burow, M.; Rollin, P.; Agerbirk, N. Glucosinolate structural diversity, identification, chemical synthesis and metabolism in plants. Phytochemistry 2020, 169, 112100. [Google Scholar] [CrossRef]
- Bones, A.M.; Rossiter, J.T. The myrosinase-glucosinolate system, its organisation and biochemistry. Physiol. Plant. 1996, 97, 194–208. [Google Scholar] [CrossRef]
- Ludikhuyze, L.; Rodrigo, L.; Hendrickx, M. The activity of myrosinase from broccoli (Brassica oleracea L. cv. italica): Influence of intrinsic and extrinsic factors. J. Food Prot. 2000, 63, 400–403. [Google Scholar] [CrossRef]
- Morimitsu, Y.; Hayashi, K.; Nakagawa, Y.; Fujii, H.; Horio, F.; Uchida, K.; Osawa, T. Antiplatelet and anticancer isothiocyanates in Japanese domestic horseradish, Wasabi. Mech. Ageing Dev. 2000, 116, 125–134. [Google Scholar] [CrossRef]
- Hanschen, F.S.; Lamy, E.; Schreiner, M.; Rohn, S. Reactivity and stability of glucosinolates and their breakdown products in foods. Angew. Chem. Int. Ed. 2014, 53, 11430–11450. [Google Scholar] [CrossRef] [PubMed]
- Tsai, S.-C.; Huang, W.-W.; Huang, W.-C.; Lu, C.-C.; Chiang, J.-H.; Peng, S.-F.; Chung, J.-G.; Lin, Y.-H.; Hsu, Y.-M.; Amagaya, S. ERK-modulated intrinsic signaling and G2/M phase arrest contribute to the induction of apoptotic death by allyl isothiocyanate in MDA-MB-468 human breast adenocarcinoma cells. Int. J. Oncol. 2012, 41, 2065–2072. [Google Scholar] [CrossRef] [PubMed]
- Leoni, O.; Iori, R.; Palmieri, S.; Esposito, E.; Menegatti, E.; Cortesi, R.; Nastruzzi, C. Myrosinase-generated isothiocyanate from glucosinolates: Isolation, characterization and in vitro antiproliferative studies. Bioorg. Med. Chem. 1997, 5, 1799–1806. [Google Scholar] [CrossRef]
- Tripathi, K.; Hussein, U.K.; Anupalli, R.; Barnett, R.; Bachaboina, L.; Scalici, J.; Rocconi, R.P.; Owen, L.B.; Piazza, G.A.; Palle, K. Allyl isothiocyanate induces replication-associated DNA damage response in NSCLC cells and sensitizes to ionizing radiation. Oncotarget 2015, 6, 5237. [Google Scholar] [CrossRef]
- Lambrix, V.; Reichelt, M.; Mitchell-Olds, T.; Kliebenstein, D.J.; Gershenzon, J. The Arabidopsis Epithiospecifier Protein Promotes the Hydrolysis of Glucosinolates to Nitriles and Influences Trichoplusia ni Herbivory. Plant Cell 2001, 13, 2793–2807. [Google Scholar] [CrossRef] [Green Version]
- Baik, H.Y.; Juvik, J.; Jeffery, E.H.; Wallig, M.A.; Kushad, M.; Klein, B.P. Relating glucosinolate content and flavor of broccoli cultivars. J. Food Sci. 2003, 68, 1043–1050. [Google Scholar] [CrossRef]
- Cox, D.N.; Melo, L.; Zabaras, D.; Delahunty, C.M. Acceptance of health-promoting Brassica vegetables: The influence of taste perception, information and attitudes. Public Health Nutr. 2012, 15, 1474–1482. [Google Scholar] [CrossRef] [Green Version]
- Beck, T.K.; Jensen, S.; Bjoern, G.K.; Kidmose, U. The Masking Effect of Sucrose on Perception of Bitter Compounds in Brassica Vegetables. J. Sens. Stud. 2014, 29, 190–200. [Google Scholar] [CrossRef]
- Doorn, H.E.v.; Kruk, G.C.v.d.; Holst, G.-J.v.; Raaijmakers-Ruijs, N.C.; Postma, E.; Groeneweg, B.; Jongen, W.H.F. The Glucosinolates Sinigrin and Progoitrin are important determination for taste. J. Sci. Food Agric. 1998, 78, 30–38. [Google Scholar] [CrossRef]
- Wei, J.; Miao, H.; Wang, Q. Effect of glucose on glucosinolates, antioxidants and metabolic enzymes in Brassica sprouts. Sci. Hortic. 2011, 129, 535–540. [Google Scholar] [CrossRef]
- Charron, C.S.; Saxton, A.M.; Sams, C.E. Relationship of climate and genotype to seasonal variation in the glucosinolate–myrosinase system. II. Myrosinase activity in ten cultivars of Brassica oleracea grown in fall and spring seasons. J. Sci. Food Agric. 2005, 85, 682–690. [Google Scholar] [CrossRef]
- Charron, C.S.; Sams, C.E. Glucosinolate content and myrosinase activity in rapid-cycling Brassica oleracea grown in a controlled environment. J. Am. Soc. Hort. Sci. 2004, 129, 321–330. [Google Scholar] [CrossRef] [Green Version]
- Rosa, E.A.; Rodrigues, P.M. The effect of light and temperature on glucosinolate concentration in the leaves and roots of cabbage seedlings. J. Sci. Food Agric. 1998, 78, 208–212. [Google Scholar] [CrossRef]
- Yen, G.-C.; Wei, Q.-K. Myrosinase activity and total glucosinolate content of cruciferous vegetables, and some properties of cabbage myrosinase in Taiwan. J. Sci. Food Agric. 1993, 61, 471–475. [Google Scholar] [CrossRef]
- Penas, E.; Frias, J.; Martinez-Villaluenga, C.; Vidal-Valverde, C. Bioactive compounds, myrosinase activity, and antioxidant capacity of white cabbages grown in different locations of Spain. J. Agric. Food Chem. 2011, 59, 3772–3779. [Google Scholar] [CrossRef] [Green Version]
- Farnham, M.W.; Wilson, P.E.; Stephenson, K.K.; Fahey, J.W. Genetic and environmental effects on glucosinolate content and chemoprotective potency of broccoli. Plant Breed. 2004, 123, 60–65. [Google Scholar] [CrossRef]
- Verkerk, R.; Schreiner, M.; Krumbein, A.; Ciska, E.; Holst, B.; Rowland, I.; Schrijver, R.D.; Hansen, M.; Gerhaeuser, G.; Mithen, R.; et al. Glucosinolates in Brassica vegetables: The influence of the food supply chain on intake, bioavailability and human health. Mol. Nutr. Food Res. 2009, 53, S219. [Google Scholar] [CrossRef] [PubMed]
- Velasco, P.; Cartea, M.E.; González, C.; Vilar, M.; Ordás, A. Factors affecting the glucosinolate content of kale (Brassica oleracea acephala group). J. Agric. Food Chem. 2007, 55, 955–962. [Google Scholar] [CrossRef]
- Steindal, A.L.H.; Rødven, R.; Hansen, E.; Mølmann, J. Effects of photoperiod, growth temperature and cold acclimatisation on glucosinolates, sugars and fatty acids in kale. Food Chem. 2015, 174, 44–51. [Google Scholar] [CrossRef] [PubMed]
- Rungapamestry, V.; Duncan, A.J.; Fuller, Z.; Ratcliffe, B. Changes in glucosinolate concentrations, myrosinase activity, and production of metabolites of glucosinolates in cabbage (Brassica oleracea var. capitata) cooked for different durations. J. Agric. Food Chem. 2006, 54, 7628–7634. [Google Scholar] [CrossRef] [PubMed]
- Rungapamestry, V.; Duncan, A.J.; Fuller, Z.; Ratcliffe, B. Influence of blanching and freezing broccoli (Brassica oleracea var. italica) prior to storage and cooking on glucosinolate concentrations and myrosinase activity. Eur. Food Res. Technol. 2008, 227, 37. [Google Scholar]
- Ghawi, S.K.; Methven, L.; Rastall, R.A.; Niranjan, K. Thermal and high hydrostatic pressure inactivation of myrosinase from green cabbage: A kinetic study. Food Chem. 2012, 131, 1240–1247. [Google Scholar] [CrossRef]
- Okunade, O.A.; Methven, L.; Niranjan, K. A comparison of myrosinase activity and stability in fresh broccoli (B. oleracea var. italica) and Brown Mustard (B. juncea) Seeds. Turk. J. Agric.-Food Sci. Technol. 2020, 8, 64. [Google Scholar] [CrossRef] [Green Version]
- Singh, J.; Rai, M.; Upadhyay, A.K.; Prasad, K. Sinigrin (2-propenyl glucosinolate) content and myrosinase activity in Brassica vegetables. Int. J. Veg. Sci. 2007, 13, 21–31. [Google Scholar] [CrossRef]
- Charron, C.S.; Saxton, A.M.; Sams, C.E. Relationship of climate and genotype to seasonal variation in the glucosinolate-myrosinase system. I. Glucosinolate content in ten cultivars of Brassica oleracea grown in fall and spring seasons. J. Sci. Food Agric. 2005, 85, 671–681. [Google Scholar] [CrossRef]
- Cartea, M.E.; Velasco, P.; Obregon, S.; Padilla, G.; de Haro, A. Seasonal variation in glucosinolate content in Brassica oleracea crops grown in northwestern Spain. Phytochemistry 2008, 69, 403–410. [Google Scholar] [CrossRef]
- Ciska, E.; Martyniak-Przybyszewska, B.; Kozlowska, H. Content of glucosinolates in cruciferous vegetables grown at the same site for two years under different climatic conditions. J. Agric. Food Chem. 2000, 48, 2862–2867. [Google Scholar] [CrossRef]
- Jasper, J.; Wagstaff, C.; Bell, L. Growth temperature influences postharvest glucosinolate concentrations and hydrolysis product formation in first and second cuts of rocket salad. Postharvest Biol. Technol. 2020, 163, 111157. [Google Scholar] [CrossRef] [PubMed]
- Hanschen, F.S.; Schreiner, M. Isothiocyanates, nitriles, and epithionitriles from glucosinolates are affected by genotype and developmental stage in Brassica oleracea varieties. Front. Plant Sci. 2017, 8, 1095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daxenbichler, M.E.; Van Etten, C.H.; Spencer, G.F. Glucosinolates and derived products in cruciferous vegetables. Identification of organic nitriles from cabbage. J. Agric. Food Chem. 1977, 25, 121–124. [Google Scholar] [CrossRef]
- Van Eylen, D.; Bellostas, N.; Strobel, B.W.; Oey, I.; Hendrickx, M.; Van Loey, A.; Sørensen, H.; Sørensen, J.C. Influence of pressure/temperature treatments on glucosinolate conversion in broccoli (Brassica oleraceae L. cv Italica) heads. Food Chem. 2009, 112, 646–653. [Google Scholar] [CrossRef]
- Church, A.; Mitchell, R.; Ravenscroft, N.; Stapleton, L.M. ‘Growing your own’: A multi-level modelling approach to understanding personal food growing trends and motivations in Europe. Ecol. Econ. 2015, 110, 71–80. [Google Scholar] [CrossRef] [Green Version]
- Tolstrup, J. The Rise of the Indoor Gardening Movement. Available online: https://blog.backtotheroots.com/2018/03/14/rise-indoor-gardening-movement/ (accessed on 25 May 2021).
- Ghawi, S.K.; Methven, L.; Niranjan, K. The potential to intensify sulforaphane formation in cooked broccoli (Brassica oleracea var. italica) using mustard seeds (Sinapis alba). Food Chem. 2013, 138, 1734–1741. [Google Scholar] [CrossRef]
- Gatfield, I.L.; Sand, T. A coupled enzymatic procedure for the determination of myrosinase activity. Lebensm.-Wiss. Technol. 1983, 16, 73–75. [Google Scholar]
- Wilkinson, A.P.; Rhodes, M.J.C.; Fenwick, R.G. Myrosinase Activity of Cruciferous Vegetables. J. Sci. Food Agric. 1984, 35, 543–552. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Bell, L.; Oruna-Concha, M.J.; Wagstaff, C. Identification and quantification of glucosinolate and flavonol compounds in rocket salad (Eruca sativa, Eruca vesicaria and Diplotaxis tenuifolia) by LC–MS: Highlighting the potential for improving nutritional value of rocket crops. Food Chem. 2015, 172, 852–861. [Google Scholar] [CrossRef] [Green Version]
- Clarke, D.B. Glucosinolates, structures and analysis in food. Anal. Methods 2010, 2, 310. [Google Scholar] [CrossRef]
- Rochfort, S.J.; Trenerry, V.C.; Imsic, M.; Panozzo, J.; Jones, R. Class targeted metabolomics: ESI ion trap screening methods for glucosinolates based on MSn fragmentation. Phytochemistry 2008, 69, 1671–1679. [Google Scholar] [CrossRef] [PubMed]
- Lelario, F.; Bianco, G.; Bufo, S.A.; Cataldi, T.R.I. Establishing the occurrence of major and minor glucosinolates in Brassicaceae by LC–ESI-hybrid linear ion-trap and Fourier-transform ion cyclotron resonance mass spectrometry. Phytochemistry 2012, 73, 74–83. [Google Scholar] [CrossRef]
- Bennett, R.N.; Mellon, F.A.; Kroon, P.A. Screening crucifer seeds as sources of specific intact glucosinolates using ion-pair high-performance liquid chromatography negative ion electrospray mass spectrometry. J. Agric. Food Chem. 2004, 52, 428–438. [Google Scholar] [CrossRef]
- Bell, L.; Yahya, H.N.; Oloyede, O.O.; Methven, L.; Wagstaff, C. Changes in rocket salad phytochemicals within the commercial supply chain: Glucosinolates, isothiocyanates, amino acids and bacterial load increase significantly after processing. Food Chem. 2017, 221, 521–534. [Google Scholar] [CrossRef]
- Al-Gendy, A.A.; Lockwood, G.B. GC-MS analysis of volatile hydrolysis products from glucosinolates in Farsetia aegyptia var. ovalis. Flavour Fragr. J. 2003, 18, 148–152. [Google Scholar] [CrossRef]
- Arora, R.; Sharma, D.; Kumar, R.; Singh, B.; Vig, A.P.; Arora, S. Evaluating extraction conditions of glucosinolate hydrolytic products from seeds of Eruca sativa (Mill.) Thell. using GC-MS. J. Food Sci. 2014, 79, C1964–C1969. [Google Scholar] [CrossRef]
- Hong, E.; Kim, G.-H. GC-MS analysis of the extracts from Korean cabbage (Brassica campestris L. ssp. pekinensis) and its seed. Prev. Nutr. Food Sci. 2013, 18, 218–221. [Google Scholar] [CrossRef] [Green Version]
- Spencer, G.F.; Daxenbichler, M.E. Gas chromatography-mass spectrometry of nitriles, isothiocyanates and oxazolidinethiones derived from cruciferous glucosinolates. J. Sci. Food Agric. 1980, 31, 359–367. [Google Scholar] [CrossRef]
- Hanschen, F.S.; Klopsch, R.; Oliviero, T.; Schreiner, M.; Verkerk, R.; Dekker, M. Optimizing isothiocyanate formation during enzymatic glucosinolate breakdown by adjusting pH value, temperature and dilution in Brassica vegetables and Arabidopsis thaliana. Sci. Rep. 2017, 7, 40807. [Google Scholar] [CrossRef]
- de Pinho, P.G.; Valentão, P.; Gonçalves, R.F.; Sousa, C.; Andrade, P.B. Volatile composition of Brassica oleracea L. var. costata DC leaves using solid-phase microextraction and gas chromatography/ion trap mass spectrometry. Rapid Commun. Mass Spectrom. 2009, 23, 2292–2300. [Google Scholar] [CrossRef] [PubMed]
- Vaughn, S.F.; Palmquist, D.E.; Duval, S.M.; Berhow, M.A. Herbicidal activity of glucosinolate-containing seedmeals. Weed Sci. 2017, 54, 743–748. [Google Scholar] [CrossRef]
- Travers-Martin, N.; Kuhlmann, F.; Muller, C. Revised determination of free and complexed myrosinase activities in plant extracts. Plant Physiol. Biochem. 2008, 46, 506–516. [Google Scholar] [CrossRef] [PubMed]
- Grace, J. 3. Plant response to wind. Agric. Ecosyst. Environ. 1988, 22-23, 71–88. [Google Scholar] [CrossRef]
- Hirai, M.Y.; Yano, M.; Goodenowe, D.B.; Kanaya, S.; Kimura, T.; Awazuhara, M.; Arita, M.; Fujiwara, T.; Saito, K. Integration of transcriptomics and metabolomics for understanding of global responses to nutritional stresses in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2004, 101, 10205–10210. [Google Scholar] [CrossRef] [Green Version]
- Yuan, G.; Wang, X.; Guo, R.; Wang, Q. Effect of salt stress on phenolic compounds, glucosinolates, myrosinase and antioxidant activity in radish sprouts. Food Chem. 2010, 121, 1014–1019. [Google Scholar] [CrossRef]
- Rodríguez-Hernández, M.D.C.; Moreno, D.A.; Carvajal, M.; Martínez-Ballesta, M.D.C. Genotype influences sulfur metabolism in broccoli (Brassica oleracea L.) under elevated CO2 and NaCl stress. Plant Cell Physiol. 2014, 55, 2047–2059. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pang, Q.; Guo, J.; Chen, S.; Chen, Y.; Zhang, L.; Fei, M.; Jin, S.; Li, M.; Wang, Y.; Yan, X. Effect of salt treatment on the glucosinolate-myrosinase system in Thellungiella salsuginea. Plant Soil 2012, 355, 363–374. [Google Scholar] [CrossRef]
- Kissen, R.; Eberl, F.; Winge, P.; Uleberg, E.; Martinussen, I.; Bones, A.M. Effect of growth temperature on glucosinolate profiles in Arabidopsis thaliana accessions. Phytochemistry 2016, 130, 106–118. [Google Scholar] [CrossRef]
- Koh, J.; Chen, G.; Yoo, M.-J.; Zhu, N.; Dufresne, D.; Erickson, J.E.; Shao, H.; Chen, S. Comparative proteomic analysis of Brassica napus in response to drought stress. J. Proteome Res. 2015, 14, 3068–3081. [Google Scholar] [CrossRef]
- Rosa, E.; Heaney, R. Seasonal variation in protein, mineral and glucosinolate composition of Portuguese cabbages and kale. Anim. Feed Sci. Technol. 1996, 57, 111–127. [Google Scholar] [CrossRef]
- Kushad, M.M.; Brown, A.F.; Kurilich, A.C.; Juvik, J.A.; Klein, B.P.; Wallig, M.A.; Jeffery, E.H. Variation of glucosinolates in vegetable crops of Brassica oleracea. J. Agric. Food Chem. 1999, 47, 1541–1548. [Google Scholar] [CrossRef]
- Park, S.; Arasu, M.V.; Lee, M.-K.; Chun, J.-H.; Seo, J.M.; Al-Dhabi, N.A.; Kim, S.-J. Analysis and metabolite profiling of glucosinolates, anthocyanins and free amino acids in inbred lines of green and red cabbage (Brassica oleracea L.). LWT-Food Sci. Technol. 2014, 58, 203–213. [Google Scholar] [CrossRef]
- Zabaras, D.; Roohani, M.; Krishnamurthy, R.; Cochet, M.; Delahunty, C.M. Characterisation of taste-active extracts from raw Brassica oleracea vegetables. Food Funct. 2013, 4, 592–601. [Google Scholar] [CrossRef]
- Fahey, J.W.; Zalcmann, A.T.; Talalay, P. The chemical diversity and distribution of glucosinolates and isothiocyanates among plants. Phytochemistry 2001, 56, 5–51. [Google Scholar] [CrossRef]
- Song, L.; Thornalley, P.J. Effect of storage, processing and cooking on glucosinolate content of Brassica vegetables. Food Chem. Toxicol. 2007, 45, 216–224. [Google Scholar] [CrossRef]
- Drewnowski, A.; Gomez-Carneros, C. Bitter taste, phytonutrients, and the consumer: A review. Am. J. Clin. Nutr. 2000, 72, 1424–1435. [Google Scholar] [CrossRef]
- Vaughn, S.F.; Berhow, M.A. Glucosinolate hydrolysis products from various plant sources: PH effects, isolation, and purification. Ind. Crop. Prod. 2005, 21, 193–202. [Google Scholar] [CrossRef]
- Rohr, F.; Ulrichs, C.; Mucha-Pelzer, T.; Mewis, I. Variability of aliphatic glucosinolates in Arabidopsis and their influence on insect resistance. Commun. Agric. Appl. Biol. Sci. 2006, 71, 507–515. [Google Scholar] [PubMed]
- Wurr, D.C.E.; Fellows, J.R.; Phelps, K. Investigating trends in vegetable crop response to increasing temperature associated with climate change. Sci. Hortic. 1996, 66, 255–263. [Google Scholar] [CrossRef]
- Gigolashvili, T.; Berger, B.; Flügge, U.-I. Specific and coordinated control of indolic and aliphatic glucosinolate biosynthesis by R2R3-MYB transcription factors in Arabidopsis thaliana. Phytochem. Rev. 2009, 8, 3–13. [Google Scholar] [CrossRef]
- Zhao, F.; Evans, E.J.; Bilsborrow, P.E.; Syers, J.K. Influence of nitrogen and sulphur on the glucosinolate profile of rapeseed (Brassica napus L). J. Sci. Food Agric. 1994, 64, 295–304. [Google Scholar] [CrossRef]
- Burow, M.; Wittstock, U. Regulation and function of specifier proteins in plants. Phytochem. Rev. 2009, 8, 87–99. [Google Scholar] [CrossRef]
- Angelino, D.; Jeffery, E. Glucosinolate hydrolysis and bioavailability of resulting isothiocyanates: Focus on glucoraphanin. J. Funct. Foods 2014, 7, 67–76. [Google Scholar] [CrossRef]
- Fenwick, G.R.; Griffiths, N.M.; Heaney, R.K. Bitterness in brussels sprouts (Brassica oleracea L. var.gemmifera): The role of glucosinolates and their breakdown products. J. Sci. Food Agric. 1983, 34, 73–80. [Google Scholar] [CrossRef]
- Traka, M.H.; Saha, S.; Huseby, S.; Kopriva, S.; Walley, P.G.; Barker, G.C.; Moore, J.; Mero, G.; van den Bosch, F.; Constant, H.; et al. Genetic regulation of glucoraphanin accumulation in Beneforté broccoli. New Phytol. 2013, 198, 1085–1095. [Google Scholar] [CrossRef] [Green Version]
- James, D.C.; Rossiter, J.T. Development and characteristics of myrosinase in Brassica napus during early seedling growth. Physiol. Plant. 1991, 82, 163–170. [Google Scholar] [CrossRef]
- Klopsch, R.; Witzel, K.; Börner, A.; Schreiner, M.; Hanschen, F.S. Metabolic profiling of glucosinolates and their hydrolysis products in a germplasm collection of Brassica rapa turnips. Food Res. Int. 2017, 100, 392–403. [Google Scholar] [CrossRef]
- Bell, L.; Kitsopanou, E.; Oloyede, O.O.; Lignou, S. Important odorants of four Brassicaceae species, and discrepancies between glucosinolate profiles and observed hydrolysis products. Foods 2021, 10, 1055. [Google Scholar] [CrossRef] [PubMed]
- Matusheski, N.V.; Swarup, R.; Juvik, J.A.; Mithen, R.; Bennett, M.; Jeffery, E.H. Epithiospecifier protein from broccoli (Brassica oleracea L. ssp. italica) inhibits formation of the anticancer agent sulforaphane. J. Agric. Food Chem. 2006, 54, 2069–2076. [Google Scholar] [CrossRef] [PubMed]
- Winkler, S.; Faragher, J.; Franz, P.; Imsic, M.; Jones, R. Glucoraphanin and flavonoid levels remain stable during simulated transport and marketing of broccoli (Brassica oleracea var. italica) heads. Postharvest Biol. Technol. 2007, 43, 89–94. [Google Scholar] [CrossRef]
- Jones, R.B.; Faragher, J.D.; Winkler, S. A review of the influence of postharvest treatments on quality and glucosinolate content in broccoli (Brassica oleracea var. italica) heads. Postharvest Biol. Technol. 2006, 41, 1–8. [Google Scholar] [CrossRef]
- van Doorn, J.E. Development of Vegetables with Improved Consumer Quality: A Case Study in Brussels Sprouts; University of Wageningen: Wageningen, The Netherlands, 1999. [Google Scholar]
- Chadwick, M.; Gawthrop, F.; Michelmore, R.W.; Wagstaff, C.; Methven, L. Perception of bitterness, sweetness and liking of different genotypes of lettuce. Food Chem. 2016, 197, 66–74. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genus/Morphotype a | Accession Name | Accession Code | Common Name | Origin | Head Formation |
|---|---|---|---|---|---|
| Black kale | |||||
| Brassica oleracea var. acephala | Cavolo nero di toscana o senza palla | BK-CNDTP (BK1) | Fodder black kale | Italy | Open leaf |
| Brassica oleracea var. acephala | Cavolo palmizio | BK-CPNT (BK2) | Black kale | Italy | Open leaf |
| Brassica oleracea var. acephala | Cavolo nero di toscana o senza testa | BK-CNDTT (BK3) | Fodder black kale | Italy | Open leaf |
| Wild | |||||
| Brassica oleracea var. oleracea | Wild cabbage | WD-8707 (WD1) | Wild cabbage | Great Britain | Open leaf |
| Brassica oleracea var. oleracea | Wild cabbage | WD-GRU (WD2) | Wild cabbage | New Zealand | Open leaf |
| Brassica oleracea var. oleracea | Wild cabbage | WD-8714 (WD3) | Wild cabbage | Great Britain | Open leaf |
| Tronchuda | |||||
| Brassica oleracea var. tronchuda | Penca mistura | TC-PCM (TC1) | Tronchuda cabbage | Portugal | Open leaf |
| Brassica oleracea var. tronchuda | Penca povoa | TC-CPDP (TC2) | Tronchuda cabbage | Portugal | Open leaf |
| Brassica oleracea var. tronchuda | Tronchuda | TC-T (TC3) | Tronchuda cabbage | Portugal | Open leaf |
| Savoy | |||||
| Brassica oleracea var. capitata | Hybrid savoy wirosa cabbage | SC-HSC (SC1) | Hybrid savoy cabbage | Great Britain | Closed heart |
| Brassica oleracea var. capitata | Pointed winter | SC-PW (SC2) | Savoy cabbage | Great Britain | Closed heart |
| Brassica oleracea var. capitata | Dark green | SC-SDG (SC3) | Savoy cabbage | Italy | Closed heart |
| Red | |||||
| Brassica oleracea var. capitata | Red langendijker | RC-RL (RC1) | Red cabbage | Great Britain | Closed heart |
| Brassica oleracea var. capitata | Rocco marner (Hybrid) | RC-RM (RC2) | Hybrid red cabbage | Great Britain | Closed heart |
| Brassica oleracea var. capitata | Red Danish | RC-RD (RC3) | Red cabbage | Netherlands | Closed heart |
| White | |||||
| Brassica oleracea var. capitata | Early market | WC-FEM (WC1) | White spring cabbage | Great Britain | Closed heart |
| Brassica oleracea var. capitata | Couve repolho | WC-CRB (WC2) | White cabbage | Portugal | Closed heart |
| Brassica oleracea var. capitata | De louviers | WC-DLI (WC3) | Hybrid white cabbage | Great Britain | Closed heart |
| Common Name | Chemical Name | Abbreviation | Mass Parent Ion | MS2 Spectrum Ion (Base Ion in Bold) a | Reference |
|---|---|---|---|---|---|
| sinigrin | 2-propenyl(allyl) GSL | SIN | 358 | 278, 275, 259, 227, 195, 180, 162 | [54,55] |
| gluconapin | 3-butenyl GSL | GPN | 372 | 292, 275, 259, 195, 194, 176 | [54,56] |
| epi/progoitrin | (R, S)-2-hydroxy-3-butenyl GSL | PROG | 388 | 332, 308, 301, 275, 259, 210, 195, 146, 136 | [54,55,56] |
| glucoiberverin | 3-(methylthio)propyl GSL | GIBVN | 406 | 326, 275, 259, 288, 228,195 | [52,54,55] |
| glucoerucin | 4-(methylthio)butyl GSL | GER | 420 | 340, 291, 275, 259, 227, 195, 178, 163 | [52,54,55] |
| glucoiberin | 3-(methylsulfinyl) propyl GSL | GIBN | 422 | 407, 358, 259 | [54,55,56] |
| glucoraphanin | 4-(methylsulfinyl) butyl GSL | GRPN | 436 | 422, 372, 291, 259, 194 | [52,54,55] |
| glucobrassicin | 3-indolylmethyl GSL | GBSN | 447 | 275, 259, 251, 205 | [54,55,56] |
| 4-hydroxyglucobrassicin | 4-hydroxy-3-indolylmethyl GSL | 4-HOH | 463 | 383, 285, 267, 259, 240, 195 | [54,55,56] |
| Precursor Glucosinolate | Glucosinolate Hydrolysis Product | Abbreviation | LRI a | ID b | MS2 Spectrum Ion (Base Ion in Bold) c | Reference | |
|---|---|---|---|---|---|---|---|
| Common name | Chemical Name | ||||||
| sinigrin | allyl thiocyanate | 2-propenyl thiocyanate | ATC | 871 | B | 99, 72, 45, 44, 41, 39 | [58] |
| allyl-ITC | 2-propenyl isothiocyanate | AITC | 884 | B | 99, 72, 71, 45, 41, 39 | [58,59] | |
| 1-cyano-2,3-epithiopropane | 3,4-epithiobutane nitrile | CETP | 1004 | B | 99, 72, 66, 59, 45, 41, 39 | [58] | |
| gluconapin | 3-butenyl-ITC | 1-butene, 4-isothiocyanate | 3BITC | 983 | B | 113, 85, 72, 64, 55, 46, 45, 41 | [58,59,60] |
| 4,5-epithiovaleronitrile | 1-cyano-3,4-epithiobutane | EVN | 1121 | B | 113, 86, 80, 73, 60, 45 | [60] | |
| progoitrin | goitrin | 5-vinyloxazolidin-2-thione | GN | 1545 | B | 129, 86, 85, 68, 57, 45, 43, 41, 39 | [61] |
| 1-cyano-2-hydroxy-3,4-epit-hiobutane isomer 1 | 2-hydroxy-3,4-epithiobutylcyanide diastereomer-1 | CHETB-1 | 1225 | B | 129, 111, 89, 84, 68, 61, 58, 55, 45 | ||
| 1-cyano-2-hydroxy-3,4-epit-hiobutane isomer 2 | 2-hydroxy-3,4-epithiobutylcyanide diastereomer-2 | CHETB-2 | 1245 | B | 129, 111, 89, 84, 68, 61, 58, 55, 45 | ||
| glucoiberverin | 4-methylthiobutyl nitrile | 4-methylthio butanenitrile | 4MBN | 1085 | B | 115, 74, 68, 61, 54, 47, 41 | [58] |
| glucoerucin | erucin | 4-(methylthio)-butyl-ITC | ER | 1427 | B | 161, 146, 115, 85, 72, 61, 55 | [58,59] |
| erucin nitrile | 1-cyano-4-(methylthio) butane | ERN | 1200 | B | 129, 87, 82, 61, 55, 48, 41, 47 | [58,59] [58] | |
| glucoiberin | iberin | 3-methylsulfinylpropyl-ITC | IB | 1617 | B | 163, 130, 116, 102, 100, 86, 72, 63, 61,41 | |
| iberin nitrile | 4-methylsulfinylbutanenitrile | IBN | 1384 | B | 131, 78, 64, 47, 41 | [58] [58] | |
| gluconasturtin | 2-phenylethyl-ITC | 2-isothiocyanatoethyl benzene | PEITC | 1458 | B | 163, 105, 91, 65, 51, 40 | |
| benzenepropanenitrile | 2-phenylethyl cyanide | BPN | 1238 | B | 131, 91, 85, 65, 63, 57, 44, 51 | [60] | |
| glucoraphanin | sulforaphane | 4-methylsulfinylbutyl-ITC | SFP | 1757 | A | 160, 114, 85, 72, 64, 63, 61, 55. 41, 39 | [57,59] |
| sulforaphane nitrile | 5-(methylsulfinyl) pentanenitrile | SFN | 1526 | B | 145, 128, 82, 64, 55, 41 | [57,59] [61] | |
| glucobrassiccin | indole-3-carbinol | 1H-indole-3-methanol | I3C | 1801 | B | 144, 145, 116, 108, 89 | |
| indoleacetonitrile | 1H-indole-3-acetonitrile | 1IAN | 1796 | B | 155, 145, 144, 130, 116, 89, 101, 63 | [62] | |
| pentyl glucosinolate | pentyl-ITC | 1-isothiocyanato-pentane | PITC | 1165 | B | 129, 114, 101, 96, 72, 55, 43, 41, 39 | [63] |
| glucotropaeolin | benzeneacetonitrile | 2-phenylacetonitrile | BAN | 1137 | A | 117, 90, 89, 77, 63, 51 | [64] |
| Cabbage Morphotype/Accession | Protein Content (mg/g ± SD) DW | Specific activity (U/mg Soluble Protein ± SD) DW | ||
|---|---|---|---|---|
| Glasshouse | Field | Glasshouse | Field | |
| Black Kale | ||||
| BK-CNDTP | 29.1 ± 0.4 gh, B | 33.7 ± 0.6 l, C | 0.5 ± 0.0 a, A | 1.3 ± 0.2 d-h, C |
| BK-CPNT | 24.5 ± 0.1 e, A | 35.4 ± 1.0 m, D | 0.5 ± 0.1 a, A | 0.9 ± 0.1 a-d, B |
| BK-CNDTT | 25.4 ± 3.9 e, A | 36.7 ± 0.7 m, E | 0.6 ± 0.1 ab, A | 1.0 ± 0.0 b-e, B |
| Wild | ||||
| WD-8707 | 27.4 ± 0.7 f, C | 31.4 ± 0.1.2 jk, E | 1.1 ± 0.1 c-f, B | 1.6 ± 0.1 ghi, C |
| WD-GRU | 25.3 ± 0.1 e, B | 29.9 ± 0.6 hi, D | 0.7 ± 0.1 abc, A | 1.7 ± 0.2 hij, C |
| WD-8714 | 18.4 ± 0.1 a, A | 30.6 ± 0.8 ij, DE | 1.3 ± 0.1 d-h, B | 2.4 ± 0.2 l, D |
| Tronchuda | ||||
| TC-PCM | 32.8 ± 0.1 kl, D | 33.6 ± 0.2 l, E | 1.2 ± 0.0 d-h, AB | 1.2 ± 0.1 d-g, AB |
| TC-CPDP | 21.2 ± 0.2 b, A | 27.8 ± 0.6 fg, B | 2.4 ± 0.1 l, C | 2.4 ± 0.3 l, C |
| TC-T | 30.5 ± 0.2 hij, C | 33.1 ± 0.8 l, DE | 1.1 ± 0.1 cde, A | 1.4 ± 0.1 e-h, B |
| Savoy | ||||
| SC-HSC | 24.5 ± 1.0 e, A | 24.6 ± 1.43 e, A | 3.7 ± 0.1 m, A | 4.7 ± 0.3 n, B |
| SC-PW | 24.1 ± 0.1 cde, A | 24.3 ± 0.3 de, A | 5.3 ± 0.1 o, BC | 6.4 ± 0.5 q, D |
| SC-SDG | dng | 24.4 ± 0.5 de, A | dng | 5.8 ± 0.7 p, CD |
| Red | ||||
| RC-RL | 21.0 ± 0.5 b, A | 33.6 ± 0.6l, C | 1.6 ± 0.1 ghi, B | 0.9 ± 0.1 a-d, A |
| RC-RM | dng | 35.4 ± 1.0 m, D | dng | 1.5 ± 0.3 f-i, B |
| RC-RD | 25.3 ± 0.1 e, B | 36.7 ± 0.7 m, E | 2.1 ± 0.0 jkl, C | 1.9 ± 0.1 ijk, C |
| White | ||||
| WC-FEM | 21.2 ± 0.9 b, A | 21.3 ± 0.4 b, A | 3.8 ± 0.2 m, C | 3.4 ± 0.2 m, B |
| WC-CRB | 22.8 ± 0.6 c, B | 23.0 ± 1.2 cd, B | 2.2 ± 0.2 kl, A | 2.1 ± 0.1 kl, A |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oloyede, O.O.; Wagstaff, C.; Methven, L. Influence of Cabbage (Brassica oleracea) Accession and Growing Conditions on Myrosinase Activity, Glucosinolates and Their Hydrolysis Products. Foods 2021, 10, 2903. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10122903
Oloyede OO, Wagstaff C, Methven L. Influence of Cabbage (Brassica oleracea) Accession and Growing Conditions on Myrosinase Activity, Glucosinolates and Their Hydrolysis Products. Foods. 2021; 10(12):2903. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10122903
Chicago/Turabian StyleOloyede, Omobolanle O., Carol Wagstaff, and Lisa Methven. 2021. "Influence of Cabbage (Brassica oleracea) Accession and Growing Conditions on Myrosinase Activity, Glucosinolates and Their Hydrolysis Products" Foods 10, no. 12: 2903. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10122903