
Development of Antioxidant and Nutritious Lentil (Lens culinaris) Flour Using Controlled Optimized Germination as a Bioprocess

 ,
,  ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Lentil Seeds
2.3. Germination Process
2.4. Experimental Design
2.5. Proximate Compositon (PC) and Fatty Acid Profile (FAP)
2.6. Determinion of Malondialdehyde (MDA)
2.7. Content of Ascorbic Acid
2.8. Content of γ-Aminobutyric Acid (GABA)
2.9. Content of Total Phenolic Compounds Determined by the Folin-Ciocalteu Method
2.10. Determination of Free and Bound Phenolic Compounds Using HPLC-Q-TOF-MS/MS and UPLC-TQD
2.11. Total Antioxidant Capacity (TAC)
2.11.1. Ferric Reducing Antioxidant Power (FRAP)
2.11.2. Oxygen Radical Absorbance Capacity (ORAC)
2.11.3. Radical Scavenging Activity (DPPH•) and Quencher Radical Scavenging Activity (Q-DPPH·)
2.11.4. Radical Cation Scavenging Activity (ABTS•+)
2.12. Glycemic Index (GI)
2.13. Colorimetric Characterization of Germinated Lentil Flours
2.14. Statistical Analysis
3. Results and Discussion
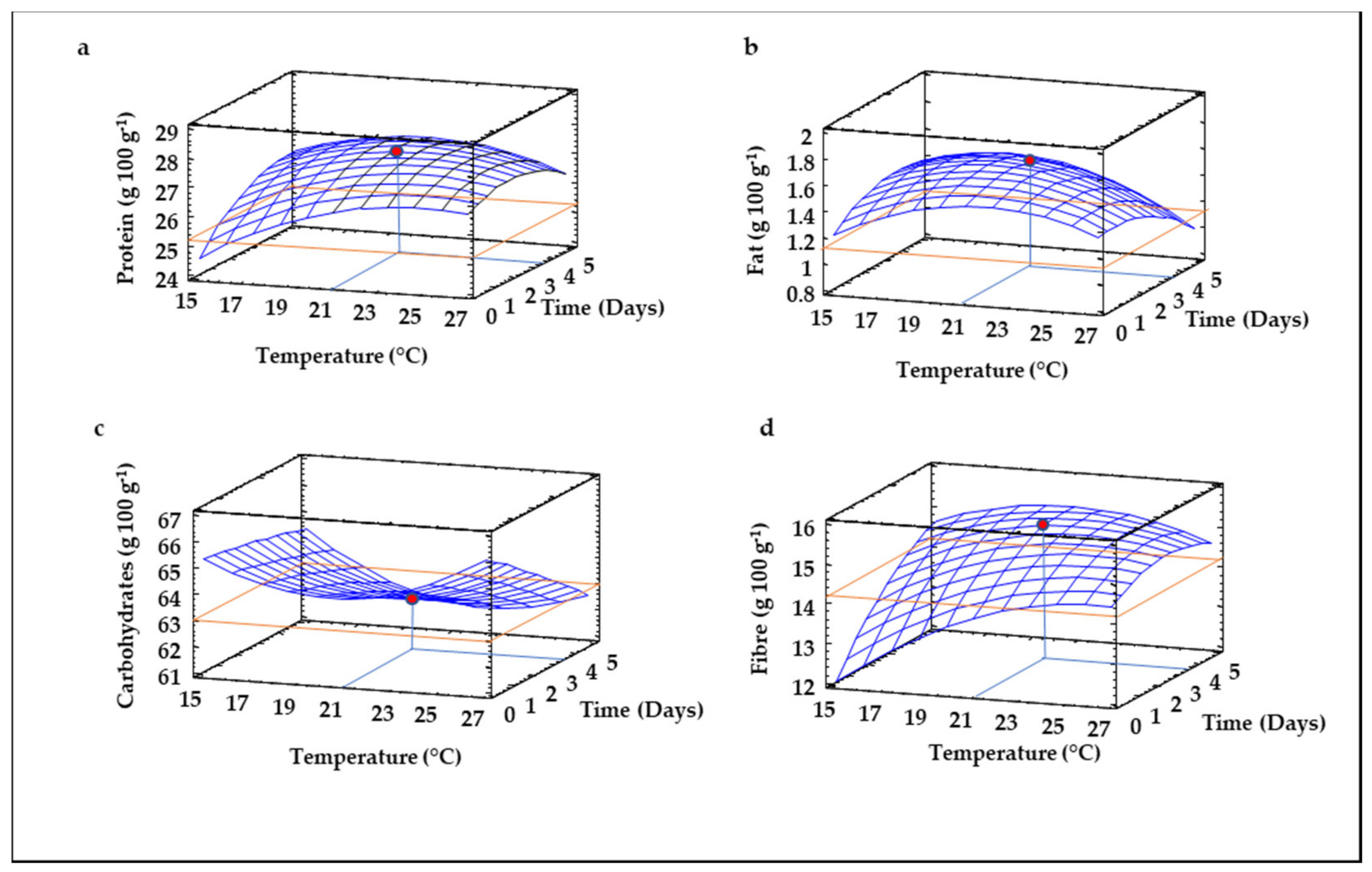
3.1. RSM Modeling of Germination Conditions and Their Impact on Sprouted Lentil Flours
3.2. Proximate Composition of Sprouted Lentil Flours
3.3. Fatty Acid of Sprouted Lentil Flours
3.4. Content of Malondialdehyde (MDA) of Sprouted Lentil Flours
3.5. Phytic Acid Content of Sprouted Lentil Flours
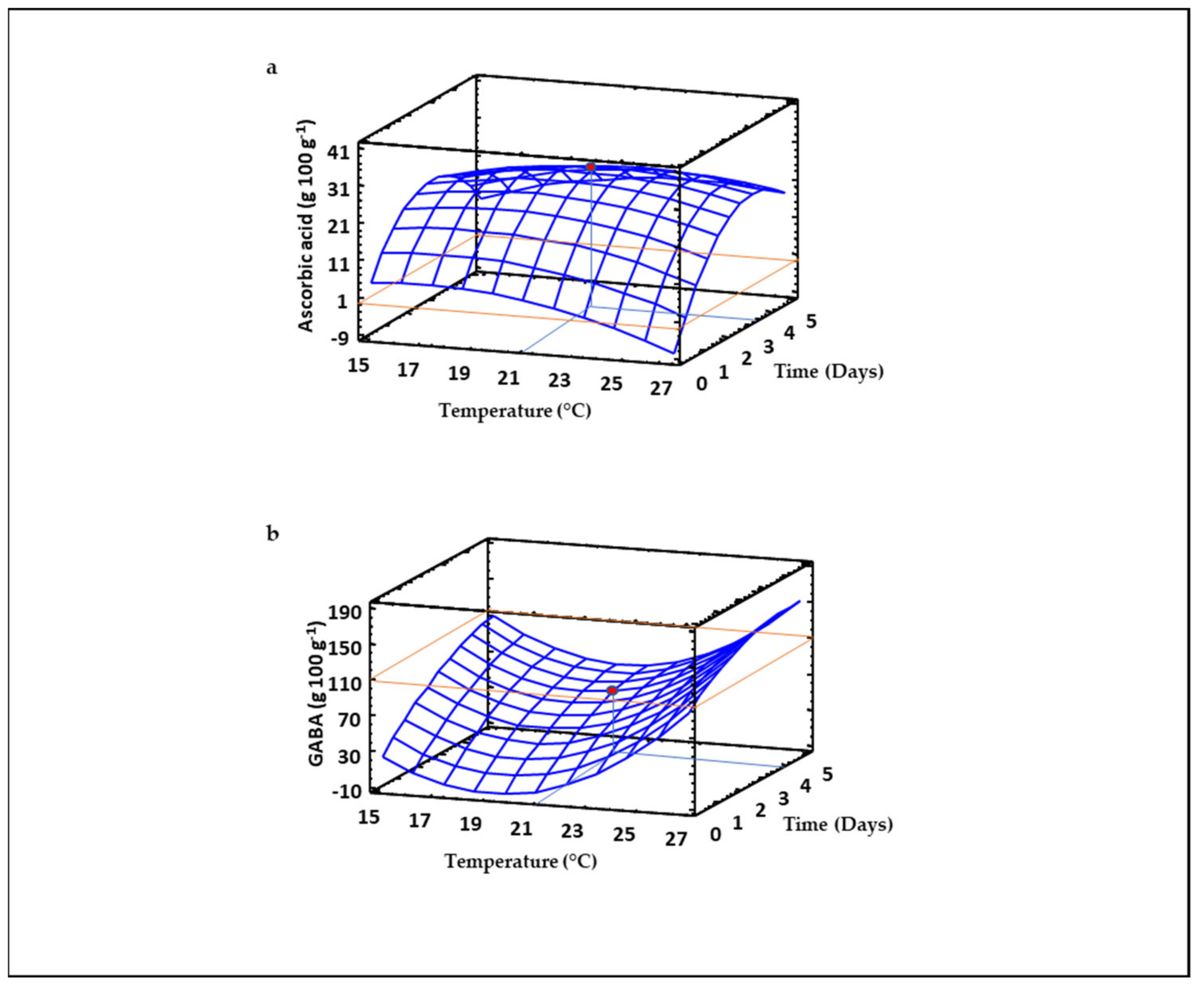
3.6. Ascorbic Acid Content of Sprouted Lentil Flours
3.7. GABA Content of Sprouted Lentil Flours
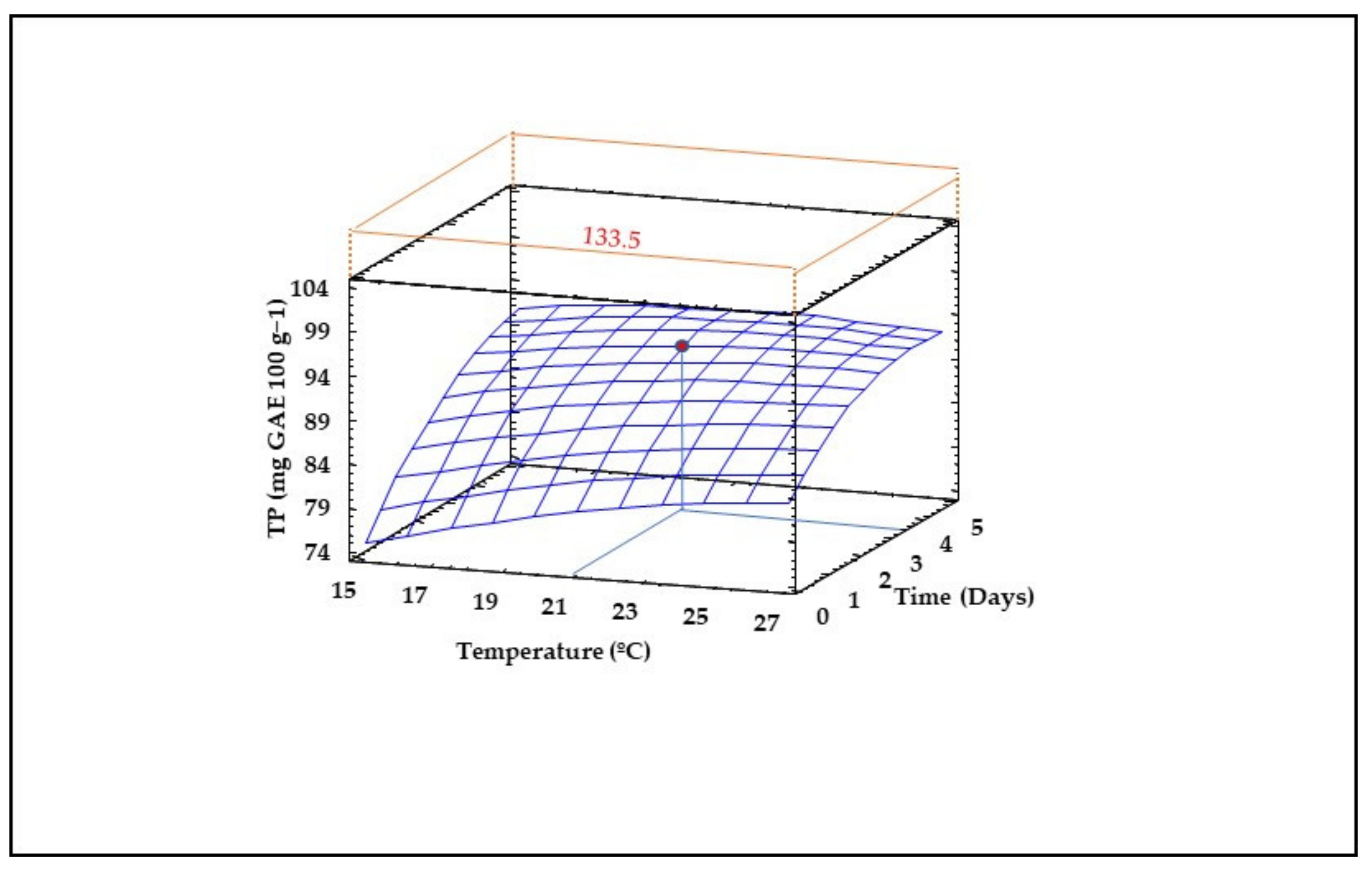
3.8. Content of Total Phenolic Compounds (TPS) in Sprouted Lentil Flours
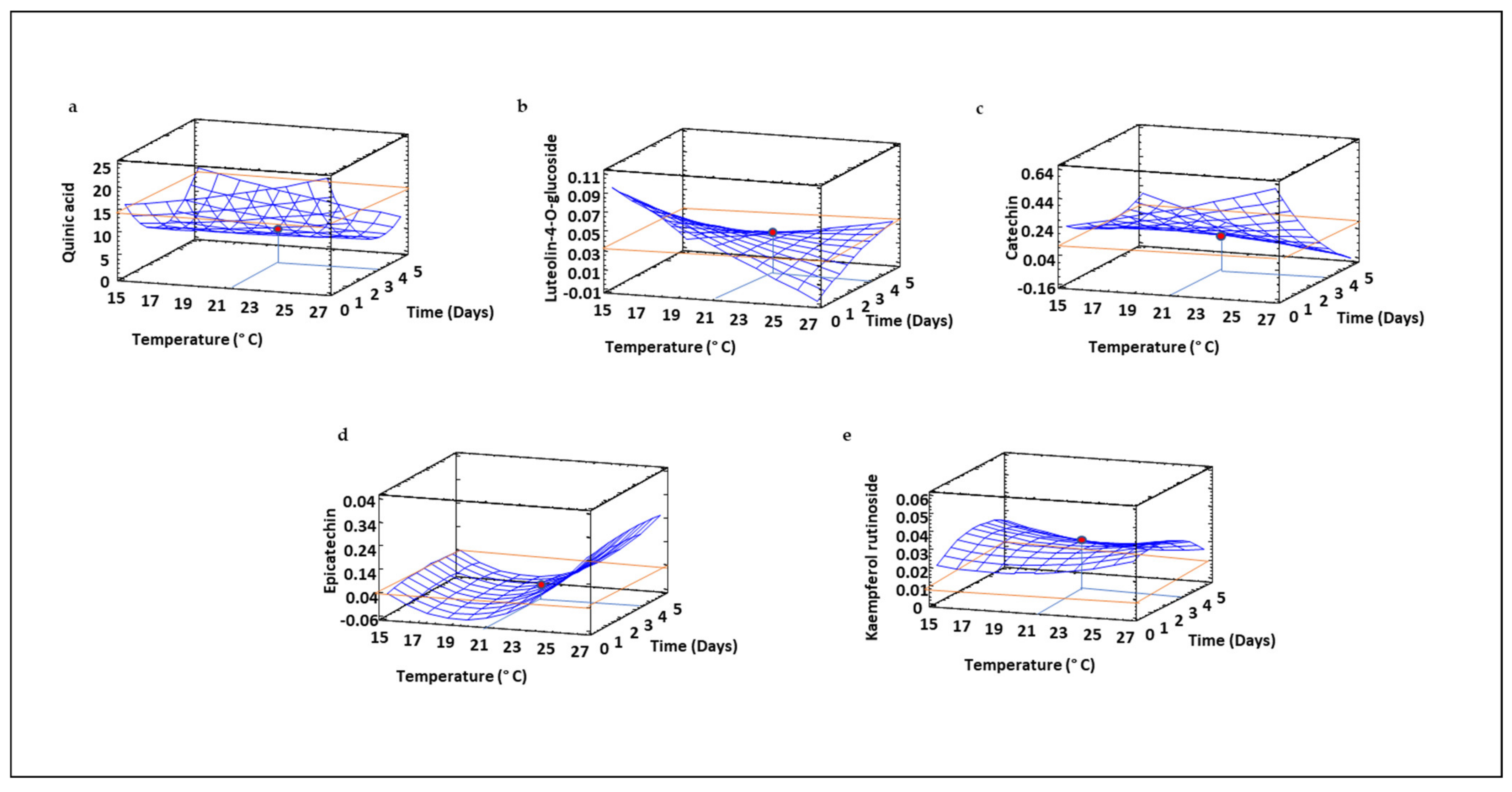
3.9. Profile of Free and Bound Phenolic Compounds in Sprouted Lentil Flours
3.10. Antioxidant Activity of Sprouted Lentil Flours
3.11. Starch Content and Glycemic Index of Sprouted Lentil Flours
3.12. Color of Sprouted Lentil Flours
3.13. RSM Optimization of Germination Conditions to Maximize the Quality of Sprouted Lentil Flours
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Teoh, S.L.; Ngorsuraches, S.; Lai, N.M.; Bangpan, M.; Chaiyakunapruk, N. Factors affecting consumers’ decisions on the use of nutraceuticals: A systematic review. Int. J. Food Sci. Nutr. 2019, 70, 491–512. [Google Scholar] [CrossRef]
- Parisi, G.F.; Carota, G.; Castruccio Castracani, C.; Spampinato, M.; Manti, S.; Papale, M.; Di Rosa, M.; Barbagallo, I.; Leonardi, S. Nutraceuticals in the prevention of viral infections, including COVID-19, among the pediatric population: A review of the literature. Int. J. Mol. Sci. 2021, 22, 2465. [Google Scholar] [CrossRef]
- Subedi, L.; Tchen, S.; Gaire, B.P.; Hu, B.; Hu, K. Adjunctive nutraceutical therapies for COVID-19. Int. J. Mol. Sci. 2021, 22, 1963. [Google Scholar] [CrossRef] [PubMed]
- Varzakas, T.; Zakynthinos, G.; Verpoort, F. Plant food residues as a source of nutraceuticals and functional foods. Foods 2016, 5, 88. [Google Scholar] [CrossRef] [Green Version]
- Mudryj, A.N.; Yu, N.; Aukema, H.M. Nutritional and health benefits of pulses. Appl. Physiol. Nutr. Metab. 2014, 39, 1197–1204. [Google Scholar] [CrossRef] [PubMed]
- United Nations. The 17 Goals—Sustainable Development Goals. Available online: https://sdgs.un.org/2030agenda (accessed on 15 November 2021).
- Joehnke, M.S.; Jeske, S.; Ispiryan, L.; Zannini, E.; Arendt, E.K.; Bez, J.; Sørensen, J.C.; Petersen, I.L. Nutritional and anti-nutritional properties of lentil (Lens culinaris) protein isolates prepared by pilot-scale processing. Food Chem. X 2021, 9, 100112. [Google Scholar] [CrossRef] [PubMed]
- Ganesan, K.; Xu, B. Polyphenol-rich lentils and their health promoting effects. Int. J. Mol. Sci. 2017, 18, 2390. [Google Scholar] [CrossRef] [Green Version]
- Zhang, B.; Deng, Z.; Ramdath, D.D.; Tang, Y.; Chen, P.X.; Liu, R.; Liu, Q.; Tsao, R. Phenolic profiles of 20 Canadian lentil cultivars and their contribution to antioxidant activity and inhibitory effects on α-glucosidase and pancreatic lipase. Food Chem. 2015, 172, 862–872. [Google Scholar] [CrossRef]
- González-Sarrías, A.; Larrosa, M.; García-Conesa, M.T.; Francisco, A.; Barberán, T.; Espín, J.C. Nutraceuticals for older people: Facts, fictions and gaps in knowledge. Maturitas 2013, 75, 313–334. [Google Scholar] [CrossRef]
- Romano, A.; Gallo, V.; Ferranti, P.; Masi, P. Lentil flour: Nutritional and technological properties, in vitro digestibility and perspectives for use in the food industry. Curr. Op. Food Sci. 2021, 40, 157–167. [Google Scholar] [CrossRef]
- Larretxi, I.; Simon, E.; Benjumea, L.; Miranda, J.; Bustamante, M.A.; Lasa, A.; Euzaguirre, F.J.; Churruca, I. Gluten-free-rendered products contribute to imbalanced diets in children and adolescents with celiac disease. Eur. J. Nutr. 2019, 58, 775–783. [Google Scholar] [CrossRef]
- Aparicio García, N.; Martínez-Villaluenga, C.; Frias, J.; Peñas, E. Sprouted oat as a potential gluten-free ingredient with enhanced nutritional and bioactive properties. Food Chem. 2021, 338, 127972. [Google Scholar] [CrossRef] [PubMed]
- Rico, D.; Peñas, E.; García, M.d.C.; Martínez-Villaluenga, C.; Rai, D.K.; Birsan, R.I.; Frias, J.; Martín-Diana, A.B. Sprouted barley flour as a nutritious and functional ingredient. Foods 2020, 9, 296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomé-Sánchez, I.; Martín-Diana, A.B.; Peñas, E.; Bautista-Expósito, S.; Frias, J.; Rico, D.; González-Maillo, L.; Martinez-Villaluenga, C. Soluble phenolic composition tailored by germination conditions accompany antioxidant and anti-Inflammatory properties of wheat. Antioxidants 2020, 9, 426. [Google Scholar] [CrossRef] [PubMed]
- Bohoua, G.L.; Yelakan, C.K. Effect of germinated sorghum flour on the performance of laying hens (Warren). Int. J. Poult. Sci. 2007, 6, 122–124. [Google Scholar] [CrossRef] [Green Version]
- Aguilera, Y.; Dueñas, M.; Estrella, I.; Hernández, T.; Benitez, V.; Esteban, R.M.; Martín-Cabrejas, M.A. Evaluation of phenolic profile and antioxidant properties of Pardina lentil as affected by industrial dehydration. J. Agric. Food Chem. 2010, 58, 10101–10108. [Google Scholar] [CrossRef]
- Pal, R.S.; Bhartiya, A.; Yadav, P.; Kant, L.; Mishra, K.K.; Aditya, J.P.; Pattanayak, A. Effect of dehulling, germination and cooking on nutrients, anti-nutrients, fatty acid composition and antioxidant properties in lentil (Lens culinaris). J. Food Sci. Technol. 2017, 54, 909–920. [Google Scholar] [CrossRef] [Green Version]
- Giami, S.Y. Effect of processing on the proximate composition and functional properties of cowpea (Vigna unguiculata) flour. Food Chem. 1993, 47, 153–158. [Google Scholar] [CrossRef]
- Box, G.E.P.; Wilson, K.G. On the experimental attainment of optimum conditions. J. Royal Stat. Soc. 1951, 13, 1–45. [Google Scholar] [CrossRef]
- AOAC. Methods 990.03, 2003.05, 985.29 & 923.03. In Official Methods of Analysis of AOAC International, 18th ed.; AOAC: Gaithersburg, MD, USA, 2005. [Google Scholar]
- Vyncke, W. Direct determination of the thiobarbituric acid value in trichloracetic acid extracts of fish as a measure of oxidative rancidity. Eur. J. Lipid Sci. Technol. 1970, 72, 1084–1087. [Google Scholar] [CrossRef]
- Cáceres, P.J.; Martínez-Villaluenga, C.; Amigo, L.; Frias, J. Maximising the phytochemical content and antioxidant activity of Ecuadorian brown rice sprouts through optimal germination conditions. Food Chem. 2014, 152, 407–414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slinkard, K.; Singleton, V.L. Total phenol analyses: Automation and comparison with manual methods. Am. J. Enol. Viticult. 1977, 28, 49–55. [Google Scholar]
- Mattila, P.; Pihlava, J.M.; Hellström, J. Contents of phenolic acids, alkyl- and alkenylresorcinols, and avenanthramides in commercial grain products. J. Agric. Food Chem. 2005, 53, 8290–8295. [Google Scholar] [CrossRef] [PubMed]
- Gangopadhyay, N.; Rai, D.K.; Brunton, N.P.; Gallagher, E.; Hossain, M.B. Antioxidant-guided isolation and mass spectrometric identification of the major polyphenols in barley (Hordeum vulgare) grain. Food Chem. 2016, 210, 212–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benzie, I.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [Green Version]
- Ou, B.; Hampsch-Woodill, M.; Prior, R.L. Development and validation of an improved oxygen radical absorbance capacity assay using fluorescein as the fluorescent probe. J. Agric. Food Chem. 2001, 49, 4619–4626. [Google Scholar] [CrossRef]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a free radical method to evaluate antioxidant activity. LWT 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Serpen, A.; Capuano, E.; Fogliano, V.; Gökmen, V. A new procedure to measure the antioxidant activity of insoluble food components. J. Agric. Food Chem. 2007, 55, 7676–7681. [Google Scholar] [CrossRef]
- Miller, N.J.; Rice-Evans, C.A. Factors influencing the antioxidant activity determined by the ABTS•+ radical cation assay. Free Radic. Res. 1997, 26, 195–199. [Google Scholar] [CrossRef]
- Martin-Diana, A.B.; Izquierdo, N.; Albertos, I.; Sanchez, M.S.; Herrero, A.; Sanz, M.A.; Rico, D. Valorization of Carob’s germ and seed peel as natural antioxidant ingredients in gluten-free crackers. J. Food Process. Preserv. 2017, 41, e12770. [Google Scholar] [CrossRef]
- Gularte, M.; Rosell, C. Physicochemical properties and enzymatic hydrolysis of different starches in the presence of hydrocolloids. Carbohydr. Polym. 2011, 85, 237–244. [Google Scholar] [CrossRef] [Green Version]
- Granfeldt, Y. Foods Factors Affecting Metabolic Responses to Cereal Products. Ph.D. Thesis, University of Lund, Lund, Sweden, 1994. [Google Scholar]
- Kaur, M.; Sandhu, K.S. Functional, thermal and pasting characteristics of flours from different lentil (Lens culinaris) cultivars. J. Food Sci. Technol. 2010, 47, 273–278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bautista-Expósito, S.; Vandenberg, A.; Peñas, E.; Frias, J.; Martínez-Villaluenga, C. Lentil and fava bean with contrasting germination kinetics: A focus on digestion of proteins and bioactivity of resistant peptides. Front. Plant. Sci. 2021, 12, 754287. [Google Scholar] [CrossRef]
- Ghumman, A.; Kaur, A.; Sing, N. Impact of germination on flour, protein and starch characteristics of lentil (Lens culinaris) and horsegram (Macrotyloma uniflorum L.) lines. LWT 2016, 65, 137–144. [Google Scholar] [CrossRef]
- Xu, M.; Jin, Z.; Simsek, S.; Hall, C.; Rao, J.; Chen, B. Effect of germination on the chemical composition, thermal, pasting, and moisture sorption properties of flours from chickpea, lentil, and yellow pea. Food Chem. 2019, 295, 579–587. [Google Scholar] [CrossRef] [PubMed]
- Moongngarm, A.; Saetung, N. Comparison of chemical compositions and bioactive compounds of germinated rough and brown rice. Food Chem. 2010, 122, 782–788. [Google Scholar] [CrossRef]
- Grasi, S.; Cardone, G.; Bigagnoli, D.; Marti, A. Monitoring the germinating process of wheat by non-conventional approaches. J. Cereal Sci. 2018, 83, 180–187. [Google Scholar] [CrossRef]
- Atudorei, D.; Stroe, S.-G.; Codină, G.G. Impact of germination on the microstructural and physicochemical properties of different legume types. Plants 2021, 10, 592. [Google Scholar] [CrossRef]
- Ghavidel, R.A.; Prakash, J. The impact of germination and dehylling on nutrients, antinutrients, in vitro iron and calcium bioavailability and in vitro starch and protein digestibility of some legume seeds. LWT 2007, 40, 1292–1299. [Google Scholar] [CrossRef]
- Olaerts, H.; Roye, C.; Derde, L.J.; Sinnaeve, G.; Meza, W.R.; Bodson, B.; Courtin C., M. Impact of preharvest sprouting of wheat (Triticum aestivum) in the field on starch, protein, and arabinoxylan properties. J. Agric. Food Chem. 2016, 64, 8324–8332. [Google Scholar] [CrossRef]
- Benítez, V.; Cantera, S.; Aguilera, Y.; Mollá, E.; Esteban, R.M.; Díaz, M.F.; Martín-Cabrejas, M.A. Impact of germination on starch, dietary fibre and physicochemical properties in non-conventional legumes. Food Res. Int. 2013, 50, 64–69. [Google Scholar] [CrossRef]
- Laxmi, G.; Chaturvedi, N.; Richa, S. The impact of malting on nutritional composition of foxtail millet, wheat and chickpea. J. Nutr. Food Sci. 2015, 5, 407. [Google Scholar]
- Dueñas, M.; Sarmento, T.; Aguilera, Y.; Benitez, V.; Mollá, E.; Esteban, R.M.; Martín-Cabrejas, M.A. Impact of cooking and germination on phenolic composition and dietary fibre fractions in dark beans (Phaseolus vulgaris L.) and lentils (Lens culinaris L.). LWT 2016, 66, 72–78. [Google Scholar] [CrossRef]
- Gunenc, A.; Yeung, M.H.; Lavergne, C.; Bertinato, J.; Hosseinian, F. Enhancements of antioxidant activity and mineral solubility of germinated wrinkled lentils during fermentation in kefir. J. Funct. Foods 2017, 32, 72–79. [Google Scholar] [CrossRef]
- Ding, J.; Hou, G.G.; Nenzer, B.V.; Xiong, S.; Dubat, A.; Feng, H. Effects of controlled germination on selected physicochemical and functional properties of whole-wheat flour and enhanced γ-aminobutyric acid accumulation by ultrasonication. Food Chem. 2018, 243, 214–221. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Yin, Y.; Liu, C.; Zhao, Z.; Guo, M. Effect of germination time on the compositional, functional and antioxidant properties of whole wheat malt and its end-use evaluation in cookie-making. Food Chem. 2021, 349, 129125. [Google Scholar] [CrossRef]
- Bewley, J.D.; Black, M. Physiology of Development and Germination. In Seeds; Bewley, J., Ed.; Springer: Boston, MA, USA, 1985; Volume 13, p. 367. [Google Scholar]
- Zhang, B.; Deng, Z.; Tang, Y.; Chen, P.; Liu, R.; Ramdath, D.D.; Liu, Q.; Hernandez, M.; Tsao, R. Fatty acid, carotenoid and tocopherol compositions of 20 Canadian lentil cultivars and synergistic contribution to antioxidant activities. Food Chem. 2014, 161, 296–304. [Google Scholar] [CrossRef]
- Montemurro, M.; Pontonio, E.; Gobbetti, M.; Rizzello, C.G. Investigation of the nutritional, functional and technological effects of the sourdough fermentation of sprouted flours. Int. J. Food Microbiol. 2019, 302, 47–58. [Google Scholar] [CrossRef]
- Bahhegyi, G.; Mandl, J. The hepatic glycogenoreticular system. Pathol. Oncol. Res. 2001, 7, 107–110. [Google Scholar] [CrossRef] [Green Version]
- Desai, A.D.; Kulkarni, S.S.; Sahoo, A.K.; Ranveer, R.C.; Dandge, P.B. Effect of supplementation of malted ragi flour on the nutritional and sensorial quality characteristics of cake. Adv. J. Food Sci. Technol. 2010, 2, 67–71. [Google Scholar]
- Xu, M.J.; Dong, J.F.; Zhu, M.Y. Effects of germination conditions on ascorbic acid level and yield of soybean sprouts. J. Sci. Food Agric. 2005, 85, 943–947. [Google Scholar] [CrossRef]
- Sholag, M.J.I.; Wei, Y.; Yang, X. Changes of folate and other potential health-promoting phytochemicals in legume seeds as affected by germination. J. Agric. Food Chem. 2012, 60, 9137–9143. [Google Scholar]
- Sahab, N.R.M.; Subroto, E.; Balia, R.; Utama, G.L. γ-aminobutyric acid found in fermented foods and beverages: Current trends. Heliyon 2020, 6, e05526. [Google Scholar] [CrossRef] [PubMed]
- Chaijan, M.; Panpinat, W. Nutritional composition and bioactivity of germinated Thai indigenous rice extracts: A feasibility study. PLoS ONE 2020, 15, e0237844. [Google Scholar] [CrossRef]
- Kuo, Y.-H.; Rozan, P.; Lambein, F.; Frias, J.; Vidal-Valverde, C. Effects of different germination conditions on the contents of free protein and non-protein amino acid of commercial legumes. Food Chem. 2004, 86, 537–545. [Google Scholar] [CrossRef]
- Oh, S.H.; Cho, W.G. Changes in the levels of gamma-aminobutyric acid and glutamate decarboxylase in developing soybean seedlings. J. Plant. Res. 2001, 114, 309–313. [Google Scholar] [CrossRef]
- Ding, L.N.; Guo, X.J.; Li, M.; Fu, Z.L.; Yan, S.Z.; Zhu, K.M.; Wang, Z.; Tan, X.L. Improving seed germination and oil contents by regulating the GDSL transcriptional level in Brassica napus. Plant. Cell Rep. 2019, 38, 243–253. [Google Scholar] [CrossRef]
- Serrano-Sandoval, S.N.; Guardado-Félix, D.; Gutiérrez-Uribe, J.A. Deglycosylation of isoflavones in selenized germinated chickpea flours due to convection drying. LWT 2022, 153, 112417. [Google Scholar] [CrossRef]
- Žilić, S.; Delić, N.; Basić, Z.; Ignjatović-Micić, D.; Janković, M.; Vančetovic, J. Effects of alkaline cooking and sprouting on bioactive compounds, their bioavailability and relation to antioxidant capacity of maize flour. J. Food Nutr. Res. 2015, 54, 155–164. [Google Scholar]
- Markham, K.; Ryan, K.G.; Gould, K.S.; Richards, G.K. Cell wall sited flavonoids in Lisianthus flower petals. Phytochemistry 2000, 54, 681–687. [Google Scholar] [CrossRef]
- Åkesson, C.; Lindgren, H.; Pero, R.W.; Leanderson, T.; Ivars, F. Quinic acid is a biologically active component of the Uncaria tomentosa extract C-Med 100®. Int. Immunopharmacol. 2005, 5, 219–229. [Google Scholar] [CrossRef] [PubMed]
- Prince, P.D.; Rodríguez-Lanci, C.; Fraga, C.G.; Galleano, M. Dietary (-)-epicatechin affects NF-κB activation and NADPH oxidases in the kidney cortex of high-fructose-fed rats. Food Funct. 2019, 10, 26–32. [Google Scholar] [CrossRef] [PubMed]
- Shulaev, V.; Corrs, D.; Miller, G.; Mittler, R. Metabolomics for plant stress response. Plant. Physiol. 2008, 132, 199–208. [Google Scholar] [CrossRef]
- Baczek-Kwinta, R.; Sala, A. What the antioxidant activity of sprouts depends on? Oxid. Commun. 2012, 35, 990–1000. [Google Scholar]
- Strack, D.; Heilemann, J.; Mömken, M.; Wray, V. Cell wall-conjutaged phenolics from coniferae leaves. Phytochem. 1988, 27, 3517–3521. [Google Scholar] [CrossRef]
- Nirmala, M.; Subba Rao, M.V.S.S.T.; Muralikrishna, G. Carbohydrates and their degrading enzymes from native and malted finger millet (Ragi, Eleusine coracana, Indaf-15). Food Chem. 2000, 69, 175–180. [Google Scholar] [CrossRef]
- Świeca, M.; Gawlik-Dziki, U. Effects of sprouting and postharvest storage under cool temperature conditions on starch content and antioxidant capacity of green pea, lentil and young mung bean sprouts. Food Chem. 2015, 185, 99–105. [Google Scholar] [CrossRef]
- Tian, B.; Xie, B.; Shi, J.; Wu, J.; CAi, Y.; Xu, T.; Xue, S.; Deng, Q. Physicochemical changes of oat seeds during germination. Food Chem. 2010, 119, 1195–1200. [Google Scholar] [CrossRef]
- Farzaneh, V.; Ghodsvali, A.; Bakhshabadi, H.; Zare, Z.; Carvalho, I.S. The impact of germination time on the some selected parameters through malting process. Int. J. Biol. Macromol. 2017, 94, 663–668. [Google Scholar] [CrossRef] [PubMed]
- Oskaybaş, B.; Özbey, A.; Kahraman, K. Effects of germination on the physicochemical and nutritional characteristics of lentil and its utilization potential in cookie-making. J. Food Meas. Charact. 2021, 15, 4245–4255. [Google Scholar] [CrossRef]
- Veronique, C. Polyphenols in foods are more complex than often thought. Am. J. Clin. Nutr. 2005, 81, 223S–229S. [Google Scholar]
- Rusydi, M.; Azrina, A. Effect of germination on total phenolic, tannin and phytic acid contents in soybean and peanut. IFR Lat. Am. 2012, 19, 673–677. [Google Scholar]
- Kumari, S.; Krishnan, V.; Sachdev, A. Impact of soaking and germination durations on antioxidants and anti-nutrients of black and yellow soybean (Glycine max. L.) varieties. J. Plant. Biochem. Biotechnol. 2014, 24, 355–358. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time (Days) | Temperature (°C) |
|---|---|
| 2 | 17 |
| 3 | 21 |
| 3 | 21 |
| 2 | 25 |
| 4 | 25 |
| 3 | 21 |
| 1 | 21 |
| 5 | 21 |
| 4 | 17 |
| 3 | 15.3 |
| 3 | 26.7 |
| Response Variables | Mathematical Models * | R2 |
|---|---|---|
| Protein | 12.83 + 1.12*T + 1.41*t − 0.02*T2 − 0.04*T*t − 0.09*t2 | 0.69 |
| Carbohydrates | 81.24 − 1.68*T + 0.52*t + 0.04*T2 − 0.04*T*t − 0.02*t2 | 0.72 |
| Fiber | 1.71 + 0.96*T + 1.49*t − 0.02*T2 − 0.04*T*t − 0.07*t2 | 0.63 |
| MDA | −53.00 + 3.66*T + 9.47*t − 0.06*T2 − 0.12*T*t − 0.87*t2 | 0.96 |
| Phytic Acid | 1.26 − 0.02*T − 0.11*t + 0.0008*T2 − 0.009*T*t + 0.03*t2 | 0.81 |
| GABA | 650.94 − 68.61*T + 38.90*t + 1.78*T2 − 0.57*T*t − 2.47*t2 | 0.67 |
| ORAC | −2370.23 + 494.28*T + 517.02*t − 11.22*T2 – 9.05*T*t | 0.55 |
| Starch | 72.11 − 2.20*T − 2.48*t + 0.05*T2 – 0.08*T*t + 0.46*t2 | 0.52 |
| GI | −4.40 + 4.72*T + 7.14*t − 0.11*T2 − 0.03*T*t − 0.55*t2 | 0.73 |
| a* | −1.62 + 0.18*T − 0.24*t − 0.004*T2 − 0.006*T*t + 0.05*t2 | 0.62 |
| b* | 24.53 − 0.91*T − 1.93*t + 0.02*T2 + 0.06*T*t + 0.12*t2 | 0.82 |
| Chroma | 24.57 − 0.92*T − 1.93*t + 0.02*T2 + 0.06*T*t + 0.12*t2 | 0.82 |
| Hue | 1.46 + 0.18*T − 2.80*t − 0.004*T2 + 0.0003*T*t + 0.38*T2 | 0.77 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rico, D.; Peñas, E.; del Carmen García, M.; Rai, D.K.; Martínez-Villaluenga, C.; Frias, J.; Martín-Diana, A.B. Development of Antioxidant and Nutritious Lentil (Lens culinaris) Flour Using Controlled Optimized Germination as a Bioprocess. Foods 2021, 10, 2924. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10122924
Rico D, Peñas E, del Carmen García M, Rai DK, Martínez-Villaluenga C, Frias J, Martín-Diana AB. Development of Antioxidant and Nutritious Lentil (Lens culinaris) Flour Using Controlled Optimized Germination as a Bioprocess. Foods. 2021; 10(12):2924. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10122924
Chicago/Turabian StyleRico, Daniel, Elena Peñas, María del Carmen García, Dilip K. Rai, Cristina Martínez-Villaluenga, Juana Frias, and Ana B. Martín-Diana. 2021. "Development of Antioxidant and Nutritious Lentil (Lens culinaris) Flour Using Controlled Optimized Germination as a Bioprocess" Foods 10, no. 12: 2924. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10122924