
The Effects of Enteral Nutrition on the Intestinal Environment in Patients in a Persistent Vegetative State

 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Clinical Study
2.2. 16S Amplicon Sequencing Analysis of Intestinal Microbiota
2.3. Metabolomic Analysis of Intestinal Metabolites by LC-TOFMS
2.3.1. Chemicals and Reagents
2.3.2. Fecal Sample Pretreatment and Standard Preparation
2.3.3. LC-TOFMS Condition
2.3.4. LC-TOFMS Measurement
2.4. Analysis of Bioinformatics and Statistical Analysis
3. Results
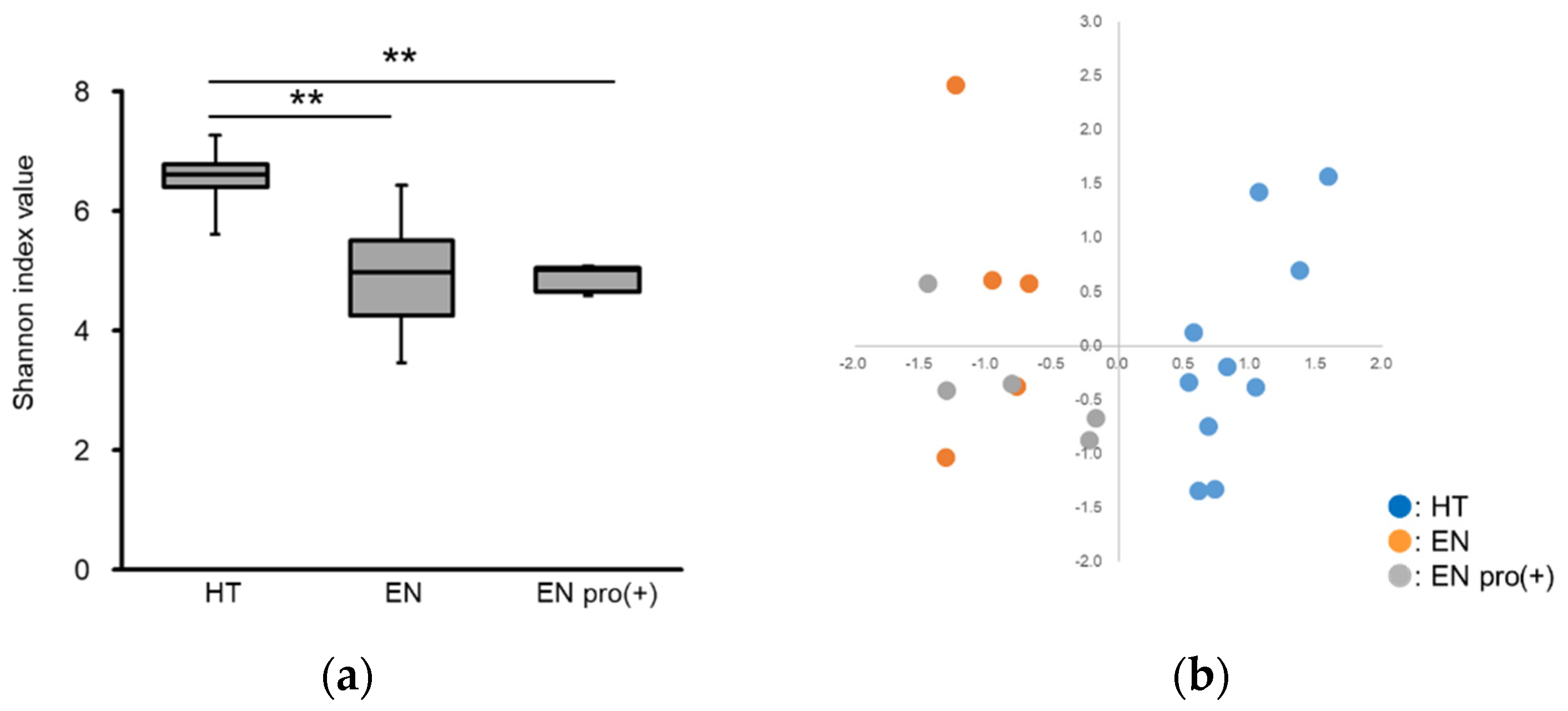
3.1. Analysis of Intestinal Microbiota
3.2. Analysis of Intestinal Metabolites
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Koh, A.; Molinaro, A.; Ståhlman, M.; Khan, M.T.; Schmidt, C.; Mannerås-Holm, L.; Wu, H.; Carreras, A.; Jeong, H.; Olofsson, L.E.; et al. Microbially produced imidazole propionate impairs insulin signaling through mTORC1. Cell 2018, 175, 947–961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feehley, T.; Plunkett, C.H.; Bao, R.; Hong, S.; Culleen, E.; Belda-Ferre, P.; Campbell, E.; Aitoro, R.; Nocerino, R.; Paparo, L.; et al. Healthy infants harbor intestinal bacteria that protect against food allergy. Nat. Med. 2019, 25, 448–453. [Google Scholar] [CrossRef] [PubMed]
- Nishiwaki, H.; Ito, M.; Ishida, T.; Hamaguchi, T.; Maeda, T.; Kashihara, K.; Tsuboi, Y.; Ueyama, J.; Shimamura, T.; Mori, H.; et al. Meta-analysis of gut dysbiosis in Parkinson’s disease. Mov. Disord. 2020, 35, 1626–1635. [Google Scholar] [CrossRef] [PubMed]
- Gramlich, L.; Hurt, R.H.; Jin, J.; Mundi, M.S. Home Enteral Nutrition: Towards a standard of care. Nutrients 2018, 10, 1020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, A.; Sowa, P.M.; Banks, M.D.; Bauer, J.D. Home Enteral Nutrition in Singapore’s Long-Term Care Homes-Incidence, Prevalence, Cost, and Staffing. Nutrients 2019, 11, 2492. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.W.; Suda, W.; Kim, S.; Oshima, K.; Fukuda, S.; Ohno, H.; Morita, H.; Hattori, M. Robustness of Gut Microbiota of Healthy Adults in Response to Probiotic Intervention Revealed by High-Throughput Pyrosequencing. DNA Res. 2013, 20, 241–253. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Kechin, A.; Boyarskikh, U.; Kel, A.; Filipenko, M. A New Tool for Accurate Cutting of Primers from Reads of Targeted Next Generation Sequencing. J. Comput. Biol. 2017, 24, 1138–1143. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [Green Version]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, 590–596. [Google Scholar] [CrossRef]
- Hippe, B.; Remely, M.; Aumueller, E.; Pointner, A.; Magnet, U.; Haslberger, A. Faecalibacterium prausnitzii phylotypes in type two diabetic, obese, and lean control subjects. Benef. Microbes 2016, 7, 511–517. [Google Scholar] [CrossRef]
- Candela, M.; Rampelli, S.; Turroni, S.; Severgnini, M.; Consolandi, C.; Bellis, G.D.; Masetti, R.; Ricci, G.; Pession, A.; Brigidi, P. Unbalance of intestinal microbiota in atopic children. BMC Microbiol. 2012, 12, 95. [Google Scholar] [CrossRef] [Green Version]
- Schwiertz, A.; Hold, G.L.; Duncan, S.H.; Gruhl, B.; Collins, M.D.; Lawson, P.A.; Flint, H.J.; Blaut, M. Anaerostipes caccae gen. nov., sp. nov., a new saccharolytic, acetate-utilising, butyrate-producing bacterium from human faeces. Syst. Appl. Microbiol. 2002, 25, 46–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Machiels, K.; Joossens, M.; Sabino, J.; Preter, V.D.; Arijs, I.; Eeckhaut, V.; Ballet, V.; Claes, K.; Immerseel, F.V.; Verbeke, K.; et al. A decrease of the butyrate-producing species Roseburia hominis and Faecalibacterium prausnitzii defines dysbiosis in patients with ulcerative colitis. Gut 2014, 63, 1275–1283. [Google Scholar] [CrossRef]
- Nishijima, S.; Suda, W.; Oshima, K.; Kim, S.W.; Hirose, Y.; Morita, H.; Hattori, M. The gut microbiome of healthy Japanese and its microbial and functional uniqueness. DNA Res. 2016, 23, 125–133. [Google Scholar] [CrossRef] [Green Version]
- Morita, N.; Umemoto, E.; Fujita, S.; Hayashi, A.; Kikuta, J.; Kimura, I.; Haneda, T.; Imai, T.; Inoue, A.; Mimuro, H.; et al. GPR31-dependent dendrite protrusion of intestinal CX3CR1 + cells by bacterial metabolites. Nature 2019, 566, 110–114. [Google Scholar] [CrossRef] [PubMed]
- Niv, Z.; Stavros, B.; Maayan, L.; Eran, E. The Role of the Immune System in Metabolic Health and Disease. Cell Metab. 2017, 25, 506–521. [Google Scholar]
- Schneider, S.; Gall, L.; Girard-Pipau, F.; Piche, T.; Pompei, A.; Nano, J.; Hébuterne, X.; Rampal, P. Total artificial nutrition is associated with major changes in the fecal flora. Eur. J. Nutr. 2000, 39, 248–255. [Google Scholar] [CrossRef] [PubMed]
- Nakanishi, S.; Kataoka, K.; Kuwahara, T.; Ohnishi, Y. Effects of High Amylose Maize Starch and Clostridium butyricum on Metabolism in Colonic Microbiota and Formation of Azoxymethane-Induced Aberrant Crypt Foci in the Rat Colon. Microbiol. Immunol. 2003, 47, 951–958. [Google Scholar] [CrossRef]
- Seki, H.; Shiohara, M.; Matsumura, T.; Miyagawa, N.; Tanaka, M.; Komiyama, A.; Kurata, S. Prevention of antibiotic-associated diarrhea in children by Clostridium butyricum MIYAIRI. Pediatr. Int. 2003, 45, 8690. [Google Scholar] [CrossRef]
- Gibson, G.R.; Roberfroid, M.B. Dietary modulation of the human colonic microbiota, introducing the concept of prebiotics. J. Nutr. 1995, 125, 1401–1412. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Intestinal Bacteria (Genus) | HT | EN | EN Pro (+) |
|---|---|---|---|
| Bacteroides | 15.66 (11.35–20.67) | 14.07 (8.09–41.49) | 31.68 (4.20–37.73) |
| Blautia | 10.95 (8.99–12.32) | 0.31 (0.00–1.76) ** | 1.52 (0.00–4.30) ** |
| Faecalibacterium | 10.37 (3.89–14.95) | 0.00 (0.00–0.00) ** | 0.00 (0.00–0.00) ** |
| Bifidobacterium | 3.34 (2.14–6.20) | 0.06 (0.00–6.36) ** | 0.05 (0.00–0.10) ** |
| Anaerostipes | 2.93 (2.28–4.75) | 0.00 (0.00–0.00) ** | 0.29 (0.00–1.16) ** |
| Agathobacter | 2.66 (0.61–7.10) | 0.00 (0.00–0.00) ** | 0.00 (0.00–0.00) ** |
| Fusicatenibacter | 2.49 (1.56–7.71) | 0.00 (0.00–0.00) ** | 0.00 (0.00–0.00) ** |
| Eubacterium hallii group | 1.63 (0.36–2.24) | 0.00 (0.00–0.07) | 0.00 (0.00–0.00) |
| Roseburia | 1.62 (1.20–4.98) | 0.00 (0.00–0.00) ** | 0.00 (0.00–0.00) ** |
| Parabacteroides | 1.59 (1.06–3.19) | 9.39 (4.35–19.89) ** | 0.37 (0.04–3.05) † |
| Unclassified | 1.35 (0.65–1.62) | 6.76 (5.39–7.48) ** | 7.89 (3.28–9.12) |
| Collinsella | 1.28 (0.45–1.45) | 0.00 (0.00–3.27) | 0.00 (0.00–0.92) |
| Lachnoclostridium | 1.02 (0.63–1.40) | 1.32 (0.92–4.77) | 2.97 (2.21–3.55) |
| Lachnospiraceae NK4A136 group | 0.99 (0.02–1.44) | 0.00 (0.00–0.00) | 0.00 (0.00–0.00) |
| Ruminococcus 2 | 0.84 (0.00–4.07) | 0.00 (0.00–0.00) | 0.00 (0.00–0.00) |
| Ruminococcus torques group | 0.74 (0.02–1.56) | 0.55 (0.44–0.69) | 0.40 (0.00–2.75) |
| Alistipes | 0.55 (0.19–0.93) | 2.62 (0.94–3.30) | 0.48 (0.41–1.60) |
| Subdoligranulum | 0.45 (0.03–3.99) | 0.00 (0.00–0.00) | 0.00 (0.00–0.00) |
| Ruminococcus gnavus group | 0.37 (0.06–1.41) | 0.71 (0.00–3.12) | 1.35 (0.00–10.30) |
| Ruminococcaceae UCG-002 | 0.13 (0.00–1.96) | 0.00 (0.00–0.00) | 0.00 (0.00–0.40) |
| Sutterella | 0.00 (0.00–0.86) | 1.31 (0.00–1.44) | 1.23 (0.00–3.84) |
| Clostridium innocuum group | 0.00 (0.00–0.03) | 2.71 (0.06–3.32) | 0.10 (0.02–0.89) |
| Intestinal Metabolites (nmol/g) | HT | EN | EN Pro (+) |
|---|---|---|---|
| Acetic acid | 252,580.13 (180,372.79–573,905.12) | 129,897.19(112,520.47–209,572.18) | 447,528.97 (250,086.40–611437.28) |
| Propionic acid | 47,302.18 (35,579.74–65,092.26) | 29,917.86 (27,741.12–37,093.41) | 95,408.42 (19,889.65–103,542.40) |
| Pyruvic acid | 1134.66 (481.72–1501.72) | 0.00 (0.00–0.00) ** | 496.94 (296.98–946.55) † |
| Isobutyric acid | 2707.42 (1519.09–4458.13) | 4050.38 (3755.59–4878.96) | 6012.13 (5238.17–14,997.66) |
| Butyric acid | 29,652.65 (15,291.35–44,766.02) | 2924.76 (521.84–6709.13) ** | 19,645.04 (15,059.10–50,424.18) † |
| Lactic acid | 239.79 (64.23–752.95) | 0.00 (0.00–906.65) | 154.32 (44.14–265.63) |
| Isovaleric acid | 1753.43 (580.36–3396.05) | 3343.35 (2626.70–6838.71) | 5457.41 (5423.17–11,377.23) |
| Valeric acid | 3168.25 (2565.22–4186.83) | 2009.47 (572.39–3964.96) | 3272.69 (2074.14–12,968.91) |
| 2-Hydroxyisobutyric acid | 2.29 (0.00–8.51) | 33.08 (30.63–60.85) ** | 0.00 (0.00–42.20) |
| 2-Methylvaleric acid | 0.00 (0.00–0.00) | 0.00 (0.00–0.00) | 0.00 (0.00–0.00) |
| 4-Methylvaleric acid | 0.00 (0.00–0.00) | 0.00 (0.00–0.00) | 0.00 (0.00–0.00) |
| Caproic acid | 0.00 (0.00–825.07) | 0.00 (0.00–715.71) | 959.72 (614.58–969.63) |
| Malonic acid | 0.00 (0.00–201.50) | 0.00 (0.00–370.86) | 1041.24 (216.11–1440.59) |
| Succinic acid | 717.54 (627.78–1113.96) | 6424.71 (707.86–38,814.07) | 6565.67 (5106.95–137,974.49) |
| LCA | 172.05 (113.95–278.99) | 211.27 (142.34–973.96) | 493.75 (205.04–832.45) |
| UDCA | 6.48 (0.36–50.22) | 1.08 (0.57–12.68) | 1.42 (1.36–76.44) |
| CDCA | 9.13 (0.47–70.97) | 8.04 (6.47–9.57) | 1.64 (0.38–37.44) |
| DCA | 201.08 (82.73–267.01) | 119.56 (29.05–146.37) | 550.86 (499.64–585.78) |
| CA | 5.65 (1.11–41.95) | 1.37 (1.30–1.88) | 5.88 (1.78–14.70) |
| GUDCA | 0.15 (0.04–0.91) | 0.01 (0.01–0.07) | 0.06 (0.01–1.39) |
| GCDCA | 1.91 (0.85–8.01) | 0.05 (0.04–0.49) | 2.11 (2.04–8.37) |
| GDCA | 0.89 (0.50–1.64) | 0,05 (0.04–0.33) | 1.12 (0.33–2.18) |
| GCA | 1.59 (0.55–4.99) | 0.04 (0.03–0.44) | 0.95 (0.31–38.20) |
| TLCA | 0.02 (0.00–0.14) | 0.02 (0.02–0.03) | 0.02 (0.01–0.11) |
| TUDCA | 0.08 (0.01–0.33) | 0.01 (0.00–0.01) | 0.01 (0.01–0.13) |
| TCDCA | 3.81 (0.27–10.73) | 0.02 (0.01–0.06) ** | 2.61 (0.99–3.02) †† |
| TDCA | 0.96 (0.13–4.79) | 0.49 (0.05–0.54) | 0.71 (0.64–0.91) |
| TCA | 1.32 (0.19–3.86) | 0.18 (0.06–0.75) | 1.56 (0.18–6.30) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Matsuoka, H.; Tochio, T.; Watanabe, A.; Funasaka, K.; Hirooka, Y.; Hartanto, T.; Togashi, Y.; Saito, M.; Nishimoto, Y.; Mizuguchi, Y.; et al. The Effects of Enteral Nutrition on the Intestinal Environment in Patients in a Persistent Vegetative State. Foods 2022, 11, 549. https://0-doi-org.brum.beds.ac.uk/10.3390/foods11040549
Matsuoka H, Tochio T, Watanabe A, Funasaka K, Hirooka Y, Hartanto T, Togashi Y, Saito M, Nishimoto Y, Mizuguchi Y, et al. The Effects of Enteral Nutrition on the Intestinal Environment in Patients in a Persistent Vegetative State. Foods. 2022; 11(4):549. https://0-doi-org.brum.beds.ac.uk/10.3390/foods11040549
Chicago/Turabian StyleMatsuoka, Hiroshi, Takumi Tochio, Ayako Watanabe, Kohei Funasaka, Yoshiki Hirooka, Tenagy Hartanto, Yuka Togashi, Misa Saito, Yuichiro Nishimoto, Yoshinori Mizuguchi, and et al. 2022. "The Effects of Enteral Nutrition on the Intestinal Environment in Patients in a Persistent Vegetative State" Foods 11, no. 4: 549. https://0-doi-org.brum.beds.ac.uk/10.3390/foods11040549