
ε-Poly-l-Lysine Enhances Fruit Disease Resistance in Postharvest Longans (Dimocarpus longan Lour.) by Modulating Energy Status and ATPase Activity

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Treatments
2.2. Measurement of Fruit Disease Index
2.3. Determination of ADP, AMP, ATP, and Energy Charge
2.4. Measurement of ATPase Activity
2.5. Statistical Analyses
3. Results
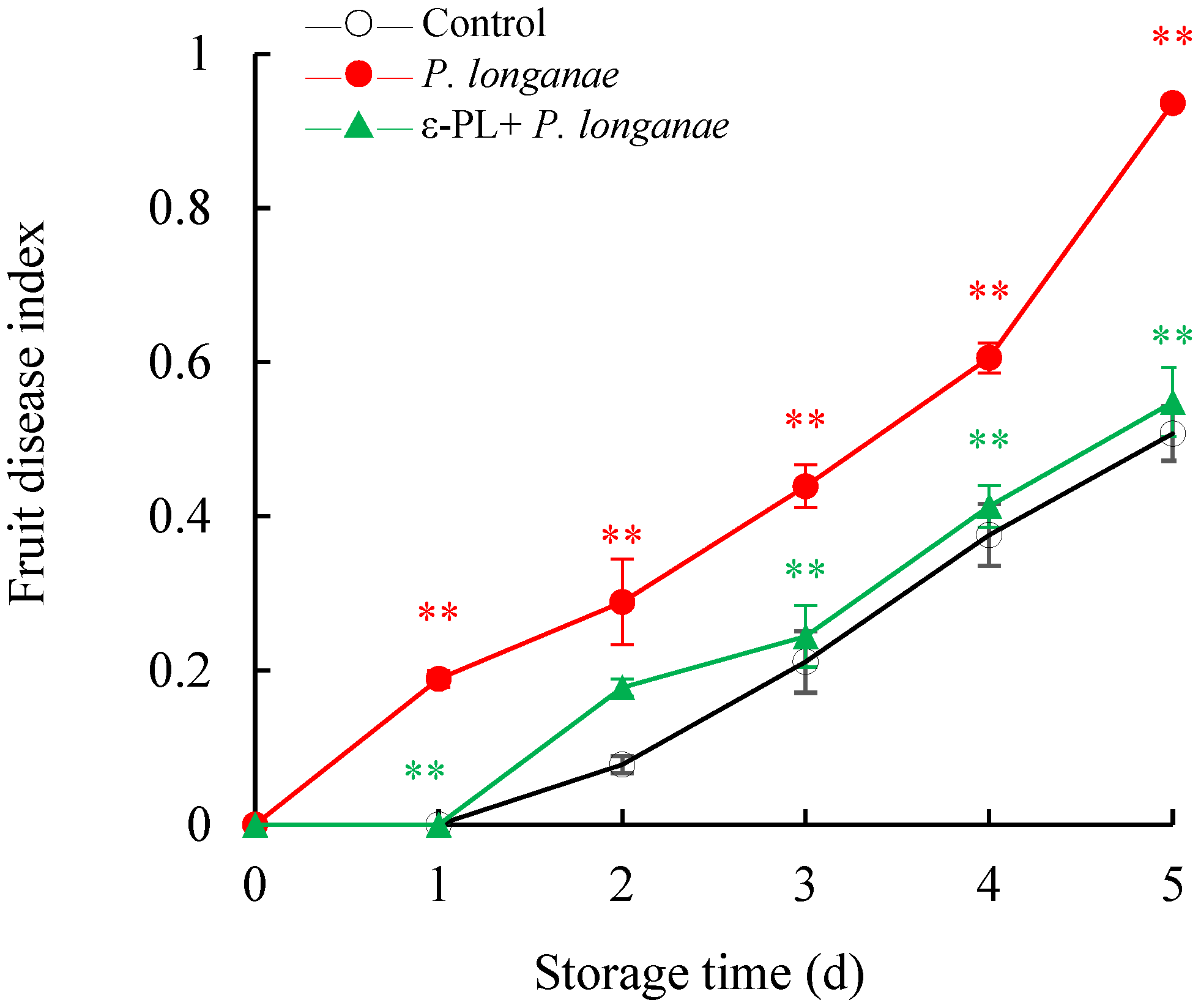
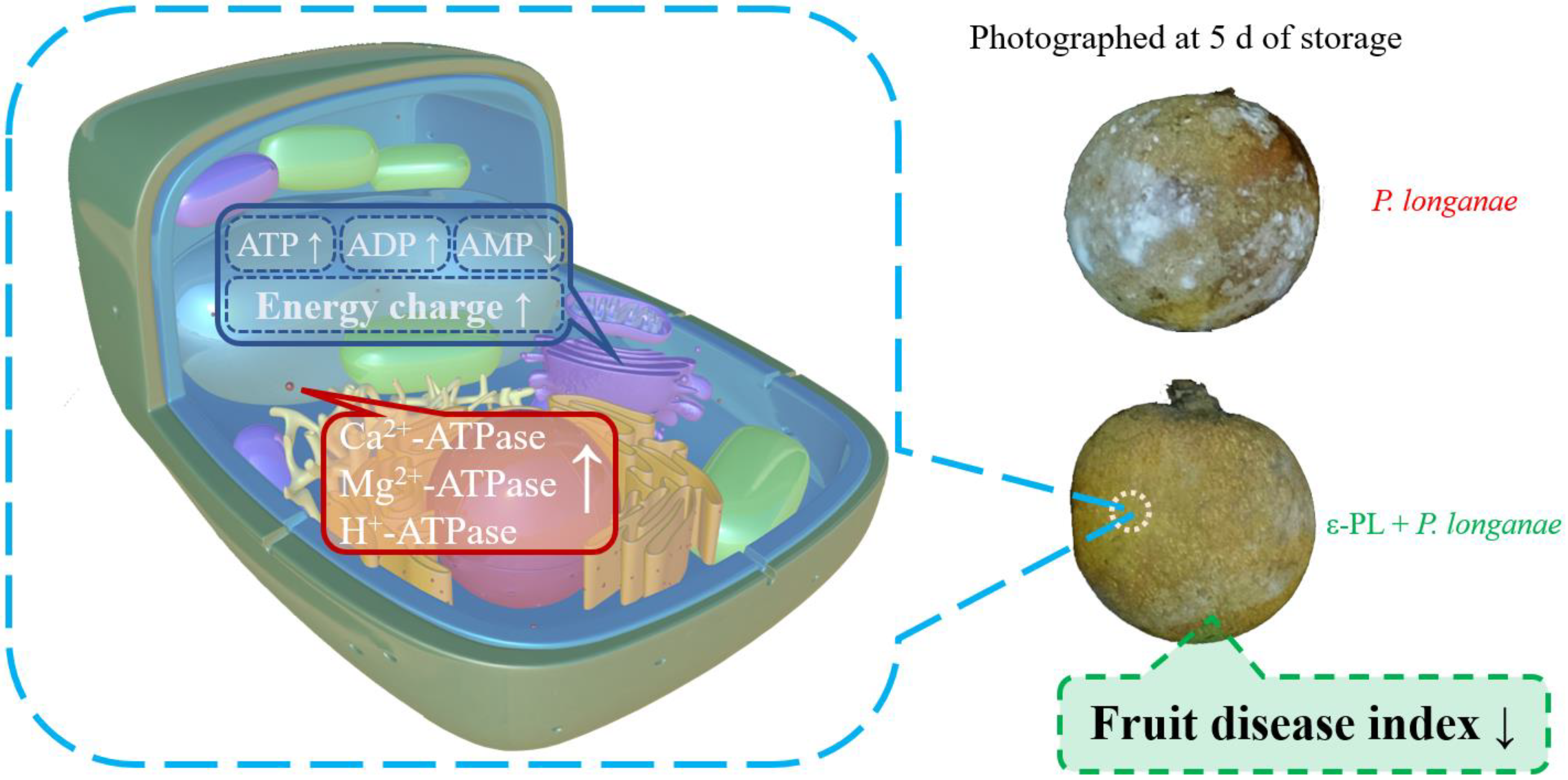
3.1. Effect of ε-PL Treatment on the Fruit Disease Index
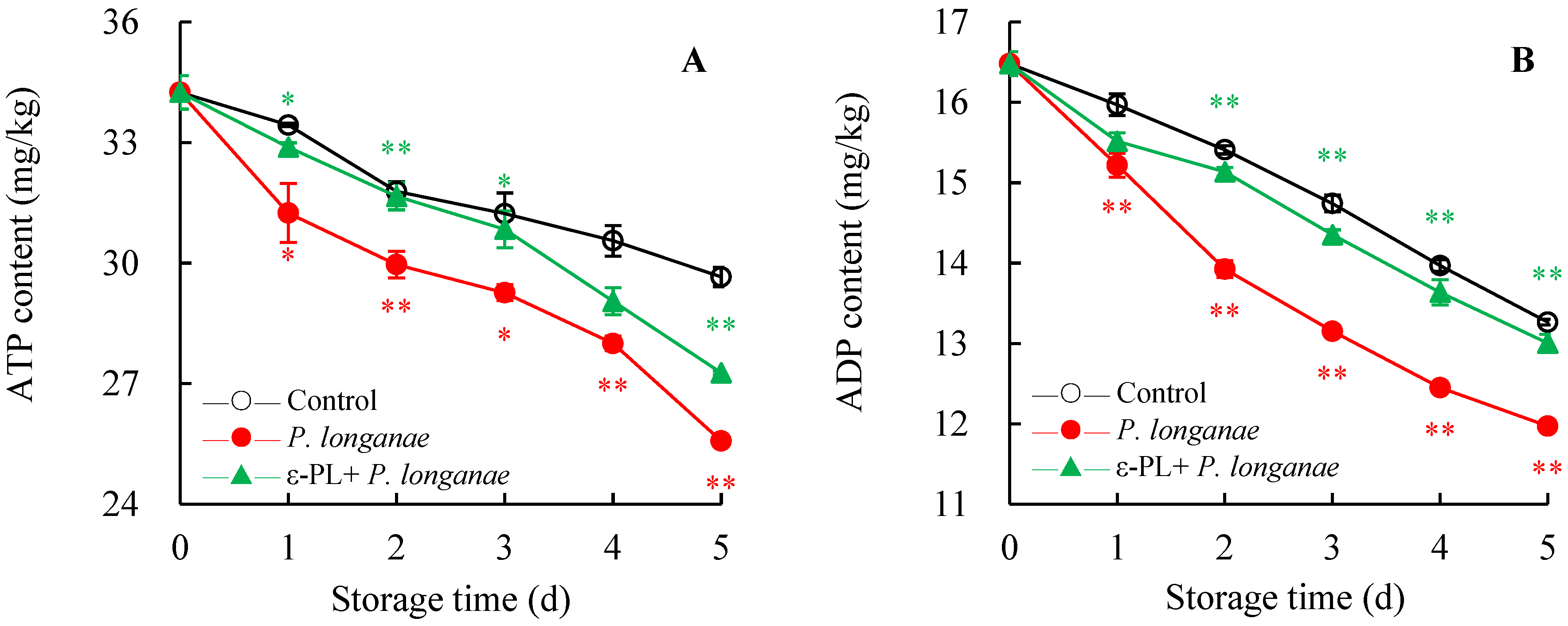
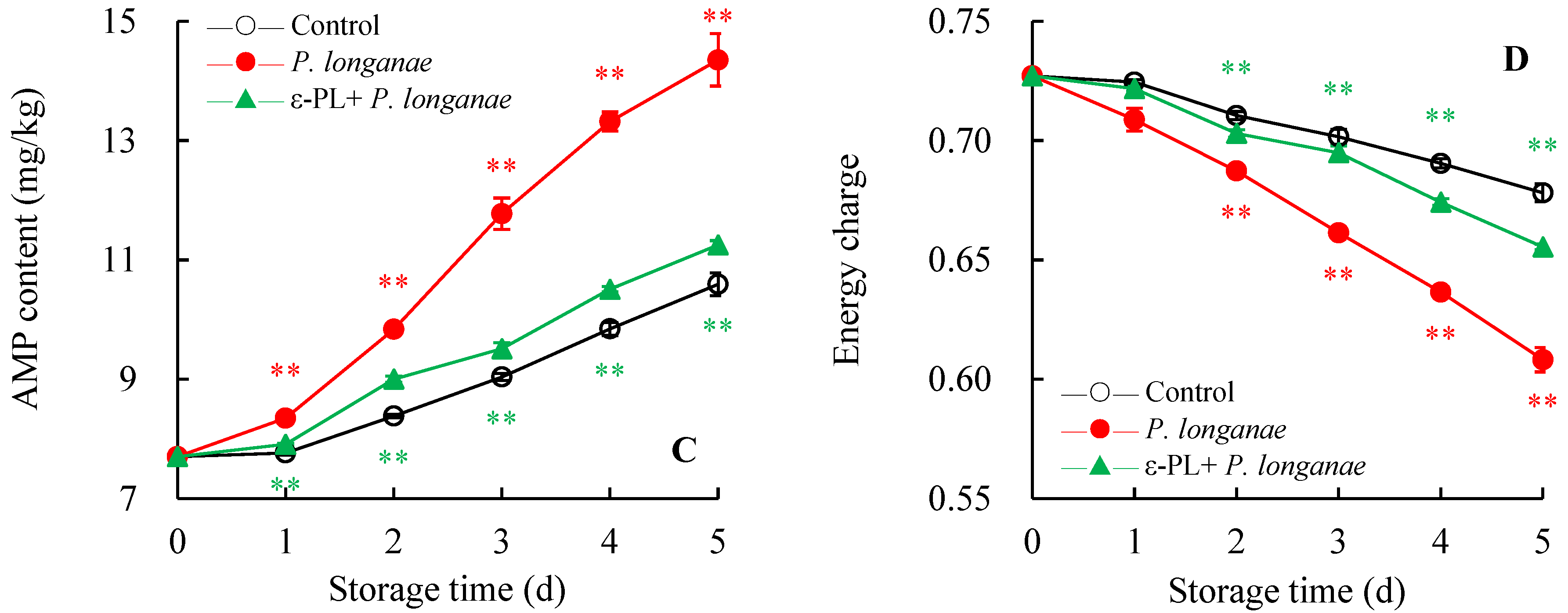
3.2. Effect of ε-PL treatment on ADP, AMP, ATP, and Energy Charge
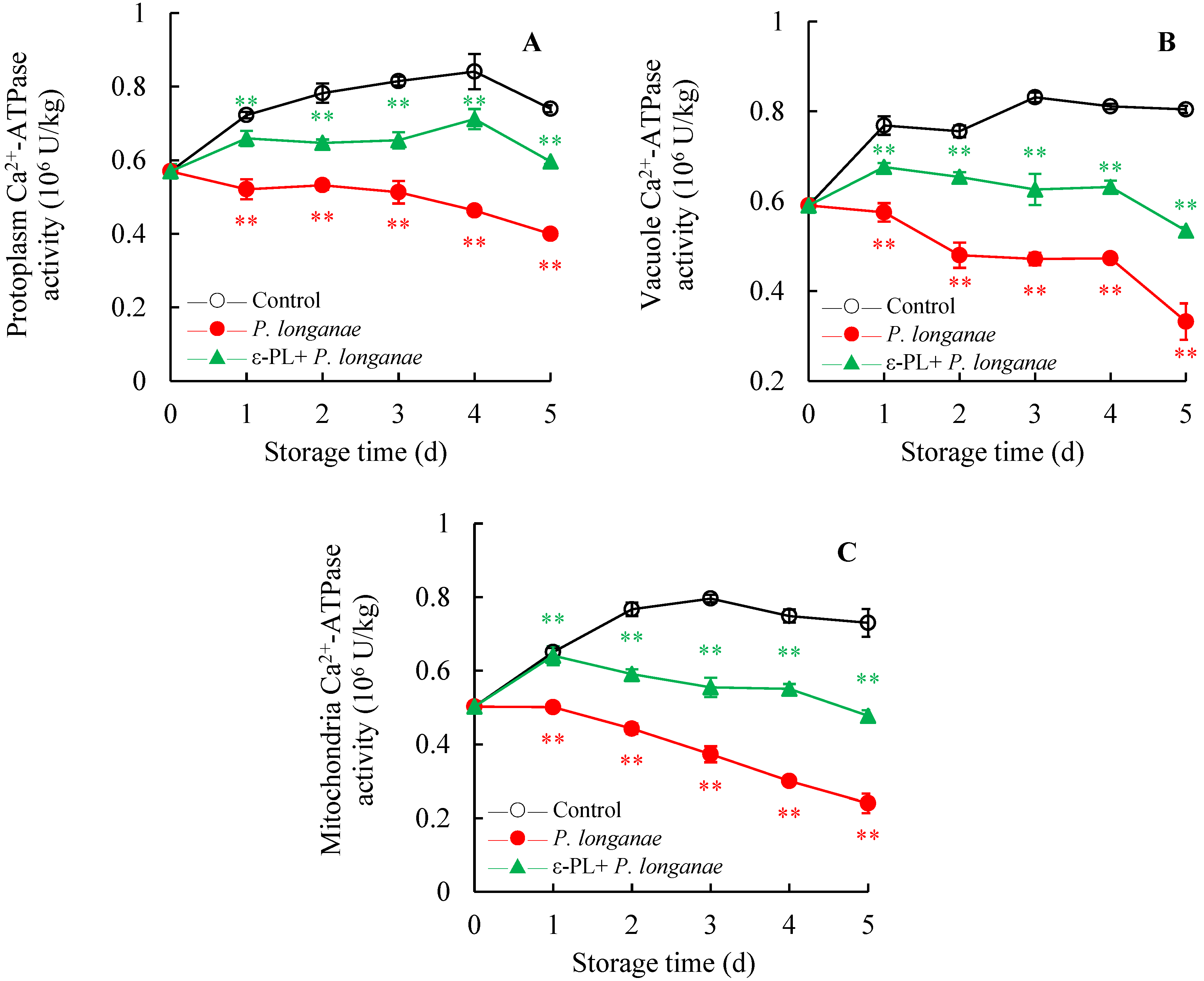
3.3. Effect of ε-PL Treatment on Ca2+-ATPase Activity
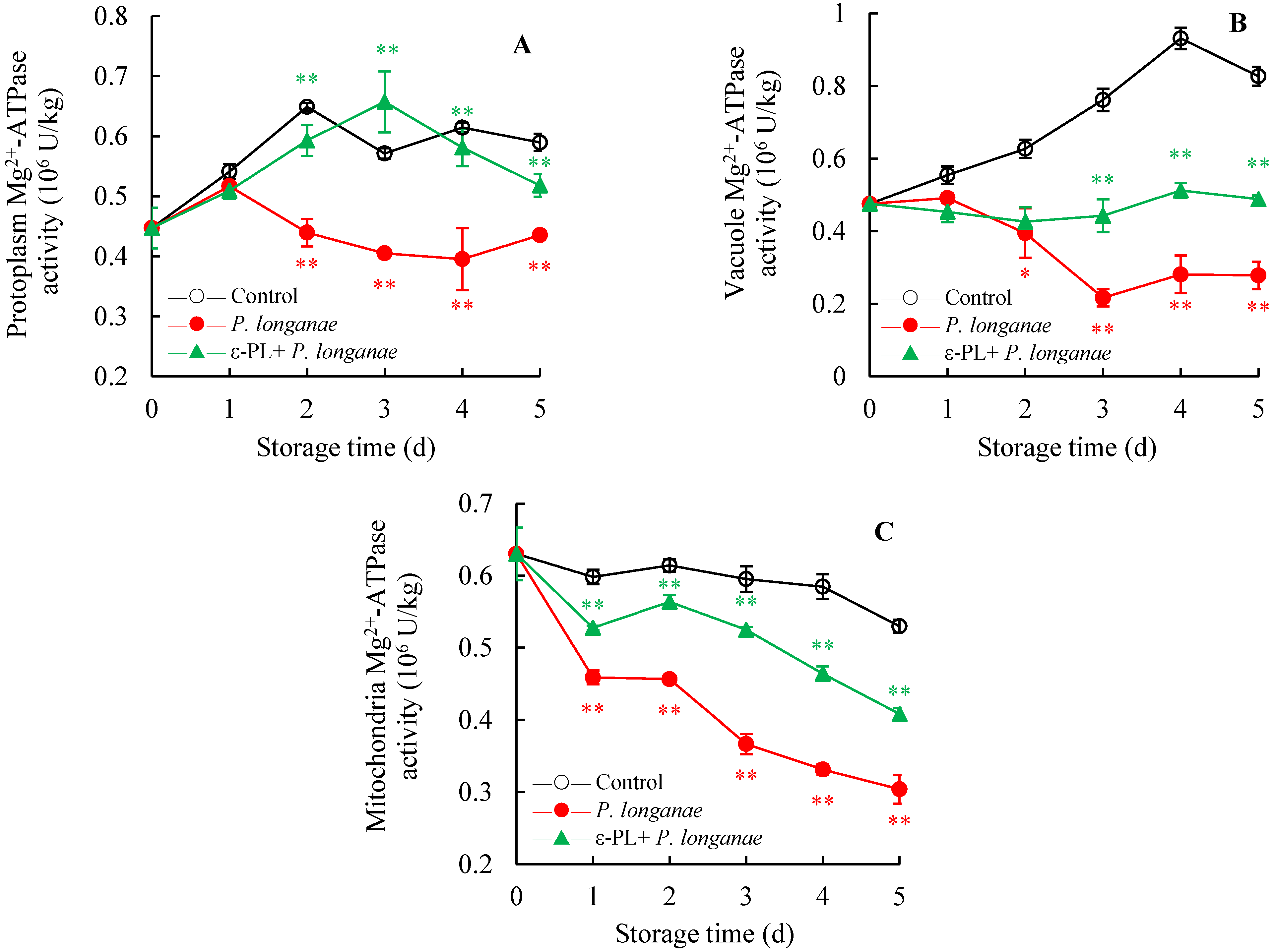
3.4. Effect of ε-PL Treatment on Mg2+-ATPase Activity
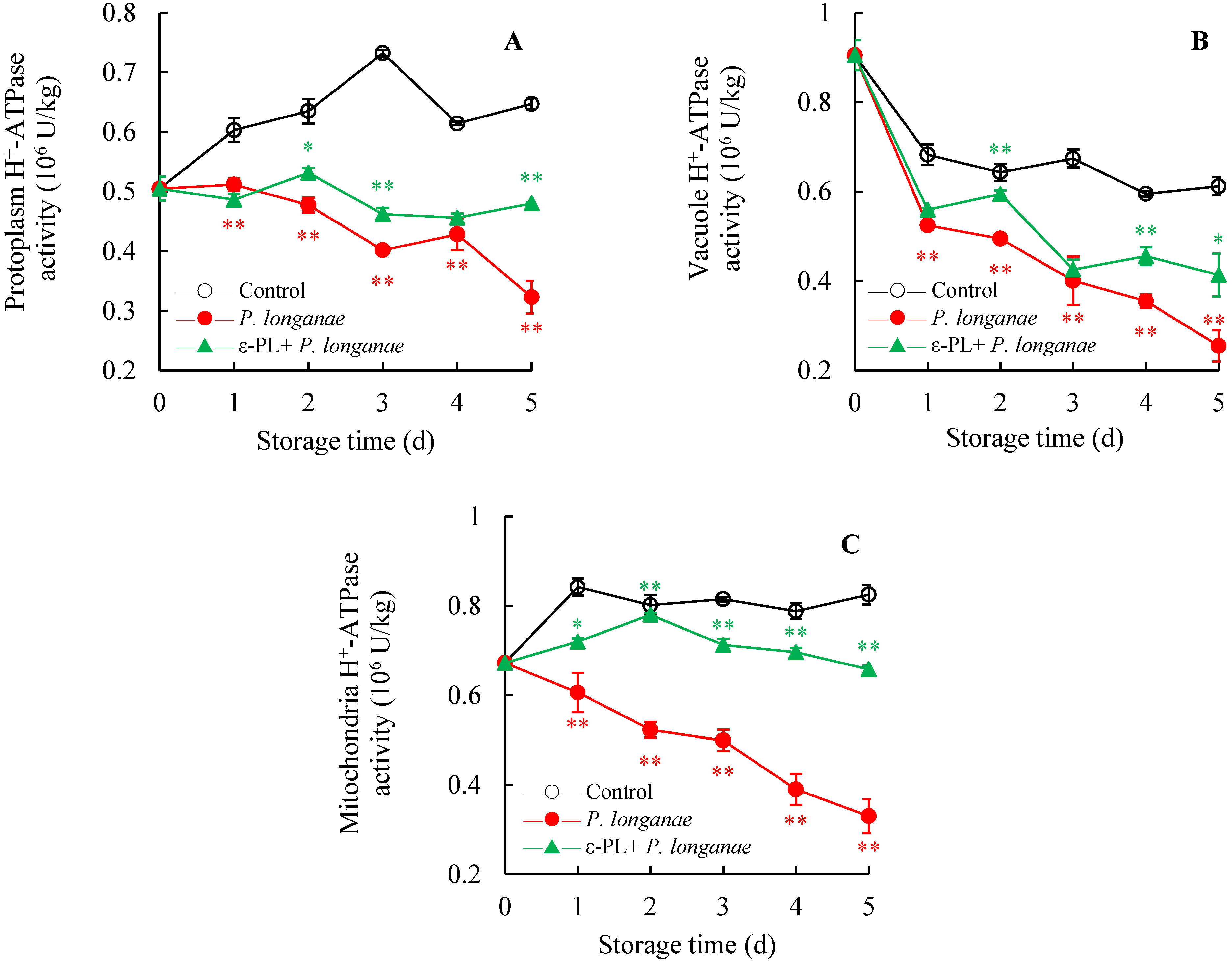
3.5. Effect of ε-PL Treatment on H+-ATPase Activity
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chen, Y.Z.; Zhang, S.; Lin, H.T.; Lu, W.J.; Wang, H.; Chen, Y.H.; Lin, Y.F.; Fan, Z.Q. The role of cell wall polysaccharides disassembly in Lasiodiplodia theobromae-induced disease occurrence and softening of fresh longan fruit. Food Chem. 2021, 351, 129294. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.Y.; Chen, H.B.; Lin, H.T.; Hung, Y.C.; Xie, H.L.; Chen, Y.H. Acidic electrolyzed water treatment delayed fruit disease development of harvested longans through inducing the disease resistance and maintaining the ROS metabolism systems. Postharvest Biol. Technol. 2021, 171, 111349. [Google Scholar] [CrossRef]
- Lin, Y.F.; Lin, Y.X.; Lin, H.T.; Ritenour, M.A.; Shi, J.; Zhang, S.; Chen, Y.H.; Wang, H. Hydrogen peroxide-induced pericarp browning of harvested longan fruit in association with energy metabolism. Food Chem. 2017, 225, 31–36. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.Z.; Lin, H.T.; Zhang, S.; Lin, Y.F.; Wang, H.; Lin, M.S.; Hung, Y.C.; Chen, Y.H. The roles of ROS production-scavenging system in Lasiodiplodia theobromae (Pat.) Griff. & Maubl.-induced pericarp browning and disease development of harvested longan fruit. Food Chem. 2018, 247, 16–22. [Google Scholar] [CrossRef]
- Somjai, C.; Siriwoharn, T.; Kulprachakarn, K.; Chaipoot, S.; Phongphisutthinant, R.; Chaiyana, W.; Srinuanpan, S.; Wiriyacharee, P. Effect of drying process and long-term storage on characterization of longan pulps and their biological aspects: Antioxidant and cholinesterase inhibition activities. LWT-Food Sci. Technol. 2022, 154, 112692. [Google Scholar] [CrossRef]
- Chen, Y.H.; Sun, J.Z.; Lin, H.T.; Lin, M.S.; Lin, Y.F.; Wang, H.; Hung, Y.C. Salicylic acid treatment suppresses Phomopsis longanae Chi-induced disease development of postharvest longan fruit by modulating membrane lipid metabolism. Postharvest Biol. Technol. 2020, 164, 111168. [Google Scholar] [CrossRef]
- Wang, H.; Chen, Y.H.; Sun, J.Z.; Lin, Y.F.; Lin, Y.X.; Lin, M.S.; Hung, Y.C.; Ritenour, M.A.; Lin, H.T. The changes in metabolisms of membrane lipids and phenolics induced by Phomopsis longanae Chi infection in association with pericarp browning and disease occurrence of postharvest longan fruit. J. Agric. Food Chem. 2018, 66, 12794–12804. [Google Scholar] [CrossRef]
- Hu, W.Y.; Zhang, X.Y.; Godana, E.A.; Gu, X.Y.; Zhao, L.N.; Zhang, H.Y. Yarrowia lipolytica reduces the disease incidence of asparagus infected by Fusarium proliferatum by affecting respiratory metabolism and energy status. Biol. Control 2021, 159, 104625. [Google Scholar] [CrossRef]
- Shi, J.Y.; Liu, N.; Gu, R.X.; Zhu, L.Q.; Zhang, C.; Wang, Q.G.; Lei, Z.H.; Liu, Y.Y.; Ren, J.Y. Signals induced by exogenous nitric oxide and their role in controlling brown rot disease caused by Monilinia fructicola in postharvest peach fruit. J. Gen. Plant Pathol. 2015, 81, 68–76. [Google Scholar] [CrossRef]
- Ge, Y.H.; Wei, M.L.; Li, C.Y.; Chen, Y.R.; Lv, J.Y.; Li, J.R. Effect of acibenzolar-S-methyl on energy metabolism and blue mould of Nanguo pear fruit. Sci. Hortic. 2017, 225, 221–225. [Google Scholar] [CrossRef]
- Lyu, L.; Bi, Y.; Li, S.G.; Xue, H.L.; Li, Y.C.; Prusky, D.B. Sodium silicate prime defense responses in harvested muskmelon by regulating mitochondrial energy metabolism and reactive oxygen species production. Food Chem. 2019, 289, 369–376. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.K.; Wang, T.; Liu, G.S.; Hu, M.J.; Yun, Z.; Duan, X.W.; Cai, K.; Jiang, G.X. Inhibition of downy blight and enhancement of resistance in litchi fruit by postharvest application of melatonin. Food Chem. 2021, 347, 129009. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.L.; Liu, H.; Bai, Y.; Yan, Y.T.; Li, L.G. Manipulation of cellular energy reveals the relationship between ultraweak luminescence and cellular energy during senescence of strawberry (Fragaria × ananassa) fruits. Acta Physiol. Plant. 2018, 40, 134. [Google Scholar] [CrossRef]
- Li, S.G.; Jiang, H.; Wang, Y.; Lyu, L.; Prusky, D.; Ji, Y.; Zheng, X.L.; Bi, Y. Effect of benzothiadiazole treatment on improving the mitochondrial energy metabolism involved in induced resistance of apple fruit during postharvest storage. Food Chem. 2020, 302, 125288. [Google Scholar] [CrossRef]
- Liu, K.W.; Zhou, X.J.; Fu, M.R. Inhibiting effects of epsilon-poly-lysine (ε-PL) on Pencillium digitatum and its involved mechanism. Postharvest Biol. Technol. 2017, 123, 94–101. [Google Scholar] [CrossRef]
- Yoshida, T.; Nagasawa, T. ε-Poly-L-lysine: Microbial production, biodegradation and application potential. Appl. Microbiol. Biotechnol. 2003, 62, 21–26. [Google Scholar] [CrossRef]
- Sun, H.T.; Shao, X.R.; Zhang, M.F.; Wang, Z.Y.; Dong, J.T.; Yu, D. Mechanical, barrier and antimicrobial properties of corn distarch phosphate/nanocrystalline cellulose films incorporated with Nisin and ε-polylysine. Int. J. Biol. Macromol. 2019, 136, 839–846. [Google Scholar] [CrossRef]
- Wu, C.H.; Sun, J.S.; Lu, Y.Z.; Wu, T.T.; Pang, J.; Hu, Y.Q. In situ self-assembly chitosan/ε-polylysine bionanocomposite film with enhanced antimicrobial properties for food packaging. Int. J. Biol. Macromol. 2019, 132, 385–392. [Google Scholar] [CrossRef]
- Hou, W.F.; Zhang, Y.; Zhang, Y.J.; Yue, Q.Q.; Wang, L.M.; Min, T.; Wang, H.X. Label-free proteomics study on Shewanella putrefaciens regulated by epsilon-poly-lysine treatment. J. Appl. Microbiol. 2021, 131, 791–800. [Google Scholar] [CrossRef]
- Cai, L.Y.; Cao, A.L.; Bai, F.L.; Li, J.R. Effect of ε-polylysine in combination with alginate coating treatment on physicochemical and microbial characteristics of Japanese sea bass (Lateolabrax japonicas) during refrigerated storage. LWT-Food Sci. Technol. 2015, 62, 1053–1059. [Google Scholar] [CrossRef]
- Lin, L.; Gu, Y.L.; Li, C.Z.; Vittayapadung, S.; Cui, H.Y. Antibacterial mechanism of ε-Poly-lysine against Listeria monocytogenes and its application on cheese. Food Control 2018, 91, 76–84. [Google Scholar] [CrossRef]
- Hou, Y.; Wang, F.P.; Tan, Z.L.; Cui, J.D.; Jia, S.R. Antifungal mechanisms of ε-poly-L-Lysine with different molecular weights on Saccharomyces cerevisiae. Korean J. Chem. Eng. 2020, 37, 482–492. [Google Scholar] [CrossRef]
- Jiao, W.X.; Liu, X.; Chen, Q.M.; Du, Y.M.; Li, Y.Y.; Yue, F.L.; Dong, X.Q.; Fu, M.R. Epsilon-poly-l-lysine (ε-PL) exhibits antifungal activity in vivo and in vitro against Botrytis cinerea and mechanism involved. Postharvest Biol. Technol. 2020, 168, 111270. [Google Scholar] [CrossRef]
- Ning, H.Q.; Li, Y.Q.; Lin, H.; Wang, J.X. Apoptosis-induction effect of ε-poly-lysine against Staphylococcus aureus and its application on pasteurized milk. LWT-Food Sci. Technol. 2021, 137, 110493. [Google Scholar] [CrossRef]
- Shu, C.; Cui, K.B.; Li, Q.Q.; Cao, J.K.; Jiang, W.B. Epsilon-poly-l-lysine (epsilon-PL) exhibits multifaceted antifungal mechanisms of action that control postharvest Alternaria rot. Int. J. Food Microbiol. 2021, 348, 109224. [Google Scholar] [CrossRef]
- Li, H.; He, C.; Li, G.J.; Zhang, Z.Q.; Li, B.Q.; Tian, S.P. The modes of action of epsilon-polylysine (ε-PL) against Botrytis cinerea in jujube fruit. Postharvest Biol. Technol. 2019, 147, 1–9. [Google Scholar] [CrossRef]
- Dou, Y.; Routledge, M.N.; Gong, Y.Y.; Godana, E.A.; Dhanasekaran, S.; Yang, Q.Y.; Zhang, X.Y.; Zhang, H.Y. Efficacy of epsilon-poly-L-lysine inhibition of postharvest blue mold in apples and potential mechanisms. Postharvest Biol. Technol. 2021, 171, 111346. [Google Scholar] [CrossRef]
- Ge, Y.H.; Wei, M.L.; Li, C.Y.; Chen, Y.R.; Lv, J.Y.; Meng, K.; Wang, W.H.; Li, J.R. Reactive oxygen species metabolism and phenylpropanoid pathway involved in disease resistance against Penicillium expansum in apple fruit induced by ε-poly-l-lysine. J. Sci. Food Agric. 2018, 98, 5082–5088. [Google Scholar] [CrossRef]
- Chen, Y.H.; Lin, H.T.; Jiang, Y.M.; Zhang, S.; Lin, Y.F.; Wang, Z.H. Phomopsis longanae Chi-induced pericarp browning and disease development of harvested longan fruit in association with energy status. Postharvest Biol. Technol. 2014, 93, 24–28. [Google Scholar] [CrossRef]
- Chen, Y.H.; Lin, H.T.; Zhang, S.; Sun, J.Z.; Lin, Y.F.; Wang, H.; Lin, M.S.; Shi, J. Phomopsis longanae Chi-induced disease development and pericarp browning of harvested longan fruit in association with energy metabolism. Food Microbiol. 2018, 9, 1454. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Lin, H.T.; Lin, M.S.; Lin, Y.F.; Chen, Y.H.; Wang, H.; Lin, Y.X.; Shi, J. Lasiodiplodia theobromae (Pat.) Griff. & Maubl. reduced energy status and ATPase activity and its relation to disease development and pericarp browning of harvested longan fruit. Food Chem. 2019, 275, 239–245. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.G.; Yuan, M.Q.; Zhang, W.M.; Zhang, Z.K. Effect of low temperatures on chilling injury in relation to energy status in papaya fruit during storage. Postharvest Biol. Technol. 2017, 125, 181–187. [Google Scholar] [CrossRef]
- Tang, R.F.; Zhou, Y.J.; Chen, Z.S.Z.; Wang, L.; Lai, Y.K.; Chang, S.K.; Wang, Y.F.; Qu, H.X.; Jiang, Y.M.; Huang, H. Regulation of browning and senescence of litchi fruit mediated by phenolics and energy status: A postharvest comparison on three different cultivars. Postharvest Biol. Technol. 2020, 168, 111280. [Google Scholar] [CrossRef]
- Chen, M.Y.; Lin, H.T.; Zhang, S.; Lin, Y.F.; Chen, Y.H.; Lin, Y.X. Effects of adenosine triphosphate (ATP) treatment on postharvest physiology, quality and storage behavior of longan fruit. Food Bioprocess Technol. 2015, 8, 971–982. [Google Scholar] [CrossRef]
- Zhang, S.; Lin, H.T.; Lin, Y.F.; Lin, Y.X.; Hung, Y.C.; Chen, Y.H.; Wang, H.; Shi, J. Energy status regulates disease development and respiratory metabolism of Lasiodiplodia theobromae (Pat.) Griff. & Maubl.-infected longan fruit. Food Chem. 2017, 231, 238–246. [Google Scholar] [CrossRef]
- Jin, P.; Zhu, H.; Wang, J.; Chen, J.J.; Wang, X.L.; Zheng, Y.H. Effect of methyl jasmonate on energy metabolism in peach fruit during chilling stress. J. Sci. Food Agric. 2013, 93, 1827–1832. [Google Scholar] [CrossRef]
- Lin, Y.F.; Lin, Y.X.; Lin, H.T.; Chen, Y.H.; Wang, H.; Shi, J. Application of propyl gallate alleviates pericarp browning in harvested longan fruit by modulating metabolisms of respiration and energy. Food Chem. 2018, 240, 863–869. [Google Scholar] [CrossRef]
- Vichaiya, T.; Uthaibutra, J.; Saengnil, K. Gaseous chlorine dioxide increases energy status and energy metabolism related enzyme activities leading to reduction in pericarp browning of longan fruit during storage. Sci. Hortic. 2020, 263, 109118. [Google Scholar] [CrossRef]
- Liu, Z.L.; Li, L.; Luo, Z.S.; Zeng, F.F.; Jiang, L.; Tang, K.C. Effect of brassinolide on energy status and proline metabolism in postharvest bamboo shoot during chilling stress. Postharvest Biol. Technol. 2016, 111, 240–246. [Google Scholar] [CrossRef]
- Elmore, J.M.; Coaker, G. The role of the plasma membrane H+-ATPase in plant-microbe interactions. Mol. Plant 2011, 4, 416–427. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.F.; Chen, M.Y.; Lin, H.T.; Lin, M.S.; Hung, Y.C.; Lin, Y.X.; Chen, Y.H.; Wang, H.; Ritenour, M.A. Phomopsis longanae-induced pericarp browning and disease development of longan fruit can be alleviated or aggravated by regulation of ATP-mediated membrane lipid metabolism. Food Chem. 2018, 269, 644–651. [Google Scholar] [CrossRef] [PubMed]
- Yi, C.; Qu, H.X.; Jiang, Y.M.; Shi, J.; Duan, X.W.; Joyce, D.C.; Li, Y.B. ATP-induced changes in energy status and membrane integrity of harvested litchi fruit and its relation to pathogen resistance. J. Phytopathol. 2008, 156, 365–371. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, J.; Chen, H.; Chen, Y.; Lin, M.; Hung, Y.-C.; Jiang, Y.; Lin, H. ε-Poly-l-Lysine Enhances Fruit Disease Resistance in Postharvest Longans (Dimocarpus longan Lour.) by Modulating Energy Status and ATPase Activity. Foods 2022, 11, 773. https://0-doi-org.brum.beds.ac.uk/10.3390/foods11050773
Sun J, Chen H, Chen Y, Lin M, Hung Y-C, Jiang Y, Lin H. ε-Poly-l-Lysine Enhances Fruit Disease Resistance in Postharvest Longans (Dimocarpus longan Lour.) by Modulating Energy Status and ATPase Activity. Foods. 2022; 11(5):773. https://0-doi-org.brum.beds.ac.uk/10.3390/foods11050773
Chicago/Turabian StyleSun, Junzheng, Hongbin Chen, Yihui Chen, Mengshi Lin, Yen-Con Hung, Yuji Jiang, and Hetong Lin. 2022. "ε-Poly-l-Lysine Enhances Fruit Disease Resistance in Postharvest Longans (Dimocarpus longan Lour.) by Modulating Energy Status and ATPase Activity" Foods 11, no. 5: 773. https://0-doi-org.brum.beds.ac.uk/10.3390/foods11050773