
Lacticaseibacillus rhamnosus: A Suitable Candidate for the Construction of Novel Bioengineered Probiotic Strains for Targeted Pathogen Control


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
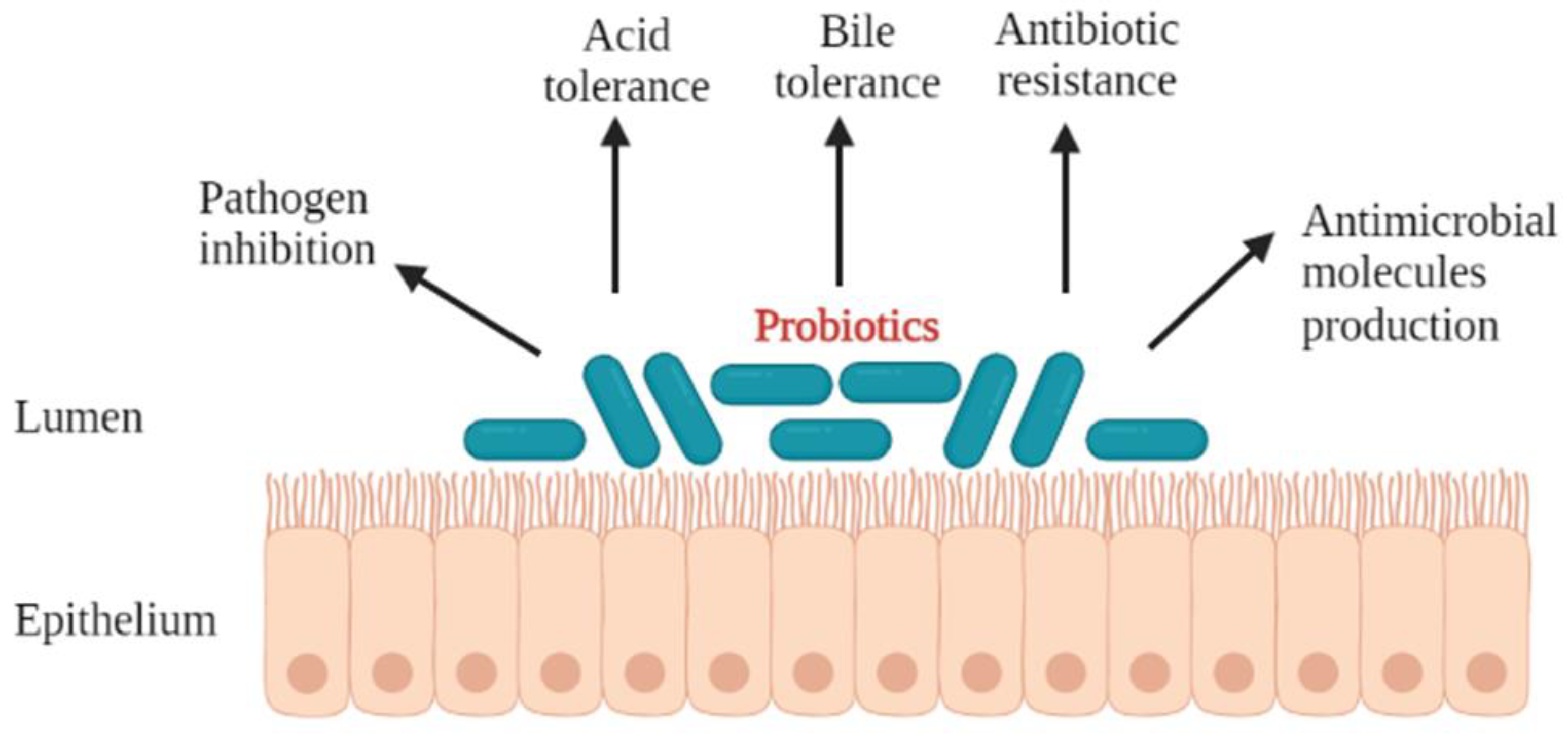
2. Desirable Properties of Probiotics
3. How Well Does L. rhamnosus Fulfil the Probiotic Selection Criteria?
3.1. Ability to Endure Gastrointestinal Stresses, Acid and Bile Tolerance
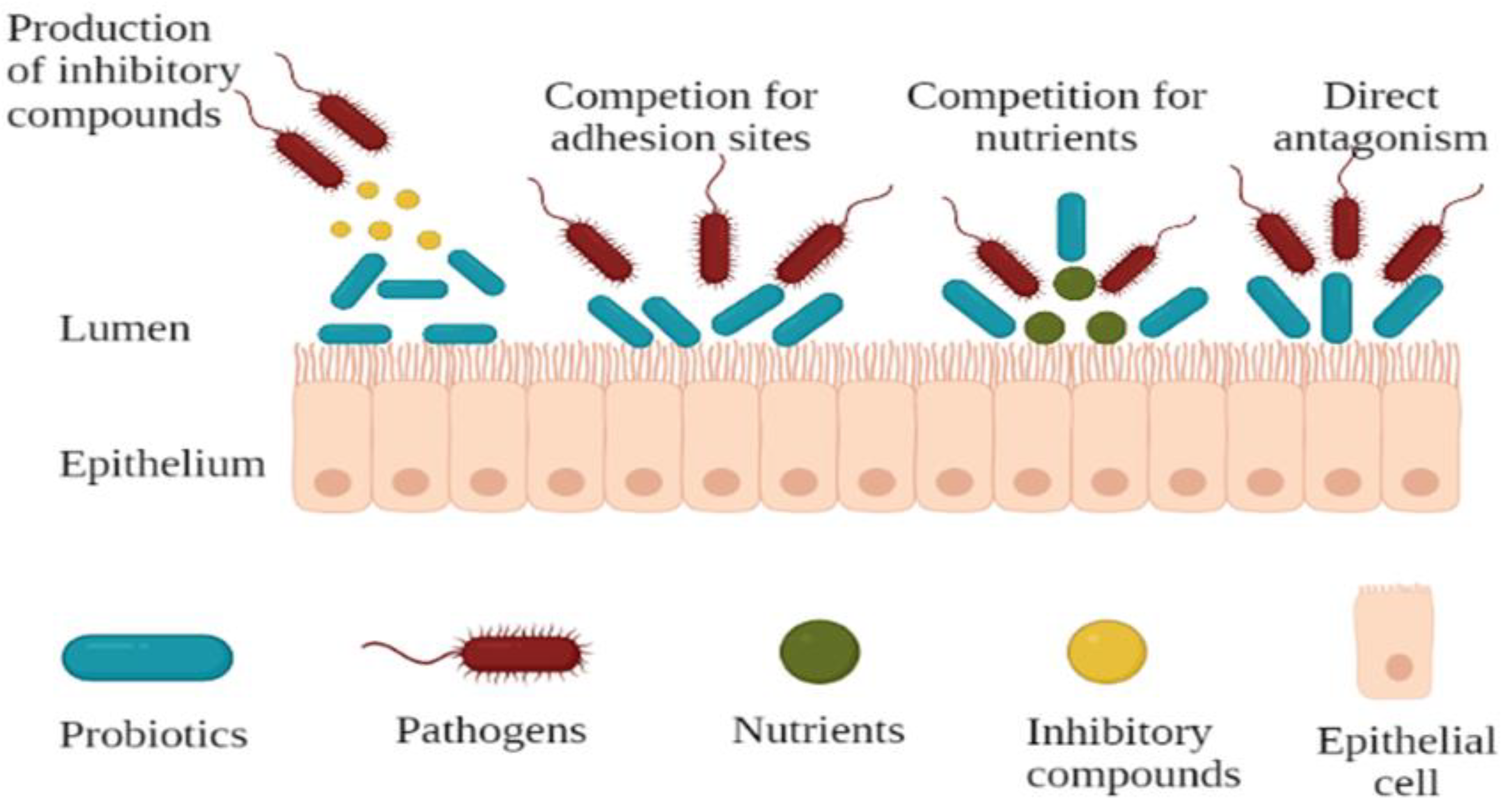
3.2. Antipathogenicity Effects
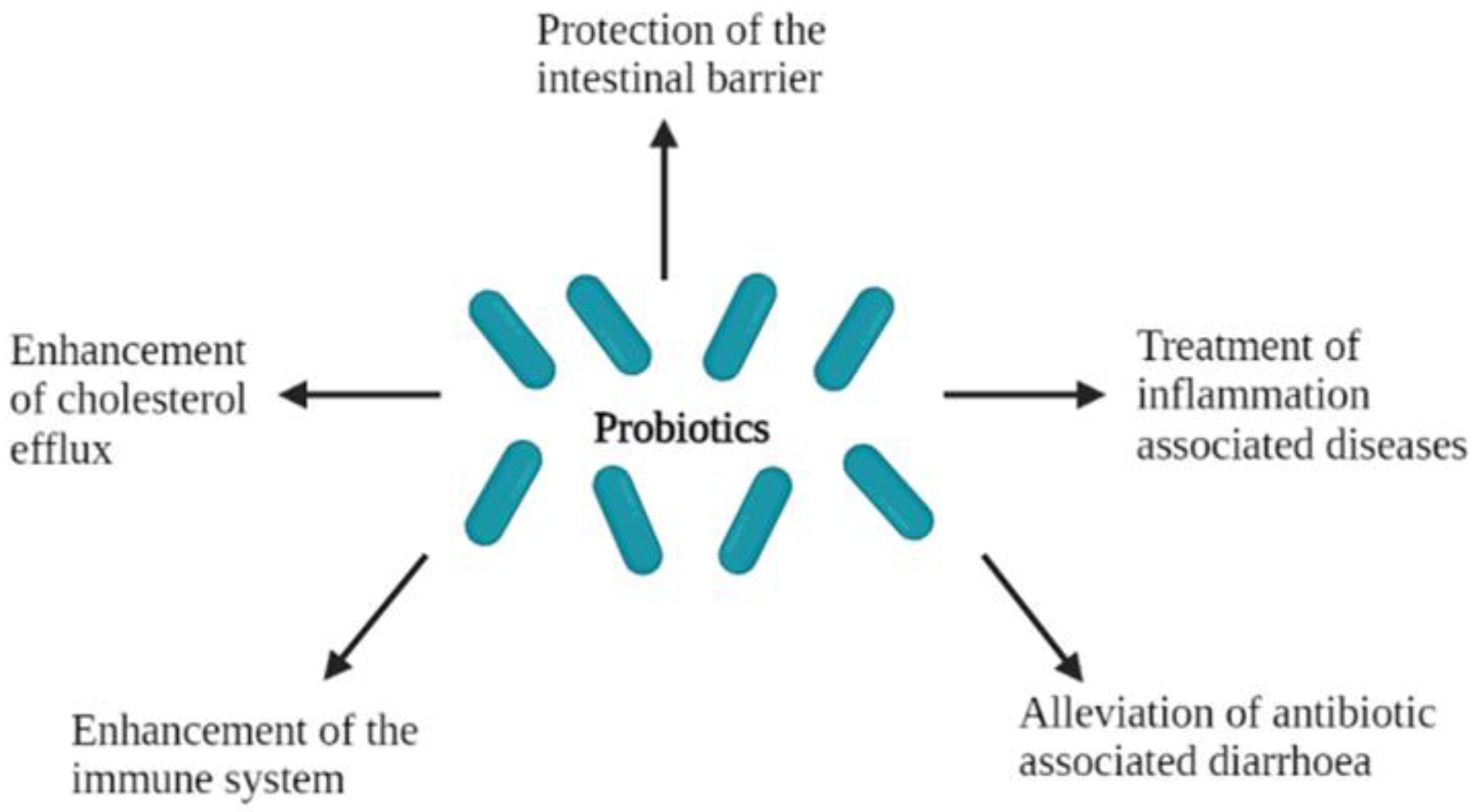
3.3. Protection of the Epithelial Barrier
3.4. Enhancement of Immune Response
4. Clinical Suitability: The Use of L. rhamnosus for Treatment of Disease
4.1. Alleviation of Antibiotic-Associated Diarrhoea
4.2. Treatment of Inflammation-Associated Diseases
4.3. Enhancement of Cholesterol Efflux
5. Safety Profile of L. rhamnosus
6. Additional Desirable Attributes of L. rhamnosus
6.1. L. rhamnosus Can Be Incorporated into Varied Delivery Food Vehicles
6.2. L. rhamnosus Is Amenable to the Enhancement of Stability Using Microencapsulation
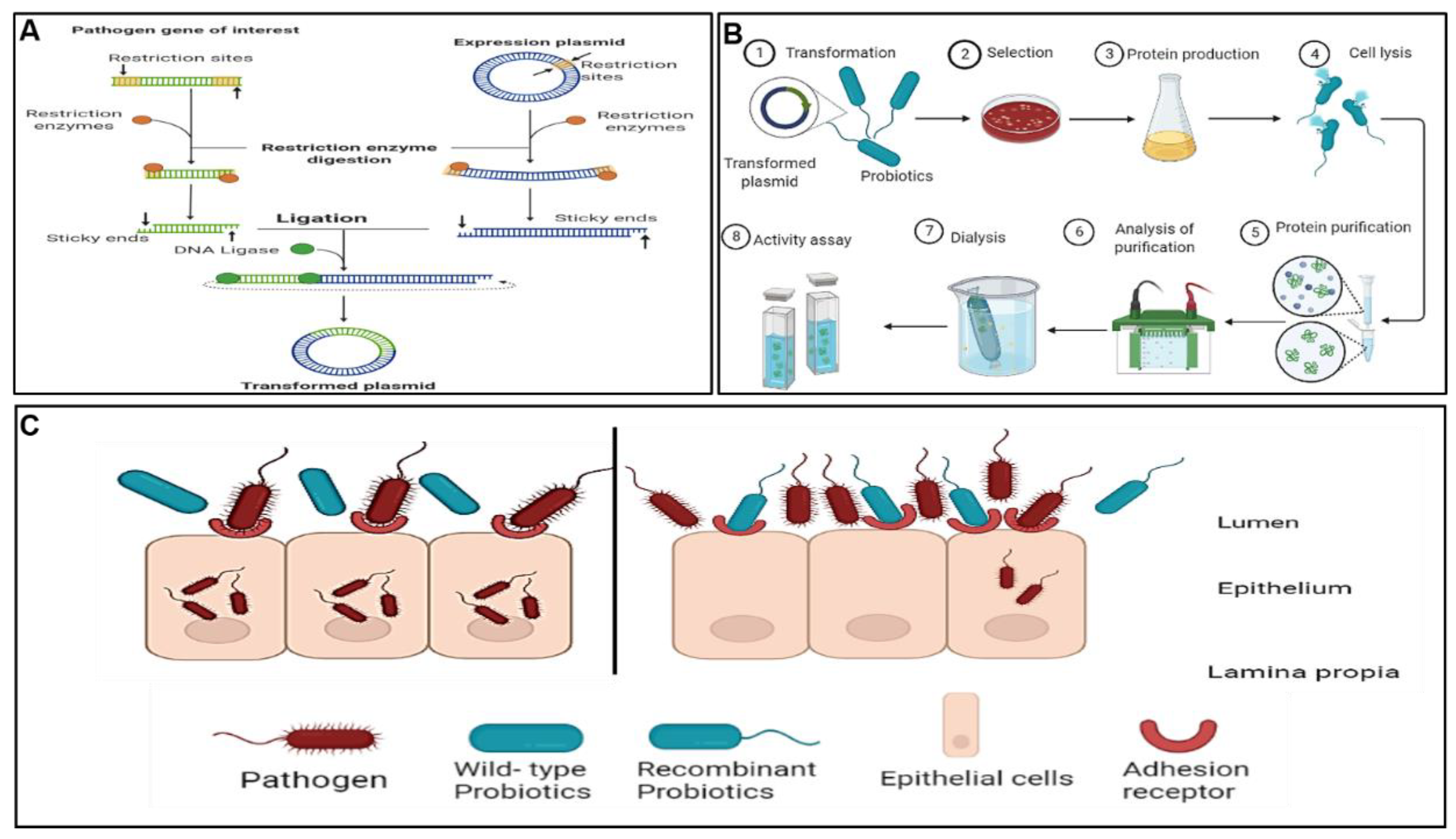
7. Present Studies on Bioengineering of L. rhamnosus
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alshammari, E.; Patel, M.; Sachidanandan, M.; Kumar, P.; Adnan, M. Potential evaluation and health fostering intrinsic traits of novel probiotic strain Enterococcus durans F3 isolated from the gut of fresh water fish Catla catla. Food Sci. Anim. Resour. 2019, 39, 844–861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mishra, S.; Acharya, S. A brief overview on probiotics: The health friendly microbes. Biomed. Pharmacol. J. 2021, 14, 1869–2285. [Google Scholar] [CrossRef]
- Binda, S.; Hill, C.; Johansen, E.; Obis, D.; Pot, B.; Sanders, M.E.; Tremblay, A.; Ouwenhand, A.C. Criteria to qualify microorganisms as “probiotic” in foods and dietary supplements. Front Microbiol. 2020, 11, 1662. [Google Scholar] [CrossRef]
- Zheng, J.; Wittouck, S.; Salvetti, E.; Franz, C.M.; Harris, H.; Mattarelli, P.; O’Toole, P.W.; Pot, B.; Vandamme, P.; Walter, J.; et al. A taxonomic note on the genus Lactobacillus: Description of 23 novel genera, emended description of the genus Lactobacillus Beijerinck 1901, and union of Lactobacillaceae and Leuconostocaceae. Int. J. Syst. Evol. Microbiol. 2020, 70, 2782–2858. [Google Scholar] [CrossRef]
- Kalliomäki, M.; Salminen, S.; Arvilommi, H.; Kero, P.; Koskinen, P.; Isolauri, E. Probiotics in Primary Prevention of Atopic Disease: A Randomised Placebo-Controlled Trial. Lancet 2001, 357, 1076–1079. [Google Scholar] [CrossRef]
- Moslem, P.; Hossein, N.; Mahdi, R.; Seyed, N.H.; Seyed, A.S. Lactobacillus rhamnosus Gorbach-Goldin (GG): A top well-researched probiotic strain. J. Med. Microbiol. 2017, 5, 46–59. [Google Scholar]
- De Champs, C.; Maroncle, N.; Balestrino, D.; Rich, C.; Forestier, C. Persistence of colonization of intestinal mucosa by a probiotic strain, Lactobacillus casei subsp. rhamnosus LCR35, after oral consumption. J. Clin. Microbiol. 2003, 41, 1270–1273. [Google Scholar]
- Forestier, C.; De Champs, C.; Vatoux, C.; Joly, B. Probiotic activities of Lactobacillus casei rhamnosus: In vitro adherence to intestinal cells and antimicrobial properties. Res. Microbiol. 2001, 152, 167–173. [Google Scholar] [CrossRef]
- Doron, S.; Snydman, D.R.; Gorbach, S.L. Lactobacillus GG: Bacteriology and clinical applications. Gastroenterol. Clin. N. Am. 2005, 34, 483–498. [Google Scholar] [CrossRef]
- Tuomola, E.M.; Ouwehand, A.C.; Salminenm, S.J. The effect of probiotic bacteria on the adhesion of pathogens to human intestinal mucus. FEMS Immunol. Med. Microbiol. 1999, 26, 137–142. [Google Scholar] [CrossRef] [PubMed]
- Martín, R.; Chamignon, C.; Mhedbi-Hajri, N.; Chain, F.; Derrien, M.; Escribano-Vázquez, U.; Garault, P.; Cotillard, A.; Pham, H.P.; Chervaux, C.; et al. The potential probiotic Lactobacillus rhamnosus CNCM I-3690 strain protects the intestinal barrier by stimulating both mucus production and cytoprotective response. Sci. Rep. 2019, 9, 5398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Segers, M.E.; Lebeer, S. Towards a better understanding of Lactobacillus rhamnosus GG-host interactions. Microb. Cell Fact. 2014, 13, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vélez, M.P.; De Keersmaecker, S.C.; Vanderleyden, J. Adherence factors of Lactobacillus in the human gastrointestinal tract. FEMS Microbiol. Lett. 2007, 276, 140–148. [Google Scholar] [CrossRef] [Green Version]
- Marianelli, C.; Cifani, N.; Pasquali, P. Evaluation of antimicrobial activity of probiotic bacteria against Salmonella enterica subsp. enterica serovar Typhimurium 1344 in a common medium under different environmental conditions. Res. Microbiol. 2010, 161, 673–680. [Google Scholar]
- Zhang, Z.G.; Ye, Z.Q.; Yu, L.; Shi, P. Phylogenomic reconstruction of lactic acid bacteria: An update. BMC Evol. Biol. 2011, 11, 1. [Google Scholar] [CrossRef] [Green Version]
- Szajewska, H.; Skórka, A.; Ruszczyński, M.; Gieruszczak-Białek, D. Meta-analysis: Lactobacillus GG for treating acute diarrhoea in children. Aliment. Pharmacol. Ther. 2007, 25, 871–881. [Google Scholar] [CrossRef]
- Szajewska, H.; Wanke, M.; Patro, B. Meta-analysis: The effects of Lactobacillus rhamnosus GG supplementation for the prevention of healthcare-associated diarrhoea in children. Aliment. Pharmacol. Ther. 2011, 34, 1079–1087. [Google Scholar] [CrossRef]
- Hojsak, I.; Abdović, S.; Szajewska, H.; Milosević, M.; Krznarić, Z.; Kolacek, S. Lactobacillus GG in the prevention of nosocomial gastrointestinal and respiratory tract infections. Pediatrics 2010, 125, 1171–1177. [Google Scholar] [CrossRef] [Green Version]
- Horvath, A.; Dziechciarz, P.; Szajewska, H. Meta-analysis: Lactobacillus rhamnosus GG for abdominal pain-related functional gastrointestinal disorders in childhood: Meta-analysis: Lactobacillus GG and abdominal pain-related functional disorders. Aliment. Pharmacol. Ther. 2011, 33, 1302–1310. [Google Scholar] [CrossRef]
- Manzoni, P.; Mostert, M.; Leonessa, M.L.; Priolo, C.; Farina, D.; Monetti, C.; Latino, M.A.; Gomirato, G. Oral supplementation with Lactobacillus casei subspecies rhamnosus prevents enteric colonization by Candida species in preterm neonates: A randomized study. Clin. Infect. Dis. 2006, 42, 1735–1742. [Google Scholar] [CrossRef] [PubMed]
- Allonsius, C.N.; van den Broek, M.F.L.; De Boeck, I.; Kiekens, S.; Oerlemans, E.F.M.; Kiekens, F.; Foubert, K.; Vandenheuvel, D.; Cos, P.; Delputte, P.; et al. Interplay between Lactobacillus rhamnosus GG and Candida and the involvement of exopolysaccharides. Microb. Biotechnol. 2017, 10, 1753–1763. [Google Scholar] [CrossRef]
- Boonma, P.; Spinler, J.K.; Venable, S.F.; Versalovic, J.; Tumwasorn, S. Lactobacillus rhamnosus l34 and Lactobacillus casei l39 suppress Clostridium difficile-induced il-8 production by colonic epithelial cells. BMC Microbiol. 2014, 14, 177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slykerman, R.F.; Hood, F.; Wickens, K.; Thompson, J.M.D.; Barthow, C.; Murphy, R.; Kang, J.; Rowden, J.; Stone, P.; Crane, J.; et al. Effect of Lactobacillus rhamnosus HN001 in pregnancy on postpartum symptoms of depression and anxiety: A randomised double-blind placebo-controlled trial. EBioMedicine 2017, 24, 159–165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Succi, M.; Tremonte, P.; Pannella, G.; Tipaldi, L.; Cozzolino, A.; Romaniello, R.; Sorrentino, E.; Coppola, R. Pre-cultivation with selected prebiotics enhances the survival and the stress response of Lactobacillus rhamnosus strains in simulated gastrointestinal transit. Front. Microbiol. 2017, 8, 1067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- FAO/WHO. Guidelines for the Evaluation of Probiotics in Food; Report of a Joint FAO/WHO Working Group on Drafting Guidelines for the Evaluation of Probiotics in Food; FAO: Gatineau, ON, Canada, 2002. [Google Scholar]
- Ljung, A.; Wadstöm, T. Lactic acid bacteria as probiotic. Curr. Iss. Intest. Microbiol. 2006, 7, 73–90. [Google Scholar]
- Rönkä, E.; Malinen, E.; Saarela, M.; Rinta-Koski, M.; Aarnikunnas, J.; Palva, A. Probiotic and milk technological properties of Lactobacillus brevis. Int. J. Food Microbiol. 2003, 83, 63–74. [Google Scholar] [CrossRef]
- Jacobsen, C.N.; Rosenfeldt, N.V.; Hayford, A.E.; Møller, P.L.; Michaelsen, K.F.; Paerregaard, A.; Sandström, B.; Tvede, M.; Jakobsen, M. Screening of probiotic activities of forty-seven strains of Lactobacillus spp. by in vitro techniques and evaluation of the colonization ability of five selected strains in humans. Appl. Environ. Microbiol. 1999, 65, 4949–4956. [Google Scholar]
- Gardiner, G.E.; O’sullivan, E.; Kelly, J.; Auty, M.A.E.; Fitzgerald, G.F.; Collins, J.K.; Ross, R.P.; Stanton, C. Comparative survival rates of human-derived probiotic Lactobacillus paracasei and L. salivarius strains during heat treatment and spray drying. Appl. Environ. Microbiol. 2000, 66, 2605–2612. [Google Scholar] [CrossRef] [Green Version]
- Pereira, D.I.; Gibson, G.R. Cholesterol assimilation by lactic acid bacteria and Bifidobacteria isolated from the human gut. Appl. Environ. Microbiol. 2002, 68, 4689–4693. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Adams, M.C. In vitro assessment of the upper gastrointestinal tolerance of potential probiotic dairy propionibacteria. Int. J. Food Microbiol. 2004, 91, 253–260. [Google Scholar] [CrossRef] [PubMed]
- Ranadheera, C.S.; Evans, C.A.; Adams, M.C.; Baines, S.K. In vitro analysis of gastrointestinal tolerance and intestinal cell adhesion of probiotics in goat’s milk ice cream and yogurt. Int. Food Res. J. 2012, 49, 619–625. [Google Scholar] [CrossRef]
- Servin, A.L. Antagonistic activities of lactobacilli and bifidobacteria against microbial pathogens. FEMS Microbiol. Rev. 2004, 28, 405–440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernet-Carnard, M.F.; Lievin, V.; Brassart, D.; Neeser, J.R.; Servin, A.L.; Hudault, S. The human L. acidophilus strain LA1 secretes a non-bacteriocin anti-bacterial substance(s) active in vitro and in vivo. Appl. Environ. Microbiol. 1997, 63, 2747–2753. [Google Scholar] [CrossRef] [Green Version]
- Salyers, A.A.; Gupta, A.; Wang, Y. Human intestinal bacteria as reservoirs for antibiotic resistance genes. Trends Microbiol. 2004, 12, 412–416. [Google Scholar] [CrossRef]
- Senok, A.C.; Ismaeel, A.Y.; Botta, G.A. Probiotics: Facts and myths. Clin. Microbiol. Infect. 2005, 11, 958–966. [Google Scholar] [CrossRef] [Green Version]
- Shah, N.P. Probiotic bacteria: Selective enumeration and survival in dairy foods. J. Dairy Sci. 2000, 83, 894–907. [Google Scholar] [CrossRef]
- Corcoran, B.M.; Stanton, C.; Fitzgerald, G.F.; Ross, R.P. Survival of probiotic lactobacilli in acidic environments is enhanced in the presence of metabolizable sugars. Appl. Environ. Microbiol. 2005, 71, 3060–3067. [Google Scholar] [CrossRef] [Green Version]
- Holzapfel, W.H.; Haberer, P.; Snel, J.; Schillinger, U.; Huis in’t Veld, J.H.J. Overview of gut flora and probiotics. Int. J. Food Microbiol. 1998, 41, 85–101. [Google Scholar] [CrossRef]
- Guan, N.; Liu, L. Microbial response to acid stress: Mechanisms and applications. Appl. Microbiol. Biotechnol. 2020, 104, 51–65. [Google Scholar] [CrossRef] [Green Version]
- Verdenelli, M.C.; Ghelfi, F.; Silvi, S.; Orpianesi, C.; Cecchini, C.; Cresci, A. Probiotic properties of Lactobacillus rhamnosus and Lactobacillus paracasei isolated from human faeces. Eur. J. Nutr. 2009, 48, 355–363. [Google Scholar] [CrossRef] [PubMed]
- Saarela, M.; Mogensen, G.; Fonden, R.; Matto, J.; Mattila-Sandholm, T. Probiotic bacteria: Safety; functional and technological properties. J. Biotechnol. 2000, 84, 197–215. [Google Scholar] [CrossRef]
- Ding, W.K.; Shah, N.P. Acid, bile, and heat tolerance of free and microencapsulated probiotic bacteria. J. Food Sci. 2007, 72, 446–450. [Google Scholar] [CrossRef]
- Leite, A.M.; Miguel, M.A.L.; Peixoto, R.S.; Ruas-Madiedo, P.; Paschoalin, V.M.F.; Mayo, B.; Delgado, S. Probiotic potential of selected lactic acid bacteria strains isolated from Brazilian kefir grains. J. Dairy Sci. 2015, 98, 3622–3632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kimoto-Nira, H.; Kobayashi, M.; Nomura, M.; Sasaki, K.; Suzuki, C. Bile resistance in Lactococcus lactis strains varies with cellular fatty acid composition: Analysis by using different growth media. Int. J. Food Microbiol. 2009, 131, 183–188. [Google Scholar] [CrossRef] [PubMed]
- Su, J.; Wang, T.; Li, Y.Y.; Li, J.; Zhang, Y.; Wang, Y.; Wang, H.; Li, H. Antioxidant properties of wine lactic acid bacteria: Oenococcus oeni. Appl. Microbiol. Biotechnol. 2015, 99, 5189–5202. [Google Scholar] [CrossRef] [PubMed]
- Bongaerts, G.P.A.; Severijnen, R.S.V.M. The beneficial; antimicrobial effect of probiotics. Med. Hypotheses 2001, 56, 174–177. [Google Scholar] [CrossRef]
- Guarner, F.; Khan, A.G.; Garisch, J.; Eliakim, R.; Gangl, A.; Thomson, A.; Krabshuis, J.; Lemair, T.; Kaufmann, P.; de Paula, J.A.; et al. World gastroenterology organisation global guidelines: Probiotics and prebiotics October 2011. J. Clin. Gastroenterol. 2012, 46, 468–481. [Google Scholar] [CrossRef]
- Annuk, H.; Shchepetova, J.; Kullisaar, T.; Songisepp, E.; Zilmer, M.; Mikelsaar, M. Characterization of intestinal lactobacilli as putative probiotic candidates. J. Appl. Microbiol. 2003, 94, 403–412. [Google Scholar] [CrossRef]
- Bron, P.A.; Tomita, S.; Mercenier, A.M.E.; Kleerebezem, M. Cell surface-associated compounds of probiotic lactobacilli sustain the strain-specificity dogma. Curr. Opin. Microbiol. 2013, 16, 262–269. [Google Scholar] [CrossRef]
- Collado, M.; Meriluoto, J.; Salminen, S. Adhesion and aggregation properties of probiotic and pathogen strains. Eur. Food Res. Technol. 2008, 226, 1065–1073. [Google Scholar] [CrossRef]
- Haidar, R.; Fermaud, M.; Calvo-Garrido, C.; Roudet, J.; Deschamps, A. Modes of action for biological control of Botrytis cinerea by antagonistic bacteria. Phytopathol. Mediterr. 2016, 55, 301–322. [Google Scholar]
- Honoré, A.H.; Aunsbjerg, S.D.; Ebrahimi, P.; Thorsen, M.; Benfeldt, C.; Knøchel, S.; Skov, T. Metabolic footprinting for investigation of antifungal properties of Lactobacillus paracasei. Anal. Bioanal. Chem. 2016, 408, 83–96. [Google Scholar] [CrossRef] [PubMed]
- Golowczyc, M.A.; Silva, J.; Abraham, A.G.; De Antoni, G.L.; Teixeira, P. Preservation of probiotic strains isolated from kefir by spray drying. Lett. Appl. Microbiol. 2010, 50, 7–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siedler, S.; Rau, M.H.; Bidstrup, S.; Vento, J.M.; Aunsbjerg, S.D.; Bosma, E.F.; McNair, L.M.; Beisel, C.L.; Neves, A.R.; Drake, H.L. Competitive exclusion is a major bioprotective mechanism of lactobacilli against fungal spoilage in fermented milk products. Appl. Environ. Microbiol. 2020, 86, e02312-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerwien, F.; Skrahina, V.; Kasper, L.; Hube, B.; Brunke, S. Metals in fungal virulence. FEMS Microbiol. Rev. 2018, 42, fux050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pitino, I.; Randazzo, C.L.; Cross, K.L.; Parker, M.L.; Bisignano, C.; Wickham, M.S.J.; Mandalari, G.; Caggia, C. Survival of Lactobacillus rhamnosus strains inoculated in cheese matrix during simulated human digestion. Food Microbiol. 2012, 31, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Monteagudo-Mera, A.; Rastall, R.A.; Gibson, G.R.; Charalampopoulos, D.; Chatzifragkou, A. Adhesion mechanisms mediated by probiotics and prebiotics and their potential impact on human health. Appl. Microbiol. Biotechnol. 2019, 103, 6463–6472. [Google Scholar] [CrossRef] [Green Version]
- Madsen, K.; Cornish, A.; Soper, P.; McKaigney, C.; Jijon, H.; Yachimec, C.; Doyle, J.; Jewell, L.; De Simone, C. Probiotic bacteria enhance murine and human intestinal epithelial barrier function. Gastroenterology 2001, 121, 580–591. [Google Scholar] [CrossRef] [Green Version]
- Alvarez, S.; Villena, J.; Tohno, M.; Salva, S.; Kitazawa, H. Modulation of innate immunity by lactic acid bacteria: Impact on host response to infections. Curr. Opin. Immunol. 2009, 3, 87–126. [Google Scholar]
- Villena, J.; Oliveira, M.L.S.; Ferreira, P.C.; Salva, S.; Alvarez, S. Lactic acid bacteria in the prevention of pneumococcal respiratory infection: Future opportunities and challenges. Int. Immunopharmacol. 2011, 11, 1633–1645. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.M.; Yu, L. Opposing gatekeepers of apical sterol transport: Niemann-pick C1-like 1 (NPC1l1) and ATP-binding cassette transporters G5 and G8 (ABCG5/ABCG8). Immunol. Endocr. Metab. Agents Med. Chem. 2009, 9, 18–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Y.; Zheng, Y. The probiotic Lactobacillus acidophilus reduces cholesterol absorption through the down- regulation of Niemann-Pick C1-like 1 in Caco-2 cells. Br. J. Nutr. 2010, 103, 473–478. [Google Scholar] [CrossRef] [Green Version]
- Rose, M.A.; Stieglitz, F.; Köksal, A.; Schubert, R.; Schulze, J.; Zielen, S. Efficacy of probiotic Lactobacillus GG on allergic sensitization and asthma in infants at risk: Probiotics and allergic asthma prevention. Clin. Exp. Allergy 2010, 40, 1398–1405. [Google Scholar] [CrossRef]
- Cabana, M.D.; McKean, M.; Caughey, A.B.; Fong, L.; Lynch, S.; Wong, A.; Leong, R.; Boushey, H.A.; Hilton, J.F. Early probiotic supplementation for eczema and asthma prevention: A randomized controlled trial. Pediatrics 2017, 140, e20163000. [Google Scholar] [CrossRef] [Green Version]
- Szajewska, H.; Canani, R.B.; Guarino, A.; Hojsak, I.; Indrio, F.; Kolacek, S.; Orel, R.; Shamir, R.; Vandenplas, Y.; van Goudoever, J.B.; et al. Probiotics for the prevention of antibiotic-associated diarrhea in children. J. Pediatr. Gastroenterol. Nutr. 2016, 62, 495–506. [Google Scholar] [CrossRef] [Green Version]
- Alander, M.; Satokari, R.; Korpela, R.; Saxelin, M.; Vilpponen-Salmela, T.; Mattila-Sandholm, T.; von Wright, A. Persistence of colonization of human colonic mucosa by a probiotic strain; Lactobacillus rhamnosus GG; after oral consumption. Appl. Environ. Microbiol. 1999, 65, 351–354. [Google Scholar] [CrossRef] [Green Version]
- Schiffrin, E.J.; Rochat, F.; Link-Amster, H.; Aeschlimann, J.M.; Donnet-Hughes, A. Immunomodulation of human blood cells following the ingestion of lactic acid bacteria. J. Dairy Sci. 1995, 78, 491–497. [Google Scholar] [CrossRef]
- Coconnier, M.H.; Bernet, M.F.; Kerneis, S.; Chauviere, G. Inhibition of adhesion of enteroinvasive pathogens to human intestinal caco-2 cells by Lactobacillus acidophilus strain LB decreases bacterial invasion. FEMS Microbiol. Lett. 1993, 110, 299–305. [Google Scholar] [CrossRef]
- Alander, M.; Korpela, R.; Saxelin, M.; Vilpponen-Salmela, T.; Mattila-Sandholm, T.; von Wright, A. Recovery of Lactobacillus rhamnosus GG from human colonic biopsies. Lett. Appl. Microbiol. 1997, 24, 361–364. [Google Scholar] [CrossRef]
- Nollet, L.; Demeyer, D.; Verstraete, W. Effect of 2-bromoethanesulfonic acid and Peptostreptococcus productus ATCC 35244 addition on stimulation of reductive acetogenesis in the ruminal ecosystem by selective inhibition of methanogenesis. Appl. Environ. Microbiol. 1997, 63, 194–200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiu, I.M.; Heesters, B.A.; Ghasemlou, N.; Von Hehn, C.A.; Zhao, F.; Tran, J.; Wainger, B.; Strominger, A.; Muralidharan, S.; Horswill, A.R.; et al. Bacteria activate sensory neurons that modulate pain and inflammation. Nature 2013, 501, 52–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salva, S.; Villena, J.; Alvarez, S. Immunomodulatory activity of Lactobacillus rhamnosus strains isolated from goat milk: Impact on intestinal and respiratory infections. Int. J. Food Microbiol. 2010, 141, 82–89. [Google Scholar] [CrossRef] [PubMed]
- Dong, H.; Rowland, I.; Yaqoob, P. Comparative effects of six probiotic strains on immune function in vitro. Br. J. Nutr. 2012, 108, 459–470. [Google Scholar] [CrossRef] [Green Version]
- Takeda, S.; Kawahara, S.; Hidaka, M.; Yoshida, H.; Watanabe, W.; Takeshita, M.; Kikuchi, Y.; Bumbein, D.; Muguruma, M.; Kurokawa, M. Effects of oral administration of probiotics from Mongolian dairy products on the Th1 immune response in mice. Biosci. Biotechnol. Biochem. 2013, 77, 1372–1378. [Google Scholar] [CrossRef] [Green Version]
- Lebeer, S.; Vanderleyden, J.; De Keersmaecker, S.C.J. Genes and molecules of lactobacilli supporting probiotic action. Microbiol. Mol. Biol. Rev. 2008, 72, 728–764. [Google Scholar] [CrossRef] [Green Version]
- Bermudez-Brito, M.; Muñoz-Quezada, S.; Gomez-Llorente, C.; Romero, F.; Gil, A. Lactobacillus rhamnosus and its cell-free culture supernatant differentially modulate inflammatory biomarkers in Escherichia coli-challenged human dendritic cells. Br. J. Nutr. 2014, 111, 1727–1737. [Google Scholar] [CrossRef] [Green Version]
- Bernardeau, M.; Guguen, M.; Vernoux, J.P. Beneficial lactobacilli in food and feed: Long-term use; biodiversity and proposals for specific and realistic safety assessments. FEMS Microbiol. Rev. 2006, 30, 487–513. [Google Scholar] [CrossRef]
- Guarino, A.; Vecchio, A.L.; Canani, R.B. Probiotics as prevention and treatment for diarrhea. Curr. Opin. Gastroenterol. 2009, 25, 18–23. [Google Scholar] [CrossRef]
- Marteau, P.; Pochart, P.; Bouhnik, Y.; Rambaud, J.C. The fate and effects of transiting; nonpathogenic microorganisms in the human intestine. World Rev. Nutr. Diet. 1993, 74, 1–21. [Google Scholar]
- Vamanu, E.; Vamanu, A. Viability of the Lactobacillus rhamnosus IL1 strain in simulated gastrointestinal conditions. Int. J. Pharmacol. 1993, 6, 732–737. [Google Scholar] [CrossRef]
- Koskenniemi, K.; Laakso, K.; Koponen, J.; Kankainen, M.; Greco, D.; Auvinen, P.; Savijoki, K.; Nyman, T.A.; Surakka, A.; Salusjärvi, T.; et al. Proteomics and transcriptomics characterization of bile stress response in probiotic Lactobacillus rhamnosus GG. Mol. Cell. Proteom. 2011, 10, S1–S18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Succi, M.; Tremonte, P.; Reale, A.; Sorrentino, E.; Grazia, L.; Pacifico, S.; Coppola, R. Bile salt and acid tolerance of lactobacillus rhamnosus strains isolated from Parmigiano Reggiano cheese. FEMS Microbiol. Lett. 2005, 244, 129–137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reale, A.; Di Renzo, T.; Rossi, F.; Zotta, T.; Iacumin, L.; Preziuso, M.; Parente, E.; Sorrentino, E.; Coppola, R. Tolerance of Lactobacillus casei; Lactobacillus paracasei and Lactobacillus rhamnosus strains to stress factors encountered in food processing and in the gastro-intestinal tract. LWT-Food Sci. Technol. 2015, 60, 721–728. [Google Scholar] [CrossRef]
- Mirlohi, M.; Soleimanian-Zad, S.; Dokhani, S.; Sheikh-Zeinodin, M.; Abghary, A. Investigation of Acid and Bile Tolerance of Native Lactobacilli Isolated from Fecal Samples and Commercial Probiotics by Growth and Survival Studies. Iranian J. Biotechnol. 2009, 7, 233–240. [Google Scholar]
- Pace, F.; Pace, M.; Quartarone, G. Probiotics in digestive diseases: Focus on Lactobacillus GG. Minerva Gastroenterol. Dietol. 2015, 61, 273–292. [Google Scholar]
- Shi, T.; Aryantini, N.P.D.; Urashima, T.; Fukuda, K.; Nishiyama, K.; Nakamata, K.; Yamamoto, Y.; Mukai, T.; Mikumo, D.; Oda, Y.; et al. Isolation of potential probiotic Lactobacillus rhamnosus strains from traditional fermented mare milk produced in Sumbawa Island of Indonesia. Biosci. Biotechnol. Biochem. 2012, 76, 1897–1903. [Google Scholar] [CrossRef] [Green Version]
- Rolfe, R.D. The role of probiotic cultures in the control of gastrointestinal health. J. Nutr. 2000, 130, 396S–402S. [Google Scholar] [CrossRef]
- Schiffrin, E.J.; Blum, S. Interactions between the microbiota and the intestinal mucosa. Eur. J. Clin. Nutr. 2002, 56, S60–S64. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, S.; Macfarlane, G.T.; Fite, A.; McBain, A.J.; Gilbert, P.; Macfarlane, S. Mucosa-associated bacterial diversity in relation to human terminal ileum and colonic biopsy samples. Appl. Environ. Microbiol. 2007, 73, 7435–7442. [Google Scholar] [CrossRef] [Green Version]
- De Keersmaecker, S.C.J.; Verhoeven, T.L.A.; Desair, J.; Marchal, K.; Vanderleyden, J.; Nagy, I. Strong antimicrobial activity of Lactobacillus rhamnosus GG against Salmonella typhimurium is due to accumulation of lactic acid. FEMS Microbiol. Lett. 2006, 259, 89–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beristain-Bauza, S.C.; Mani-López, E.; Palou, E.; López-Malo, A. Antimicrobial activity and physical properties of protein films added with cell-free supernatant of Lactobacillus rhamnosus. Food Control 2016, 62, 44–51. [Google Scholar] [CrossRef]
- de Alcântara, A.L.D.A.; Bruzaroski, S.R.; Luiz, L.L.; de Souza, C.H.B.; Poli-Frederico, R.C.; Fagnani, R.; de Santana, E.H.W. Antimicrobial activity of Lactobacillus rhamnosus against Pseudomonas fluorescens and Pseudomonas putida from raw milk. J. Food Process. Preserv. 2019, 43, e14082. [Google Scholar] [CrossRef]
- Vesterlund, S.; Karp, M.; Salminen, S.; Ouwehand, A.C. Staphylococcus aureus adheres to human intestinal mucus but can be displaced by certain lactic acid bacteria. Microbiology 2006, 152, 1819–1826. [Google Scholar]
- Wong, S.S.; Toh, Z.Q.; Dunne, E.M.; Mulholland, E.K.; Tang, M.L.K.; Robins-Browne, R.M.; Licciardi, P.V.; Satzke, C. Inhibition of Streptococcus pneumoniae adherence to human epithelial cells in vitro by the probiotic Lactobacillus rhamnosus GG. BMC Res. Notes 2013, 6, 135. [Google Scholar] [CrossRef] [Green Version]
- Mohammedsaeed, W.; McBain, A.J.; Cruickshank, S.M.; O’Neill, C.A. Lactobacillus rhamnosus GG inhibits the toxic effects of Staphylococcus aureus on epidermal keratinocytes. Appl. Environ. Microbiol. 2014, 80, 5773–5781. [Google Scholar] [CrossRef] [Green Version]
- Johnson-Henry, K.C.; Donato, K.A.; Shen-Tu, G.; Gordanpour, M.; Sherman, P.M. Lactobacillus rhamnosus strain GG prevents enterohemorrhagic Escherichia coli O157:H7-induced changes in epithelial barrier function. Infect. Immun. 2008, 76, 1340–1348. [Google Scholar] [CrossRef] [Green Version]
- Ephraim, E.; Schultz, R.D.; Safdar, N. Lactobacillus rhamnosus GG Protects Cells from Clostridium difficile Toxins. Br. Microbiol. Res. J. 2013, 3, 165–175. [Google Scholar] [CrossRef]
- Turovskiy, Y.; Ludescher, R.D.; Aroutcheva, A.A.; Faro, S.; Chikindas, M.L. Lactocin 160; a bacteriocin produced by vaginal Lactobacillus rhamnosus; targets cytoplasmic membranes of the vaginal pathogen; Gardnerella vaginalis. Probiotics Antimicrob. Proteins 2009, 1, 67–74. [Google Scholar] [CrossRef] [Green Version]
- Yamaguchi, M.; Terao, Y.; Mori, Y.; Hamada, S.; Kawabata, S. Pfba; a novel plasmin- and fibronectin-binding protein of Streptococcus pneumoniae; contributes to fibronectin-dependent adhesion and antiphagocytosis. J. Biol. Chem. 2008, 283, 36272–36279. [Google Scholar] [CrossRef] [Green Version]
- Jeong, Y.J.; Moon, G.S. Antilisterial bacteriocin from Lactobacillus rhamnosus CJNU 0519 presenting a narrow antimicrobial spectrum. Korean J. Food Sci. Anim. Resour. 2015, 35, 137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biswas, S.; Keightley, A.; Biswas, I. Characterization of a stress tolerance-defective mutant of Lactobacillus rhamnosus LRB. Mol. Oral Microbiol. 2019, 34, 153–167. [Google Scholar] [CrossRef] [PubMed]
- Patel, M.; Siddiqui, A.J.; Hamadou, W.S.; Surti, M.; Awadelkareem, A.M.; Ashraf, S.A.; Alreshidi, M.; Snoussi, M.; Rizvi, S.M.; Bardakci, F.; et al. Inhibition of bacterial adhesion and antibiofilm activities of a glycolipid biosurfactant from Lactobacillus rhamnosus with its physicochemical and functional properties. Antibiotics 2021, 10, 1546. [Google Scholar] [CrossRef] [PubMed]
- Makras, L.; Triantafyllou, V.; Fayol-Messaoudi, D.; Adriany, T.; Zoumpopoulou, G.; Tsakalidou, E.; Servin, A.; De Vuyst, L. Kinetic analysis of the antibacterial activity of probiotic lactobacilli towards Salmonella enterica serovar Typhimurium reveals a role for lactic acid and other inhibitory compounds. Res. Microbiol. 2006, 157, 241–247. [Google Scholar] [CrossRef]
- Hütt, P.; Shchepetova, J.; Loivukene, K.; Kullisaar, T.; Mikelsaar, M. Antagonistic activity of probiotic lactobacilli and bifidobacteria against entero-and uropathogens. J. Appl. Microbiol. 2006, 100, 1324–1332. [Google Scholar] [CrossRef] [PubMed]
- Tytgat, H.L.P.; van Teijlingen, N.H.; Sullan, R.M.A.; Douillard- François, P.; Rasinkangas, P.; Messing, M.; Reunanen, J.; Satokari, J.; Vanderleyden, J.; Dufrene, Y.F.; et al. Probiotic gut microbiota isolate interacts with dendritic cells via glycosylated heterotrimeric pili. PLoS ONE 2016, 11, e0151824. [Google Scholar] [CrossRef] [Green Version]
- Zhou, B.; Zhang, D. Antibacterial effects of bacteriocins isolated from lactobacillus rhamnosus (ATCC 53103) in a rabbit model of knee implant infection. Exp. Ther. Med. 2018, 15, 2985–2989. [Google Scholar] [CrossRef]
- Naik, A.K.; Pandey, U.; Mukherjee, R.; Mukhopadhyay, S.; Chakraborty, S.; Ghosh, A.; Aich, P. Lactobacillus rhamnosus GG reverses mortality of neonatal mice against Salmonella challenge. Toxicol. Res. 2019, 8, 361–372. [Google Scholar] [CrossRef]
- Miyauchi, E.; O’Callaghan, J.; Buttó, L.F.; Hurley, G.; Melgar, S.; Tanabe, S.; Shanahan, F.; Nally, K.; O’Toole, P.W. Mechanism of protection of transepithelial barrier function by Lactobacillus salivarius: Strain dependence and attenuation by bacteriocin production. Am. J. Physiol. Gastrointest. Liver Physiol. 2012, 303, G1029–G1041. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Kirpich, I.; Liu, Y.; Ma, Z.; Barve, S.; McClain, C.J.; Feng, W. Lactobacillus rhamnosus GG treatment potentiates intestinal hypoxia-inducible factor; promotes intestinal integrity and ameliorates alcohol-induced liver injury. Am. J. Pathol. 2011, 179, 2866–2875. [Google Scholar] [CrossRef]
- Orlando, A.; Linsalata, M.; D’Attoma, B.; Russo, F. Changes in paracellular permeability induced by Pepsin-Trypsin digested Gliadin (PTG): Role of polyamines in the Lactobacillus rhamnosus GG protective action. J. Funct. Foods 2017, 36, 52–62. [Google Scholar] [CrossRef]
- Van Tassell, M.L.; Miller, M.J. Lactobacillus adhesion to mucus. Nutrients 2011, 3, 613–636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- von Ossowski, I.; Reunanen, J.; Satokari, R.; Vesterlund, S.; Kankainen, M.; Huhtinen, H.; Tynkkynen, S.; Salminen, S.; de Vos, W.M.; Palva, A. Mucosal adhesion properties of the probiotic Lactobacillus rhamnosus GG SpaCBA and SpaFED pilin subunits. Appl. Environ. Microbiol. 2010, 76, 2049–2057. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vélez, M.P.; Petrova, M.I.; Lebeer, S.; Verhoeven, T.L.; Claes, I.; Lambrichts, I.; Tynkkynen, S.; Vanderleyden, J.; De Keersmaecker, S.C. Characterization of MabA; a modulator of Lactobacillus rhamnosus GG adhesion and biofilm formation. FEMS Microbiol. Immunol. 2010, 59, 386–398. [Google Scholar] [CrossRef] [Green Version]
- Huang, S.H.; He, L.; Zhou, Y.; Wu, C.H.; Jong, A. Lactobacillus rhamnosus GG suppresses meningitic E. coli K1 penetration across human intestinal epithelial cells in vitro and protects neonatal rats against experimental hematogenous meningitis. Int. J. Microbiol. 2009, 2009, 647862. [Google Scholar] [CrossRef] [Green Version]
- Han, X.; Lee, A.; Huang, S.; Gao, J.; Spence, J.R.; Owyang, C. Lactobacillus rhamnosus GG prevents epithelial barrier dysfunction induced by interferon-gamma and fecal supernatants from irritable bowel syndrome patients in human intestinal enteroids and colonoids. Gut Microbes 2019, 10, 59–76. [Google Scholar] [CrossRef] [Green Version]
- Seth, A.; Yan, F.; Polk, D.B.; Rao, R.K. Probiotics ameliorate the hydrogen peroxide-induced epithelial barrier disruption by a PKC- and MAP kinase-dependent mechanism. Am. J. Physiol. Gastrointest. Liver Physiol. 2008, 294, 1060–1069. [Google Scholar] [CrossRef] [Green Version]
- Herrera, M.; Salva, S.; Villena, J.; Barbieri, N.; Alvarez, S. Lactobacillus rhamnosus CRL1505 enhances systemic and respiratory innate immune response in immunocompromised malnourished mice. J. Funct. Foods 2013, 5, 1693–1704. [Google Scholar] [CrossRef]
- Villena, J.; Chiba, E.; Vizoso-Pinto, M.; Tomosada, Y.; Takahashi, T.; Ishizuka, T.; Aso, H.; Salva, S.; Alvarez, S.; Kitazawa, H. Immunobiotic Lactobacillus rhamnosus strains differentially modulate antiviral immune response in porcine intestinal epithelial and antigen presenting cells. BMC Microbiol. 2014, 14, 126. [Google Scholar] [CrossRef] [Green Version]
- Lebeer, S.; Claes, I.; Tytgat, H.L.; Verhoeven, T.L.; Marien, E.; von Ossowski, I.; Reunanen, J.; Palva, A.; Vos, W.M.; Keersmaecker, S.C.; et al. Functional analysis of Lactobacillus rhamnosus GG pili in relation to adhesion and immunomodulatory interactions with intestinal epithelial cells. Appl. Environ. Microbiol. 2012, 78, 185–193. [Google Scholar] [CrossRef] [Green Version]
- Vargas García, C.E.; Petrova, M.; Claes, I.J.; De Boeck, I.; Verhoeven, T.L.; Dilissen, E.; von Ossowski, I.; Palva, A.; Bullens, D.M.; Vanderleyden, J.; et al. Piliation of Lactobacillus rhamnosus GG promotes adhesion; phagocytosis; and cytokine modulation in macrophages. Appl. Environ. Microbiol. 2015, 81, 2050–2062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.O.; Sheikh, H.I.; Ha, S.D.; Martins, A.; Reid, G. G-CSF-mediated inhibition of JNK is a key mechanism for Lactobacillus rhamnosus -induced suppression of TNF production in macrophages. Cell. Microbiol. 2006, 8, 1958–1971. [Google Scholar] [CrossRef] [PubMed]
- Bleau, C.; Monges, A.; Rashidan, K.; Laverdure, J.P.; Lacroix, M.; Van Calsteren, M.R.; Millette, M.; Savard, R.; Lamontagne, L. Intermediate chains of exopolysaccharides from Lactobacillus rhamnosus RW-9595M increase IL-10 production by macrophages. J. Appl. Microbiol. 2010, 108, 666–675. [Google Scholar] [CrossRef] [PubMed]
- Banna, G.L.; Torino, F.; Marletta, F.; Santagati, M.; Salemi, R.; Cannarozzo, E.; Falzone, L.; Ferraù, F.; Libra, M. Lactobacillus rhamnosus GG: An overview to explore the rationale of its use in cancer. Front. Pharmacol. 2017, 8, 603. [Google Scholar]
- Mahnet, S.; Kirin, S.; Kos, B.; Frece, J.; Šušković, J. Immunomodulatory effect of Lactobacillus rhamnosus GG from low fat fresh cheese ”BioAktiv LGG”. Mljekarstvo: Časopis Za Unaprjeđenje Proizv. I Prerade Mlijeka 2004, 54, 5–18. [Google Scholar]
- Goyal, N.; Shukla, G. Probiotic Lactobacillus rhamnosus GG modulates the mucosal immune response in giardia intestinalis-infected BALB/C mice. Dig. Dis. Sci. 2013, 58, 1218–1225. [Google Scholar] [CrossRef]
- Harb, H.; Tol, E.A.F.; Heine, H.; Braaksma, M.; Gross, G.; Overkamp, K.; Hennen, M.; Alrifai, M.; Conrad, M.L.; Renz, H.; et al. Neonatal supplementation of processed supernatant from Lactobacillus rhamnosus GG improves allergic airway inflammation in mice later in life. Clin. Exp. Allergy 2013, 43, 353–364. [Google Scholar] [CrossRef]
- Dimitrijevic, R.; Petrusic, V.; Zivkovic, I.; Dimitrijevic, L.; Ivanovic, N.; Djordjevic, B.; Mathiesen, G. Effects of Lactobacillus rhamnosus LA68 on the immune system of c57bl/6 mice upon oral administration. J. Dairy Res. 2014, 81, 202–207. [Google Scholar] [CrossRef]
- Saliganti, V.; Kapila, R.; Kapila, S. Consumption of probiotic Lactobacillus rhamnosus (MTCC: 5897) containing fermented milk plays a key role in development of the immune system in newborn mice during the suckling-weaning transition. Microbiol. Immunol. 2016, 60, 261–267. [Google Scholar] [CrossRef] [Green Version]
- Pimentel-González, D.J.; Campos-Montiel, R.G.; Lobato-Calleros, C.; Pedroza-Islas, R.; Vernon-Carter, E.J. Encapsulation of Lactobacillus rhamnosus in double emulsions formulated with sweet whey as emulsifier and survival in simulated gastrointestinal conditions. Int. Food Res. J. 2009, 42, 292–297. [Google Scholar] [CrossRef]
- Chai, G.; Governale, L.; McMahon, A.W.; Trinidad, J.P.; Staffa, J.; Murphy, D. Trends of outpatient prescription drug utilization in us children; 2002–2010. Pediatrics 2012, 130, 23–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hilton, E.; Kolakowski, P.; Singer, C.; Smith, M. Efficacy of Lactobacillus GG as a Diarrheal Preventive in Travelers. J. Travel Med. 1997, 4, 41–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vanderhoof, J.A.; Whitney, D.B.; Antonson, D.L.; Hanner, T.L.; Lupo, J.V.; Young, R.J. Lactobacillus GG in the prevention of antibiotic-associated diarrhea in children. J. Pediatrics 1999, 135, 564–568. [Google Scholar] [CrossRef]
- Arvola, T.; Laiho, K.; Torkkeli, S.; Mykkänen, H.; Salminen, S.; Maunula, L.; Isolauri, E. Prophylactic Lactobacillus GG reduces antibiotic-associated diarrhea in children with respiratory infections: A randomized study. Pediatrics 1999, 104, e64. [Google Scholar] [CrossRef] [Green Version]
- Canani, R.B.; Cirillo, P.; Terrin, G.; Cesarano, L.; Spagnuolo, M.I.; De Vincenzo, A.; Albano, F.; Passariello, A.; De Marco, G.; Manguso, F.; et al. Probiotics for treatment of acute diarrhoea in children: Randomised clinical trial of five different preparations. BMJ Brit Med. 2007, 335, 340–342. [Google Scholar] [CrossRef] [Green Version]
- Grandy, G.; Medina, M.; Soria, R.; Terán, C.G.; Araya, M. Probiotics in the treatment of acute rotavirus diarrhoea. A randomized; double-blind; controlled trial using two different probiotic preparations in Bolivian children. BMC Infect. Dis. 2010, 10, 253. [Google Scholar] [CrossRef] [Green Version]
- Hojsak, I.; Snovak, N.; Abdović, S.; Szajewska, H.; Mišak, Z.; Kolaček, S. Lactobacillus GG in the prevention of gastrointestinal and respiratory tract infections in children who attend day care centers: A randomized; double-blind; placebo-controlled trial. Clin. Nutr. 2010, 29, 312–316. [Google Scholar] [CrossRef]
- Wolvers, D.; Antoine, J.M.; Myllyluoma, E.; Schrezenmeir, J.; Szajewska, H.; Rijkers, G.T. Guidance for substantiating the evidence for beneficial effects of probiotics: Prevention and management of infections by probiotics. J. Nutr. 2010, 140, 698–712. [Google Scholar] [CrossRef]
- Goldenberg, J.Z.; Lytvyn, L.; Steurich, J.; Parkin, P.; Mahant, S.; Johnston, B.C. Probiotics for the prevention of pediatric antibiotic-associated diarrhea. Cochrane Database Syst. Rev. 2015, 12, CD004827. [Google Scholar] [CrossRef]
- Szajewska, H.; Kołodziej, M. Systematic review with meta-analysis: Lactobacillus rhamnosus GG in the prevention of antibiotic-associated diarrhoea in children and adults. Aliment. Pharmacol. Ther. 2015, 42, 1149–1157. [Google Scholar] [CrossRef] [Green Version]
- Evans, M.; Salewski, R.P.; Christman, M.C.; Girard, S.A.; Tompkins, T.A. Effectiveness of Lactobacillus helveticus and Lactobacillus rhamnosus for the management of antibiotic-associated diarrhoea in healthy adults: A randomised; double-blind; placebo-controlled trial. Br. J. Nutr. 2016, 116, 94–103. [Google Scholar] [CrossRef] [PubMed]
- Pant, N.; Marcotte, H.; Brüssow, H.; Svensson, L.; Hammarström, L. Effective prophylaxis against rotavirus diarrhea using a combination of Lactobacillus rhamnosus GG and antibodies. BMC Microbiol. 2007, 7, 86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szajewska, H.; Skórka, A.; Ruszczyński, M.; Gieruszczak-Białek, D. Meta-analysis: Lactobacillus GG for treating acute gastroenteritis in children--updated analysis of randomised controlled trials. Aliment. Pharmacol. Ther. 2013, 38, 467–476. [Google Scholar] [CrossRef]
- Arnbjerg, C.J.; Vestad, B.; Hov, J.R.; Pedersen, K.K.; Jespersen, S.; Johannesen, H.H.; Holm, K.; Halvorsen, B.; Fallentin, E.; Hansen, A.E.; et al. Effect of Lactobacillus rhamnosus GG supplementation on intestinal inflammation assessed by PET/MRI scans and gut microbiota composition in HIV-infected individuals. J. Acquir. Immune Defic. Syndr. 2018, 78, 450–457. [Google Scholar] [CrossRef] [PubMed]
- Radford-Smith, G.; Jewell, D.P. The role of cytokines in inflammatory bowel disease. Mediators Inflamm. 1994, 3, 3–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pascual, R.M.; Peters, S.P. The irreversible component of persistent asthma. J. Allergy Clin. Immunol. 2009, 124, 883–890. [Google Scholar] [CrossRef] [Green Version]
- To, T.; Stanojevic, S.; Moores, G.; Gershon, A.S.; Bateman, E.D.; Cruz, A.A.; Boulet, L.P. Global asthma prevalence in adults: Findings from the cross-sectional world health survey. BMC Public Health 2012, 12, 204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koplin, J.J.; Martin, P.E.; Allen, K.J. An update on epidemiology of anaphylaxis in children and adults. Curr. Opin. Allergy Clin. Immunol. 2011, 11, 492–496. [Google Scholar] [CrossRef]
- Liu, M.; Wu, Q.; Wang, M.; Fu, Y.; Wang, J. Lactobacillus rhamnosus GR-1 limits Escherichia coli-induced inflammatory responses via attenuating MYD88-dependent and MYD88-independent pathway activation in bovine endometrial epithelial cells. Inflammation 2016, 39, 1483–1494. [Google Scholar] [CrossRef]
- Machado, V.S.; Bicalho, M.L.; Pereira, R.V.; Caixeta, L.S.; Bittar, J.H.; Oikonomou, G.; Gilbert, R.O.; Bicalho, R.C. The effect of intrauterine administration of mannose or bacteriophage on uterine health and fertility of dairy cows with special focus on Escherichia coli and Arcanobacterium pyogenes. J. Dairy Sci. 2012, 95, 3100–3109. [Google Scholar] [CrossRef] [Green Version]
- Williams, E.J.; Fischer, D.P.; Noakes, D.E.; England, G.C.W.; Rycroft, A.; Dobson, H.; Sheldon, I.M. The relationship between uterine pathogen growth density and ovarian function in the postpartum dairy cow. Theriogenology 2007, 68, 549–559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Navarro, S.; Cossalter, G.; Chiavaroli, C.; Kanda, A.; Fleury, S.; Lazzari, A.; Cazareth, J.; Sparwasser, T.; Dombrowicz, D.; Glaichenhaus, N.; et al. The oral administration of bacterial extracts prevents asthma via the recruitment of regulatory T cells to the airways. Mucosal Immunol. 2011, 4, 53–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sagar, S.; Morgan, M.E.; Chen, S.; Vos, A.P.; Garssen, J.; van Bergenhenegouwen, J.; Boon, L.; Georgiou, N.A.; Kraneveld, A.D.; Folkerts, G. Bifidobacterium breve and Lactobacillus rhamnosus treatment is as effective as budesonide at reducing inflammation in a murine model for chronic asthma. Respir. Res. 2014, 15, 46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michail, S. The role of probiotics in allergic diseases. Allergy Asthma Clin Immunol. 2009, 5, 5. [Google Scholar] [CrossRef] [Green Version]
- Viljanen, M.; Savilahti, E.; Haahtela, T.; Juntunen-Backman, K.; Korpela, R.; Poussa, T.; Tuure, T.; Kuitunen, M. Probiotics in the treatment of atopic eczema/dermatitis syndrome in infants: A double-blind placebo-controlled trial. Allergy 2005, 60, 494–500. [Google Scholar] [CrossRef]
- Yan, F.; Cao, H.; Cover, T.L.; Whitehead, R.; Washington, M.K.; Polk, D.B. Soluble proteins produced by probiotic bacteria regulate intestinal epithelial cell survival and growth. Gastroenterology 2007, 132, 562–575. [Google Scholar] [CrossRef] [Green Version]
- Wu, Q.; Liu, M.C.; Yang, J.; Wang, J.F.; Zhu, Y.H. Lactobacillus rhamnosus GR-1 ameliorates Escherichia coli-induced inflammation and cell damage via attenuation of ASC-independent NLRP3 inflammasome activation. Appl. Environ. Microbiol. 2016, 82, 1173–1182. [Google Scholar] [CrossRef] [Green Version]
- Agerbaek, M.; Gerdes, L.U.; Richelsen, B. Hypocholesterolaemic effect of a new fermented milk product in healthy middle-aged men. Eur. J. Clin. Nutr. 1995, 49, 346–352. [Google Scholar]
- Ishimwe, N.; Daliri, E.B.; Lee, B.H.; Fang, F.; Du, G. The perspective on cholesterol-lowering mechanisms of probiotics. Mol. Nutr. Food Res. 2015, 59, 94–105. [Google Scholar] [CrossRef]
- Yoon, J.A.; Han, D.H.; Noh, J.Y.; Kim, M.H.; Son, G.H.; Kim, K.; Kim, C.J.; Pak, Y.K.; Cho, S. Mealtime shift disturbs circadian rhythmicity along with metabolic and behavioral alterations in mice. PLoS ONE 2012, 7, e44053. [Google Scholar] [CrossRef]
- Chen, S.; Shimada, K.; Zhang, W.; Huang, G.; Crother, T.R.; Arditi, M. IL-17A is proatherogenic in high-fat diet-induced and Chlamydia pneumoniae infection-accelerated atherosclerosis in mice. J. Immunol. 2010, 185, 5619–5627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, M.; Nagpal, R.; Kumar, R.; Hemalatha, R.; Verma, V.; Kumar, A.; Chakraborty, C.; Singh, B.; Marotta, F.; Jain, S.; et al. Cholesterol-lowering probiotics as potential biotherapeutics for metabolic diseases. Exp. Diabetes Res. 2012, 2012, 902917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramasamy, K.; Abdullah, N.; Wong, M.C.V.L.; Karuthan, C.; Ho, Y.W. Bile salt deconjugation and cholesterol removal from media by Lactobacillus strains used as probiotics in chickens. J. Sci. Food Agric. 2010, 90, 65–69. [Google Scholar] [CrossRef] [PubMed]
- Mathara, J.M.; Schillinger, U.; Guigas, C.; Franz, C.; Kutima, P.M.; Mbugua, S.K.; Shin, H.K.; Holzapfel, W.H. Functional characteristics of Lactobacillus spp. from traditional Maasai fermented milk products in Kenya. Int. J. Food Microbiol. 2008, 126, 57–64. [Google Scholar] [CrossRef]
- Park, S.; Kang, J.; Choi, S.; Park, H.; Hwang, E.; Kang, Y.; Kim, A.; Holzapfel, W.; Ji, Y.; Feng, W. Cholesterol-lowering effect of Lactobacillus rhamnosus BFE5264 and its influence on the gut microbiome and propionate level in a murine model. PLoS ONE 2018, 13, e0203150. [Google Scholar] [CrossRef] [Green Version]
- Isolauri, E.; Salminen, S.; Ouwehand, A.C. Probiotics. Best Pract. Res. Clin. Gastroenterol. 2004, 18, 299–313. [Google Scholar] [CrossRef]
- Pagnini, C.; Corleto, V.D.; Martorelli, M.; Lanini, C.; D’Ambra, G.; Di, G.E.; Delle, F.G. Mucosal adhesion and anti-inflammatory effects of Lactobacillus rhamnosus GG in the human colonic mucosa: A proof-of-concept study. World J. Gastroenterol. 2018, 24, 4652–4662. [Google Scholar] [CrossRef]
- Snydman, D.R. The safety of probiotics. Clin. Infect. Dis. 2008, 46, 104–111. [Google Scholar] [CrossRef] [Green Version]
- Sharma, P.; Tomar, S.K.; Goswami, P.; Sangwan, V.; Singh, R. Antibiotic resistance among commercially available probiotics. Int. Food Res. J. 2014, 57, 176–195. [Google Scholar] [CrossRef]
- Aguirre, M.; Collins, M.D. Lactic acid bacteria and human clinical infection. J. Appl. Bacteriol. 1993, 75, 95–107. [Google Scholar] [CrossRef]
- Felten, A.; Barreau, C.; Bizet, C.; Lagrange, P.H.; Philippon, A. Lactobacillus species identification; H2O2 production; and antibiotic resistance and correlation with human clinical status. J. Clin. Microbiol. 1999, 37, 729–733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salminen, S.; Von Wright, A.; Ouwehand, A.; Holzapfel, W.H. Safety Assessment of Starters and Probiotics. In Fermentation and Food Safety; Aspen Publishers: Gaithersburg, MD, USA, 2000; pp. 239–252. [Google Scholar]
- Drago, L.; Rodighiero, V.; Mattina, R.; Toscano, M.; De Vecchi, E. In vitro selection and transferability of antibiotic resistance in the probiotic strain Lactobacillus reuteri DSM 17938. J Chemother. 2011, 23, 371–373. [Google Scholar] [CrossRef] [PubMed]
- Tynkkynen, S.; Singh, K.V.; Varmanen, P. Vancomycin resistance factor of Lactobacillus rhamnosus GG in relation to enterococcal vancomycin resistance (van) genes. Int. J. Food Microbiol. 1998, 41, 195–204. [Google Scholar] [CrossRef]
- Neffe-Skocińska, K.; Rzepkowska, A.; Szydłowska, A.; Kołożyn-Krajewska, D. Trends and possibilities of the use of probiotics in Food Production. In Alternative and Replacement Foods; Academic Press: Cambridge, MA, USA, 2018; pp. 65–94. [Google Scholar]
- Sunny-Roberts, E.O.; Knorr, D. Evaluation of the response of Lactobacillus rhamnosus VTT E-97800 to sucrose-induced osmotic stress. Food Microbiol. 2008, 25, 183–189. [Google Scholar] [CrossRef] [PubMed]
- Cinzia, L.R.; Iole, P.; Fabio, L.; Giuseppe, M.; Cinzia, C. Survival of Lactobacillus rhamnosus probiotic strains in peach jam during storage at different temperatures. Food Sci. Technol. 2013, 33, 652–659. [Google Scholar]
- Alamprese, C.; Foschino, R.; Rossi, M.; Pompei, C.; Corti, S. Effects of Lactobacillus rhamnosus GG addition in ice cream. Int. J. Dairy Technol. 2005, 58, 200–206. [Google Scholar] [CrossRef]
- Morelli, L. In vitro selection of probiotic lactobacilli: A critical appraisal. Curr. Iss. Intest. Microbiol. 2000, 1, 59–67. [Google Scholar]
- Charalampopoulos, D.; Wang, R.; Pandiella, S.S.; Webb, C. Application of cereals and cereal components in functional foods: A review. Int. J. Food Microbiol. 2000, 79, 131–141. [Google Scholar] [CrossRef]
- Patel, H.M.; Pandiella, S.S.; Wang, R.H.; Webb, C. Influence of malt; wheat; and barley extracts on the bile tolerance of selected strains of lactobacilli. Food Microbiol. 2004, 21, 83–89. [Google Scholar] [CrossRef]
- Varcoe, J.; Zook, C.; Sui, J.; Leighton, S.; Busta, F.; Brady, L. Variable response to exogenous Lactobacillus acidophilus NCFM® consumed in different delivery vehicles. J. Appl. Microbiol. 2002, 93, 900–906. [Google Scholar] [CrossRef]
- Champagne, C.P.; Gardner, N.J.; Roy, D. Challenges in the addition of probiotic cultures to foods. Crit. Rev. Food Sci. Nutr. 2005, 45, 61–84. [Google Scholar] [CrossRef] [PubMed]
- Mattila-Sandholm, T.; Myllärinen, P.; Crittenden, R.; Mogensen, G.; Fondén, R.; Saarela, M. Technological challenges for future probiotic foods. Int. Dairy J. 2002, 12, 173–182. [Google Scholar] [CrossRef]
- Granato, D.; Branco, G.F.; Nazzaro, F.; Cruz, A.G.; Faria, J.A.F. Functional foods and nondairy probiotic food development: Trends; concepts; and products. Compr. Rev. Food Sci. 2010, 9, 292–302. [Google Scholar] [CrossRef] [PubMed]
- Holzapfel, W.H.; Haberer, P.; Geisen, R.; Björkroth, J.; Schillinger, U. Taxonomy and important features of probiotic microorganisms in food and nutrition. Am. J. Clin. Nutr. 2001, 73, 365–373. [Google Scholar] [CrossRef] [Green Version]
- McMaster, L.D.; Kokott, S.A.; Reid, S.J.; Abratt, V.R. Use of traditional African fermented beverages as delivery vehicles for Bifidobacterium lactis DSM 10140. Int. J. Food Microbiol. 2005, 102, 231–237. [Google Scholar] [CrossRef] [PubMed]
- Van Tienen, A.; Hullegie, Y.; Hummelen, R.; Hemsworth, J.; Changalucha, J.; Reid, G. Development of a locally sustainable functional food for people living with HIV in Sub-Saharan Africa: Laboratory testing and sensory evaluation. Benef. Microbes 2011, 2, 193–198. [Google Scholar] [CrossRef]
- Innocente, N.; Biasutti, M.; Rita, F.; Brichese, R.; Comi, G.; Iacumin, L. Effect of indigenous Lactobacillus rhamnosus isolated from bovine milk on microbiological characteristics and aromatic profile of traditional yogurt. LWT-Food Sci. Technol. 2016, 66, 158–164. [Google Scholar] [CrossRef]
- Anukam, K.C.; Osazuwa, E.O.; Osadolor, H.B.; Bruce, A.W.; Reid, G. Yogurt containing probiotic Lactobacillus rhamnosus GR-1 and L. reuteri RC-14 helps resolve moderate diarrhea and increases CD4 count in HIV/AIDS patients. J. Clin. Gastroenterol. 2008, 42, 239–243. [Google Scholar] [CrossRef]
- Carlsson, M.; Gustafson, Y.; Haglin, L.; Eriksson, S. The feasibility of serving liquid yoghurt supplemented with probiotic bacteria; Lactobacillus rhamnosus LB 21; and Lactococcus lactis L1A—A pilot study among old people with dementia in a residential care facility. J. Nutr. Health Aging 2009, 13, 813–819. [Google Scholar] [CrossRef]
- Burns, P.; Patrignani, F.; Serrazanetti, D.; Vinderola, G.C.; Reinheimer, J.A.; Lanciotti, R.; Guerzoni, M.E. Probiotic Crescenza cheese containing Lactobacillus casei and Lactobacillus acidophilus manufactured with high-pressure homogenized milk. J. Dairy Sci. 2008, 91, 500–512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scheller, M.; O’Sullivan, D.J. Comparative analysis of an intestinal strain of Bifidobacterium longum and a strain of Bifidobacterium animalis subspecies lactis in cheddar cheese. J. Dairy Sci. 2011, 94, 1122–1131. [Google Scholar] [CrossRef] [PubMed]
- Cuffia, F.; George, G.; Renzulli, P.; Reinheimer, J.; Meinardi, C.; Burns, P. Technological challenges in the production of a probiotic pasta filata soft cheese. LWT-Food Sci. Technol. 2017, 81, 111–117. [Google Scholar] [CrossRef]
- Phillips, M.; Kailasapathy, K.; Tran, L. Viability of commercial probiotic cultures (L. acidophilus; Bifidobacterium sp.; L. casei; L. paracasei and L. rhamnosus) in cheddar cheese. Int. J. Food Microbiol. 2006, 108, 276–280. [Google Scholar] [CrossRef] [PubMed]
- Boylston, T.D.; Vinderola, C.G.; Ghoddusi, H.B.; Reinheimer, J.A. Incorporation of Bifidobacteria into cheeses: Challenges and rewards. Int. Dairy J. 2004, 14, 375–387. [Google Scholar] [CrossRef]
- Cichosz, G.; Aljewicz, M.; Nalepa, B. Viability of the Lactobacillus rhamnosus HN001 probiotic strain in Swiss- and Dutch-type cheese and cheese-like products. J. Food Sci. 2014, 79, 1181–1188. [Google Scholar] [CrossRef]
- Liu, L.; Qu, X.; Xia, Q.; Wang, H.; Chen, P.; Li, X.; Wang, L.; Yang, W. Effect of Lactobacillus rhamnosus on the antioxidant activity of cheddar cheese during ripening and under simulated gastrointestinal digestion. LWT Food Sci. Technol. 2018, 95, 99–106. [Google Scholar] [CrossRef]
- Ningtyas, D.W.; Bhandari, B.; Bansal, N.; Prakash, S. The viability of probiotic Lactobacillus rhamnosus (non-encapsulated and encapsulated) in functional reduced-fat cream cheese and its textural properties during storage. Food Control 2019, 100, 8–16. [Google Scholar] [CrossRef]
- Ogunremi, O.R.; Agrawal, R.; Sanni, A.I. Development of cereal-based functional food using cereal-mix substrate fermented with probiotic strain-Pichia kudriavzevii OG32. Food Sci. Nutrit. 2015, 3, 486–494. [Google Scholar] [CrossRef] [Green Version]
- Mpofu, A.; Linnemann, A.R.; Nout, M.J.R.; Smid, E.J.; Sybesma, W.; Kort, R. Development of a locally sustainable functional food based on mutandabota; a traditional food in Southern Africa. J. Dairy Sci. 2014, 97, 2591–2599. [Google Scholar] [CrossRef] [Green Version]
- Salimei, E.; Capilongo, V.; Simoni, A.; Peiretti, P.G.; Maglieri, C.; Romano, C.A.; Mannina, L.; Coppola, R.; Sorrentino, E. Lactobacillus rhamnosus as additive for maize and sorghum ensiling. J. Agric. Food Chem. 2007, 55, 9600–9607. [Google Scholar] [CrossRef]
- Matejčeková, Z.; Liptáková, D.; Valík, Ľ. Functional probiotic products based on fermented buckwheat with Lactobacillus rhamnosus. LWT-Food Sci. Technol. 2017, 81, 35–41. [Google Scholar] [CrossRef]
- Wacoo, A.P.; Mukisa, I.M.; Meeme, R.; Byakika, S.; Wendiro, D.; Sybesma, W.; Kort, R. Probiotic enrichment and reduction of aflatoxins in a traditional African maize-based fermented food. Nutrients 2019, 11, 265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitra, S.; Ghosh, B.C. Quality characteristics of kefir as a carrier for probiotic lactobacillus rhamnosus gg. Int. J. Dairy Technol. 2020, 73, 384–391. [Google Scholar] [CrossRef]
- Perricone, M.; Bevilacqua, A.; Altieri, C.; Sinigaglia, M.; Corbo, M. Challenges for the production of probiotic fruit juices. Beverages 2015, 1, 95–103. [Google Scholar] [CrossRef] [Green Version]
- Sohail, A.; Turner, M.S.; Coombes, A.; Bhandari, B. The viability of Lactobacillus rhamnosus GG and Lactobacillus acidophilus NCFM following double encapsulation in alginate and maltodextrin. Food Bioproc. Tech. 2013, 6, 2763–2769. [Google Scholar] [CrossRef]
- Worku, K.F.; Kurabachew, H.; Hassen, Y. Probiotication of fruit juices by supplemented culture of Lactobacillus acidophilus. Int. J. Food Sci. Nutr. 2019, 9, 45–48. [Google Scholar]
- Ding, W.K.; Shah, N.P. Survival of free and microencapsulated probiotic bacteria in orange and apple juices. Int. Food Res. J. 2008, 15, 219–232. [Google Scholar]
- Sheehan, V.M.; Ross, P.; Fitzgerald, G.F. Assessing the acid tolerance and the technological robustness of probiotic cultures for fortification in fruit juices. Innov. Food Sci. Emerg. Technol. 2007, 8, 279–284. [Google Scholar] [CrossRef]
- Champagne, C.P.; Gardner, N.J. The growth and recovery of an exopolysaccharide-producing Lactobacillus rhamnosus culture on growth media containing apple juice or molasses. J. Gen. Appl. Microbiol. 2008, 54, 237–241. [Google Scholar] [CrossRef] [Green Version]
- Ying, D.Y.; Sun, J.; Sanguansri, L.; Weerakkody, R.; Augustin, M.A. Enhanced survival of spray-dried microencapsulated Lactobacillus rhamnosus GG in the presence of glucose. J. Food Eng. 2012, 109, 597–602. [Google Scholar] [CrossRef]
- Champagne, C.P.; Raymond, Y.; Gagnon, R. Viability of Lactobacillus rhamnosus R0011 in an apple-based fruit juice under simulated storage conditions at the consumer level. J. Food Sci. 2008, 73, 221–226. [Google Scholar] [CrossRef] [PubMed]
- Bedani, R.; Rossi, E.A.; Isay Saad, S.M. Impact of inulin and okara on Lactobacillus acidophilus LA-5 and Bifidobacterium animalis BB-12 viability in a fermented soy product and probiotic survival under in vitro simulated gastrointestinal conditions. Food Microbiol. 2013, 34, 382–389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farias, N.; Soares, M.; Gouveia, E. Enhancement of the viability of Lactobacillus rhamnosus ATCC 7469 in passion fruit juice: Application of a central composite rotatable design. LWT-Food Sci. Technol. 2016, 71, 149–154. [Google Scholar] [CrossRef]
- Campos, R.C.D.A.B.; Martins, E.M.F.; de Andrade Pires, B.; Peluzio, M.D.C.G.; da Rocha Campos, A.N.; Ramos, A.M.; Júnior, B.R.D.C.L.; de Oliveira Martins, A.D.; da Silva, R.R.; Martins, M.L. In vitro and in vivo resistance of Lactobacillus rhamnosus GG carried by a mixed pineapple (Ananas comosus L. Merril) and jussara (Euterpe edulis Martius) juice to the gastrointestinal tract. Int. Food Res. J. 2019, 116, 1247–1257. [Google Scholar] [CrossRef]
- Homayouni, A.; Azizi, A.; Ehsani, M.R.; Yarmand, M.S.; Razavi, S.H. Effect of microencapsulation and resistant starch on the probiotic survival and sensory properties of synbiotic ice cream. Food Chem. 2008, 111, 50–55. [Google Scholar] [CrossRef]
- Schoug, Å. A Dry Phase of Life: Freeze Drying and Storage Stability of Lactobacillus coryniformis SI3 in Sucrose-Based Formulations; University of Uppsala: Uppsala, Sweden, 2009; pp. 41–46. [Google Scholar]
- Adhikari, K.; Mustapha, A.; Grün, I.U.; Fernando, L. Viability of microencapsulated Bifidobacteria in set yogurt during refrigerated storage. J. Dairy Sci. 2000, 83, 1946–1951. [Google Scholar] [CrossRef]
- Siuta-Cruce, P.; Goulet, J. Improving probiotic survival rates: Microencapsulation preserves the potency of probiotic microorganisms in food systems. Food Technol. 2001, 55, 36–42. [Google Scholar]
- Champagne, C.P.; Fustier, P. Microencapsulation for the improved delivery of bioactive compounds into foods. Curr. Opin. Biotechnol. 2007, 18, 184–190. [Google Scholar] [CrossRef]
- Anal, A.K.; Singh, H. Recent advances in microencapsulation of probiotics for industrial applications and targeted delivery. Trends Food Sci. Technol. 2007, 18, 240–251. [Google Scholar] [CrossRef]
- Rokka, S.; Rantamäki, P. Protecting probiotic bacteria by microencapsulation: Challenges for industrial applications. Eur. Food Res. Technol. Zeitschrift Fur Lebensmittel-Untersuchung Und-Forschung A 2010, 231, 1–12. [Google Scholar] [CrossRef]
- Krasaekoopt, W.; Bhandari, B.; Deeth, H. Evaluation of encapsulation techniques of probiotics for yoghurt. Int. Dairy J. 2003, 13, 3–13. [Google Scholar] [CrossRef]
- Gebara, C.; Chaves, K.S.; Ribeiro, M.C.E.; Souza, F.N.; Grosso, C.R.; Gigante, M.L. Viability of Lactobacillus acidophilus LA5 in pectin–whey protein microparticles during exposure to simulated gastrointestinal conditions. Int. Food Res. J. 2013, 51, 872–878. [Google Scholar] [CrossRef] [Green Version]
- Broeckx, G.; Vandenheuvel, D.; Henkens, T.; Kiekens, S.; van den Broek, M.F.; Lebeer, S.; Kiekens, F. Enhancing the viability of Lactobacillus rhamnosus GG after spray drying and during storage. Int. J. Pharm. 2017, 534, 35–41. [Google Scholar] [CrossRef] [PubMed]
- Jalali, M.; Abedi, D.; Varshosaz, J.; Najjarzadeh, M.; Mirlohi, M.; Tavakoli, N. Stability evaluation of freeze-dried Lactobacillus paracasei subsp. tolerance and Lactobacillus delbrueckii subsp. bulgaricus in oral capsules. Res. Pharm. Sci. 2012, 7, 31–36. [Google Scholar] [PubMed]
- Kailasapathy, K. Microencapsulation of probiotic bacteria: Technology and potential applications. Curr. Iss. Intest. Microbiol. 2002, 3, 39–48. [Google Scholar]
- Corcoran, B.M.; Ross, R.P.; Fitzgerald, G.F.; Stanton, C. Comparative survival of probiotic lactobacilli spray-dried in the presence of prebiotic substances. J. Appl. Microbiol. 2004, 96, 1024–1039. [Google Scholar] [CrossRef]
- Broeckx, G.; Kiekens, S.; Jokicevic, K.; Byl, E.; Henkens, T.; Kiekens, F.; Vandenheuvel, D.; Lebeer, S. Effects of initial cell concentration; growth phase; and process parameters on the viability of Lactobacillus rhamnosus GG after spray drying. Dry. Technol. 2019, 38, 1474–1492. [Google Scholar] [CrossRef]
- Kiekens, S.; Vandenheuvel, D.; Broeckx Géraldine, C.I.; Allonsius, C.; De Boeck, I.; Thys, S.; Timmermans, J.P.; Kiekens, F.; Lebeer, S. Impact of spray-drying on the pili of Lactobacillus rhamnosus GG. Microb. Biotechnol. 2019, 12, 849–855. [Google Scholar] [CrossRef] [Green Version]
- Shaharuddin, S.; Muhamad, I.I. Microencapsulation of alginate-immobilized bagasse with Lactobacillus rhamnosus NRRL 442: Enhancement of survivability and thermotolerance. Carbohydr. Polym. 2015, 119, 173–181. [Google Scholar] [CrossRef]
- Li, C.; Wang, C.L.; Sun, Y.; Li, A.L.; Liu, F.; Meng, X.C. Microencapsulation of Lactobacillus rhamnosus GG by transglutaminase cross-linked soy protein isolate to improve survival in simulated gastrointestinal conditions and yoghurt. J. Food Sci. 2016, 81, 1726–1734. [Google Scholar] [CrossRef]
- Song, H.; Zhang, J.; Qu, J.; Liu, J.; Yin, P.; Zhang, G.; Shang, D. Lactobacillus rhamnosus GG microcapsules inhibit Escherichia coli biofilm formation in coculture. Biotechnol. Lett. 2019, 41, 1007–1014. [Google Scholar] [CrossRef] [PubMed]
- Lavari, L.; Burns, P.; Páez, R.; Reinheimer, J.; Vinderola, G. Study of the effects of spray drying in whey-starch on the probiotic capacity of Lactobacillus rhamnosus 64 in the gut of mice. J. Appl. Microbiol. 2017, 123, 992–1002. [Google Scholar] [CrossRef] [PubMed]
- Azizi, S.; Rezazadeh-Bari, M.; Almasi, H.; Amiri, S. Microencapsulation of Lactobacillus rhamnosus using sesame protein isolate: Effect of encapsulation method and transglutaminase. Food Biosci. 2021, 41, 101012. [Google Scholar] [CrossRef]
- Lai, K.; How, Y.; Pui, L. Microencapsulation of Lactobacillus rhamnosus GG with flaxseed mucilage using co-extrusion technique. J. Microencapsul. 2021, 38, 134–148. [Google Scholar] [CrossRef]
- Barajas-Álvarez, P.; González-Ávila, M.; Espinosa-Andrews, H. Microencapsulation of Lactobacillus rhamnosus HN001 by spray drying and its evaluation under gastrointestinal and storage conditions. LWT Food Sci. Technol. 2022, 153, 112485. [Google Scholar] [CrossRef]
- Sleator, R.D.; Hill, C. Patho-biotechnology: Using bad bugs to do good things. Curr. Opin. Biotechnol. 2006, 17, 211–216. [Google Scholar] [CrossRef]
- Sleator, R.D.; Hill, C. “Bioengineered bugs”—A patho-biotechnology approach to probiotic research and applications. Med. Hypotheses 2008, 70, 167–169. [Google Scholar] [CrossRef]
- Kumar, M.; Yadav, A.K.; Verma, V.; Singh, B.; Mal, G.; Nagpal, R.; Hemalatha, R. Bioengineered probiotics as a new hope for health and diseases: An overview of potential prospects. Future Microbiol. 2016, 11, 585–600. [Google Scholar] [CrossRef]
- Mathipa, M.G.; Thantsha, M.S. Probiotic engineering: Towards development of robust probiotic strains with enhanced functional properties and for targeted control of enteric pathogens. Gut Pathog. 2017, 9, 28. [Google Scholar] [CrossRef]
- Roberts, M.; Chatfield, S.; Pickard, D.; Li, J.; Bacon, A. Comparison of abilities of Salmonella enterica serovar Typhimurium aroA aroD and aroA htrA mutants to act as live vectors. Infect. Immun. 2000, 68, 6041–6043. [Google Scholar] [CrossRef] [Green Version]
- Saklani-Jusforgues, H.; Fontan, E.; Soussi, N.; Milon, G.; Goossens, P.L. Enteral immunization with attenuated recombinant Listeria monocytogenes as a live vaccine vector: Organ-dependent dynamics of CD4 T lymphocytes reactive to a leishmania major tracer epitope. Infect. Immun. 2003, 71, 1083–1090. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stevenson, A.; Roberts, M. Use of Bordetella bronchiseptica and Bordetella pertussis as live vaccines and vectors for heterologous antigens. FEMS Immunol. Med. Microbiol. 2003, 37, 121–128. [Google Scholar] [CrossRef] [Green Version]
- Petrova, M.I.; Imholz, N.C.; Verhoeven, T.L.; Balzarini, J.; Van Damme, E.J.; Schols, D.; Vanderleyden, J.; Lebeer, S. Lectin-like molecules of Lactobacillus rhamnosus GG inhibit pathogenic Escherichia coli and Salmonella biofilm formation. PLoS ONE 2016, 11, e0161337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petrova, M.I.; Lievens, E.; Verhoeven, T.L.A.; Macklaim, J.M.; Gloor, G.; Schols, D.; Vanderleyden, J.; Reid, G.; Lebeer, S. The lectin-like protein 1 in Lactobacillus rhamnosus GR-1 mediates tissue-specific adherence to vaginal epithelium and inhibits urogenital pathogens. Sci. Rep. 2016, 6, 37437. [Google Scholar] [CrossRef]
- Spacova, I.; Lievens, E.; Verhoeven, T.; Steenackers, H.; Vanderleyden, J.; Lebeer, S.; Petrova, M.I. Expression of fluorescent proteins in Lactobacillus rhamnosus to study host–microbe and microbe–microbe interactions. Microb. Biotechnol. 2018, 11, 317–331. [Google Scholar] [CrossRef] [Green Version]
- Günaydın, G.; Zhang, R.; Hammarström, L.; Marcotte, H. Engineered Lactobacillus rhamnosus gg expressing IgG-binding domains of protein G: Capture of hyperimmune bovine colostrum antibodies and protection against diarrhea in a mouse pup rotavirus infection model. Vaccine 2014, 32, 470–477. [Google Scholar] [CrossRef]
- Beltran, S.; Munoz-Bergmann, C.A.; Elola-Lopez, A.; Quintana, J.; Segovia, C.; Trombert, A.N. The expression of heterologous MAM-7 in Lactobacillus rhamnosus reduces its intrinsic capacity to inhibit colonization of pathogen Vibrio parahaemolyticus in vitro. Biol. Res. 2016, 49, 2. [Google Scholar] [CrossRef] [Green Version]
- Petrova, M.I.; van den Broek, M.F.; Spacova, I.; Verhoeven, T.L.A.; Balzarini, J.; Vanderleyden, J.; Schols, D.; Lebeer, S. Engineering Lactobacillus rhamnosus GG and GR-1 to express HIV-inhibiting Griffithsin. Int. J. Antimicrob. Agents 2018, 52, 599–607. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mathipa-Mdakane, M.G.; Thantsha, M.S. Lacticaseibacillus rhamnosus: A Suitable Candidate for the Construction of Novel Bioengineered Probiotic Strains for Targeted Pathogen Control. Foods 2022, 11, 785. https://0-doi-org.brum.beds.ac.uk/10.3390/foods11060785
Mathipa-Mdakane MG, Thantsha MS. Lacticaseibacillus rhamnosus: A Suitable Candidate for the Construction of Novel Bioengineered Probiotic Strains for Targeted Pathogen Control. Foods. 2022; 11(6):785. https://0-doi-org.brum.beds.ac.uk/10.3390/foods11060785
Chicago/Turabian StyleMathipa-Mdakane, Moloko G., and Mapitsi S. Thantsha. 2022. "Lacticaseibacillus rhamnosus: A Suitable Candidate for the Construction of Novel Bioengineered Probiotic Strains for Targeted Pathogen Control" Foods 11, no. 6: 785. https://0-doi-org.brum.beds.ac.uk/10.3390/foods11060785