
Nutritional Compositions, Phenolic Contents, and Antioxidant Potentials of Ten Original Lineage Beans in Thailand

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Selection, Preparation, and Extraction
2.2. Determination of Nutrients
2.2.1. Proximate Compositions
2.2.2. Minerals
2.2.3. Vitamins
2.3. Determination of Total Phenolic Contents
2.4. Determination of Antioxidant Activities
2.5. Statistical Analysis
3. Results
3.1. Nutritive Values
3.1.1. Proximate Compositions
3.1.2. Vitamin Contents
3.1.3. Mineral Contents
3.2. Total Phenolic Contents and Antioxidant Activities
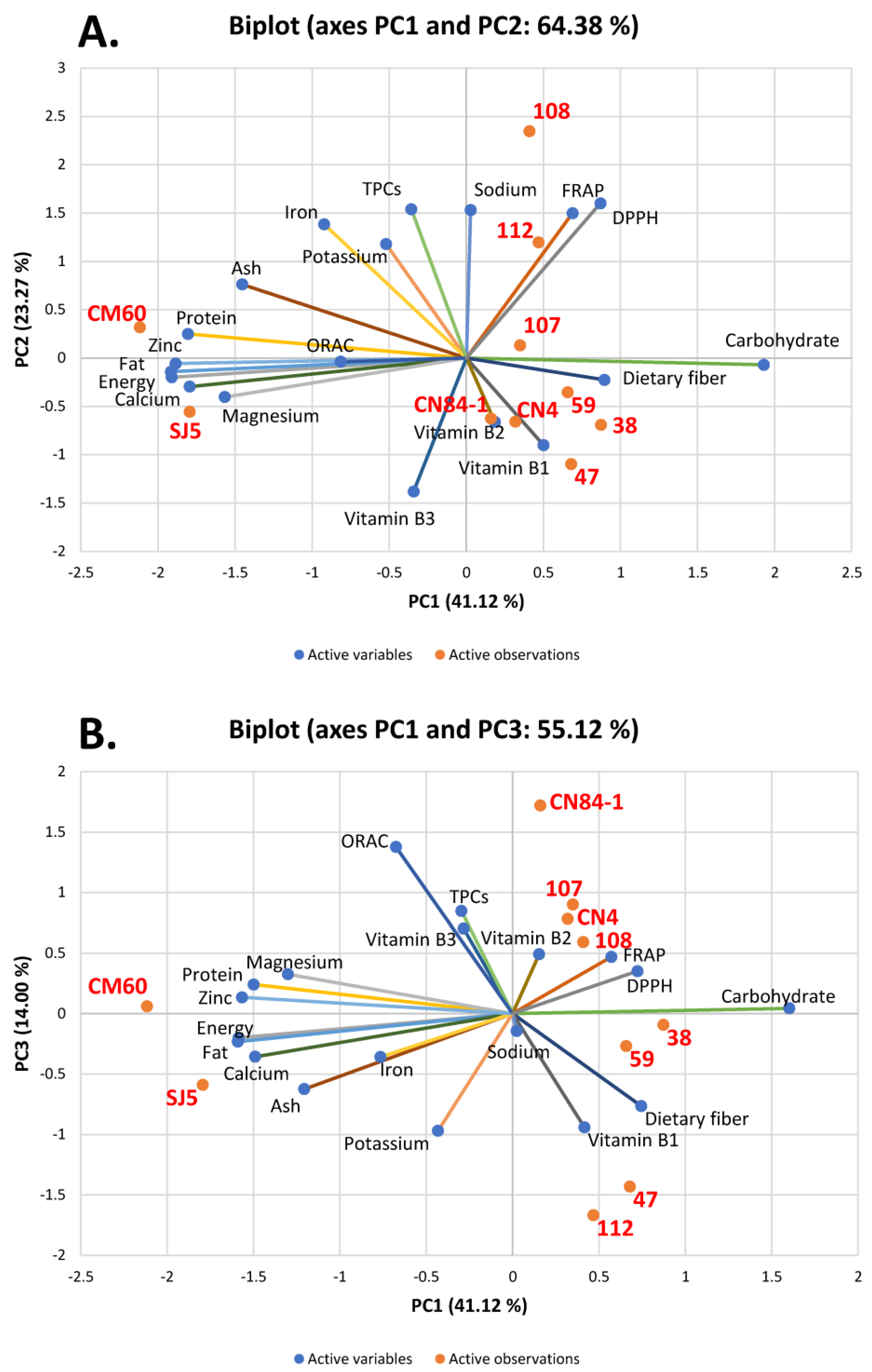
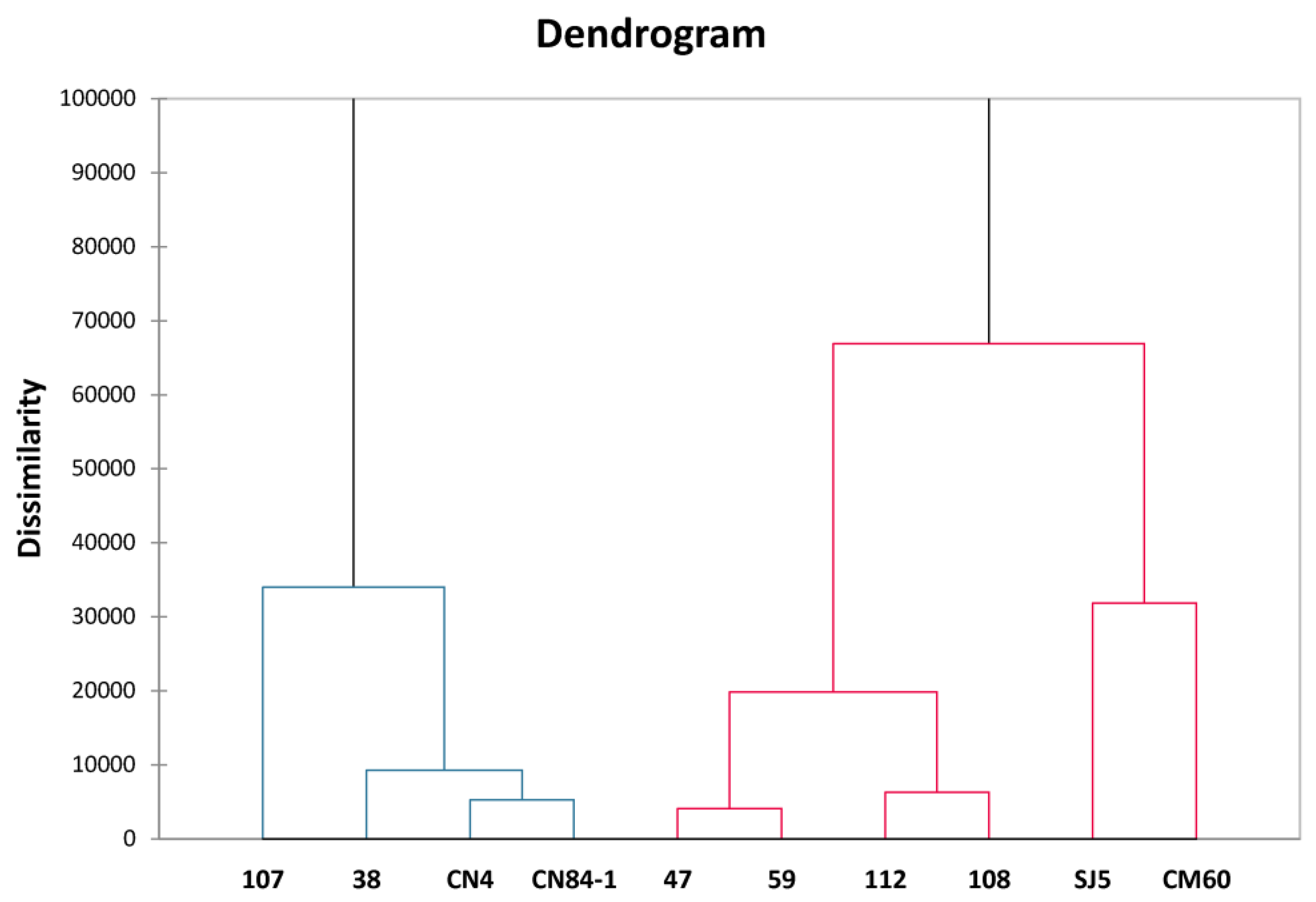
3.3. Correlation Analysis by Principal Component Analysis (PCA) and Hierarchical Cluster Analysis (HCA)
4. Discussion
4.1. Nutritional Compositions
4.2. Total Phenolic Contents and Antioxidant Potentials
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- World Health Organization (WHO): Malnutrition. 2021. Available online: https://www.who.int/news-room/fact-sheets/detail/malnutrition (accessed on 4 May 2022).
- World Health Organization; United Nations Children’s Fund (UNICEF); World Bank. Levels and Trends in Child Malnutrition: UNICEF/WHO/The World Bank Group Joint Child Malnutrition Estimates: Key Findings of the 2021 Edition; World Health Organization: Geneva, Switzerland, 2021.
- Global Nutrition Report: The State of Global Nutrition; Development Initiatives: Bristol, UK, 2021; Available online: https://globalnutritionreport.org/ (accessed on 27 May 2022).
- Wells, J.C.K.; Marphatia, A.A.; Amable, G.; Siervo, M.; Friis, H.; Miranda, J.J.; Haisma, H.H.; Raubenheimer, D. The future of human malnutrition: Rebalancing agency for better nutritional health. Glob. Health 2021, 17, 119. [Google Scholar] [CrossRef]
- Crosser, N. Plant-Based Meat, Eggs, and Dairy: 2019 U.S. State of the Industry Report. 1–72; Good Food Institute: Washington, DC, USA, 2020; Available online: https://www.ditchley.com/sites/default/files/2019-10/SOI-Report-Plant-Based.pdf (accessed on 28 May 2022).
- McClements, D.J.; Grossmann, L. A brief review of the science behind the design of healthy and sustainable plant-based foods. Npj Sci. Food 2021, 5, 17. [Google Scholar] [CrossRef]
- Semba, R.D.; Ramsing, R.; Rahman, N.; Kraemer, K.; Bloem, M.W. Legumes as a sustainable source of protein in human diets. Glob. Food Secur. 2021, 28, 100520. [Google Scholar] [CrossRef]
- Rawal, V.; Charrondiere, R.; Xipsiti, M.; Grande, F. Pulses: Nutritional Benefits and Consumption Patterns. In The Global Economy of Pulses; Rawal, V., Navarro, D.K., Eds.; FAO: Rome, Italy, 2019; pp. 9–19. [Google Scholar]
- Geraldo, R.; Santos, C.S.; Pinto, E.; Vasconcelos, M.W. Widening the Perspectives for Legume Consumption: The Case of Bioactive Non-nutrients. Front. Plant Sci. 2022, 13, 2054. [Google Scholar] [CrossRef]
- Corzo-Ríos, L.J.; Sánchez-Chino, X.M.; Cardador-Martínez, A.; Martínez-Herrera, J.; Jiménez-Martínez, C. Effect of cooking on nutritional and non-nutritional compounds in two species of Phaseolus (P. vulgaris and P. coccineus) cultivated in Mexico. Int. J. Gastron. Food Sci. 2020, 20, 100206. [Google Scholar] [CrossRef]
- Ciabotti, S.; Silva, A.C.B.B.; Juhasz, A.C.P.; Mendonça, C.D.; Tavano, O.L.; Mandarino, J.M.G.; Gonçalves, C.A.A. Chemical composition, protein profile, and isoflavones content in soybean genotypes with different seed coat colors. Int. Food Res. J. 2016, 23, 621–629. [Google Scholar]
- Singh, B.; Singh, J.P.; Shevkani, K.; Singh, N.; Kaur, A. Bioactive constituents in pulses and their health benefits. J. Food Sci. Technol. 2017, 54, 858–870. [Google Scholar] [CrossRef] [Green Version]
- Chaieb, N.; González, J.L.; López-Mesas, M.; Bouslama, M.; Valiente, M. Polyphenols content and antioxidant capacity of thirteen faba bean (Vicia faba L.) genotypes cultivated in Tunisia. Food Res. Int. 2011, 44, 970–977. [Google Scholar] [CrossRef]
- Kan, L.; Nie, S.; Hu, J.; Wang, S.; Cui, S.W.; Li, Y.; Xu, S.; Wu, Y.; Wang, J.; Bai, Z.; et al. Nutrients, phytochemicals and antioxidant activities of 26 kidney bean cultivars. Food Chem. Toxicol. 2017, 108, 467–477. [Google Scholar] [CrossRef]
- Latimer, G.W. Official Method of Analysis of AOAC International, 21st ed.; AOAC International: Rockville, ML, USA, 2019. [Google Scholar]
- Sritalahareuthai, V.; Aursalung, A.; On-nom, N.; Temviriyanukul, P.; Charoenkiatkul, S.; Suttisansanee, U. Nutritional composition of conserved Kadsura spp. plants in Northern Thailand. Heliyon 2020, 6, e04451. [Google Scholar] [CrossRef]
- Jones, D.B. Factors for Converting Percentages of Nitrogen in Foods and Feeds into Percentages of Proteins; U.S. Dept. of Agriculture: Washington, DC, USA, 1941.
- Mariotti, F.; Tomé, D.; Mirand, P.P. Converting Nitrogen into Protein—Beyond 6.25 and Jones’ Factors. Crit. Rev. Food Sci. Nutr. 2008, 48, 177–184. [Google Scholar] [CrossRef] [PubMed]
- Wimalasiri, P.; Wills, R.B. Simultaneous analysis of thiamin and riboflavin in foods by high-performance liquid chromatography. J. Chromatogr. 1985, 318, 412–416. [Google Scholar] [CrossRef]
- Ward, C.M.; Trenerry, V.C. The determination of niacin in cereals, meat and selected foods by capillary electrophoresis and high performance liquid chromatography. Food Chem. 1997, 60, 667–674. [Google Scholar] [CrossRef]
- Windahl, K.L.; Trenerry, V.C.; Ward, C.M. The determination of niacin in selected foods by capillary electrophoresis and high performance liquid chromatography: Acid extraction. Food Chem. 1999, 65, 263–270. [Google Scholar] [CrossRef]
- Sripum, C.; Kukreja, R.; Charoenkiatkul, S.; Kriengsinyos, W.; Suttisansanee, U. The effect of extraction conditions on antioxidant activities and total phenolic contents of different processed Thai Jasmine rice. Int. Food Res. J. 2017, 24, 1644–1650. [Google Scholar]
- Wannasaksri, W.; On-Nom, N.; Chupeerach, C.; Temviriyanukul, P.; Charoenkiatkul, S.; Suttisansanee, U. In Vitro Phytotherapeutic Properties of Aqueous Extracted Adenia viridiflora Craib. towards Civilization Diseases. Molecules 2021, 26, 1082. [Google Scholar] [CrossRef]
- Thai Recommended Daily Intakes (Thai RDIs), Appendix No.3. In MOPH Notification No. 182 B.E. 2541 Re: Nutrition Labelling; Royal Thai Government Gazette: Nonthaburi, Thailand, 1998.
- MOPH Notification No. 392 B.E. 2561 Re: Nutrition Labelling (No.3); Royal Thai Government Gazette: Nonthaburi, Thailand, 2018.
- U.S. Department of Agriculture; U.S. Department of Health and Human Services. Dietary Guidelines for Americans, 2020–2025, 9th ed.; U.S. Department of Agriculture: Washington, DC, USA; U.S. Department of Health and Human Services: Atlanta, GA, USA, 2020. Available online: DietaryGuidelines.gov (accessed on 3 May 2022).
- Bowman, S.A.; Clemens, J.C.; Shimizu, M.; Friday, J.E.; Moshfegh, A.J. Food Patterns Equivalents Database 2015–2016: Methodology and User Guide; Food Surveys Research Group, Beltsville Human Nutrition Research Center, Agricultural Research Service, U.S. Department of Agriculture: Beltsville, MD, USA, 2018. Available online: https://www.ars.usda.gov/ARSUserFiles/80400530/pdf/fped/FPED_1516.pdf (accessed on 28 May 2022).
- Judprasong, K.; Puwastien, P.; Rojroongwasinkul, N.; Nitithamyong, A.; Sridonpai, P.; Somjai, A.; Institute of Nutrition, Mahidol University (2015). Thai Food Composition Database, Online Version 2, September, 2018, Thailand. Available online: http://www.inmu.mahidol.ac.th/thaifcd (accessed on 2 April 2022).
- Joint FAO/WHO/UNU Expert Consultation on Protein and Amino Acid Requirements in Human Nutrition (2002: Geneva, Switzerland), Food and Agriculture Organization of the United Nations, World Health Organization & United Nations University, 2007. Available online: https://apps.who.int/iris/handle/10665/43411 (accessed on 28 May 2022).
- Zhang, T.; Dou, W.; Zhang, X.; Zhao, Y.; Zhang, Y.; Jiang, L.; Sui, X. The development history and recent updates on soy protein-based meat alternatives. Trends Food Sci. Technol. 2021, 109, 702–710. [Google Scholar] [CrossRef]
- Zaaboul, F.; Zhao, Q.; Xu, Y.; Liu, Y. Soybean oil bodies: A review on composition, properties, food applications, and future research aspects. Food Hydrocoll. 2022, 124, 107296. [Google Scholar] [CrossRef]
- U.S. Department of Agriculture (USDA), Agricultural Research Service. FoodData Central: Foundation Foods. Version Current: April, 2021. Available online: Fdc.nal.usda.gov (accessed on 15 May 2022).
- Alcázar-Valle, M.; Lugo-Cervantes, E.; Mojica, L.; Morales-Hernández, N.; Reyes-Ramírez, H.; Enríquez-Vara, J.N.; García-Morales, S. Bioactive Compounds, Antioxidant Activity, and Antinutritional Content of Legumes: A Comparison between Four Phaseolus Species. Molecules 2020, 25, 3528. [Google Scholar] [CrossRef]
- Okekunle, M.O.; Adebowale, K.O.; Olu-Owolabi, B.I.; Lamprecht, A. Physicochemical, morphological and thermal properties of oxidized starches from Lima bean (Phaseolus lunatus). Sci. Afr. 2020, 8, e00432. [Google Scholar] [CrossRef]
- Phillips, K.M.; Haytowitz, D.B.; Pehrsson, P.R. Implications of two different methods for analyzing total dietary fiber in foods for food composition databases. J. Food Compos. Anal. 2019, 84, 103253. [Google Scholar] [CrossRef] [Green Version]
- Dhingra, D.; Michael, M.; Rajput, H.; Patil, R.T. Dietary fibre in foods: A review. J. Food Sci. Technol. 2012, 49, 255–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- FAO/INFOODS Database for Pulses on Dry Matter Basis Version 1.0—PulsesDM1.0. Rome. 2017. Available online: https://www.fao.org/infoods/infoods/tables-and-databases/faoinfoods-databases/en/ (accessed on 22 May 2022).
- Bepary, R.H.; Wadikar, D.D.; Neog, S.B.; Patki, P.E. Studies on physico-chemical and cooking characteristics of rice bean varieties grown in NE region of India. J. Food Sci. Technol. 2017, 54, 973–986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshida, H.; Tomiyama, Y.; Yoshida, N.; Shibata, K.; Mizushina, Y. Regiospecific profiles of fatty acids in triacylglycerols and phospholipids from Adzuki beans (Vigna angularis). Nutrients 2010, 2, 49–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ganesan, K.; Xu, B. Polyphenol-Rich Dry Common Beans (Phaseolus vulgaris L.) and Their Health Benefits. Int. J. Mol. Sci. 2017, 18, 2331. [Google Scholar] [CrossRef] [Green Version]
- Cardador-Martínez, A.; Castaño-Tostado, E.; Loarca-Piña, G. Antimutagenic activity of natural phenolic compounds present in the common bean (Phaseolus vulgaris) against aflatoxin B1. Food Addit. Contam. 2002, 19, 62–69. [Google Scholar] [CrossRef]
- Luthria, D.L.; Pastor-Corrales, M.A. Phenolic acids content of fifteen dry edible bean (Phaseolus vulgaris L.) varieties. J. Food Compos. Anal. 2006, 19, 205–211. [Google Scholar] [CrossRef]
- Oomah, B.D.; Cardador-Martínez, A.; Loarca-Piña, G. Phenolics and antioxidative activities in common beans (Phaseolus vulgaris L). J. Sci. Food Agric. 2005, 85, 935–942. [Google Scholar] [CrossRef]
- Ha, P.T.T.; Tran, N.; Tram, N.; Kha, V. Total phenolic, total flavonoid contents and antioxidant potential of Common Bean (Phaseolus vulgaris L.) in Vietnam. AIMS Agric. Food 2020, 5, 635–648. [Google Scholar] [CrossRef]
- Xu, B.J.; Chang, S.K. Total phenolic content and antioxidant properties of eclipse black beans (Phaseolus vulgaris L.) as affected by processing methods. J. Food Sci. 2008, 73, H19–H27. [Google Scholar] [CrossRef]
- Zhao, Y.; Du, S.K.; Wang, H.; Cai, M. In Vitro antioxidant activity of extracts from common legumes. Food Chem. 2014, 152, 462–466. [Google Scholar] [CrossRef] [PubMed]
- Agostini-Costa, T.; Teodoro, A.; Alves, R.B.N.; Braga, L.; Ribeiro, I.; Silva, J.; Quintana, L.; Burle, M. Total phenolics, flavonoids, tannins and antioxidant activity of lima beans conserved in a Brazilian Genebank. Cienc. Rural. 2015, 45, 335–341. [Google Scholar] [CrossRef] [Green Version]
- Weng, Y.; Ravelombola, W.; Yang, W.; Qin, J.; Zhou, W.; Wang, Y.-J.; Mou, B.; Shi, A. Screening of Seed Soluble Sugar Content in Cowpea (Vigna unguiculata (L.) Walp). Am. J. Plant Sci. 2018, 09, 1455–1466. [Google Scholar] [CrossRef] [Green Version]
- Yao, Y.; Cheng, X.Z.; Wang, L.X.; Wang, S.H.; Ren, G. Major phenolic compounds, antioxidant capacity and antidiabetic potential of rice bean (Vigna umbellata L.) in China. Int. J. Mol. Sci. 2012, 13, 2707–2716. [Google Scholar] [CrossRef] [PubMed]
- Kaga, A.; Ishii, T.; Tsukimoto, K.; Tokoro, E.; Kamijima, O. Comparative molecular mapping in Ceratotropis species using an interspecific cross between azuki bean (Vigna angularis) and rice bean (V. umbellata). Theor. Appl. Genet. 2000, 100, 207–213. [Google Scholar] [CrossRef]
- Isemura, T.; Tomooka, N.; Kaga, A.; Vaughan, D. Comparison of the Pattern of Crop Domestication between Two Asian Beans, Azuki Bean (Vigna angularis) and Rice Bean (V. umbellata). Jpn. Agric. Res. Q. JARQ 2011, 45, 23–30. [Google Scholar] [CrossRef] [Green Version]
- Yao, Y.; Cheng, X.; Wang, S.; Wang, L.; Ren, G. Influence of altitudinal variation on the antioxidant and antidiabetic potential of azuki bean (Vigna angularis). Int. J. Food Sci. Nutr. 2012, 63, 117–124. [Google Scholar] [CrossRef]
- Girish, T.K.; Pratape, V.M.; Prasada Rao, U.J.S. Nutrient distribution, phenolic acid composition, antioxidant and alpha-glucosidase inhibitory potentials of black gram (Vigna mungo L.) and its milled by-products. Food Res. Int. 2012, 46, 370–377. [Google Scholar] [CrossRef]
- Kim, J.K.; Kim, E.H.; Lee, O.K.; Park, S.Y.; Lee, B.; Kim, S.H.; Park, I.; Chung, I.M. Variation and correlation analysis of phenolic compounds in mungbean (Vigna radiata L.) varieties. Food Chem. 2013, 141, 2988–2997. [Google Scholar] [CrossRef]
- Jia, Y.; Ma, Y.; Zou, P.; Cheng, G.; Zhou, J.; Cai, S. Effects of Different Oligochitosans on Isoflavone Metabolites, Antioxidant Activity, and Isoflavone Biosynthetic Genes in Soybean (Glycine max) Seeds during Germination. J. Agric. Food Chem. 2019, 67, 4652–4661. [Google Scholar] [CrossRef]
- Malencić, D.; Popović, M.; Miladinović, J. Phenolic content and antioxidant properties of soybean (Glycine max (L.) Merr.) seeds. Molecules 2007, 12, 576–581. [Google Scholar] [CrossRef] [PubMed]
- Suttisansanee, U.; Thiyajai, P.; Chalermchaiwat, P.; Wongwathanarat, K.; Pruesapan, K.; Charoenkiatkul, S.; Temviriyanukul, P. Phytochemicals and In Vitro Bioactivities of Aqueous Ethanolic Extracts from Common Vegetables in Thai Food. Plants 2021, 10, 1563. [Google Scholar] [CrossRef] [PubMed]
- Gohara, A.; Souza, A.; Gomes, S.; Souza, N.; Visentainer, J.; Matsushita, M. Nutritional and bioactive compounds of adzuki bean cultivars using chemometric approach. Ciênc. Agrotecnol. 2016, 40, 104–113. [Google Scholar] [CrossRef]
- Kumar, Y.; Basu, S.; Goswami, D.; Devi, M.; Shivhare, U.S.; Vishwakarma, R.K. Anti-nutritional compounds in pulses: Implications and alleviation methods. Legume Sci. 2022, 4, e111. [Google Scholar] [CrossRef]
- Sharma, A. A review on traditional technology and safety challenges with regard to antinutrients in legume foods. J. Food Sci. Technol. 2021, 58, 2863–2883. [Google Scholar] [CrossRef] [PubMed]
- De Angelis, D.; Pasqualone, A.; Allegretta, I.; Porfido, C.; Terzano, R.; Squeo, G.; Summo, C. Antinutritional factors, mineral composition and functional properties of dry fractionated flours as influenced by the type of pulse. Heliyon 2021, 7, e06177. [Google Scholar] [CrossRef]
- Sharma, A.; Kumari, S.; Nout, M.J.R.; Sarkar, P.K. Minimization of Antinutrients in Idli by Using Response Surface Process Optimization. J. Food Processing Preserv. 2017, 41, e13099. [Google Scholar] [CrossRef]
- Mohan, V.R.; Tresina, P.S.; Daffodil, E.D. Antinutritional Factors in Legume Seeds: Characteristics and Determination. In Encyclopedia of Food and Health; Academic Press: New York, NY, USA, 2016. [Google Scholar]
- Das, G.; Sharma, A.; Sarkar, P.K. Conventional and emerging processing techniques for the post-harvest reduction of antinutrients in edible legumes. Appl. Food Res. 2022, 2, 100112. [Google Scholar] [CrossRef]
- Shimelis, E.A.; Rakshit, S.K. Effect of processing on antinutrients and in vitro protein digestibility of kidney bean (Phaseolus vulgaris L.) varieties grown in East Africa. Food Chem. 2007, 103, 161–172. [Google Scholar] [CrossRef]
- Acquah, C.; Ohemeng-Boahen, G.; Power, K.A.; Tosh, S.M. The Effect of Processing on Bioactive Compounds and Nutritional Qualities of Pulses in Meeting the Sustainable Development Goal 2. Front. Sustain. Food Syst. 2021, 5, 681662. [Google Scholar] [CrossRef]
- Huertas, R.; William Allwood, J.; Hancock, R.D.; Stewart, D. Iron and zinc bioavailability in common bean (Phaseolus vulgaris) is dependent on chemical composition and cooking method. Food Chem. 2022, 387, 132900. [Google Scholar] [CrossRef] [PubMed]
- Didinger, C.; Thompson, H.J. Defining Nutritional and Functional Niches of Legumes: A Call for Clarity to Distinguish a Future Role for Pulses in the Dietary Guidelines for Americans. Nutrients 2021, 13, 1100. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, D.C.; Webster, A.; Garrison, B. Terminology Matters: Advancing Science to Define an Optimal Pulse Intake. Nutrients 2022, 14, 655. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Scientific Name | English Common Name | Local Thai Name | Cultivar | Voucher Specimen (BK No.) |
|---|---|---|---|---|
| Phaseolus lunatus L. | Lima bean | Thua Lima | 38 | 083065 |
| Thua Kao | 47 | 083066 | ||
| Thua Boy | 59 | 083064 | ||
| Phaseolus vulgaris L. | Red kidney bean | Thua Daeng Luang | 112 | 083063 |
| Vigna umbellata (Thunb.) Ohwi and H.Ohashi | Red bean, rice bean | Thua Nio Nang Daeng | 107 | 083067 |
| Vigna angularis (Wild.) Ohwi and Ohashi | Azuki bean | Thua Azuki | 108 | 083062 |
| Vigna mungo (L.) Hepper | Black gram, mungo bean | Thua Khiao Phio Dam | CN4 | 083061 |
| Vigna radiata (L.) Wilczek | Mung bean | Thua Khiao Phio Man | CN84-1 | 083072 |
| Glycine max (L.) Merrill | Soybean | Thua Lueang | SJ5 | 083060 |
| CM60 | 083057 |
| Cultivar | Energy (kcal) | Protein (g) | Fat (g) | Carbohydrate (g) | Dietary Fiber (g) | Ash (g) |
|---|---|---|---|---|---|---|
| 38 | 392.28 ± 0.54 c | 22.47 ± 0.07 g | 1.27 ± 0.12 d | 72.75 ± 0.19 c | 21.34 ± 0.39 c | 3.51 ± 0.01 i |
| 47 | 391.47 ± 1.39 cd | 16.62 ± 0.25 i | 1.72 ± 0.28 c | 77.39 ± 0.54 a | 21.93 ± 0.17 b | 4.28 ± 0.00 f |
| 59 | 389.75 ± 0.39 d | 19.20 ± 0.27 h | 1.33 ± 0.55 d | 75.25 ± 0.25 b | 19.21 ± 0.28 f | 4.22 ± 0.35 f |
| 112 | 386.39 ± 1.60 e | 23.00 ± 0.03 f | 1.38 ± 0.26 cd | 70.48 ± 0.14 e | 20.93 ± 0.23 d | 5.13 ± 0.09 c |
| 107 | 386.76 ± 0.12 e | 22.59 ± 0.07 g | 1.04 ± 0.02 de | 71.75 ± 0.08 d | 19.95 ± 0.01 e | 4.62 ± 0.01 d |
| 108 | 386.07 ± 0.60 e | 26.95 ± 0.11 c | 0.77 ± 0.12 e | 67.84 ± 0.01 g | 17.75 ± 0.31 g | 4.44 ± 0.01 e |
| CN4 | 390.96 ± 0.34 cd | 25.42 ± 0.22 e | 1.10 ± 0.12 de | 69.83 ± 0.40 f | 23.23 ± 0.02 a | 3.64 ± 0.06 h |
| CN84-1 | 391.37 ± 0.32 cd | 26.30 ± 0.05 d | 1.40 ± 0.05 cd | 68.39 ± 0.09 g | 12.56 ± 0.14 j | 3.91 ± 0.01 g |
| SJ5 | 485.32 ± 0.32 b | 40.40 ± 0.05 a | 21.22 ± 0.11 b | 33.17 ± 0.20 i | 16.59 ± 0.05 i | 5.20 ± 0.05 b |
| CM60 | 489.78 ± 2.41 a | 36.38 ± 0.24 b | 22.45 ± 0.48 a | 35.24 ± 0.71 h | 17.06 ± 0.18 h | 5.62 ± 0.01 a |
| Cultivar | Vitamin B1 (mg) | Vitamin B2 (mg) | Vitamin B3 (mg) |
|---|---|---|---|
| 38 | 0.51 ± 0.02 a | 0.03 ± 0.00 c | 1.31 ± 0.01 c |
| 47 | 0.37 ± 0.02 c | 0.03 ± 0.01 c | 1.42 ± 0.01 b |
| 59 | 0.36 ± 0.01 c | 0.03 ± 0.00 c | 1.34 ± 0.01 bc |
| 112 | 0.50 ± 0.01 a | 0.03 ± 0.01 c | 0.51 ± 0.07 e |
| 107 | 0.14 ± 0.02 e | 0.03 ± 0.00 c | 1.55 ± 0.06 a |
| 108 | 0.04 ± 0.00 g | 0.02 ± 0.00 d | 0.96 ± 0.15 d |
| CN4 | 0.44 ± 0.02 b | 0.11 ± 0.00 a | 1.39 ± 0.04 bc |
| CN84-1 | 0.16 ± 0.02 d | 0.04 ± 0.00 b | 1.35 ± 0.01 bc |
| SJ5 | 0.45 ± 0.02 b | 0.03 ± 0.00 c | 1.43 ± 0.02 b |
| CM60 | 0.07 ± 0.01 f | 0.03 ± 0.00 c | 1.30 ± 0.01 c |
| Cultivar | Macromineral (mg) | Micromineral (mg) | ||||
|---|---|---|---|---|---|---|
| Calcium | Sodium | Potassium | Magnesium | Iron | Zinc | |
| 38 | 86.40 ± 1.15 f | 33.34 ± 8.55 c | 990.13 ± 39.68 f | 115.46 ± 0.13 g | 4.30 ± 0.45 ef | 1.93 ± 0.04 f |
| 47 | 94.87 ± 0.14 e | 28.36 ± 0.59 cd | 1336.65 ± 39.39 c | 138.76 ± 1.58 d | 4.75 ± 0.63 e | 2.05 ± 0.07 e |
| 59 | 100.47 ± 0.51 cd | 19.59 ± 2.14 de | 1375.48 ± 27.10 c | 128.91 ± 1.58 e | 4.12 ± 0.16 f | 2.03 ± 0.00 e |
| 112 | 104.12 ± 1.78 c | 53.48 ± 9.95 b | 1517.36 ± 17.67 a | 118.95 ± 0.23 f | 7.00 ± 0.09 b | 2.38 ± 0.01 d |
| 107 | 31.97 ± 0.24 h | 16.75 ± 1.40 e | 1150.33 ± 12.64 e | 174.83 ± 0.30 b | 5.39 ± 0.28 d | 2.90 ± 0.01 b |
| 108 | 71.53 ± 1.29 g | 73.97 ± 8.54 a | 1449.51 ± 25.10 b | 114.79 ± 0.25 g | 7.56 ± 0.33 a | 2.33 ± 0.02 d |
| CN4 | 117.15 ± 1.68 b | 45.98 ± 4.30 b | 915.34 ± 31.61 g | 170.20 ± 0.58 c | 6.04 ± 0.25 c | 2.95 ± 0.00 b |
| CN84-1 | 96.85 ± 2.48 de | 21.54 ± 0.65 de | 972.51 ± 6.37 fg | 139.78 ± 0.57 d | 3.97 ± 0.13 f | 2.55 ± 0.02 c |
| SJ5 | 241.39 ± 0.74 a | 20.63 ± 1.69 de | 1249.61 ± 5.82 d | 173.74 ± 1.27 b | 6.91 ± 0.26 b | 4.41 ± 0.07 a |
| CM60 | 242.17 ± 8.14 a | 47.61 ± 8.74 b | 1487.96 ± 84.79 ab | 224.81 ± 3.67 a | 6.63 ± 0.27 b | 4.31 ± 0.15 a |
| Cultivar | TPCs (mg GAE/g DW) | Antioxidant Activities | ||
|---|---|---|---|---|
| ORAC Assay (µmol TE/g DW) | FRAP Assay (µmol TE/g DW) | DPPH Radical Scavenging Assay (µmol TE/100 g DW) | ||
| 38 | 1.60 ± 0.07 g | 114.95 ± 6.65 f | 8.03 ± 0.34 d | 0.48 ± 0.03 d |
| 47 | 0.72 ± 0.04 h | 21.00 ± 1.90 i | 2.74 ± 0.11 h | 0.15 ± 0.01 g |
| 59 | 1.87 ± 0.08 f | 100.89 ± 5.25 g | 7.38 ± 0.32 e | 0.55 ± 0.05 c |
| 112 | 1.99 ± 0.16 e | 59.42 ± 5.40 h | 8.53 ± 0.59 c | 0.78 ± 0.07 b |
| 107 | 2.43 ± 0.10 c | 128.46 ± 11.36 e | 14.38 ± 0.76 b | 0.59 ± 0.05 c |
| 108 | 3.12 ± 0.10 a | 139.15 ± 12.21 d | 18.00 ± 0.60 a | 1.06 ± 0.08 a |
| CN4 | 2.09 ± 0.06 d | 177.54 ± 10.72 c | 5.12 ± 0.17 f | 0.54 ± 0.04 c |
| CN84-1 | 1.92 ± 0.10 ef | 249.95 ± 5.53 a | 5.19 ± 0.08 f | 0.48 ± 0.04 d |
| SJ5 | 1.60 ± 0.05 g | 138.27 ± 3.48 d | 4.31 ± 0.19 g | 0.28 ± 0.02 e |
| CM60 | 2.54 ± 0.08 b | 197.83 ± 5.64 b | 4.04 ± 0.22 g | 0.24 ± 0.02 f |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sahasakul, Y.; Aursalung, A.; Thangsiri, S.; Wongchang, P.; Sangkasa-ad, P.; Wongpia, A.; Polpanit, A.; Inthachat, W.; Temviriyanukul, P.; Suttisansanee, U. Nutritional Compositions, Phenolic Contents, and Antioxidant Potentials of Ten Original Lineage Beans in Thailand. Foods 2022, 11, 2062. https://0-doi-org.brum.beds.ac.uk/10.3390/foods11142062
Sahasakul Y, Aursalung A, Thangsiri S, Wongchang P, Sangkasa-ad P, Wongpia A, Polpanit A, Inthachat W, Temviriyanukul P, Suttisansanee U. Nutritional Compositions, Phenolic Contents, and Antioxidant Potentials of Ten Original Lineage Beans in Thailand. Foods. 2022; 11(14):2062. https://0-doi-org.brum.beds.ac.uk/10.3390/foods11142062
Chicago/Turabian StyleSahasakul, Yuraporn, Amornrat Aursalung, Sirinapa Thangsiri, Pitthaya Wongchang, Parichart Sangkasa-ad, Aphinya Wongpia, Auytin Polpanit, Woorawee Inthachat, Piya Temviriyanukul, and Uthaiwan Suttisansanee. 2022. "Nutritional Compositions, Phenolic Contents, and Antioxidant Potentials of Ten Original Lineage Beans in Thailand" Foods 11, no. 14: 2062. https://0-doi-org.brum.beds.ac.uk/10.3390/foods11142062