
Comparison of Production and Fluorescence Characteristics of Phycoerythrin from Three Strains of Porphyridium

 ,
, 


Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Culture Conditions
2.2. Experimental Design
2.3. Morphological Observation
2.4. Construction of Phylogenetic Tree
2.5. Growth Measurement
2.6. Determination of Phycobiliproteins Content
2.7. Purification of Phycoerythrin
2.8. Determination of Fluorescence Characteristics of Phycoerythrin
2.9. RNA-seq
2.9.1. Total RNA Extraction, cDNA Library Construction and Sequencing
2.9.2. Data Analysis
2.10. Statistical Analysis
3. Results
3.1. Molecular Identification and Cell Morphology
3.2. Growth Characteristic
3.3. Phycobiliproteins Content
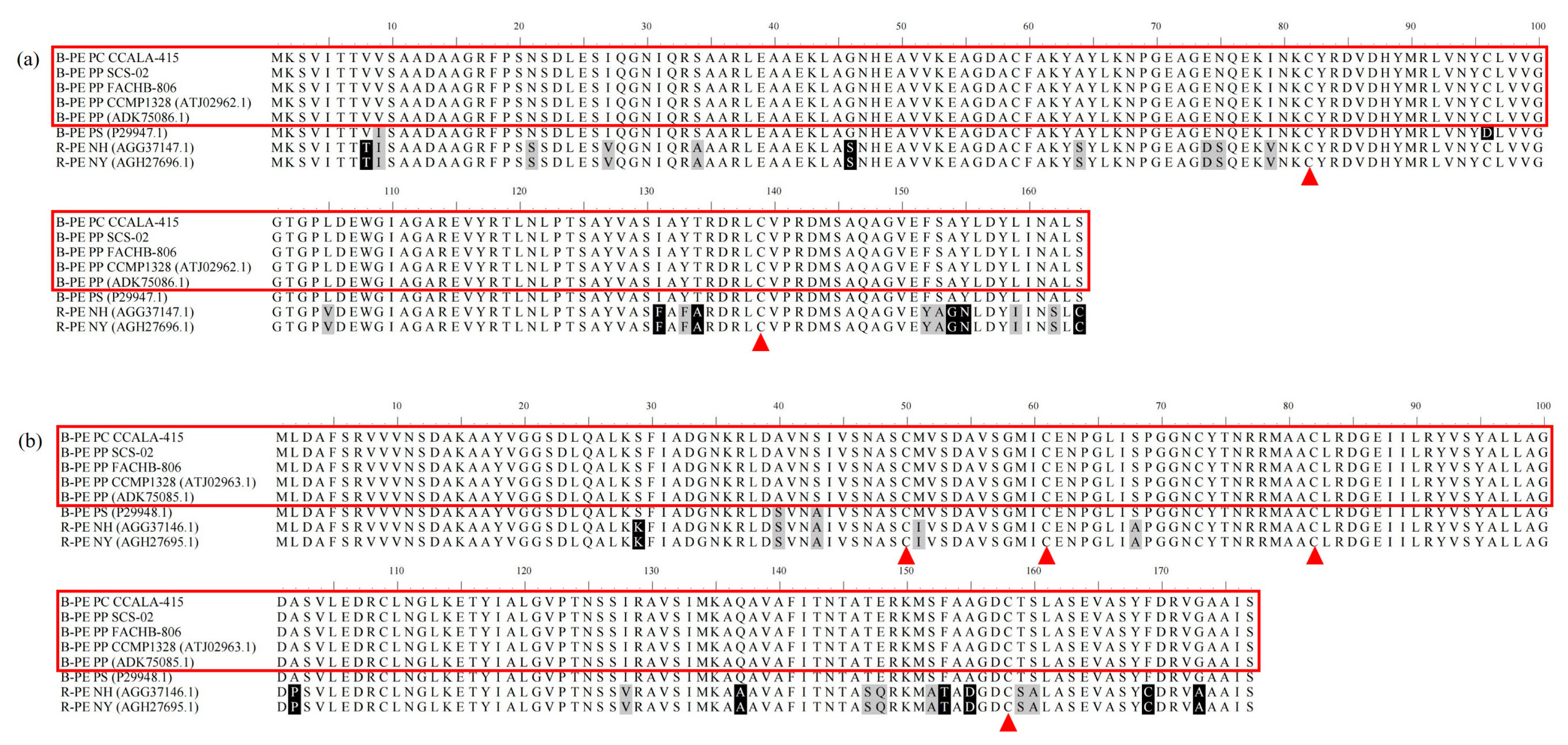
3.4. Amino acid Sequences of Phycoerythrin
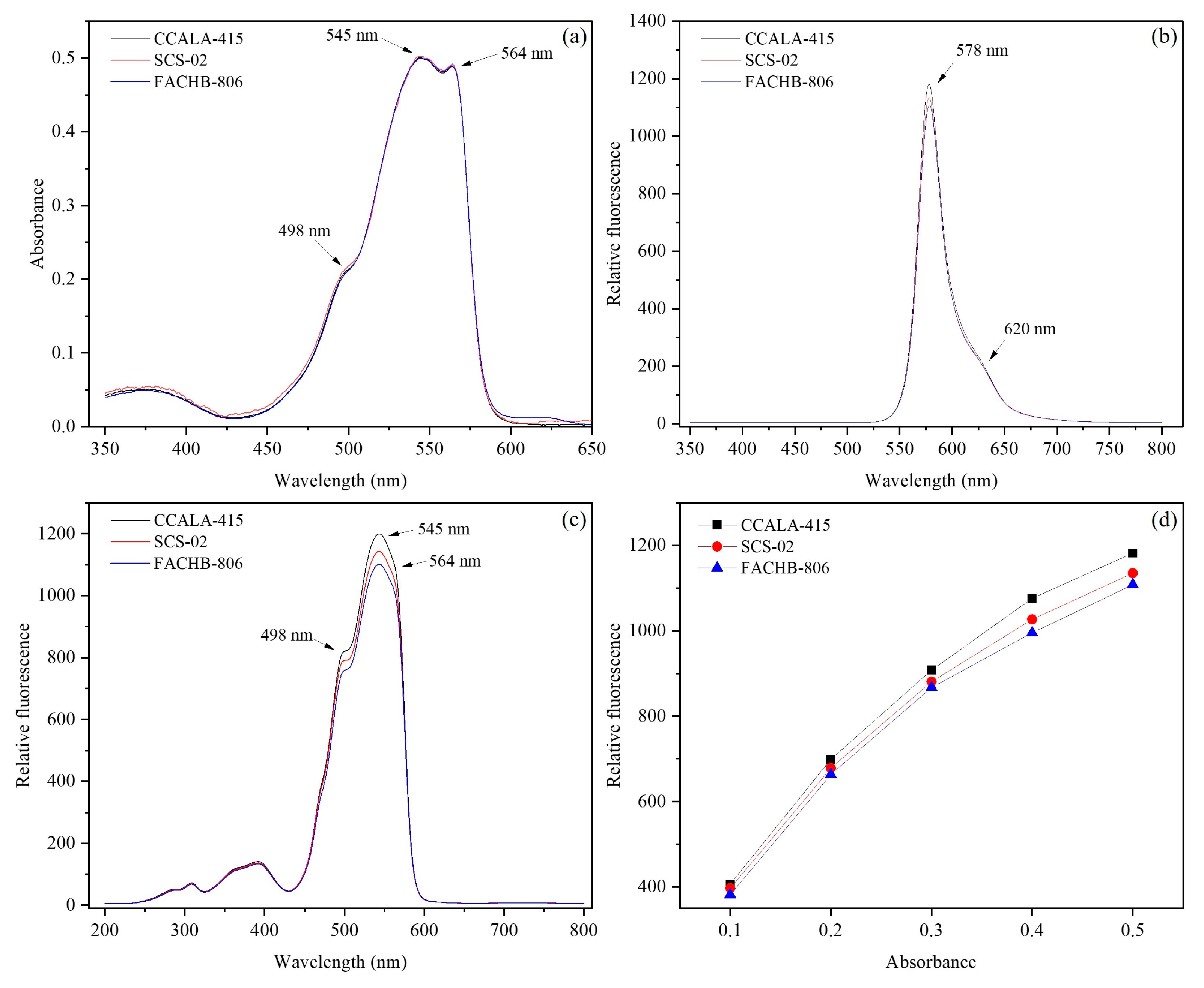
3.5. Fluorescence Characteristics of Phycoerythrin
3.6. Transcriptome Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Qi, H.; Liu, Y.; Qi, X.; Liang, H.; Chen, H.; Jiang, P.; Wang, D. Dietary Recombinant Phycoerythrin Modulates the Gut Microbiota of H22 Tumor-Bearing Mice. Mar. Drugs 2019, 17, 665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ying, J.; Tang, Z.; Zhao, G.; Li, X.; Pan, R.; Lin, S.; Lou, Y.; Wang, W.; Li, P.; Bao, Q.; et al. Transcriptionomic Study on Apoptosis of Skov-3 Cells Induced by Phycoerythrin from Gracilaria lemaneiformis. Anti Cancer Agents Med. Chem. 2021, 21, 1240–1249. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Shen, Z.; Li, L.; Li, Y.; Zhao, H.; Jiang, X. Immunomodulatory Activity of R-Phycoerythrin from Porphyra haitanensis Via Tlr4/Nf-Κb-Dependent Immunocyte Differentiation. Food Funct. 2020, 11, 2173–2185. [Google Scholar] [CrossRef] [PubMed]
- Lee, P.T.; Huang, J.; Huang, C.Y.; Liu, Z.X.; Yeh, H.Y.; Huang, H.T.; Chen, L.L.; Nan, F.H.; Lee, M.C. Phycoerythrin from Colaconema sp. Has Immunostimulatory Effects on the Whiteleg Shrimp Litopenaeus vannamei and Increases Resistance to Vibrio parahaemolyticus and White Spot Syndrome Virus. Animals 2021, 11, 2371. [Google Scholar] [CrossRef]
- Lee, P.T.; Yeh, H.Y.; Lung, W.Q.C.; Huang, J.; Chen, Y.J.; Chen, B.Y.; Nan, F.H.; Lee, M.C. R-Phycoerythrin from Colaconema formosanum (Rhodophyta), an Anti-Allergic and Collagen Promoting Material for Cosmeceuticals. Appl. Sci. 2021, 11, 9425. [Google Scholar] [CrossRef]
- Nath, P.C.; Tiwari, O.N.; Devi, I.; Bandyopadhyay, T.K.; Bhunia, B. Biochemical and Morphological Fingerprints of Isolated Anabaena sp.: A Precious Feedstock for Food Additives. Biomass Convers 2020, 11, 2723–2733. [Google Scholar] [CrossRef]
- Khan, Z.; Maznah, W.O.W.; Merican, M.S.M.F.; Convey, P.; Najimudin, N.; Alias, S.A. A Comparative Study of Phycobilliprotein Production in Two Strains of Pseudanabaena Isolated from Arctic and Tropical Regions in Relation to Different Light Wavelengths and Photoperiods. Polar Sci. 2019, 20, 3–8. [Google Scholar] [CrossRef]
- Mishra, S.K.; Shrivastav, A.; Maurya, R.R.; Patidar, S.K.; Haldar, S.; Mishra, S. Effect of Light Quality on the C-Phycoerythrin Production in Marine Cyanobacteria Pseudanabaena sp. Isolated from Gujarat Coast, India. Protein Expr. Purif. 2012, 81, 5–10. [Google Scholar] [CrossRef]
- Fethi, M.; Ben Ghedifa, A. Optimum Ranges of Combined Abiotic Factor for Gracilaria gracilis Aquaculture. J. Appl. Phycol. 2019, 31, 3025–3040. [Google Scholar] [CrossRef]
- Mensi, F.; Nasraoui, S.; Bouguerra, S.; Ben Ghedifa, A.; Chalghaf, M. Effect of Lagoon and Sea Water Depth on Gracilaria gracilis Growth and Biochemical Composition in the Northeast of Tunisia. Sci. Rep. 2020, 10, 10014. [Google Scholar] [CrossRef]
- Ghedifa, A.B.; Vega, J.; Korbee, N.; Mensi, F.; Figueroa, F.; Sadok, S. Effects of Light Quality on the Photosynthetic Activity and Biochemical Composition of Gracilaria gracilis (Rhodophyta). J. Appl. Phycol. 2021, 33, 3413–3425. [Google Scholar] [CrossRef]
- Liu, C.; Zou, D.; Li, Y. Nutrient Availability Modulates the Effects of Climate Change on Growth and Photosynthesis of Marine Macroalga Pyropia haitanensis (Bangiales, Rhodophyta). J. Appl. Phycol. 2020, 32, 3285–3294. [Google Scholar] [CrossRef]
- Zhang, T.; Shen, Z.G.; Xu, P.; Zhu, J.Y.; Lu, Q.Q.; Shen, Y.; Wang, Y.; Yao, C.Y.; Li, J.F.; Wang, Y.X.; et al. Analysis of Photosynthetic Pigments and Chlorophyll Fluorescence Characteristics of Different Strains of Porphyra yezoensis. J. Appl. Phycol. 2012, 24, 881–886. [Google Scholar] [CrossRef]
- Aouir, A.; Amiali, M.; Bitam, A.; Benchabane, A.; Raghavan, V.G. Comparison of the Biochemical Composition of Different Arthrospira platensis Strains from Algeria, Chad and the USA. J. Food Meas. Charact. 2017, 11, 913–923. [Google Scholar] [CrossRef]
- Xu, J.; Hu, H. Screening High Oleaginous Chlorella Strains from Different Climate Zones. Bioresour. Technol. 2013, 144, 637–643. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Sun, H.; Ren, Y.; Wu, T.; He, Y.; Chen, F. Chlorella zofingiensis as a Promising Strain in Wastewater Treatment. Bioresour. Technol. 2018, 268, 286–291. [Google Scholar] [CrossRef]
- Rodolfi, L.; Chini Zittelli, G.; Bassi, N.; Padovani, G.; Biondi, N.; Bonini, G.; Tredici, M.R. Microalgae for Oil: Strain Selection, Induction of Lipid Synthesis and Outdoor Mass Cultivation in a Low-Cost Photobioreactor. Biotechnol. Bioeng. 2009, 102, 100–112. [Google Scholar] [CrossRef]
- Li, T.; Xu, J.; Wu, H.; Jiang, P.; Chen, Z.; Xiang, W. Growth and Biochemical Composition of Porphyridium purpureum SCS-02 under Different Nitrogen Concentrations. Mar. Drugs 2019, 17, 124. [Google Scholar] [CrossRef] [Green Version]
- Gudvilovich, I.N.; Lelekov, A.S.; Maltsev, E.I.; Kulikovskii, M.S.; Borovkov, A.B. Growth of Porphyridium purpureum (Porphyridiales, Rhodophyta) and Production of B-Phycoerythrin under Varying Illumination. Russ. J. Plant Physiol. 2021, 68, 188–196. [Google Scholar] [CrossRef]
- Li, S.; Ji, L.; Chen, C.; Zhao, S.; Sun, M.; Gao, Z.; Wu, H.; Fan, J. Efficient Accumulation of High-Value Bioactive Substances by Carbon to Nitrogen Ratio Regulation in Marine Microalgae Porphyridium purpureum. Bioresour. Technol. 2020, 309, 123362. [Google Scholar] [CrossRef]
- Risjani, Y.; Mutmainnah, N.; Manurung, P.; Wulan, S.N.; Yunianta. Exopolysaccharide from Porphyridium cruentum (purpureum) Is Not Toxic and Stimulates Immune Response against Vibriosis: The Assessment Using Zebrafish and White Shrimp Litopenaeus vannamei. Mar. Drugs 2021, 19, 133. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharya, D.; Price, D.C.; Chan, C.X.; Qiu, H.; Rose, N.; Ball, S.; Weber, A.P.; Arias, M.C.; Henrissat, B.; Coutinho, P.M.; et al. Genome of the Red Alga Porphyridium purpureum. Nat. Commun. 2013, 4, 1941. [Google Scholar] [CrossRef] [Green Version]
- Csogor, Z.; Kiessling, B.; Perner, I.; Fleck, P.; Posten, C. Growth and Product Formation of Porphyridium purpureum. J. Appl. Phycol. 2001, 13, 317–324. [Google Scholar] [CrossRef]
- Ott, F.D. A Brief Review of the Species of Porphyridium cruentum with Additional Records for the Rarely Collected Alga Porphyridium sordidum Geitler, 1932 (Rhodophycophyta, Porphyridiales). Arch. Fur Protistenkd. 1987, 134, 35–41. [Google Scholar] [CrossRef]
- Huang, Z.; Guo, S.; Guo, Z.; He, Y.; Chen, B. Integrated Green One-Step Strategy for Concurrent Recovery of Phycobiliproteins and Polyunsaturated Fatty Acids from Wet Porphyridium Biomass. Food Chem. 2022, 389, 133103. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.J.; Cheung, P.C.K. Cold Stress Treatment Enhances Production of Metabolites and Biodiesel Feedstock in Porphyridium cruentum Via Adjustment of Cell Membrane Fluidity. Sci. Total Environ. 2021, 780, 146612. [Google Scholar] [CrossRef] [PubMed]
- Lang, I.; Hodac, L.; Friedl, T.; Feussner, I. Fatty Acid Profiles and Their Distribution Patterns in Microalgae: A Comprehensive Analysis of More Than 2000 Strains from the Sag Culture Collection. BMC Plant Biol. 2011, 11, 124. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. Mega X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- De Marsac, N.; Tandeau; Houmard, J. Complementary Chromatic Adaptation: Physiological Conditions and Action Spectra. In Cyanobacteria; Elsevier: Amsterdam, The Netherlands, 1988; pp. 318–328. [Google Scholar]
- Rito-Palomares, M.; Nuñez, L.; Amador, D. Practical Application of Aqueous Two-Phase Systems for the Development of a Prototype Process for C-Phycocyanin Recovery from Spirulina maxima. J. Chem. Technol. Biotechnol. 2001, 76, 1273–1280. [Google Scholar] [CrossRef]
- Patil, G.; Chethana, S.; Sridevi, A.S.; Raghavarao, K.S. Method to Obtain C-Phycocyanin of High Purity. J. Chromatogr. A 2006, 1127, 76–81. [Google Scholar] [CrossRef]
- Camara-Artigas, A.; Bacarizo, J.; Andujar-Sanchez, M.; Ortiz-Salmeron, E.; Mesa-Valle, C.; Cuadri, C.; Martin-Garcia, J.M.; Martinez-Rodriguez, S.; Mazzuca-Sobczuk, T.; Ibanez, M.J.; et al. Ph-Dependent Structural Conformations of B-Phycoerythrin from Porphyridium cruentum. FEBS J. 2012, 279, 3680–3691. [Google Scholar] [CrossRef] [PubMed]
- Seghiri, R.; Legrand, J.; Hsissou, R.; Essamri, A. Comparative Study of the Impact of Conventional and Unconventional Drying Processes on Phycobiliproteins from Arthrospira platensis. Algal Res. 2021, 53, 102165. [Google Scholar] [CrossRef]
- Sosa-Hernandez, J.E.; Rodas-Zuluaga, L.I.; Castillo-Zacarias, C.; Rostro-Alanis, M.; De La Cruz, R.; Carrillo-Nieves, D.; Salinas-Salazar, C.; Fuentes Grunewald, C.; Llewellyn, C.A.; Olguin, E.J.; et al. Light Intensity and Nitrogen Concentration Impact on the Biomass and Phycoerythrin Production by Porphyridium purpureum. Mar. Drugs. 2019, 17, 460. [Google Scholar] [CrossRef] [Green Version]
- Su, G.; Jiao, K.; Chang, J.; Li, Z.; Guo, X.; Sun, Y.; Zeng, X.; Lu, Y.; Lin, L. Enhancing Total Fatty Acids and Arachidonic Acid Production by the Red Microalgae Porphyridium purpureum. Bioresour. Bioprocess. 2016, 3, 33. [Google Scholar] [CrossRef] [Green Version]
- Medina-Cabrera, E.V.; Ruhmann, B.; Schmid, J.; Sieber, V. Characterization and Comparison of Porphyridium sordidum and Porphyridium purpureum Concerning Growth Characteristics and Polysaccharide Production. Algal Res. 2020, 49, 9. [Google Scholar] [CrossRef]
- Ritz, M.; Lichtle, C.; Spilar, A.; Joder, A.; Thomas, J.C.; Etienne, A.L. Characterization of Phycocyanin-Deficient Phycobilisomes from a Pigment Mutant of Porphyridium sp. (Rhodophyta). J. Phycol. 1998, 34, 835–843. [Google Scholar] [CrossRef]
- Xie, M.; Li, W.; Lin, H.; Wang, X.; Dong, J.; Qin, S.; Zhao, F. Difference in Light Use Strategy in Red Alga between Griffithsia pacifica and Porphyridium purpureum. Sci. Rep. 2021, 11, 14367. [Google Scholar] [CrossRef]
- Lee, M.C.; Yeh, H.Y.; Jhang, F.J.; Lee, P.T.; Lin, Y.K.; Nan, F.H. Enhancing Growth, Phycoerythrin Production, and Pigment Composition in the Red Alga Colaconema sp. Through Optimal Environmental Conditions in an Indoor System. Bioresour. Technol. 2021, 333, 125199. [Google Scholar] [CrossRef]
- Hsieh-Lo, M.; Castillo, G.; Ochoa-Becerra, M.A.; Mojica, L. Phycocyanin and Phycoerythrin: Strategies to Improve Production Yield and Chemical Stability. Algal Res. 2019, 42, 101600. [Google Scholar] [CrossRef]
- Kim, J.K.; Kraemer, G.P.; Neefus, C.D.; Chung, I.K.; Yarish, C. Effects of Temperature and Ammonium on Growth, Pigment Production and Nitrogen Uptake by Four Species of Porphyra (Bangiales, Rhodophyta) Native to the New England Coast. J. Appl. Phycol. 2007, 19, 431–440. [Google Scholar] [CrossRef]
- Varela-Álvarez, E.; Tobin, P.; Guihéneuf, F.; Fitzgerald, R.; Stengel, D.B. Phycobiliproteins, Nitrogenous Compounds and Fatty Acid Contents in Field-Collected and Cultured Gametophytes of Porphyra dioica, a Red Sea Vegetable. J. Appl. Phycol. 2019, 31, 3849–3860. [Google Scholar] [CrossRef]
- Yanshin, N.; Kushnareva, A.; Lemesheva, V.; Birkemeyer, C.; Tarakhovskaya, E. Chemical Composition and Potential Practical Application of 15 Red Algal Species from the White Sea Coast (the Arctic Ocean). Molecules 2021, 26, 2489. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Jiao, K.; Zhong, H.; Wu, S.; Ho, S.H.; Zeng, X.; Li, J.; Tang, X.; Sun, Y.; Lin, L. Induced Cultivation Pattern Enhanced the Phycoerythrin Production in Red Alga Porphyridium purpureum. Bioprocess Biosyst. Eng. 2020, 43, 347–355. [Google Scholar] [CrossRef] [PubMed]
- Friedman, C.; Dubinsky, Z.; Arad, S.M. Effect of Light-Intensity on Growth and Polysaccharide Production in Red and Blue-Green Rhodophyta Unicells. Bioresour. Technol. 1991, 38, 105–110. [Google Scholar] [CrossRef]
- Ficner, R.; Lobeck, K.; Schmidt, G.; Huber, R. Isolation, Crystallization, Crystal Structure Analysis and Refinement of B-Phycoerythrin from the Red Alga Porphyridium sordidum at 2·2 Å Resolution. J. Mol. Biol. 1992, 228, 935–950. [Google Scholar] [CrossRef]
- Gantt, E.; Edwards, M.R.; Conti, S.F. Ultrastructure of Porphyridium aerugineum a Blue-Green Colored Rhodophytan. J. Phycol. 1968, 4, 65–71. [Google Scholar] [CrossRef]
- Glazer, A.N.; Hixson, C.S. Subunit Structure and Chromophore Composition of Rhodophytan Phycoerythrins. Porphyridium cruentum B-Phycoerythrin and B-Phycoerythrin. J. Biol. Chem. 1977, 252, 32–42. [Google Scholar]
- Gantt, E.; Lipschultz, C.A. Phycobilisomes of Porphyridium cruentum: Pigment Analysis. Biochemistry 1974, 13, 2960–2966. [Google Scholar] [CrossRef]
- Ficner, R.; Huber, R. Refined Crystal Structure of Phycoerythrin from Porphyridium cruentum at 0.23-nm Resolution and Localization of the Gamma Subunit. Eur. J. Biochem. 1993, 218, 103–106. [Google Scholar] [CrossRef]
- Leney, A.C.; Tschanz, A.; Heck, A.J.R. Connecting Color with Assembly in the Fluorescent B-Phycoerythrin Protein Complex. FEBS J. 2018, 285, 178–187. [Google Scholar] [CrossRef] [Green Version]
- Dailey, H.A.; Dailey, T.A.; Gerdes, S.; Jahn, D.; Jahn, M.; O’brian, M.R.; Warren, M.J. Prokaryotic Heme Biosynthesis: Multiple Pathways to a Common Essential Product. Microbiol. Mol. Biol. Rev. 2017, 81, e00048-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, X.X.; Wang, H.T.; Zang, X.N.; Liu, Z.; Xu, D.; Jin, Y.M.; Zhang, F.; Wang, Z.D. Changes in the Photosynthetic Pigment Contents and Transcription Levels of Phycoerythrin-Related Genes in Three Gracilariopsis lemaneiformis Strains under Different Light Intensities. J. Ocean Univ. China 2021, 20, 661–668. [Google Scholar] [CrossRef]
- Huang, X.; Zang, X.; Wu, F.; Jin, Y.; Wang, H.; Liu, C.; Ding, Y.; He, B.; Xiao, D.; Song, X.; et al. Transcriptome Sequencing of Gracilariopsis lemaneiformis to Analyze the Genes Related to Optically Active Phycoerythrin Synthesis. PLoS ONE 2017, 12, e0170855. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Primer | Primer Sequence |
|---|---|---|
| 18s rDNA | 18s-F | 5′-CTGGTTGATCCTGCCAGT-3′ |
| 18s-R | 5′-CACCTACGCAAACCTTGTTACGACTT-3′ | |
| ITS | ITS-F | 5′-GGAAGGAGAAGTCGTAACAAGG-3′ |
| ITS-R | 5′-TCCTCCGCTTATTGATATGC-3′ | |
| α subunit of phycoerythrin | AS | 5′-ATGAAATCAGTTATTACTACTGTTGTAAGTGCAGCTG-3′ |
| AAS | 5′-CTATGAAAGTGCGTTAATTAAGTAG-3 | |
| β subunit of phycoerythrin | BS | 5′-ATGCTTGACGCATTTTCTAGAGT-3′ |
| BAS | 5′-TTAGCTAATTGCTGCACCAA-3′ |
| Strain | Fitted Curves | R2 |
|---|---|---|
| CCALA-415 | y = 486.33ln(x) + 1508.6 | 0.9960 |
| SCS-02 | y = 461.16ln(x) + 1444.0 | 0.9972 |
| FACHB-806 | y = 452.21ln(x) + 1411.4 | 0.9980 |
| Samples | Total Raw Reads | Total Clean Reads | Total Clean Nucleotides (nt) | Q20 (%) | N (%) | Map to Reference Genome (%) |
|---|---|---|---|---|---|---|
| CCALA-415 | 47,261,608 | 42,274,636 | 6341,195,400 | 97.23 | 0.001258 | 92.86% |
| SCS-02 | 53,769,454 | 48,996,844 | 7349,526,600 | 97.42 | 0.001235 | 95.06% |
| FACHB-806 | 47,496,606 | 42,821,236 | 6423,185,400 | 97.51 | 0.001244 | 90.65% |
| KEGG Entry | Protein | Abbreviation | SCS-02 vs. CCALA-415 | FACHB-806 vs. CCALA-415 | |
|---|---|---|---|---|---|
| Phycoerythrobilin metabolism | K01885 | Glutamyl-tRNA synthetase | - | Up | Up |
| K02492 | Glutamyl-tRNA reductase | - | Down | Down | |
| K01845 | Glutamate-1-semialdehyde-2,1-aminomutase | - | Down | Down | |
| K01698 | Porphobilinogen synthase | - | Down | Down | |
| K01749 | Porphobilinogen deaminase, chloroplastic | - | Up | Up | |
| K01719 | Uroporphyrinogen-III synthase | - | Down | Up | |
| K01599 | Uroporphyrinogen decarboxylase | - | Down | Down | |
| K00228 | Oxygen-dependent coproporphyrinogen-III oxidase | - | Down | Down | |
| K00231 | Protoporphyrinogen oxidase | - | Up | Up | |
| K01772 | Protoporphyrin/coproporphyrin ferrochelatase | - | Down | Down | |
| K00510 | Heme oxygenase 1 | - | Down | Up | |
| K05369 | 15,16-dihydrobiliverdin: ferredoxin oxidoreductase | - | Down | Down | |
| K05370 | Phycoerythrobilin: ferredoxin oxidoreductase | - | Down | Down | |
| Phycobilisome components | K05376 | Phycoerythrin alpha subunit | PE-α | Down | Down |
| K05377 | Phycoerythrin beta subunit | PE-β | Down | Up | |
| K05378 | Phycoerythrin gamma chain | PE-γ | Down | Down | |
| K02092 | Allophycocyanin alpha subunit | APC-α | Down | Down | |
| K02093 | Allophycocyanin beta subunit | APC-β | Up | Up | |
| K02094 | Phycobilisome 7.8 kDa linker polypeptide, allophycocyanin-associated, core | apcC | Down | Down | |
| K02095 | Phycobilisome linker polypeptide | apcD | Down | Down | |
| K02096 | Phycobilisome linker polypeptide | apcE | Down | Down | |
| K02286 | Phycobilisome 32.1 kDa linker polypeptide, phycocyanin-associated, rod | cpcC | Down | Down | |
| K05378 | Phycobilisome 31.8 kDa linker polypeptide, phycoerythrin-associated, rod | cpeC | Down | Down | |
| K05379 | Phycobilisome 27.9 kDa linker polypeptide, phycoerythrin-associated, rod | cpeD | Down | Down |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, C.; Wu, H.; Xiang, W.; Wu, H.; Wang, N.; Wu, J.; Li, T. Comparison of Production and Fluorescence Characteristics of Phycoerythrin from Three Strains of Porphyridium. Foods 2022, 11, 2069. https://0-doi-org.brum.beds.ac.uk/10.3390/foods11142069
Li C, Wu H, Xiang W, Wu H, Wang N, Wu J, Li T. Comparison of Production and Fluorescence Characteristics of Phycoerythrin from Three Strains of Porphyridium. Foods. 2022; 11(14):2069. https://0-doi-org.brum.beds.ac.uk/10.3390/foods11142069
Chicago/Turabian StyleLi, Chulin, Houbo Wu, Wenzhou Xiang, Hualian Wu, Na Wang, Jiayi Wu, and Tao Li. 2022. "Comparison of Production and Fluorescence Characteristics of Phycoerythrin from Three Strains of Porphyridium" Foods 11, no. 14: 2069. https://0-doi-org.brum.beds.ac.uk/10.3390/foods11142069