

Metabolomics Characterize the Differential Metabolic Markers between Bama Xiang Pig and Debao Pig to Identify Pork

Abstract
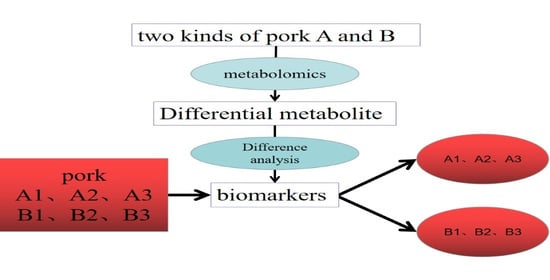
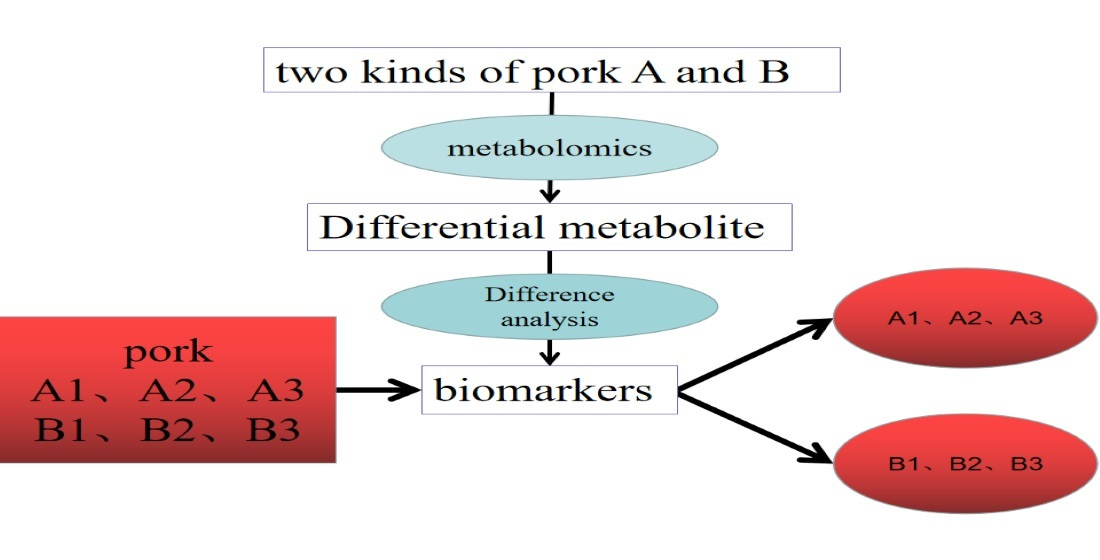
:
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Detection of Metabolites by LC-MS
- We accurately weighed 30 mg of longissimus dorsi into a 1.5 mL EP tube and added an internal standard (l-2-chlorophenyl alanine, 0.06 mg/mL; methanol configuration) 20 μL. Joined 400 μL methanol–water (v:v = 4:1);
- We added two small steel balls, pre-cooling them in the refrigerator at −20 °C for 2 min, and then putting them into the grinder for grinding (60 Hz, 2 min);
- The next step was ultrasonic extraction in an ice water bath for 10 min, stood at −20 °C for 30 min;
- The sample was centrifuged for 10 min (13,000 rpm, 4 °C), 300 μL supernatant was put into the LC-MS injection vial and evaporated;
- Then 300 μL methanol–water (v:v = 1:4) was used for re-dissolution (vortexed for 30 s, ultrasound for 3 min); stood for 2 h at −20 °C;
- The sample was centrifuged for 10 min (13,000 rpm, 4 °C), and 150 μL of supernatant was removed with a syringe, then using a 0.22 μm organic phase pinhole filter, the supernatant was transferred to an LC injection vial and stored at −80 °C until LC-MS analysis.
- Quality control samples (QC) were prepared by mixing an extract of all the samples in equal volumes.
2.3. Data Analysis Methods
3. Results
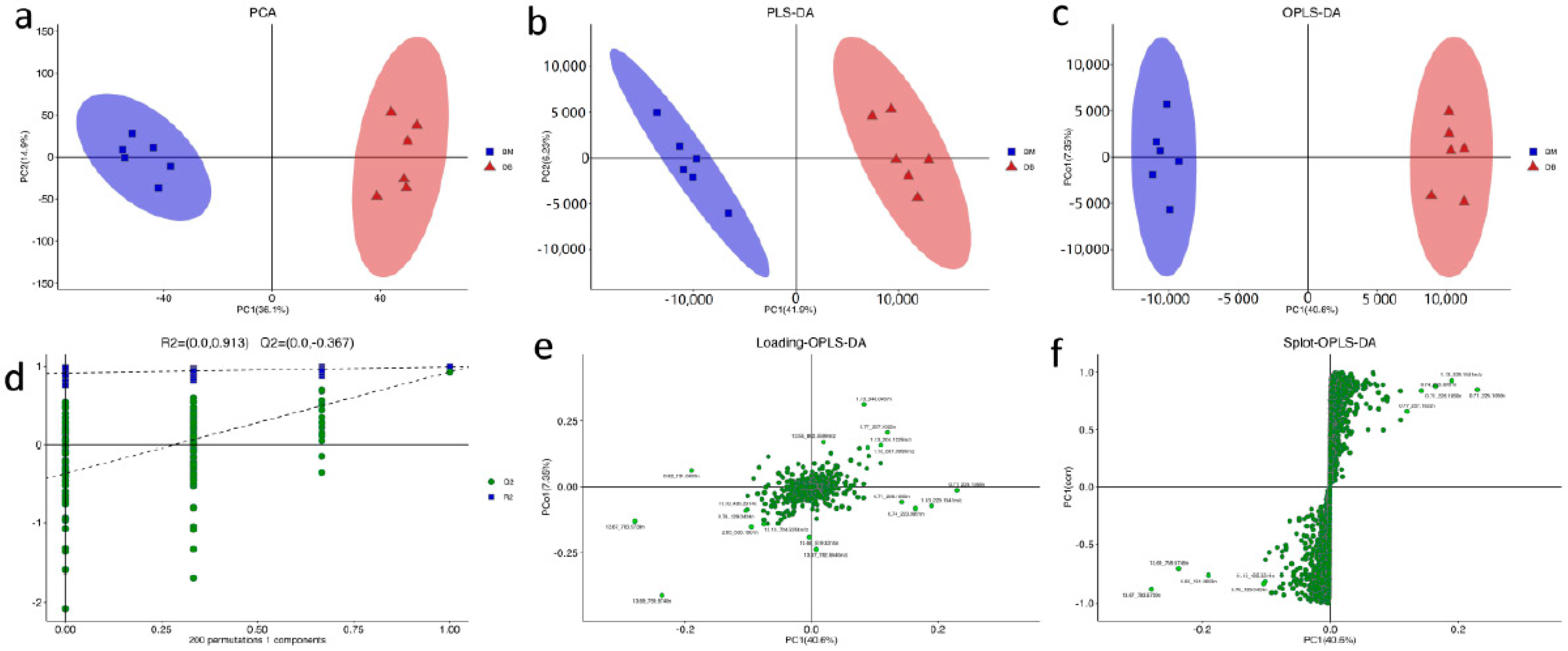
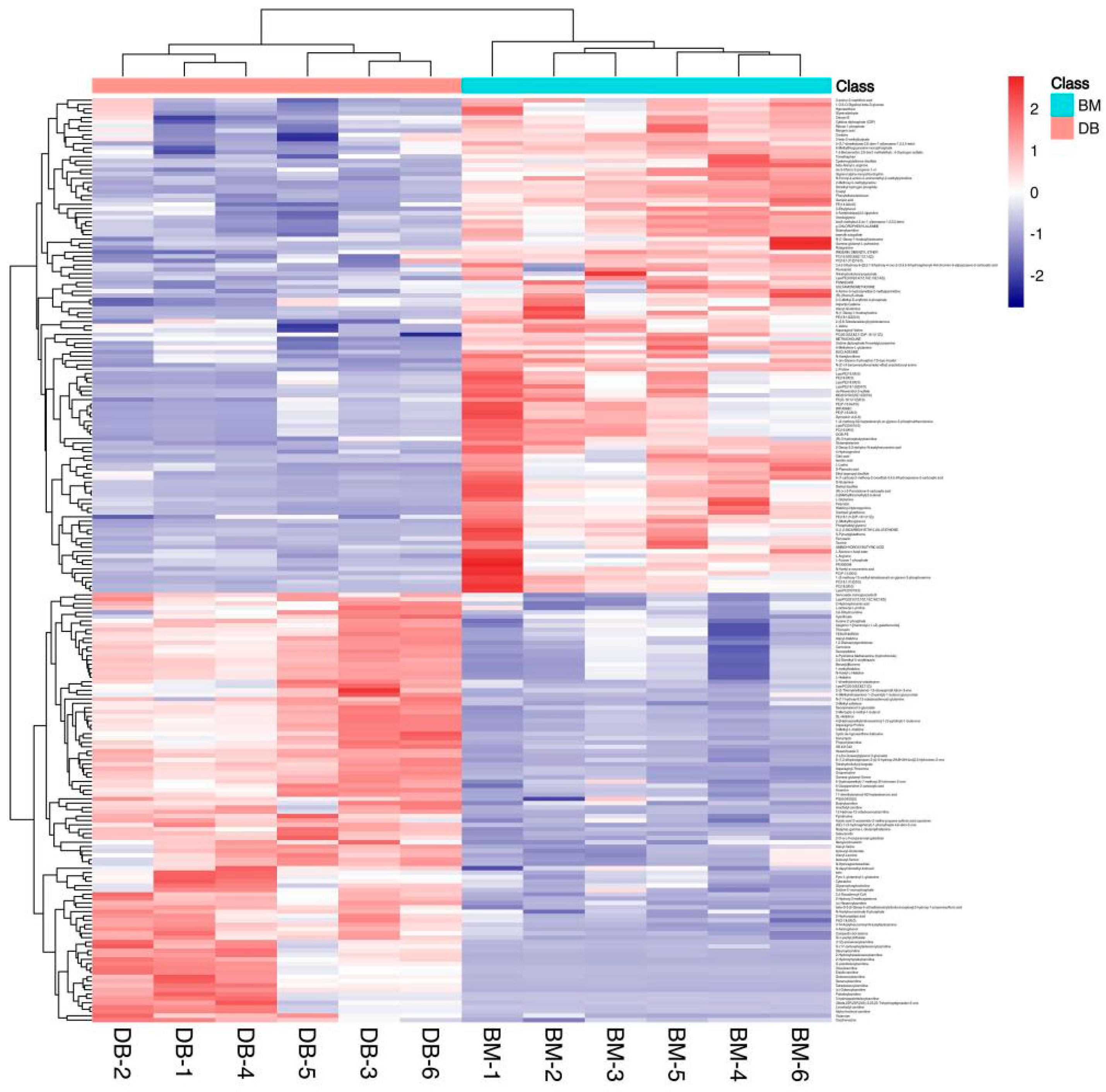
3.1. Multivariate Statistical Analysis and Univariate Statistical Analysis
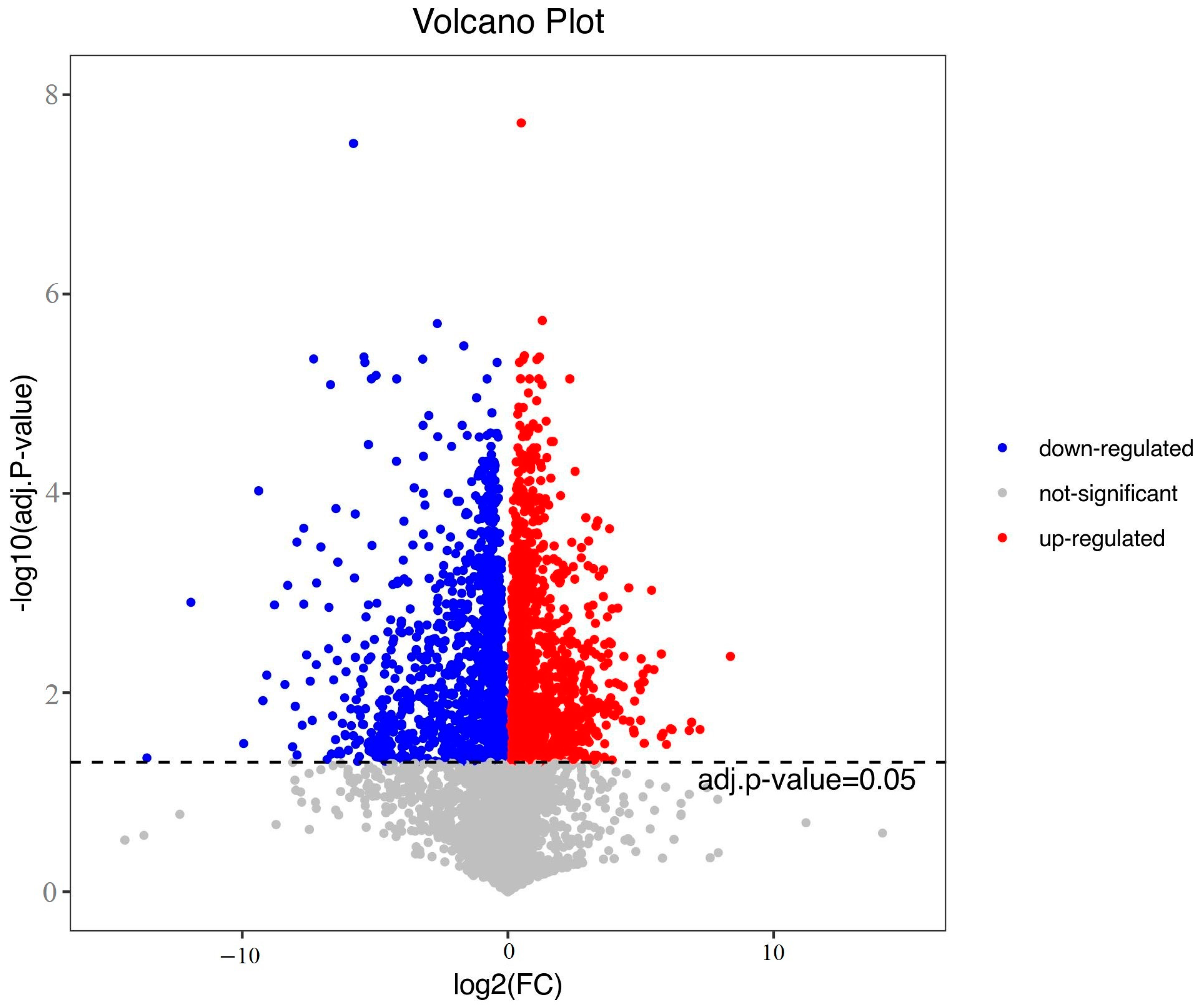
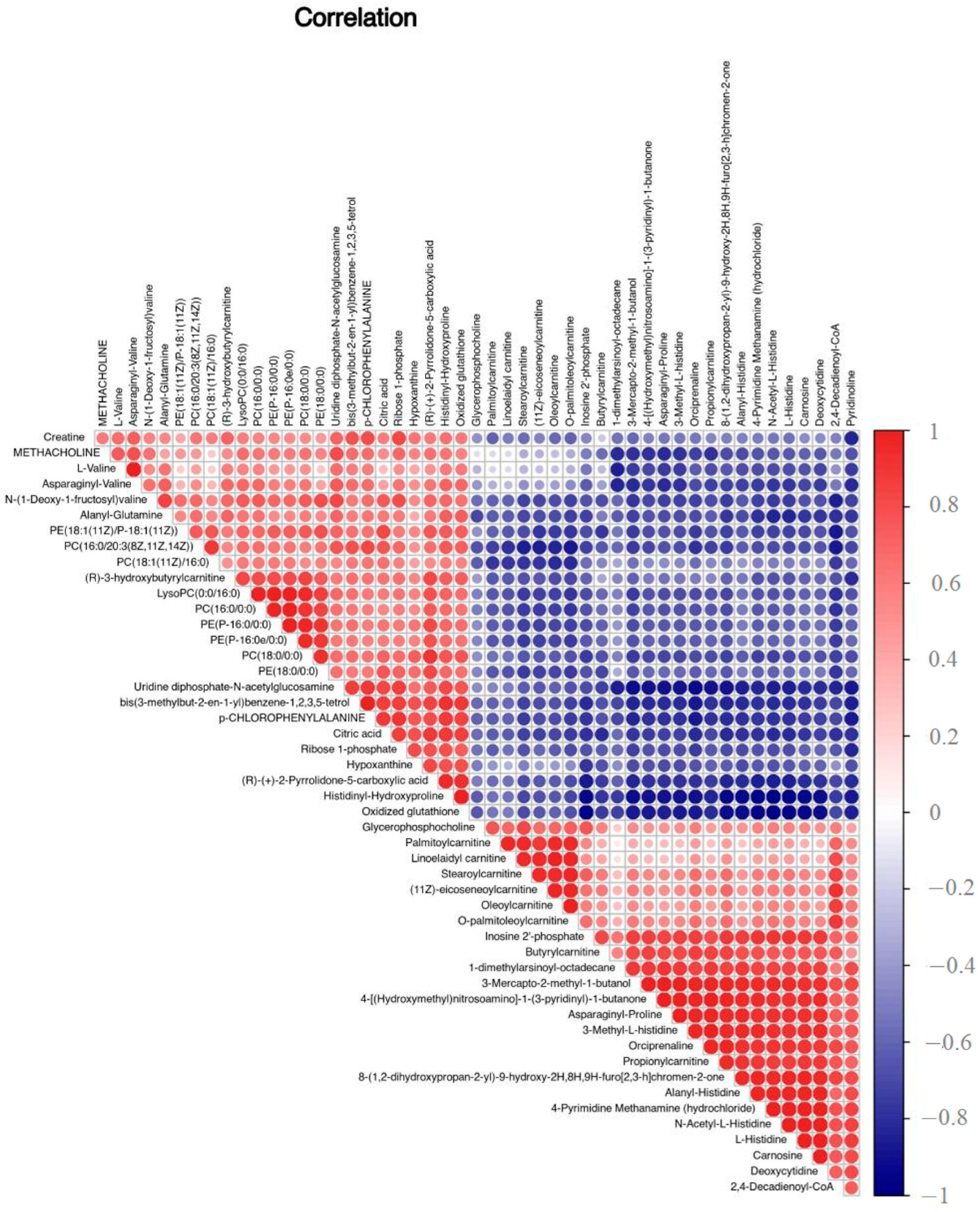
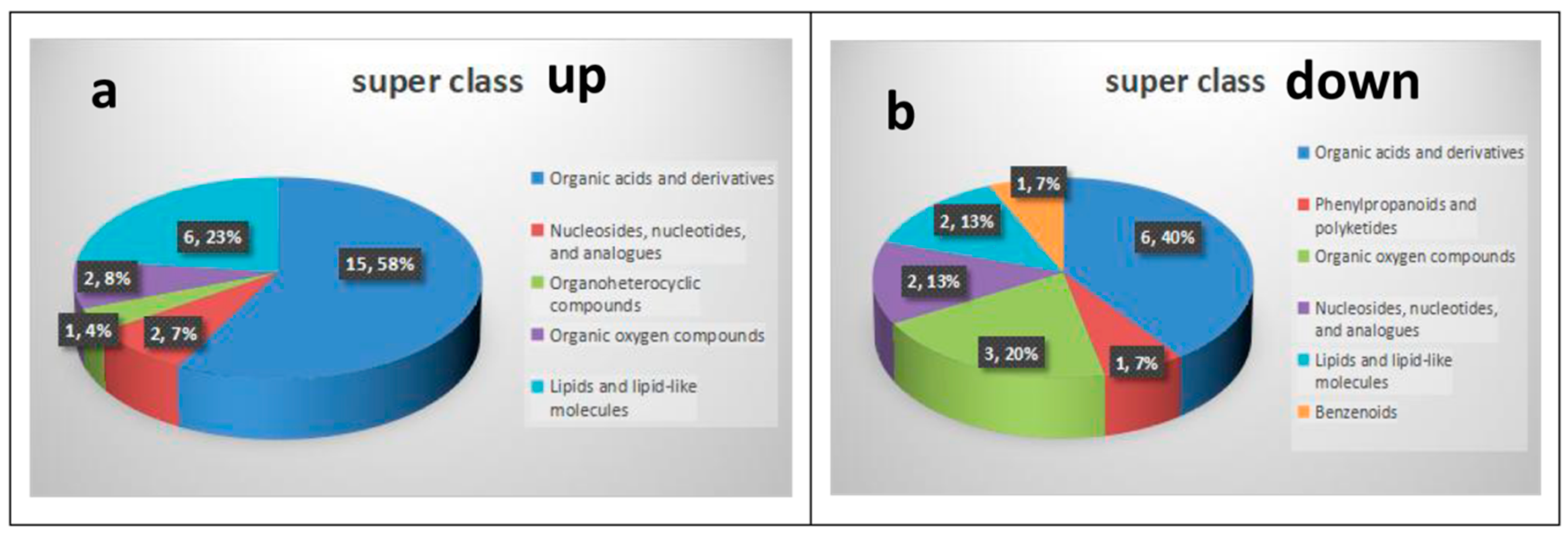
3.2. Differential Metabolites and Correlation Analysis
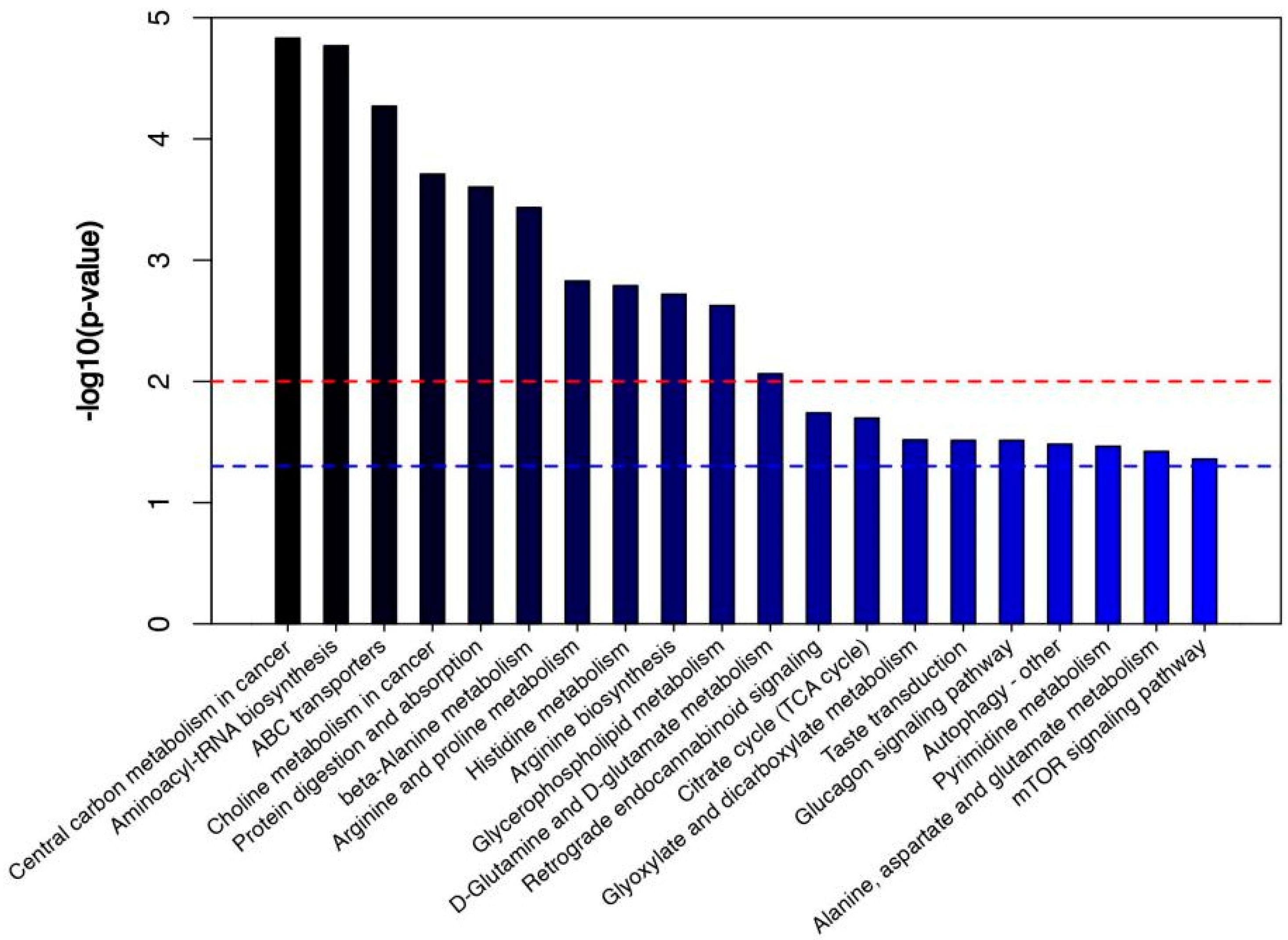
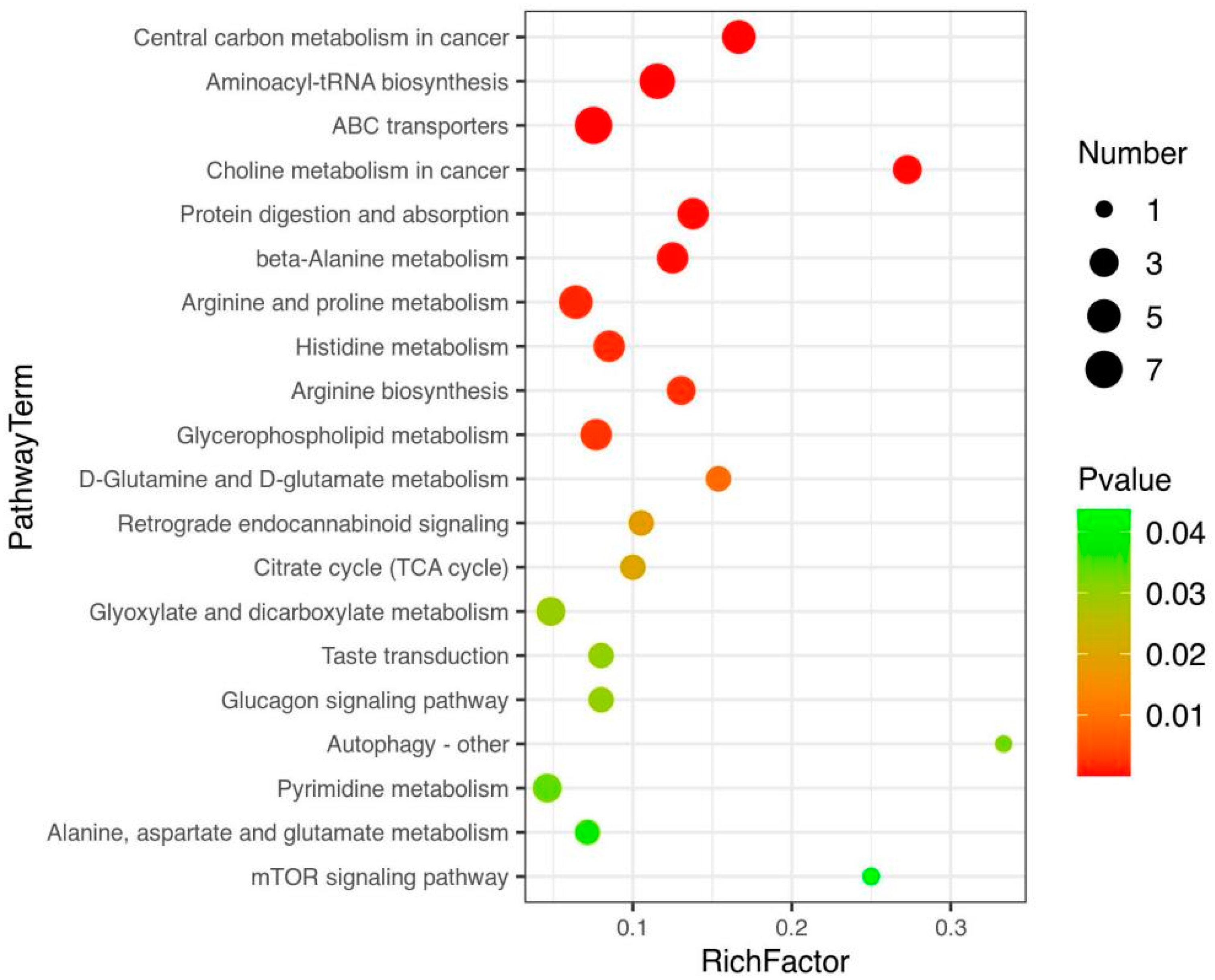
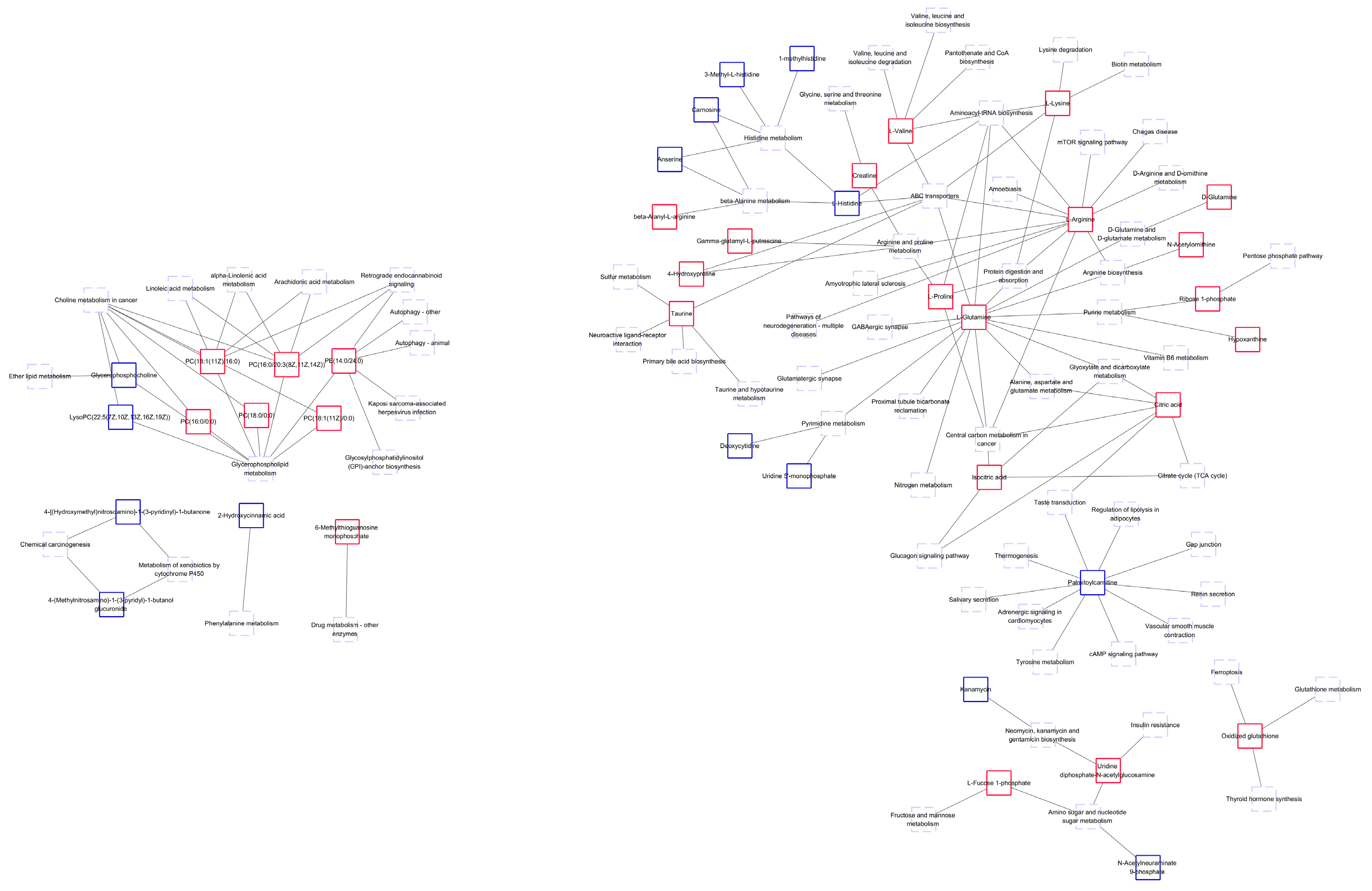
3.3. Path Analysis and Network Diagram
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Driessen, B.; Freson, L.; Buyse, J. Fasting Finisher Pigs before Slaughter Influences Pork Safety, Pork Quality and Animal Welfare. Animals 2020, 10, 2206. [Google Scholar] [CrossRef] [PubMed]
- Acevedo-Giraldo, J.D.; Sánchez, J.A.; Romero, M.H. Effects of feed withdrawal times prior to slaughter on some animal welfare indicators and meat quality traits in commercial pigs. Meat Sci. 2020, 167, 107993. [Google Scholar] [CrossRef] [PubMed]
- Faucitano, L. Preslaughter handling practices and their effects on animal welfare and pork quality. J. Anim. Sci. 2018, 96, 728–738. [Google Scholar] [CrossRef] [PubMed]
- Hambrecht, E.; Eissen, J.J.; Newman, D.J.; Smits, C.H.; den Hartog, L.A.; Verstegen, M.W. Negative effects of stress immediately before slaughter on pork quality are aggravated by suboptimal transport and lairage conditions. J. Anim. Sci. 2005, 83, 440–448. [Google Scholar] [CrossRef] [PubMed]
- Hambrecht, E.; Eissen, J.J.; Newman, D.J.; Smits, C.H.; Verstegen, M.W.; den Hartog, L.A. Preslaughter handling effects on pork quality and glycolytic potential in two muscles differing in fiber type composition. J. Anim. Sci. 2005, 83, 900–907. [Google Scholar] [CrossRef] [Green Version]
- Sardi, L.; Gastaldo, A.; Borciani, M.; Bertolini, A.; Musi, V.; Martelli, G.; Cavallini, D.; Rubini, G.; Nannoni, E. Identification of Possible Pre-Slaughter Indicators to Predict Stress and Meat Quality: A Study on Heavy Pigs. Animals 2020, 10, 945. [Google Scholar] [CrossRef]
- Klont, R.E.; Hulsegge, B.; Hoving-Bolink, A.H.; Gerritzen, M.A.; Kurt, E.; Winkelman-Goedhart, H.A.; de Jong, I.C.; Kranen, R.W. Relationships between behavioral and meat quality characteristics of pigs raised under barren and enriched housing conditions. J. Anim. Sci. 2001, 79, 2835–2843. [Google Scholar] [CrossRef]
- Lebret, B.; Meunier-Salaün, M.C.; Foury, A.; Mormède, P.; Dransfield, E.; Dourmad, J.Y. Influence of rearing conditions on performance, behavioral, and physiological responses of pigs to preslaughter handling, carcass traits, and meat quality. J. Anim. Sci. 2006, 84, 2436–2447. [Google Scholar] [CrossRef] [Green Version]
- Lebret, B.; Prunier, A.; Bonhomme, N.; Foury, A.; Mormède, P.; Dourmad, J.Y. Physiological traits and meat quality of pigs as affected by genotype and housing system. Meat Sci. 2011, 88, 14–22. [Google Scholar] [CrossRef]
- Lefaucheur, L.; Lebret, B.; Ecolan, P.; Louveau, I.; Damon, M.; Prunier, A.; Billon, Y.; Sellier, P.; Gilbert, H. Muscle characteristics and meat quality traits are affected by divergent selection on residual feed intake in pigs. J. Anim. Sci. 2011, 89, 996–1010. [Google Scholar] [CrossRef]
- Smith, R.M.; Gabler, N.K.; Young, J.M.; Cai, W.; Boddicker, N.J.; Anderson, M.J.; Huff-Lonergan, E.; Dekkers, J.C.; Lonergan, S.M. Effects of selection for decreased residual feed intake on composition and quality of fresh pork. J. Anim. Sci. 2011, 89, 192–200. [Google Scholar] [CrossRef] [Green Version]
- Boddicker, N.; Gabler, N.K.; Spurlock, M.E.; Nettleton, D.; Dekkers, J.C. Effects of ad libitum and restricted feed intake on growth performance and body composition of Yorkshire pigs selected for reduced residual feed intake. J. Anim. Sci. 2011, 89, 40–51. [Google Scholar] [CrossRef] [PubMed]
- Ngapo, T.M.; Gariepy, C. Factors affecting the eating quality of pork. Crit. Rev. Food Sci. Nutr. 2008, 48, 599–633. [Google Scholar] [CrossRef] [PubMed]
- Marcon, A.V.; Caldara, F.R.; de Oliveira, G.F.; Gonçalves, L.M.P.; Garcia, R.G.; Paz, I.C.L.A.; Crone, C.; Marcon, A. Pork quality after electrical or carbon dioxide stunning at slaughter. Meat Sci. 2019, 156, 93–97. [Google Scholar] [CrossRef] [PubMed]
- Arkfeld, E.K.; Mohrhauser, D.A.; King, D.A.; Wheeler, T.L.; Dilger, A.C.; Shackelford, S.D.; Boler, D.D. Characterization of variability in pork carcass composition and primal quality. J. Anim. Sci. 2017, 95, 697–708. [Google Scholar] [CrossRef]
- Garrido, M.D.; Pedauyé, J.; Bañón, S.; Laencina, J. Objective assessment of pork quality. Meat Sci. 1994, 37, 411–420. [Google Scholar] [CrossRef]
- López-Pedrouso, M.; Lorenzo, J.M.; Gagaoua, M.; Franco, D. Application of Proteomic Technologies to Assess the Quality of Raw Pork and Pork Products: An Overview from Farm-To-Fork. Biology 2020, 9, 393. [Google Scholar] [CrossRef]
- Te Pas, M.F.; Kruijt, L.; Pierzchala, M.; Crump, R.E.; Boeren, S.; Keuning, E.; Hoving-Bolink, R.; Hortós, M.; Gispert, M.; Arnau, J.; et al. Identification of proteomic biomarkers in M. Longissimus dorsi as potential predictors of pork quality. Meat Sci. 2013, 95, 679–687. [Google Scholar] [CrossRef]
- Chen, Q.; Xu, H.; Huang, D. Analysis of free amino acids in bamaxiang pork and three different varieties of pork. Anhui Agric. Sci. 2016, 44, 97–99. [Google Scholar] [CrossRef]
- Gan, M.; Shen, L.; Chen, L.; Jiang, D.; Jiang, Y.; Li, Q.; Chen, Y.; Ge, G.; Liu, Y.; Xu, X.; et al. Meat Quality, Amino Acid, and Fatty Acid Composition of Liangshan Pigs at Different Weights. Animals 2020, 10, 822. [Google Scholar] [CrossRef]
- Guo, Q.; Kong, X.; Hu, C.; Zhou, B.; Wang, C.; Shen, Q.W. Fatty Acid Content, Flavor Compounds, and Sensory Quality of Pork Loin as Affected by Dietary Supplementation with l-arginine and Glutamic Acid. J. Food. Sci. 2019, 84, 3445–3453. [Google Scholar] [CrossRef] [PubMed]
- Madeira, M.S.; Alfaia, C.M.; Costa, P.; Lopes, P.A.; Lemos, J.P.; Bessa, R.J.; Prates, J.A. The combination of arginine and leucine supplementation of reduced crude protein diets for boars increases eating quality of pork. J. Anim. Sci. 2014, 92, 2030–2040. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.J.; Jiang, Q.Y.; Zhang, T.; Yin, Y.L.; Li, F.N.; Deng, J.P.; Wu, G.Y.; Kong, X.F. Dietary supplementation with arginine and glutamic acid modifies growth performance, carcass traits, and meat quality in growing-finishing pigs. J. Anim. Sci. 2017, 95, 2680–2689. [Google Scholar] [CrossRef]
- Yu, Q.; Cooper, B.; Sobreira, T.; Kim, Y.H.B. Utilizing Pork Exudate Metabolomics to Reveal the Impact of Aging on Meat Quality. Foods 2021, 10, 668. [Google Scholar] [CrossRef]
- Cao, M.; Han, Q.; Zhang, J.; Zhang, R.; Wang, J.; Gu, W.; Kang, W.; Lian, K.; Ai, L. An untargeted and pseudotargeted metabolomic combination approach to identify differential markers to distinguish live from dead pork meat by liquid chromatography-mass spectrometry. J. Chromatogr. A 2020, 1610, 460553. [Google Scholar] [CrossRef] [PubMed]
- Ueda, S.; Iwamoto, E.; Kato, Y.; Shinohara, M.; Shirai, Y.; Yamanoue, M. Comparative metabolomics of Japanese Black cattle beef and other meats using gas chromatography-mass spectrometry. Biosci. Biotechnol. Biochem. 2019, 83, 137–147. [Google Scholar] [CrossRef]
- Amin, A.B.; Zhang, L.; Zhang, J.; Mao, S. Metagenomic and Metabolomic Insights into the Mechanism Underlying the Disparity in Milk Yield of Holstein Cows. Front. Microbiol. 2022, 13, 844968. [Google Scholar] [CrossRef]
- Reecy, J.M.; Williams, J.E.; Kerley, M.S.; MacDonald, R.S.; Thornton, W.H., Jr.; Davis, J.L. The effect of postruminal amino acid flow on muscle cell proliferation and protein turnover. J. Anim. Sci. 1996, 74, 2158–2169. [Google Scholar] [CrossRef]
- Lobley, G.E.; Hoskin, S.O.; McNeil, C.J. Glutamine in animal science and production. J. Nutr. 2001, 131, 2525S–2531S. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Wang, C.; Huasai, S.; Chen, A. Metabolomics Reveals the Effects of High Dietary Energy Density on the Metabolism of Transition Angus Cows. Animals 2022, 12, 1147. [Google Scholar] [CrossRef]
- Holeček, M. Side effects of amino acid supplements. Physiol. Res. 2022, 71, 29–45. [Google Scholar] [CrossRef] [PubMed]
- Rivera-Antonio, A.; Rosales-Hernández, M.C.; Balbuena-Rebolledo, I.; Santiago-Quintana, J.M.; Mendieta-Wejebe, J.E.; Correa-Basurto, J.; García-Vázquez, J.B.; García-Báez, E.V.; Padilla-Martínez, I.I. Myeloperoxidase Inhibitory and Antioxidant Activities of (E)-2-Hydroxy-α-aminocinnamic Acids Obtained through Microwave-Assisted Synthesis. Pharmaceuticals 2021, 14, 513. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Zhang, W.; Cai, J.; Ni, Y.; Xiao, L.; Zhang, J. Transcriptome analysis in comparing carcass and meat quality traits of Jiaxing Black Pig and Duroc × Duroc × Berkshire × Jiaxing Black Pig crosses. Gene 2022, 808, 145978. [Google Scholar] [CrossRef] [PubMed]
- McFarland, G.A.; Holliday, R. Further evidence for the rejuvenating effects of the dipeptide l-carnosine on cultured human diploid fibroblasts. Exp. Gerontol. 1999, 34, 35–45. [Google Scholar] [CrossRef] [PubMed]
- Hipkiss, A.R. Glycation, ageing and carnosine: Are carnivorous diets beneficial? Mech. Ageing Dev. 2005, 126, 1034–1039. [Google Scholar] [CrossRef] [PubMed]
- Lan, S.; Qing-huan, L.; Zheng, T. L-Carnosine reduces telomere damage and shortening rate in cultured normal fibroblasts. Biochem. Biophys. Res. Commun. 2004, 324, 931–936. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Metabolites | KEGG ID | Super Class |
|---|---|---|
| Hypoxanthine | C00262 | Organoheterocyclic compounds |
| L-Fucose 1-phosphate | C02985 | Organic oxygen compounds |
| Ribose 1-phosphate | C00620 | Organic oxygen compounds |
| 4-Hydroxyproline | C01157 | Organic acids and derivatives |
| Beta-Alanyl-L-arginine | C05340 | Organic acids and derivatives |
| Citric acid | C00158 | Organic acids and derivatives |
| Creatine | C00300 | Organic acids and derivatives |
| D-Glutamine | C00819 | Organic acids and derivatives |
| Gamma-glutamyl-L-putrescine | C15699 | Organic acids and derivatives |
| Isocitric acid | C00311 | Organic acids and derivatives |
| L-Arginine | C00062 | Organic acids and derivatives |
| L-Glutamine | C00064 | Organic acids and derivatives |
| L-Lysine | C00047 | Organic acids and derivatives |
| L-Proline | C00148 | Organic acids and derivatives |
| L-Valine | C00183 | Organic acids and derivatives |
| N-Acetylornithine | C00437 | Organic acids and derivatives |
| Oxidized glutathione | C00127 | Organic acids and derivatives |
| Taurine | C00245 | Organic acids and derivatives |
| 6-Methylthioguanosine monophosphate | C16620 | Nucleosides, nucleotides, and analogues |
| Uridine diphosphate-N-acetylglucosamine | C00043 | Nucleosides, nucleotides, and analogues |
| PC (18:1(11Z)/0:0) | C04230 | Lipids and lipid-like molecules |
| PC (16:0/0:0) | C04230 | Lipids and lipid-like molecules |
| PC (18:0/0:0) | C04230 | Lipids and lipid-like molecules |
| PC (18:1(11Z)/16:0) | C00157 | Lipids and lipid-like molecules |
| PC (16:0/20:3(8Z,11Z,14Z)) | C00157 | Lipids and lipid-like molecules |
| PE (14:0/24:0) | C00350 | Lipids and lipid-like molecules |
| Metabolites | KEGG ID | Super Class |
|---|---|---|
| 2-Hydroxycinnamic acid | C01772 | Phenylpropanoids and polyketides |
| 4-[(Hydroxymethyl)nitrosoamino]-1-(3-pyridinyl)-1-butanone | C19563 | Organic oxygen compounds |
| Kanamycin | C01822 | Organic oxygen compounds |
| N-Acetylneuraminate 9-phosphate | C06241 | Organic oxygen compounds |
| 3-Methyl-L-histidine | C01152 | Organic acids and derivatives |
| 1-methylhistidine | C01152 | Organic acids and derivatives |
| 4-(Methylnitrosamino)-1-(3-pyridyl)-1-butanol glucuronide | C19605 | Organic acids and derivatives |
| Anserine | C01262 | Organic acids and derivatives |
| Carnosine | C00386 | Organic acids and derivatives |
| L-Histidine | C00135 | Organic acids and derivatives |
| Deoxycytidine | C00881 | Nucleosides, nucleotides, and analogues |
| Uridine 5′-monophosphate | C00105 | Nucleosides, nucleotides, and analogues |
| Glycerophosphocholine | C00670 | Lipids and lipid-like molecules |
| LysoPC (22:5(7Z,10Z,13Z,16Z,19Z)) | C04230 | Lipids and lipid-like molecules |
| Palmitoylcarnitine | C00547 | Benzenoids |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, C.; Zheng, J.; Xiong, C.; Zhou, H.; Wei, C.; Hu, X.; Qian, X.; He, M.; Shi, Y.; Liu, Y.; et al. Metabolomics Characterize the Differential Metabolic Markers between Bama Xiang Pig and Debao Pig to Identify Pork. Foods 2023, 12, 5. https://0-doi-org.brum.beds.ac.uk/10.3390/foods12010005
Chen C, Zheng J, Xiong C, Zhou H, Wei C, Hu X, Qian X, He M, Shi Y, Liu Y, et al. Metabolomics Characterize the Differential Metabolic Markers between Bama Xiang Pig and Debao Pig to Identify Pork. Foods. 2023; 12(1):5. https://0-doi-org.brum.beds.ac.uk/10.3390/foods12010005
Chicago/Turabian StyleChen, Changyi, Junwen Zheng, Chenyong Xiong, Hongjin Zhou, Chuntao Wei, Xin Hu, Xinxiu Qian, Mengyi He, Yandi Shi, Yuwen Liu, and et al. 2023. "Metabolomics Characterize the Differential Metabolic Markers between Bama Xiang Pig and Debao Pig to Identify Pork" Foods 12, no. 1: 5. https://0-doi-org.brum.beds.ac.uk/10.3390/foods12010005