
In Silico Evidence of the Multifunctional Features of Lactiplantibacillus pentosus LPG1, a Natural Fermenting Agent Isolated from Table Olive Biofilms

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strain, Culture Conditions, and DNA Isolation
2.2. Genome Sequencing, Assembly, and Annotation
2.3. Phylogenetic and Pan-Genome Analysis
2.4. Food Safety Assessment
2.5. Technological and Probiotic Genomic Assessment
2.6. Identification of Prophage, Genomic Island, and Other Insertion Sequences
2.7. Analysis of CRISPR−Cas Sequences
3. Results and Discussion
3.1. Genome Features
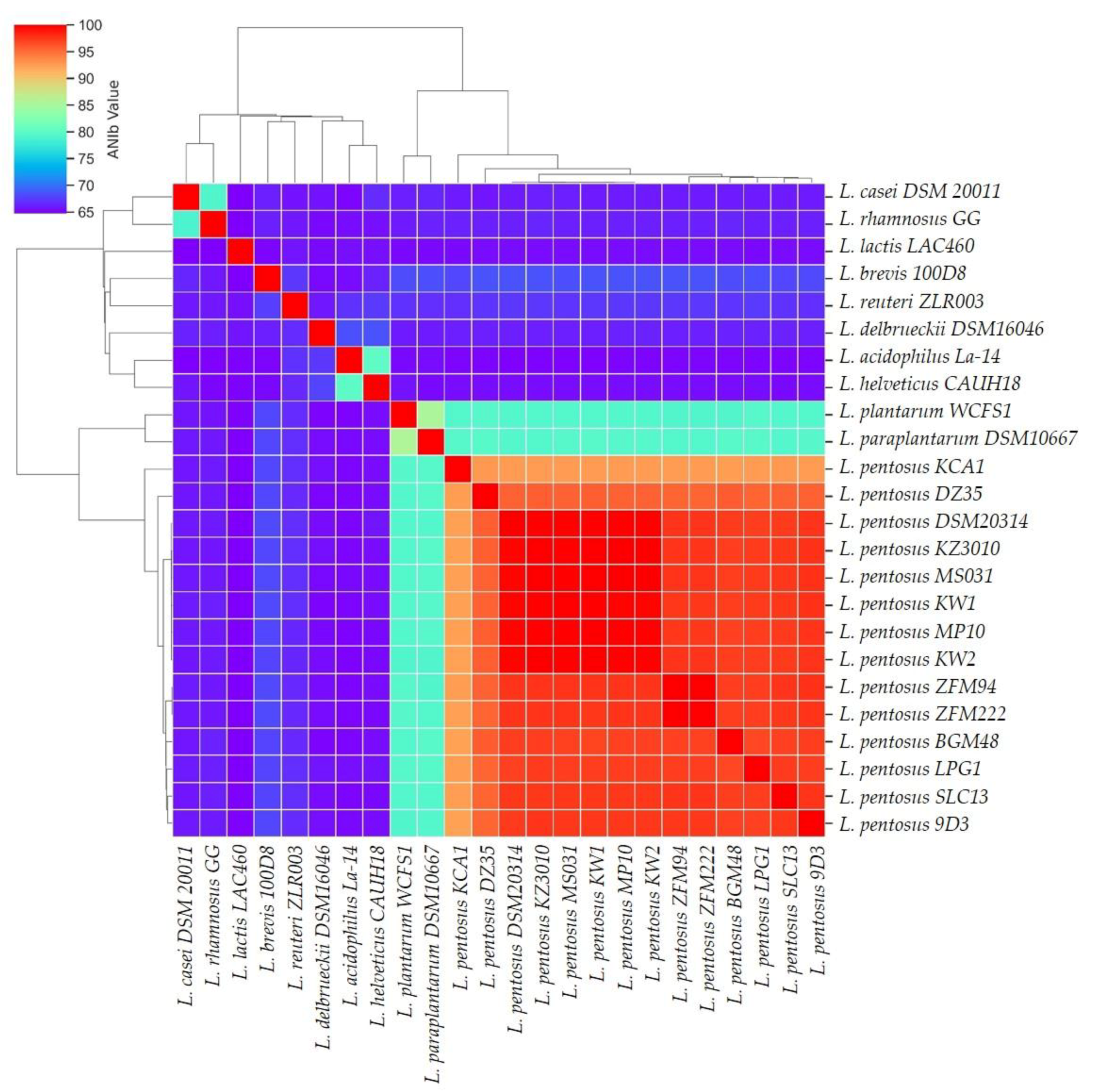
3.2. Phylogenetic and Comparative Analyses
3.3. Food Safety Assessment
3.4. Multifunctional Assessment
3.5. Identification of Mobile Genetic Elements
3.6. Identification of CRISPR−Cas Sequences
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Inglin, R.C.; Meile, L.; Stevens, M.J.A. Clustering of Pan- and Core-genome of Lactobacillus provides novel evolutionary insights for differentiation. BMC Genom. 2018, 19, 284. [Google Scholar] [CrossRef] [PubMed]
- IOC Economic Affairs & Promotion Unit—International Olive Council. Available online: https://www.internationaloliveoil.org/what-we-do/economic-affairs-promotion-unit/#figures%0Ahttps://www.internationaloliveoil.org/what-we- (accessed on 10 July 2022).
- Garrido-Fernandez, A.; Fernández Díez, M.J.; Adams, R.M. Table Olives: Production and Processing; Chapman & Hall: London, UK, 1997; pp. 67–109. [Google Scholar]
- Hurtado, A.; Reguant, C.; Bordons, A.; Rozès, N. Lactic acid bacteria from fermented table olives. Food Microbiol. 2012, 31, 1–8. [Google Scholar] [CrossRef]
- Arroyo-López, F.N.; Romero-Gil, V.; Bautista-Gallego, J.; Rodríguez-Gómez, F.; Jiménez-Díaz, R.; García-García, P.; Querol, A.; Garrido-Fernández, A. Potential benefits of the application of yeast starters in table olive processing. Front. Microbiol. 2012, 3, 161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grand View Research. Available online: https://www.grandviewresearch.com/industry-analysis/probiotics-market (accessed on 3 November 2022).
- Arroyo-López, F.N.; Bautista-Gallego, J.; Domínguez-Manzano, J.; Romero-Gil, V.; Rodríguez-Gómez, F.; García-García, P.; Garrido-Fernández, A.; Jiménez-Díaz, R. Formation of lactic acid bacteria-yeasts communities on the olive Surface during Spanish-style Manzanilla fermentations. Food Microbiol. 2012, 32, 295–301. [Google Scholar] [CrossRef] [PubMed]
- Benítez-Cabello, A.; Calero-Delgado, B.; Rodríguez-Gómez, F.; Garrido-Fernández, A.; Jiménez-Díaz, R.; Arroyo-López, F.N. Biodiversity and multifunctional features of lactic acid bacteria isolated from table olive biofilms. Front. Microbiol. 2019, 10, 836. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benítez-Cabello, A.; Torres-Maravilla, E.; Bermúdez-Humarán, L.; Langella, P.; Martín, R.; Jiménez-Díaz, R.; Arroyo-López, F.N. Probiotic Properties of Lactobacillus Strains Isolated from Table Olive Biofilms. Probiotics Antimicrob. Proteins 2020, 12, 1071–1082. [Google Scholar] [CrossRef]
- Maldonado-Barragán, A.; Caballero-Guerrero, B.; Lucena-Padrós, H.; Ruiz-Barba, J.L. Genome sequence of Lactobacillus pentosus IG1, a strain isolated from Spanish-style green olive fermentations. J. Bacteriol. 2011, 193, 5605. [Google Scholar] [CrossRef] [Green Version]
- Abriouel, H.; Benomar, N.; Pulido, R.P.; Cañamero, M.M.; Gálvez, A. Annotated genome sequence of Lactobacillus pentosus MP-10, which has probiotic potential, from naturally fermented Aloreña green table olives. J. Bacteriol. 2011, 193, 4559–4560. [Google Scholar] [CrossRef] [Green Version]
- Calero-Delgado, B.; Martín-Platero, A.M.; Pérez-Pulido, A.J.; Benítez-Cabello, A.; Casimiro-Soriguer, C.S.; Martínez-Bueno, M.; Arroyo-López, F.N.; Rodríguez-Gómez, F.; Bautista-Gallego, J.; Garrido-Fernández, A.; et al. Draft genome sequences of six Lactobacillus pentosus strains isolated from brines of traditionally fermented Spanish-style green table olives. Genome Announc. 2018, 6, e00379-18. [Google Scholar] [CrossRef] [Green Version]
- Calero-Delgado, B.; Pérez-Pulido, A.J.; Benítez-Cabello, A.; Martín-Platero, A.M.; Casimiro-Soriguer, C.S.; Martínez-Bueno, M.; Arroyo-López, F.N.; Díaz, R.J. Multiple Genome Sequences of Lactobacillus pentosus Strains Isolated from Biofilms on the Skin of Fermented Green Table Olives. Microbiol. Resour. Announc. 2019, 8, e01546-18. [Google Scholar] [CrossRef] [Green Version]
- Abriouel, H.; Manestsberger, J.; Caballero-Cómez, N.; Benomar, N. In silico genomic analysis of the potential probiotic Lactiplantibacillus pentosus CF2-10N reveals promising beneficial effects with health promoting properties. Front. Microbiol. 2022, 13, 4201. [Google Scholar] [CrossRef] [PubMed]
- Page, C.A.; Pérez-Díaz, I.M. Whole-Genome Sequencing and Annotation of Selected Lactobacillales Isolated from Commercial Cucumber Fermentation. Microbiol. Resour. Announc. 2021, 10, e00625-21. [Google Scholar] [CrossRef] [PubMed]
- EFSA. Guidance on the characterisation of microorganisms used as feed additives or 30 as production organisms. EFSA J. 2018, 16, 5206. [Google Scholar] [CrossRef]
- Martín-Platero, A.M.; Valdivia, E.; Maqueda, M.; Martínez-Bueno, M. Fast, convenient, and economical method for isolating genomic DNA from lactic acid bacteria using a modification of the protein “salting-out” procedure. Anal. Biochem. 2007, 366, 102–104. [Google Scholar] [CrossRef]
- Son, S.; Lee, R.; Park, S.-M.; Lee, S.H.; Lee, H.-K.; Kim, Y.; Shin, D. Complete genome sequencing and comparative genomic analysis of Lactobacillus acidophilus C5 as a potential canine probiotics. J. Anim. Sci. Technol. 2021, 63, 1411–1422. [Google Scholar] [CrossRef]
- Seemann, T. Prokka: Rapid prokaryotic genome annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef] [Green Version]
- Stothard, P.; Wishart, D.S. Circular genome visualization and exploration using CGView. Bioinformatics 2005, 21, 537–539. [Google Scholar] [CrossRef] [Green Version]
- Richter, M.; Rosselló-Móra, R.; Oliver Glöckner, F.; Peplies, J. JSpeciesWS: A web server for prokaryotic species circumscription based on pairwise genome comparison. Bioinformatics 2016, 32, 929–931. [Google Scholar] [CrossRef]
- Ning, W.; Wei, Y.; Gao, L.; Han, C.; Gou, Y.; Fu, S.; Liu, D.; Zhang, C.; Huang, X.; Wu, S.; et al. HemI 2.0: An online service for heatmap illustration. Nucleic Acids Res. 2022, 50, W405–W411. [Google Scholar] [CrossRef]
- Page, A.J.; Cummins, C.A.; Hunt, M.; Wong, V.K.; Reuter, S.; Holden, M.T.G.; Fookes, M.; Falush, D.; Keane, J.A.; Parkhill, J. Roary: Rapid large-scale prokaryote pan genome analysis. Bioinformatics 2015, 31, 3691–3693. [Google Scholar] [CrossRef] [Green Version]
- Alcock, B.P.; Raphenya, A.R.; Lau, T.T.; Tsang, K.K.; Bouchard, M.; Edalatmand, A.; Huynh, W.; Nguyen, A.L.V.; Cheng, A.A.; Liu, S.; et al. CARD 2020: Antibiotic resistome surveillance with the comprehensive antibiotic resistance database. Nucleic Acids Res. 2020, 48, D517–D525. [Google Scholar] [CrossRef] [PubMed]
- Bortolaia, V.; Kaas, R.S.; Ruppe, E.; Roberts, M.C.; Schwarz, S.; Cattoir, V.; Philippon, A.; Allesoe, R.L.; Rita Rebelo, A.; Ferrer Florensa, A.; et al. Mette Pinholt 16, Muna F. Anjum 18, Nicholas A. Duggett 18. J. Antimicrob. Chemother. 2020, 16, 3491–3500. [Google Scholar] [CrossRef] [PubMed]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zankari, E.; Allesøe, R.; Joensen, K.G.; Cavaco, L.M.; Lund, O.; Aarestrup, F.M. PointFinder: A novel web tool for WGS-based detection of antimicrobial resistance associated with chromosomal point mutations in bacterial pathogens. J. Antimicrob. Chemother. 2017, 72, 2764–2768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cosentino, S.; Voldby Larsen, M.; Møller Aarestrup, F.; Lund, O. PathogenFinder—Distinguishing Friend from Foe Using Bacterial Whole Genome Sequence Data. PLoS ONE 2013, 8, 77302. [Google Scholar] [CrossRef]
- Joensen, K.G.; Scheutz, F.; Lund, O.; Hasman, H.; Kaas, R.S.; Nielsen, E.M.; Aarestrup, F.M. Real-time whole-genome sequencing for routine typing, surveillance, and outbreak detection of verotoxigenic Escherichia coli. J. Clin. Microbiol. 2014, 52, 1501–1510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tetzschner, A.M.M.; Johnson, J.R.; Johnston, B.D.; Lund, O.; Scheutz, F. In Silico genotyping of Escherichia coli isolates for extraintestinal virulence genes by use of whole-genome sequencing data. J. Clin. Microbiol. 2020, 58, e01269-20. [Google Scholar] [CrossRef]
- Huerta-Cepas, J.; Szklarczyk, D.; Heller, D.; Hernández-Plaza, A.; Forslund, S.K.; Cook, H.; Mende, D.R.; Letunic, I.; Rattei, T.; Jensen, L.J.; et al. EggNOG 5.0: A hierarchical, functionally and phylogenetically annotated orthology resource based on 5090 organisms and 2502 viruses. Nucleic Acids Res. 2019, 47, D309–D314. [Google Scholar] [CrossRef] [Green Version]
- Aramaki, T.; Blanc-Mathieu, R.; Endo, H.; Ohkubo, K.; Kanehisa, M.; Goto, S.; Ogata, H. KofamKOALA: KEGG Ortholog assignment based on profile HMM and adaptive score threshold. Bioinformatics 2020, 36, 2251–2252. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Yohe, T.; Huang, L.; Entwistle, S.; Wu, P.; Yang, Z.; Busk, P.K.; Xu, Y.; Yin, Y. DbCAN2: A meta server for automated carbohydrate-active enzyme annotation. Nucleic Acids Res. 2018, 46, W95–W101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, Y.; Mao, X.; Yang, J.; Chen, X.; Mao, F.; Xu, Y. DbCAN: A web resource for automated carbohydrate-active enzyme annotation. Nucleic Acids Res. 2012, 40, W445–W451. [Google Scholar] [CrossRef] [PubMed]
- Van Heel, A.J.; De Jong, A.; Song, C.; Viel, J.H.; Kok, J.; Kuipers, O.P. BAGEL4: A user-friendly web server to thoroughly mine RiPPs and bacteriocins. Nucleic Acids Res. 2018, 46, W278–W281. [Google Scholar] [CrossRef] [PubMed]
- Blin, K.; Shaw, S.; Kloosterman, A.M.; Charlop-Powers, Z.; Van Wezel, G.P.; Medema, M.H.; Weber, T. AntiSMASH 6.0: Improving cluster detection and comparison capabilities. Nucleic Acids Res. 2021, 49, W29–W35. [Google Scholar] [CrossRef] [PubMed]
- Arndt, D.; Grant, J.R.; Marcu, A.; Sajed, T.; Pon, A.; Liang, Y.; Wishart, D.S. PHASTER: A better, faster version of the PHAST phage search tool. Nucleic Acids Res. 2016, 44, W16–W21. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Liang, Y.; Lynch, K.H.; Dennis, J.J.; Wishart, D.S. PHAST: A Fast Phage Search Tool. Nucleic Acids Res. 2011, 39, W347–W352. [Google Scholar] [CrossRef] [PubMed]
- Mihara, T.; Nishimura, Y.; Shimizu, Y.; Nishiyama, H.; Yoshikawa, G.; Uehara, H.; Hingamp, P.; Goto, S.; Ogata, H. Linking virus genomes with host taxonomy. Viruses 2016, 8, 66. [Google Scholar] [CrossRef] [Green Version]
- Nishimura, Y.; Yoshida, T.; Kuronishi, M.; Uehara, H.; Ogata, H.; Goto, S. ViPTree: The viral proteomic tree server. Bioinformatics 2017, 33, 2379–2380. [Google Scholar] [CrossRef]
- Bertelli, C.; Laird, M.R.; Williams, K.P.; Lau, B.Y.; Hoad, G.; Winsor, G.L.; Brinkman, F.S.L. IslandViewer 4: Expanded prediction of genomic islands for larger-scale datasets. Nucleic Acids Res. 2017, 45, W30–W35. [Google Scholar] [CrossRef]
- Néron, B.; Littner, E.; Haudiquet, M.; Perrin, A.; Cury, J.; Rocha, E.P.C. IntegronFinder 2.0: Identification and Analysis of Integrons across Bacteria, with a Focus on Antibiotic Resistance in Klebsiella. Microorganisms 2022, 10, 700. [Google Scholar] [CrossRef]
- Varani, A.M.; Siguier, P.; Gourbeyre, E.; Charneau, V.; Chandler, M. ISsaga is an ensemble of web-based methods for high throughput identification and semi-automatic annotation of insertion sequences in prokaryotic genomes. Genome Biol. 2011, 12, R30. [Google Scholar] [CrossRef] [Green Version]
- Siguier, P.; Perochon, J.; Lestrade, L.; Mahillon, J.; Chandler, M. ISfinder: The reference centre for bacterial insertion sequences. Nucleic Acids Res. 2006, 34, D32–D36. [Google Scholar] [CrossRef] [Green Version]
- Grissa, I.; Vergnaud, G.; Pourcel, C. CRISPRFinder: A web tool to identify clustered regularly interspaced short palindromic repeats. Nucleic Acids Res. 2007, 35, W52–W57. [Google Scholar] [CrossRef] [Green Version]
- Abby, S.S.; Néron, B.; Ménager, H.; Touchon, M.; Rocha, E.P.C. MacSyFinder: A program to mine genomes for molecular systems with an application to CRISPR-Cas systems. PLoS ONE 2014, 9, e110726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siezen, R.J.; Tzeneva, V.A.; Castioni, A.; Wels, M.; Phan, H.T.K.; Rademaker, J.L.W.; Starrenburg, M.J.C.; Kleerebezem, M.; van Hylckama Vlieg, J.E.T. Phenotypic and genomic diversity of Lactobacillus plantarum strains isolated from various environmental niches. Environ. Microbiol. 2010, 12, 758–773. [Google Scholar] [CrossRef] [PubMed]
- O’Sullivan, O.; O’Callaghan, J.; Sangrador-Vegas, A.; McAuliffe, O.; Slattery, L.; Kaleta, P.; Callanan, M.; Fitzgerald, G.F.; Ross, R.P.; Beresford, T. Comparative genomics of lactic acid bacteria reveals a niche-specific gene set. BMC Microbiol. 2009, 9, 50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holzapfel, W.H.; Haberer, P.; Geisen, R.; Björkroth, J.; Schillinger, U. Taxonomy and important features of probiotic microorganisms in food and nutrition. Am. J. Clin. Nutr. 2001, 73, 365s–373s. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jain, C.; Rodriguez-R, L.M.; Phillippy, A.M.; Konstantinidis, K.T.; Aluru, S. High throughput ANI analysis of 90K prokaryotic genomes reveals clear species boundaries. Nat. Commun. 2018, 9, 5114. [Google Scholar] [CrossRef] [Green Version]
- Torriani, S.; Clementi, F.; Vancanneyt, M.; Hoste, B.; Dellaglio, F.; Kersters, K. Differentiation of Lactobacillus plantarum, L. pentosus and L. paraplantarum species by RAPD-PCR and AFLP. Syst. Appl. Microbiol. 2001, 24, 554–560. [Google Scholar] [CrossRef]
- Arahal, D.R. Whole-genome analyses: Average nucleotide identity. In Methods in Microbiology; Academic Press: Cambridge, MA, USA, 2014; Volume 41, pp. 103–122. [Google Scholar]
- Schmid, J.; Sieber, V.; Rehm, B. Bacterial exopolysaccharides: Biosynthesis pathways and engineering strategies. Front. Microbiol. 2015, 6, 496. [Google Scholar] [CrossRef] [Green Version]
- Zeidan, A.A.; Poulsen, V.K.; Janzen, T.; Buldo, P.; Derkx, P.M.F.; Øregaard, G.; Neves, A.R. Polysaccharide production by lactic acid bacteria: From genes to industrial applications. FEMS Microbiol. Rev. 2017, 41, S168–S200. [Google Scholar] [CrossRef] [Green Version]
- Remus, D.M.; van Kranenburg, R.; van Swam, I.I.; Taverne, N.; Bongers, R.S.; Wels, M.; Wells, J.M.; Bron, P.A.; Kleerebezem, M. Impact of 4 Lactobacillus plantarum capsular polysaccharide clusters on surface glycan composition and host cell signaling. Microb. Cell Fact. 2012, 11, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, K.; Li, W.; Rui, X.; Chen, X.; Jiang, M.; Dong, M. Characterization of a novel exopolysaccharide with antitumor activity from Lactobacillus plantarum 70810. Int. J. Biol. Macromol. 2014, 63, 133–139. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Huang, R.; Shah, N.P.; Tao, X.; Xiong, Y.; Wei, H. Antioxidant and antibacterial activities of exopolysaccharides from Bifidobacterium bifidum WBIN03 and Lactobacillus plantarum R315. J. Dairy Sci. 2014, 97, 7334–7343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dertli, E.; Mayer, M.J.; Narbad, A. Impact of the exopolysaccharide layer on biofilms, adhesion and resistance to stress in Lactobacillus johnsonii FI9785. BMC Microbiol. 2015, 15, 8. [Google Scholar] [CrossRef] [Green Version]
- Whitehead, K.; Versalovic, J.; Roos, S.; Britton, R.A. Genomic and genetic characterization of the bile stress response of probiotic Lactobacillus reuteri ATCC 55730. Appl. Environ. Microbiol. 2008, 74, 1812–1819. [Google Scholar] [CrossRef] [Green Version]
- Lebeer, S.; Vanderleyden, J.; De Keersmaecker, S.C.J. Genes and Molecules of Lactobacilli Supporting Probiotic Action. Microbiol. Mol. Biol. Rev. 2008, 72, 728–764. [Google Scholar] [CrossRef] [Green Version]
- Choi, S.B.; Lew, L.C.; Yeo, S.K.; Nair Parvathy, S.; Liong, M.T. Probiotics and the BSH-related cholesterol lowering mechanism: A Jekyll and Hyde scenario. Crit. Rev. Biotechnol. 2015, 35, 392–401. [Google Scholar] [CrossRef]
- Buck, B.L.; Altermann, E.; Svingerud, T.; Klaenhammer, T.R. Functional analysis of putative adhesion factors in Lactobacillus acidophilus NCFM. Appl. Environ. Microbiol. 2005, 71, 8344–8351. [Google Scholar] [CrossRef] [Green Version]
- Heinemann, C.; Hylckama Vlieg, J.E.T.; Janssen, D.B.; Busscher, H.J.; Mei, H.C.; Reid, G. Purification and characterization of a surface-binding protein from Lactobacillus fermentum RC-14 that inhibits adhesion of Enterococcus faecalis 1131. FEMS Microbiol. Lett. 2000, 190, 177–180. [Google Scholar] [CrossRef]
- Kinoshita, H.; Uchida, H.; Kawai, Y.; Kawasaki, T.; Wakahara, N.; Matsuo, H.; Watanabe, M.; Kitazawa, H.; Ohnuma, S.; Miura, K.; et al. Cell surface Lactobacillus plantarum LA 318 glyceraldehyde-3-phosphate dehydrogenase (GAPDH) adheres to human colonic mucin. J. Appl. Microbiol. 2008, 104, 1667–1674. [Google Scholar] [CrossRef]
- Abriouel, H.; Pérez Montoro, B.; Casimiro-Soriguer, C.S.; Pérez Pulido, A.J.; Knapp, C.W.; Caballero Gómez, N.; Castillo-Gutiérrez, S.; Estudillo-Martínez, M.D.; Gálvez, A.; Benomar, N. Insight into potential probiotic markers predicted in Lactobacillus pentosus MP-10 genome sequence. Front. Microbiol. 2017, 8, 891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anukam, K.C.; Macklaim, J.M.; Gloor, G.B.; Reid, G.; Boekhorst, J.; Renckens, B.; van Hijum, S.A.F.T.; Siezen, R.J. Genome Sequence of Lactobacillus pentosus KCA1: Vaginal Isolate from a Healthy Premenopausal Woman. PLoS ONE 2013, 8, e59239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alayande, K.A.; Aiyegoro, O.A.; Nengwekhulu, T.M.; Katata-Seru, L.; Ateba, C.N. Integrated genome-based probiotic relevance and safety evaluation of Lactobacillus reuteri PNW1. PLoS ONE 2020, 15, e235873. [Google Scholar] [CrossRef] [PubMed]
- Zotta, T.; Giavalisco, M.; Parente, E.; Picariello, G.; Siano, F.; Ricciardi, A. Selection of Lactiplantibacillus Strains for the Production of Fermented Table Olives. Microorganisms 2022, 10, 625. [Google Scholar] [CrossRef]
- Hummel, A.S.; Hertel, C.; Holzapfel, W.H.; Franz, C.M.A.P. Antibiotic resistances of starter and probiotic strains of lactic acid bacteria. Appl. Environ. Microbiol. 2007, 73, 730–739. [Google Scholar] [CrossRef] [Green Version]
- Sharma, P.; Tomar, S.K.; Goswami, P.; Sangwan, V.; Singh, R. Antibiotic resistance among commercially available probiotics. Food Res. Int. 2014, 57, 176–195. [Google Scholar] [CrossRef]
- Gueimonde, M.; Sánchez, B.; de los Reyes-Gavilán, C.G.; Margolles, A. Antibiotic resistance in probiotic bacteria. Front. Microbiol. 2013, 4, 202. [Google Scholar] [CrossRef] [Green Version]
- Rhazi, N.; Delmarcelle, M.; Sauvage, E.; Jacquemotte, F.; Devriendt, K.; Tallon, V.; Ghosez, L.; Frère, J.-M. Specificity and reversibility of the transpeptidation reaction catalyzed by the Streptomyces R61 D-Ala-D-Ala peptidase. Protein Sci. 2005, 14, 2922–2928. [Google Scholar] [CrossRef] [Green Version]
- Arias, C.A.; Weisner, J.; Blackburn, J.M.; Reynolds, P.E. Serine and alanine racemase activities of VanT: A protein necessary for vancomycin resistance in Enterococcus gallinarum BM4174. Microbiology 2000, 146, 1727–1734. [Google Scholar] [CrossRef] [Green Version]
- Vaccalluzzo, A.; Pino, A.; Russo, N.; De Angelis, M.; Caggia, C.; Randazzo, C.L. FoodOmics as a new frontier to reveal microbial community and metabolic processes occurring on table olives fermentation. Food Microbiol. 2020, 92, 103606. [Google Scholar] [CrossRef]
- Stergiou, O.S.; Tegopoulos, K.; Kiousi, D.E.; Tsifintaris, M.; Papageorgiou, A.C.; Tassou, C.C.; Chorianopoulos, N.; Kolovos, P.; Galanis, A. Whole-Genome Sequencing, Phylogenetic and Genomic Analysis of Lactiplantibacillus pentosus L33, a Potential Probiotic Strain Isolated from Fermented Sausages. Front. Microbiol. 2021, 12, 3295. [Google Scholar] [CrossRef] [PubMed]
- CAZy—Bacteria. Available online: http://www.cazy.org/b96.html (accessed on 30 September 2022).
- Arnison, P.G.; Bibb, M.J.; Bierbaum, G.; Bowers, A.A.; Bugni, T.S.; Bulaj, G.; Camarero, J.A.; Campopiano, D.J.; Challis, G.L.; Clardy, J.; et al. Ribosomally synthesized and post-translationally modified peptide natural products: Overview and recommendations for a universal nomenclature. Nat. Prod. Rep. 2013, 30, 108–160. [Google Scholar] [CrossRef] [PubMed]
- Hurtado, A.; Ben Othman, N.; Chammem, N.; Hamdi, M.; Ferrer, S.; Reguant, C.; Bordons, A.; Rozès, N. Characterization of Lactobacillus isolates from fermented olives and their bacteriocin gene profiles. Food Microbiol. 2011, 28, 1514–1518. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, A.A.T.; Mantovani, H.C.; Jain, S. Bacteriocins from lactic acid bacteria and their potential in the preservation of fruit products. Crit. Rev. Biotechnol. 2017, 37, 852–864. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, J.M.; Martínez, M.I.; Kok, J. Pediocin PA-1, a wide-spectrum bacteriocin from lactic acid bacteria. Crit. Rev. Food Sci. Nutr. 2002, 42, 91–121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderssen, E.L.; Diep, D.B.; Nes, I.F.; Eijsink, V.G.H.; Nissen-Meyer, J. Antagonistic activity of Lactobacillus plantarum C11: Two new two- peptide bacteriocins, plantaricins EF and JK, and the induction factor plantaricin A. Appl. Environ. Microbiol. 1998, 64, 2269–2272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ekblad, B.; Kyriakou, P.K.; Oppegård, C.; Nissen-Meyer, J.; Kaznessis, Y.N.; Kristiansen, P.E. Structure-Function Analysis of the Two-Peptide Bacteriocin Plantaricin EF. Biochemistry 2016, 55, 5106–5116. [Google Scholar] [CrossRef] [PubMed]
- Azcarate-Peril, M.A.; McAuliffe, O.; Altermann, E.; Lick, S.; Russell, W.M.; Klaenhammer, T.R. Microarray analysis of a two-component regulatory system involved in acid resistance and proteolytic activity in Lactobacillus acidophilus. Appl. Environ. Microbiol. 2005, 71, 5794–5804. [Google Scholar] [CrossRef] [Green Version]
- Pfeiler, E.A.; Azcarate-Peril, M.A.; Klaenhammer, T.R. Characterization of a novel bile-inducible operon encoding a two-component regulatory system in Lactobacillus acidophilus. J. Bacteriol. 2007, 189, 4624–4634. [Google Scholar] [CrossRef] [Green Version]
- Lorca, G.L.; Font De Valdez, G.; Ljungh, Å. Characterization of the protein-synthesis dependent adaptive acid tolerance response in Lactobacillus acidophilus. J. Mol. Microbiol. Biotechnol. 2002, 4, 525–532. [Google Scholar]
- Silva, J.; Carvalho, A.S.; Ferreira, R.; Vitorino, R.; Amado, F.; Domingues, P.; Teixeira, P.; Gibbs, P.A. Effect of the pH of growth on the survival of Lactobacillus delbrueckii subsp. bulgaricus to stress conditions during spray-drying. J. Appl. Microbiol. 2005, 98, 775–782. [Google Scholar] [CrossRef] [PubMed]
- Bron, P.A.; Molenaar, D.; De Vos, W.M.; Kleerebezem, M. DNA micro-array-based identification of bile-responsive genes in Lactobacillus plantarum. J. Appl. Microbiol. 2006, 100, 728–738. [Google Scholar] [CrossRef]
- Hynönen, U.; Westerlund-Wikström, B.; Palva, A.; Korhonen, T.K. Identification by flagellum display of an epithelial cell- and fibronectin-binding function in the S1pA surface protein of Lactobacillus brevis. J. Bacteriol. 2002, 184, 3360–3367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tannock, G.W.; Ghazally, S.; Walter, J.; Loach, D.; Brooks, H.; Cook, G.; Surette, M.; Simmers, C.; Bremer, P.; Dal Bello, F.; et al. Ecological behavior of Lactobacillus reuteri 100-23 is affected by mutation of the luxS gene. Appl. Environ. Microbiol. 2005, 71, 8419–8425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vélez, M.P.; De Keersmaecker, S.C.J.; Vanderleyden, J. Adherence factors of Lactobacillus in the human gastrointestinal tract. FEMS Microbiol. Lett. 2007, 276, 140–148. [Google Scholar] [CrossRef] [Green Version]
- Walter, J.; Loach, D.M.; Alqumber, M.; Rockel, C.; Hermann, C.; Pfitzenmaier, M.; Tannock, G.W. D-Alanyl ester depletion of teichoic acids in Lactobacillus reuteri 100-23 results in impaired colonization of the mouse gastrointestinal tract. Environ. Microbiol. 2007, 9, 1750–1760. [Google Scholar] [CrossRef]
- Caggianiello, G.; Kleerebezem, M.; Spano, G. Exopolysaccharides produced by lactic acid bacteria: From health-promoting benefits to stress tolerance mechanisms. Appl. Microbiol. Biotechnol. 2016, 100, 3877–3886. [Google Scholar] [CrossRef]
- Silva, L.A.; Lopes Neto, J.H.P.; Cardarelli, H.R. Exopolysaccharides produced by Lactobacillus plantarum: Technological properties, biological activity, and potential application in the food industry. Ann. Microbiol. 2019, 69, 321–328. [Google Scholar] [CrossRef]
- Chabot, S.; Yu, H.L.; De Léséleuc, L.; Cloutier, D.; Van Calsteren, M.R.; Lessard, M.; Roy, D.; Lacroix, M.; Oth, D. Exopolysaccharides from Lactobacillus rhamnosus RW-9595M stimulate TNF, IL-6 and IL-12 in human and mouse cultured immunocompetent cells, and IFN-γ in mouse splenocytes. Lait 2001, 81, 683–697. [Google Scholar] [CrossRef] [Green Version]
- Vinderola, G.; Perdigón, G.; Duarte, J.; Farnworth, E.; Matar, C. Effects of the oral administration of the exopolysaccharide produced by Lactobacillus kefiranofaciens on the gut mucosal immunity. Cytokine 2006, 36, 254–260. [Google Scholar] [CrossRef]
- Duncan, S.H.; Louis, P.; Flint, H.J. Lactate-utilizing bacteria, isolated from human feces, that produce butyrate as a major fermentation product. Appl. Environ. Microbiol. 2004, 70, 5810–5817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scheppach, W. Effects of short chain fatty acids on gut morphology and function. Gut 1994, 35, S35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Crécy-Lagard, V.; El Yacoubi, B.; de la Garza, R.D.; Noiriel, A.; Hanson, A.D. Comparative genomics of bacterial and plant folate synthesis and salvage: Predictions and validations. BMC Genom. 2007, 8, 245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuchs, C.S.; Willett, W.C.; Colditz, G.A.; Hunter, D.J.; Stampfer, M.J.; Speizer, F.E.; Giovannucci, E.L. The influence of folate and multivitamin use on the familial risk of colon cancer in women. Cancer Epidemiol. Biomark. Prev. 2002, 11, 227–234. [Google Scholar]
- Moineau, S. Bacteriophage. In Brenner’s Encyclopedia of Genetics, 2nd ed.; Academic Press: Cambridge, MA, USA, 2013; pp. 280–283. ISBN 9780080961569. [Google Scholar]
- Pei, Z.; Sadiq, F.A.; Han, X.; Zhao, J.; Zhang, H.; Ross, R.P.; Lu, W.; Chen, W. Comprehensive Scanning of Prophages in Lactobacillus: Distribution, Diversity, Antibiotic Resistance Genes, and Linkages with CRISPR-Cas Systems. mSystems 2021, 6, e01211-20. [Google Scholar] [CrossRef]
- Ventura, M.; Canchaya, C.; Kleerebezem, M.; De Vos, W.M.; Siezen, R.J.; Brüssow, H. The prophage sequences of Lactobacillus plantarum strain WCFS1. Virology 2003, 316, 245–255. [Google Scholar] [CrossRef] [Green Version]
- Lugli, G.A.; Milani, C.; Turroni, F.; Tremblay, D.; Ferrario, C.; Mancabelli, L.; Duranti, S.; Ward, D.V.; Ossiprandi, M.C.; Moineau, S.; et al. Prophages of the genus Bifidobacterium as modulating agents of the infant gut microbiota. Environ. Microbiol. 2016, 18, 2196–2213. [Google Scholar] [CrossRef]
- Eraclio, G.; Fortina, M.G.; Labrie, S.J.; Tremblay, D.M.; Moineau, S. Characterization of prophages of Lactococcus garvieae. Sci. Rep. 2017, 7, 1856. [Google Scholar] [CrossRef] [Green Version]
- Brüssow, H.; Canchaya, C.; Hardt, W.-D. Phages and the Evolution of Bacterial Pathogens: From Genomic Rearrangements to Lysogenic Conversion. Microbiol. Mol. Biol. Rev. 2004, 68, 560–602. [Google Scholar] [CrossRef] [Green Version]
- Brüssow, H. Impact of Phages on Evolution of Bacterial Pathogenicity. Bact. Pathog. 2014, 267–300. [Google Scholar] [CrossRef]
- Quirós, P.; Colomer-Lluch, M.; Martínez-Castillo, A.; Miró, E.; Argente, M.; Jofre, J.; Navarro, F.; Muniesa, M. Antibiotic resistance genes in the bacteriophage DNA fraction of human fecal samples. Antimicrob. Agents Chemother. 2014, 58, 606–609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, Z.; Zhang, S.; Wei, Y.; Wang, M.; Ma, S.; Yang, S.; Wang, J.; Yuan, C.; Jiang, L.; Du, Y. Horizontal Gene Transfer Clarifies Taxonomic Confusion and Promotes the Genetic Diversity and Pathogenicity of Plesiomonas shigelloides. mSystems 2020, 5, e00448-20. [Google Scholar] [CrossRef] [PubMed]
- Brüssow, H. Phages of dairy bacteria. Annu. Rev. Microbiol. 2001, 55, 283–303. [Google Scholar] [CrossRef] [PubMed]
- Sulaiman, S.; Yusoff, N.S.; Mun, N.S.; Makmur, H.; Firdaus-Raih, M. Inference of horizontal gene transfer: Gaining insights into evolution via lateral acquisition of genetic material. Encycl. Bioinform. Comput. Biol. ABC Bioinform. 2018, 1–3, 953–964. [Google Scholar] [CrossRef]
- Ho Sui, S.J.; Fedynak, A.; Hsiao, W.W.L.; Langille, M.G.I.; Brinkman, F.S.L. The association of virulence factors with genomic Islands. PLoS ONE 2009, 4, e8094. [Google Scholar] [CrossRef]
- Hsiao, W.; Wan, I.; Jones, S.J.; Brinkman, F.S.L. IslandPath: Aiding detection of genomic islands in prokaryotes. Bioinformatics 2003, 19, 418–420. [Google Scholar] [CrossRef] [Green Version]
- Waack, S.; Keller, O.; Asper, R.; Brodag, T.; Damm, C.; Fricke, W.F.; Surovcik, K.; Meinicke, P.; Merkl, R. Score-based prediction of genomic islands in prokaryotic genomes using hidden Markov models. BMC Bioinform. 2006, 7, 142. [Google Scholar] [CrossRef] [Green Version]
- Langille, M.G.I.; Hsiao, W.W.L.; Brinkman, F.S.L. Evaluation of genomic island predictors using a comparative genomics approach. BMC Bioinform. 2008, 9, 329. [Google Scholar] [CrossRef] [Green Version]
- Kankainen, M.; Paulin, L.; Tynkkynen, S.; Von Ossowski, I.; Reunanen, J.; Partanen, P.; Satokari, R.; Vesterlund, S.; Hendrickx, A.P.A.; Lebeer, S.; et al. Comparative genomic analysis of Lactobacillus rhamnosus GG reveals pili containing a human-mucus binding protein. Proc. Natl. Acad. Sci. USA 2009, 106, 17193–17198. [Google Scholar] [CrossRef] [Green Version]
- Jeon, S.; Jung, J.; Kim, K.; Yoo, D.A.; Lee, C.; Kang, J.; Cho, K.; Kang, D.K.; Kwak, W.; Yoon, S.H.; et al. Comparative genome analysis of Lactobacillus plantarum GB-LP3 provides candidates of survival-related genetic factors. Infect. Genet. Evol. 2017, 53, 218–226. [Google Scholar] [CrossRef]
- Deng, Y.; Bao, X.; Ji, L.; Chen, L.; Liu, J.; Miao, J.; Chen, D.; Bian, H.; Li, Y.; Yu, G. Resistance integrons: Class 1, 2 and 3 integrons. Ann. Clin. Microbiol. Antimicrob. 2015, 14, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Portilha-Cunha, M.F.; Macedo, A.C.; Malcata, F.X. A review on adventitious lactic acid bacteria from table olives. Foods 2020, 9, 948. [Google Scholar] [CrossRef] [PubMed]
- Botta, C.; Cocolin, L. Microbial dynamics and biodiversity in table olive fermentation: Culture-dependent and -independent approaches. Front. Microbiol. 2012, 3, 245. [Google Scholar] [CrossRef] [Green Version]
- Patrick, T.; Ehrlich, S.D.; Chopin, M.C. Characterization of IS1201, an insertion sequence isolated from Lactobacillus helveticus. Gene 1994, 145, 75–79. [Google Scholar] [CrossRef]
- De Las Rivas, B.; Marcobal, Á.E.; Gómez, A.; Muñoz, R. Characterization of ISLpl4, a functional insertion sequence in Lactobacillus plantarum. Gene 2005, 363, 202–210. [Google Scholar] [CrossRef] [Green Version]
- Sybesma, W.; Molenaar, D.; van IJcken, W.; Venema, K.; Korta, R. Genome instability in Lactobacillus rhamnosus GG. Appl. Environ. Microbiol. 2013, 79, 2233–2239. [Google Scholar] [CrossRef] [Green Version]
- Abriouel, H.; Montoro, B.P.; Casado Muñoz, M.D.C.; Knapp, C.W.; Gálvez, A.; Benomar, N. In silico genomic insights into aspects of food safety and defense mechanisms of a potentially probiotic Lactobacillus pentosus MP-10 isolated from brines of naturally fermented Aloreña green table olives. PLoS ONE 2017, 12, e176801. [Google Scholar] [CrossRef] [Green Version]
- Clark, D.P.; Pazdernik, N.J.; McGehee, M.R. Genome Defense. Mol. Biol. 2019, 622–653. [Google Scholar] [CrossRef]
- Roberts, A.; Barrangou, R. Applications of CRISPR-Cas systems in lactic acid bacteria. FEMS Microbiol. Rev. 2020, 44, 523–537. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Function Involved | Gene | Locus | Annotation | Presence in Pangenome (%) | Reference |
|---|---|---|---|---|---|
| Acid stress resistance and tolerance bile salt | SecB_1 | LOPJBOPB_01128 | Protein-export chaperone SecB | 40 | [60] |
| secB_2 | LOPJBOPB_01741 | Protein-export chaperone SecB | 79 | [60] | |
| oppA_2 | LOPJBOPB_00084 | Peptide ABC transporter substrate-binding protein | 87 | [60] | |
| pva2 | LOPJBOPB_03149 | Choloylglycine hydrolase | 68 | [61] | |
| lmrA | LOPJBOPB_02793 | Multidrug resistance ABC transporter ATP-binding and permease protein | 40 | [59] | |
| emrB_1 | LOPJBOPB_00043 | Multidrug efflux MFS transporter | 24 | [59] | |
| speF | LOPJBOPB_00263 | Inducible ornithine decarboxylase | 16 | [60] | |
| Adhesion | LOPJBOPB_02921 | LOPJBOPB_02921 | MucBP domain-containing protein | 59 | [62,63] |
| LNFPBAJA_00018 | LNFPBAJA_00018 | MucBP domain-containing protein | 37 | [62,63] | |
| LOPJBOPB_01179 | LOPJBOPB_01179 | LPXTG cell wall anchor domain-containing protein | 24 | [60] | |
| gap_1 | LOPJBOPB_00266 | Glyceraldehyde-3-phosphate dehydrogenase | 11 | [64] | |
| Carbohydrate metabolism | glgP | LOPJBOPB_00022 | Glycogen phosphorylase | 81 | [65,66] |
| malS | LOPJBOPB_00166 | Alpha-1,4-glucan:maltose-1-phosphate maltosyltransferase | 57 | [65] | |
| mal_1 | LOPJBOPB_00161 | Oligo-1,6-glucosidase/alpha-amylase | 57 | [65,67] | |
| iolI | LOPJBOPB_03268 | Inosose isomerase/epimerase | 37 | [65] | |
| treP | LOPJBOPB_00084 | Alpha-trehalose phosphorylase | 25 | [60] | |
| ycjT | LOPJBOPB_03255 | Kojibiose phosphorylase | 19 | [65] | |
| abf2 | LOPJBOPB_03286 | Intracellular exo-alpha-L-arabinofuranosidase | 8 | [60] | |
| bgaA | LOPJBOPB_03283 | beta-galactosidase | 8 | [65,66] | |
| Response to stress | dps_2 | NFJKPBJO_00003 | DNA protection during starvation protein | 10 | [60,67] |
| Degradation phenolic compound | LOPJBOPB_02786 | LOPJBOPB_02786 | Tannase | 59 | [68] |
| LOPJBOPB_00821 | LOPJBOPB_00821 | Carboxylesterase | 81 | [68] | |
| LOPJBOPB_02878 | LOPJBOPB_02878 | PadR family transcriptional regulator | 56 | [68] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
López-García, E.; Benítez-Cabello, A.; Ramiro-García, J.; Ladero, V.; Arroyo-López, F.N. In Silico Evidence of the Multifunctional Features of Lactiplantibacillus pentosus LPG1, a Natural Fermenting Agent Isolated from Table Olive Biofilms. Foods 2023, 12, 938. https://0-doi-org.brum.beds.ac.uk/10.3390/foods12050938
López-García E, Benítez-Cabello A, Ramiro-García J, Ladero V, Arroyo-López FN. In Silico Evidence of the Multifunctional Features of Lactiplantibacillus pentosus LPG1, a Natural Fermenting Agent Isolated from Table Olive Biofilms. Foods. 2023; 12(5):938. https://0-doi-org.brum.beds.ac.uk/10.3390/foods12050938
Chicago/Turabian StyleLópez-García, Elio, Antonio Benítez-Cabello, Javier Ramiro-García, Victor Ladero, and Francisco Noé Arroyo-López. 2023. "In Silico Evidence of the Multifunctional Features of Lactiplantibacillus pentosus LPG1, a Natural Fermenting Agent Isolated from Table Olive Biofilms" Foods 12, no. 5: 938. https://0-doi-org.brum.beds.ac.uk/10.3390/foods12050938