Influence of Cooking Methods on Glucosinolates and Isothiocyanates Content in Novel Cruciferous Foods

 , ,
, , 
Abstract
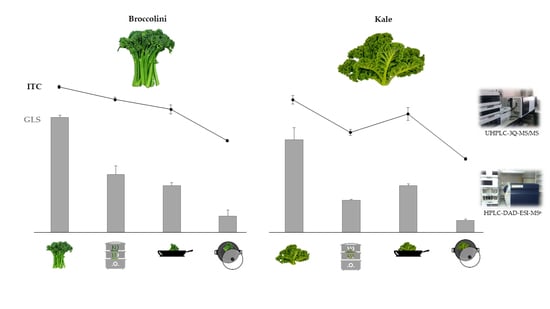
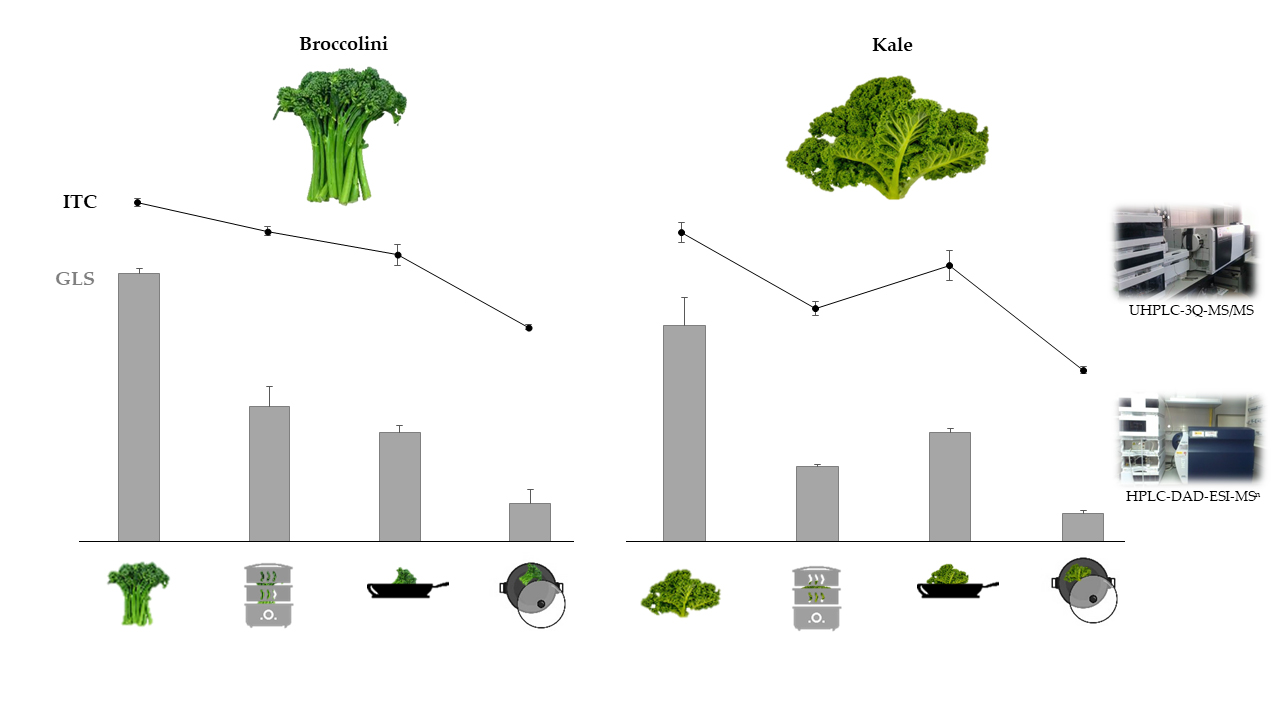
:
1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Extraction and Determination of Glucosinolates (GLS)
2.3. Extraction and Determination of Isothiocyanates (ITC)
2.4. Statistical Analysis
3. Results and Discussion
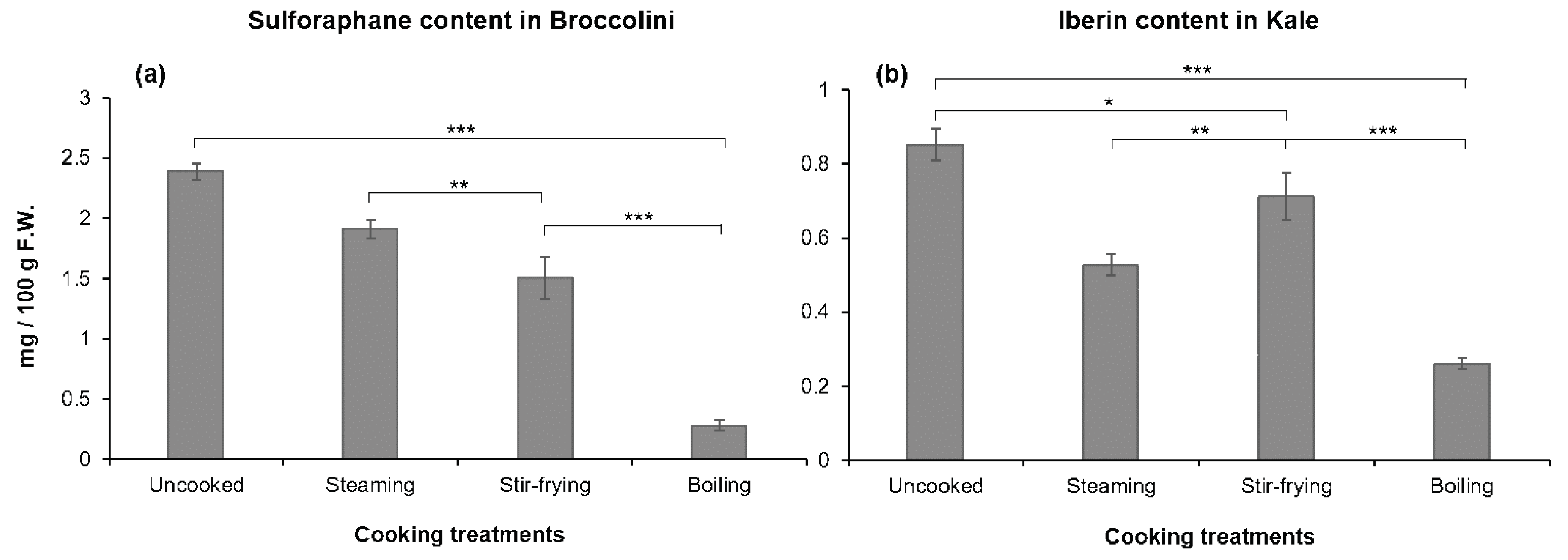
3.1. Glucosinolates Content of Vegetables: Effects of Cooking Methods
3.2. Isothiocyanate Content of Vegetables: Effects of Cooking Methods
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Royston, K.J.; Tollefsbol, T.O. The Epigenetic Impact of Cruciferous Vegetables on Cancer Prevention. Curr. Pharmacol. Rep. 2015, 1, 46–51. [Google Scholar] [CrossRef] [PubMed]
- Sita, G.; Hrelia, P.; Tarozzi, A.; Morroni, F. Isothiocyanates Are Promising Compounds against Oxidative Stress, Neuroinflammation and Cell Death that May Benefit Neurodegeneration in Parkinson’s Disease. Int. J. Mol. Sci. 2016, 17, 1454. [Google Scholar] [CrossRef] [PubMed]
- Fahey, J.W.; Zalcmann, A.T.; Talalay, P. The chemical diversity and distribution of glucosinolates and isothiocyanates among plants. Phytochemistry 2001, 56, 5–51. [Google Scholar] [CrossRef]
- Stefanson, A.; Bakovic, M. Dietary Regulation of Keap1/Nrf2/ARE Pathway: Focus on Plant-Derived Compounds and Trace Minerals. Nutrients 2014, 6, 3777–3801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Angelino, D.; Jeffery, E. Glucosinolate hydrolysis and bioavailability of resulting isothiocyanates: Focus on glucoraphanin. J. Funct. Foods 2014, 7, 67–76. [Google Scholar] [CrossRef]
- Dosz, E.B.; Ku, K.-M.; Juvik, J.A.; Jeffery, E.H. Total Myrosinase Activity Estimates in Brassica Vegetable Produce. J. Agric. Food Chem. 2014, 62, 8094–8100. [Google Scholar] [CrossRef]
- Soares, A.; Carrascosa, C.; Raposo, A. Influence of Different Cooking Methods on the Concentration of Glucosinolates and Vitamin C in Broccoli. Food Bioprocess. Technol. 2017, 10, 1387–1411. [Google Scholar] [CrossRef]
- Tabart, J.; Pincemail, J.; Kevers, C.; Defraigne, J.-O.; Dommes, J. Processing effects on antioxidant, glucosinolate, and sulforaphane contents in broccoli and red cabbage. Eur. Food Res. Technol. 2018, 244, 2085–2094. [Google Scholar] [CrossRef]
- Garcia-Viguera, C.; Soler-Rivas, C. Effect of Cooking on the Bioactive Compounds. In Frontiers in Bioactive Compounds; Bentham Science Publishers: Sharjah, United Arab Emirates, 2017; pp. 383–411. [Google Scholar]
- Pellegrini, N.; Chiavaro, E.; Gardana, C.; Mazzeo, T.; Contino, D.; Gallo, M.; Riso, P.; Fogliano, V.; Porrini, M. Effect of Different Cooking Methods on Color, Phytochemical Concentration, and Antioxidant Capacity of Raw and Frozen Brassica Vegetables. J. Agric. Food Chem. 2010, 58, 4310–4321. [Google Scholar] [CrossRef]
- Oliviero, T.; Verkerk, R.; Dekker, M. Isothiocyanates from Brassica Vegetables-Effects of Processing, Cooking, Mastication, and Digestion. Mol. Nutr. Food Res. 2018, 62, 1701069. [Google Scholar] [CrossRef]
- Block, E. Challenges and Artifact Concerns in Analysis of Volatile Sulfur Compounds. In Volatile Sulfur Compounds in Food; Qian, M.C., Fan, X., Mahattanatawee, K., Eds.; American Chemical Society Symposium Series; Oxford University Press: Oxford, UK, 2011; Volume 1068, pp. 35–63. [Google Scholar]
- Martínez-Hernández, G.B.; Artés-Hernández, F.; Gómez, P.A.; Artés, F. Comparative behaviour between kailan-hybrid and conventional fresh-cut broccoli throughout shelf-life. LWT-Food Sci. Technol. 2013, 50, 298–305. [Google Scholar] [CrossRef]
- Martínez-Hernández, G.B.; Gómez, P.A.; Artés, F.; Artés-Hernández, F. Nutritional quality changes throughout shelf-life of fresh-cut kailan-hybrid and ‘Parthenon’ broccoli as affected by temperature and atmosphere composition. Food Sci. Technol. Int. 2015, 21, 14–23. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Hernández, G.B.; Artés-Hernández, F.; Gómez, P.A.; Artés, F. Induced changes in bioactive compounds of kailan-hybrid broccoli after innovative processing and storage. J. Funct. Foods 2013, 5, 133–143. [Google Scholar] [CrossRef]
- Yang, Y.; Zhang, X. Extraction, Identification and Comparison of Glucosinolates Profiles in the Seeds of Broccolini, Broccoli and Chinese Broccoli. Solvent Extr. Res. Dev. Jpn. 2012, 19, 153–160. [Google Scholar] [CrossRef] [Green Version]
- Nilsson, J.; Olsson, K.; Engqvist, G.; Ekvall, J.; Olsson, M.; Nyman, M.; Akesson, B. Variation in the content of glucosinolates, hydroxycinnamic acids, carotenoids, total antioxidant capacity and low-molecular-weight carbohydrates in Brassica vegetables. J. Sci. Food Agric. 2006, 86, 528–538. [Google Scholar] [CrossRef]
- Giorgetti, L.; Giorgi, G.; Cherubini, E.; Gervasi, P.G.; Della Croce, C.M.; Longo, V.; Bellani, L. Screening and identification of major phytochemical compounds in seeds, sprouts and leaves of Tuscan black kale Brassica oleracea (L.) ssp acephala (DC) var. sabellica L. Nat. Prod. Res. 2018, 32, 1617–1626. [Google Scholar] [CrossRef] [PubMed]
- Vallejo, F.; Tomás-Barberán, B.; García-Viguera, C. Glucosinolates and vitamin C content in edible parts of broccoli florets after domestic cooking. Eur. Food Res. Technol. 2002, 215, 310–316. [Google Scholar] [CrossRef]
- Moreno, D.A.; López-Berenguer, C.; García-Viguera, C. Effects of Stir-Fry Cooking with Different Edible Oils on the Phytochemical Composition of Broccoli. J. Food Sci. 2007, 72, S064–S068. [Google Scholar] [CrossRef] [PubMed]
- Baenas, N.; García-Viguera, C.; Moreno, D.A. Biotic Elicitors Effectively Increase the Glucosinolates Content in Brassicaceae Sprouts. J. Agric. Food Chem. 2014, 62, 1881–1889. [Google Scholar] [CrossRef]
- Baenas, N.; Suárez-Martínez, C.; García-Viguera, C.; Moreno, D.A. Bioavailability and new biomarkers of cruciferous sprouts consumption. Food Res. Int. 2017, 100 Pt 1, 497–503. [Google Scholar] [CrossRef]
- Rodríguez-Hernández, M.C.; Medina, S.; Gil-Izquierdo, A.; Martínez-Ballesta, C.; Moreno, D.A. Broccoli isothiocyanates content and in vitro availability according to variety and origin. Maced. J. Chem. Chem. Eng. 2013, 32, 251–264. [Google Scholar] [CrossRef] [Green Version]
- Cartea, M.E.; Velasco, P.; Obregón, S.; Padilla, G.; de Haro, A. Seasonal variation in glucosinolate content in Brassica oleracea crops grown in northwestern Spain. Phytochemistry 2008, 69, 403–410. [Google Scholar] [CrossRef] [PubMed]
- Korus, A.; Słupski, J.; Gębczyński, P.; Banaś, A. Effect of preliminary processing and method of preservation on the content of glucosinolates in kale (Brassica oleracea L. var. acephala) leaves. LWT-Food Sci. Technol. 2014, 59, 1003–1008. [Google Scholar] [CrossRef]
- Velasco, P.; Cartea, M.E.; González, C.; Vilar, M.; Ordás, A. Factors Affecting the Glucosinolate Content of Kale (Brassica oleracea acephala Group). J. Agric. Food Chem. 2007, 55, 955–962. [Google Scholar] [CrossRef] [PubMed]
- Florkiewicz, A.; Ciska, E.; Filipiak-Florkiewicz, A.; Topolska, K. Comparison of Sous-vide methods and traditional hydrothermal treatment on GLS content in Brassica vegetables. Eur. Food Res. Technol. 2017, 243, 1507–1517. [Google Scholar] [CrossRef]
- Gründemann, C.; Huber, R. Chemoprevention with isothiocyanates—From bench to bedside. Cancer Lett. 2018, 414, 26–33. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Hernández, G.B.; Artés-Hernández, F.; Colares-Souza, F.; Gómez, P.A.; García-Gómez, P.; Artés, F. Innovative Cooking Techniques for Improving the Overall Quality of a Kailan-Hybrid Broccoli. Food Bioprocess. Technol. 2013, 6, 2135–2149. [Google Scholar] [CrossRef]
- Yuan, G.; Sun, B.; Yuan, J.; Wang, Q. Effects of different cooking methods on health-promoting compounds of broccoli. J. Zhejiang Univ. Sci. B 2009, 10, 580–588. [Google Scholar] [CrossRef] [Green Version]
- Rungapamestry, V.; Duncan, A.J.; Fuller, Z.; Ratcliffe, B. Effect of cooking brassica vegetables on the subsequent hydrolysis and metabolic fate of glucosinolates. Proc. Nutr. Soc. 2007, 66, 69–81. [Google Scholar] [CrossRef]
- Cieślik, E.; Leszczyńska, T.; Filipiak-Florkiewicz, A.; Sikora, E.; Pisulewski, P.M. Effects of some technological processes on glucosinolate contents in cruciferous vegetables. Food Chem. 2007, 105, 976–981. [Google Scholar] [CrossRef]
- Sarvan, I.; Kramer, E.; Bouwmeester, H.; Dekker, M.; Verkerk, R. Sulforaphane formation and bioaccessibility are more affected by steaming time than meal composition during in vitro digestion of broccoli. Food Chem. 2017, 214, 580–586. [Google Scholar] [CrossRef] [PubMed]
- Song, L.; Thornalley, P.J. Effect of storage, processing and cooking on glucosinolate content of Brassica vegetables. Food Chem. Toxicol. 2007, 45, 216–224. [Google Scholar] [CrossRef] [PubMed]
- Russo, M.; Spagnuolo, C.; Russo, G.L.; Skalicka-Woźniak, K.; Daglia, M.; Sobarzo-Sánchez, E.; Nabavi, S.F.; Nabavi, S.M. Nrf2 targeting by sulforaphane: A potential therapy for cancer treatment. Crit. Rev. Food Sci. Nutr. 2018, 58, 1391–1405. [Google Scholar] [CrossRef] [PubMed]
- Jeffery, E.H.; Stewart, K.E. Upregulation of Quinone Reductase by Glucosinolate Hydrolysis Products from Dietary Broccoli. Methods Enzymol. 2004, 382, 457–469. [Google Scholar] [CrossRef] [PubMed]
- Biegańska-Marecik, R.; Radziejewska-Kubzdela, E.; Marecik, R. Characterization of phenolics, glucosinolates and antioxidant activity of beverages based on apple juice with addition of frozen and freeze-dried curly kale leaves (Brassica oleracea L. var. acephala L.). Food Chem. 2017, 230, 271–280. [Google Scholar] [CrossRef]
- Verkerk, R.; Schreiner, M.; Krumbein, A.; Ciska, E.; Holst, B.; Rowland, I.; De Schrijver, R.; Hansen, M.; Gerhäuser, C.; Mitchen, R.; et al. Glucosinolates in Brassica vegetables: The influence of the food supply chain on intake, bioavailability and human health. Mol. Nutr. Food Res. 2009, 53, S219. [Google Scholar] [CrossRef] [PubMed]
- Verkerk, R.; Dekker, M.; Jongen, W.M. Post-harvest increase of indolyl glucosinolates in response to chopping and storage of Brassica vegetables. J. Sci. Food Agric. 2001, 81, 953–958. [Google Scholar] [CrossRef]
- Palermo, M.; Pellegrini, N.; Fogliano, V. The effect of cooking on the phytochemical content of vegetables. J. Sci. Food Agric. 2014, 94, 1057–1070. [Google Scholar] [CrossRef] [PubMed]
- Jadhav, U.; Ezhilarasan, R.; Vaughn, S.F.; Berhow, M.A.; Mohanam, S. Iberin induces cell cycle arrest and apoptosis in human neuroblastoma cells. Int. J. Mol. Med. 2007, 19, 353–361. [Google Scholar] [CrossRef] [Green Version]
- Wagner, A.E.; Terschluesen, A.M.; Rimbach, G. Health promoting effects of brassica-derived phytochemicals: From chemopreventive and anti-inflammatory activities to epigenetic regulation. Oxid. Med. Cell. Longev. 2013, 2013, 964539. [Google Scholar] [CrossRef]
- Jones, R.B.; Frisina, C.L.; Winkler, S.; Imsic, M.; Tomkins, R.B. Cooking method significantly effects glucosinolate content and sulforaphane production in broccoli florets. Food Chem. 2010, 123, 237–242. [Google Scholar] [CrossRef]
- Ciska, E.; Drabińska, N.; Honke, J.; Narwojsz, A. Boiled Brussels sprouts: A rich source of glucosinolates and the corresponding nitriles. J. Funct. Foods 2015, 19, 91–99. [Google Scholar] [CrossRef]
- Galgano, F.; Favati, F.; Caruso, M.; Pietrafesa, A.; Natella, S. The Influence of Processing and Preservation on the Retention of Health-Promoting Compounds in Broccoli. J. Food Sci. 2007, 72, S130–S135. [Google Scholar] [CrossRef] [PubMed]
- Bayat Mokhtari, R.; Baluch, N.; Homayouni, T.S.; Morgatskaya, E.; Kumar, S.; Kazemi, P.; Herman, Y. The role of Sulforaphane in cancer chemoprevention and health benefits: A mini-review. J. Cell Commun. Signal. 2018, 12, 91–101. [Google Scholar] [CrossRef] [PubMed]
- Briones-Herrera, A.; Eugenio-Pérez, D.; Reyes-Ocampo, J.G.; Rivera-Mancía, S.; Pedraza-Chaverri, J. New highlights on the health-improving effects of sulforaphane. Food Funct. 2018, 9, 2589–2606. [Google Scholar] [CrossRef]
- Luang-In, V.; Deeseenthum, S.; Udomwong, P.; Saengha, W.; Gregori, M. Formation of Sulforaphane and Iberin Products from Thai Cabbage Fermented by Myrosinase-Positive Bacteria. Molecules 2018, 23, 955. [Google Scholar] [CrossRef] [PubMed]
- Vieites-Outes, C.; Lopez-Hernandez, J.; Lage-Yusty, M.A. Modification of glucosinolates in turnip greens (Brassica rapa subsp. rapa L.) subjected to culinary heat processes. CyTA J. Food 2009, 14, 536–540. [Google Scholar] [CrossRef]
- Nugrahedi, P.Y.; Verkerk, R.; Widianarko, B.; Dekker, M. A Mechanistic Perspective on Process-Induced Changes in Glucosinolate Content in Brassica Vegetables: A Review. Crit. Rev. Food Sci. Nutr. 2015, 55, 823–838. [Google Scholar] [CrossRef]
- Bricker, G.V.; Riedl, K.M.; Ralston, R.A.; Tober, K.L.; Oberyszyn, T.M.; Schwartz, S.J. Isothiocyanate metabolism, distribution, and interconversion in mice following consumption of thermally processed broccoli sprouts or purified sulforaphane. Mol. Nutr. Food Res. 2014, 58, 1991–2000. [Google Scholar] [CrossRef]
- Liang, H.; Wei, Y.; Li, R.; Cheng, L.; Yuan, Q.; Zheng, F. Intensifying sulforaphane formation in broccoli sprouts by using other cruciferous sprouts additions. Food Sci. Biotechnol. 2018, 27, 957–962. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Glucosinolate | Semi-Systematic Name | Rt (min) | [M-H]− (m/z) | Broccolini | Kale |
|---|---|---|---|---|---|
| Glucoiberin | 3-methylsulfinylpropyl-gls | 4.0 | 422 | 0 1 | + |
| Progoitrin | 2-hydroxy-3-butenyl-gls | 4.2 | 388 | + | 0 |
| Glucoraphanin | 4-methylsulfinylbutyl-gls | 4.6 | 436 | + | + |
| Sinigrin | 2-propenyl-gls | 5.7 | 358 | 0 | + |
| Gluconapin | 3-butenyl-gls | 7.8 | 372 | + | + |
| 4-Hydroxyglucobrassicin | 4-hydroxy-3-indolylmethyl-gls | 11.0 | 463 | + | + |
| Glucosinalbin | 4-hydroxybenzyl-gls | 13.6 | 424 | + | 0 |
| Glucobrassicanapin | 4-pentenyl-gls | 17.2 | 386 | 0 | + |
| Glucobrassicin | 3-indolylmethyl-gls | 20.0 | 447 | + | + |
| Gluconasturtin | 2-phenylethyl-gls | 22.1 | 422 | + | + |
| 4-Methoxyglucobrassicin | 4-methoxy-3-indolylmethyl-gls | 23.5 | 477 | + | + |
| Neoglucobrassicin | N-methoxy-3-indolylmethyl-gls | 25.8 | 477 | + | 0 |
| Glucosinolates | Broccolini | |||||||
| Fresh | Steaming | Stir-frying | Boiling | |||||
| Progoitrin | 31.76 | ±6.6 1 | ||||||
| Glucoraphanin | 78.74 | ±11.6 a | 58.37 | ±4.3 b | 56.27 | ±3.5 b | 16.86 | ±6.3 c |
| 4-Hydroxiglucobrassicin | 9.61 | ±1.1 a | 4.68 | ±2.4 b | 3.33 | ±1.2 b | 1.30 | ±1.5 b |
| Glucobrassicin | 72.11 | ±4.5 a | 27.07 | ±6.4 b | 10.08 | ±1.5 c | 7.16 | ±1.9 c |
| 4-Methoxyglucobrassicin | 15.10 | ±0.7 a | 5.69 | ±2.1 b | 4.17 | ±0.3 b | 1.68 | ±0.5 c |
| Neoglucobrassicin | 43.55 | ±3.3 a | 21.51 | ±4.5 b | 8.84 | ±0.7 c | 5.93 | ±1.3 c |
| Aliphatic | 110.49 | ±7.8 a | 58.37 | ±4.3 b | 56.27 | ±3.5 b | 16.86 | ±6.3 c |
| Indolic | 68.26 | ±5.1 a | 31.88 | ±9.0 b | 16.34 | ±1.8 c | 8.91 | ±3.3 c |
| Total | 178.76 | ±3.4 a | 90.24 | ±13.1 b | 72.60 | ±4.7 b | 25.77 | ±8.9 c |
| Glucosinolates | Kale | |||||||
| Fresh | Steaming | Stir-frying | Boiling | |||||
| Glucoiberin | 11.58 | ±1.3 a | 8.05 | ±0.1 b | 9.32 | ±0.6 a | 3.45 | ±0.1 b |
| Sinigrin | 37.27 | ±6.6 a | 4.09 | ±0.5 c | 6.60 | ±0.2 b | 2.09 | ±0.5 c |
| 4-Hydroxiglucobrassicin | 1.34 | ±0.2 b | 1.51 | ±0.2 b | 2.81 | ±0.4 a | 0.46 | ±0.2 c |
| Glucobrassicin | 2.44 | ±0.3 b | 3.32 | ±0.2 b | 6.13 | ±0.9 a | 0.55 | ±0.1 c |
| 4-Methoxyglucobrassicin | 1.90 | ±0.1 a | 2.14 | ±0.5 a | 2.85 | ±0.8 a | 0.57 | ±0.1 b |
| Aliphatic | 48.85 | ±7.8 a | 12.14 | ±0.6 c | 15.92 | ±0.8 b | 5.54 | ±0.6 d |
| Indolic | 5.68 | ±0.6 b | 6.96 | ±0.2 b | 11.79 | ±1.6 a | 1.58 | ±0.2 c |
| Total | 54.54 | ±7.3 a | 19.11 | ±0.3 c | 27.71 | ±0.9 b | 7.12 | ±0.7 d |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baenas, N.; Marhuenda, J.; García-Viguera, C.; Zafrilla, P.; Moreno, D.A. Influence of Cooking Methods on Glucosinolates and Isothiocyanates Content in Novel Cruciferous Foods. Foods 2019, 8, 257. https://0-doi-org.brum.beds.ac.uk/10.3390/foods8070257
Baenas N, Marhuenda J, García-Viguera C, Zafrilla P, Moreno DA. Influence of Cooking Methods on Glucosinolates and Isothiocyanates Content in Novel Cruciferous Foods. Foods. 2019; 8(7):257. https://0-doi-org.brum.beds.ac.uk/10.3390/foods8070257
Chicago/Turabian StyleBaenas, Nieves, Javier Marhuenda, Cristina García-Viguera, Pilar Zafrilla, and Diego A. Moreno. 2019. "Influence of Cooking Methods on Glucosinolates and Isothiocyanates Content in Novel Cruciferous Foods" Foods 8, no. 7: 257. https://0-doi-org.brum.beds.ac.uk/10.3390/foods8070257