Identification and Structural Elucidation of Anti-Inflammatory Compounds from Chinese Olive (Canarium Album L.) Fruit Extracts


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. General Experimental Procedures
2.2. Chemicals and Solvents
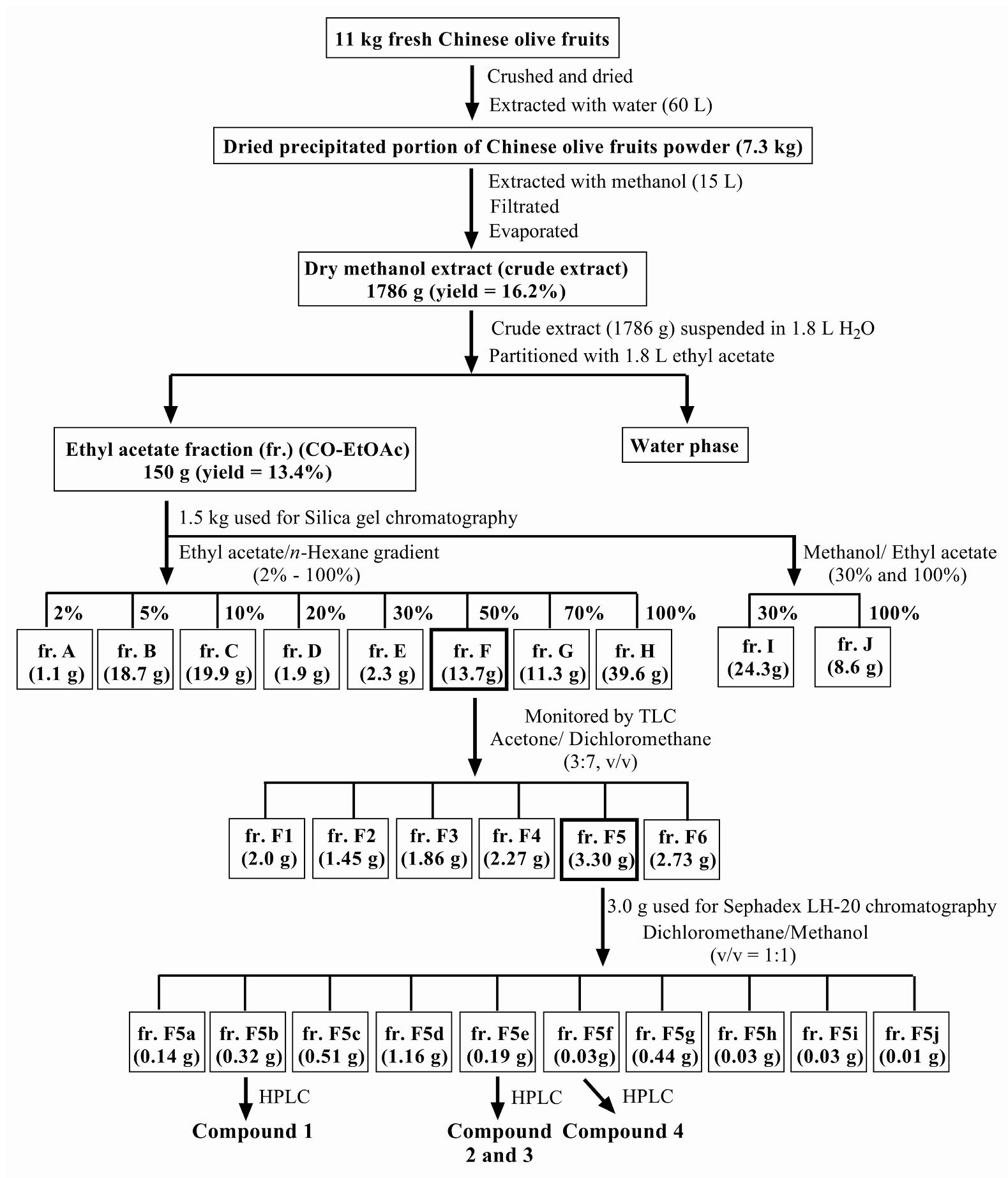
2.3. Plant Material and Extraction Procedures
2.4. Cell Culture
2.5. Water-Soluble Tetrazolium Salt (WST)-1 Cell Proliferation Assay
2.6. Nitric Oxide (NO) Assay
2.7. Statistical Analysis
3. Results
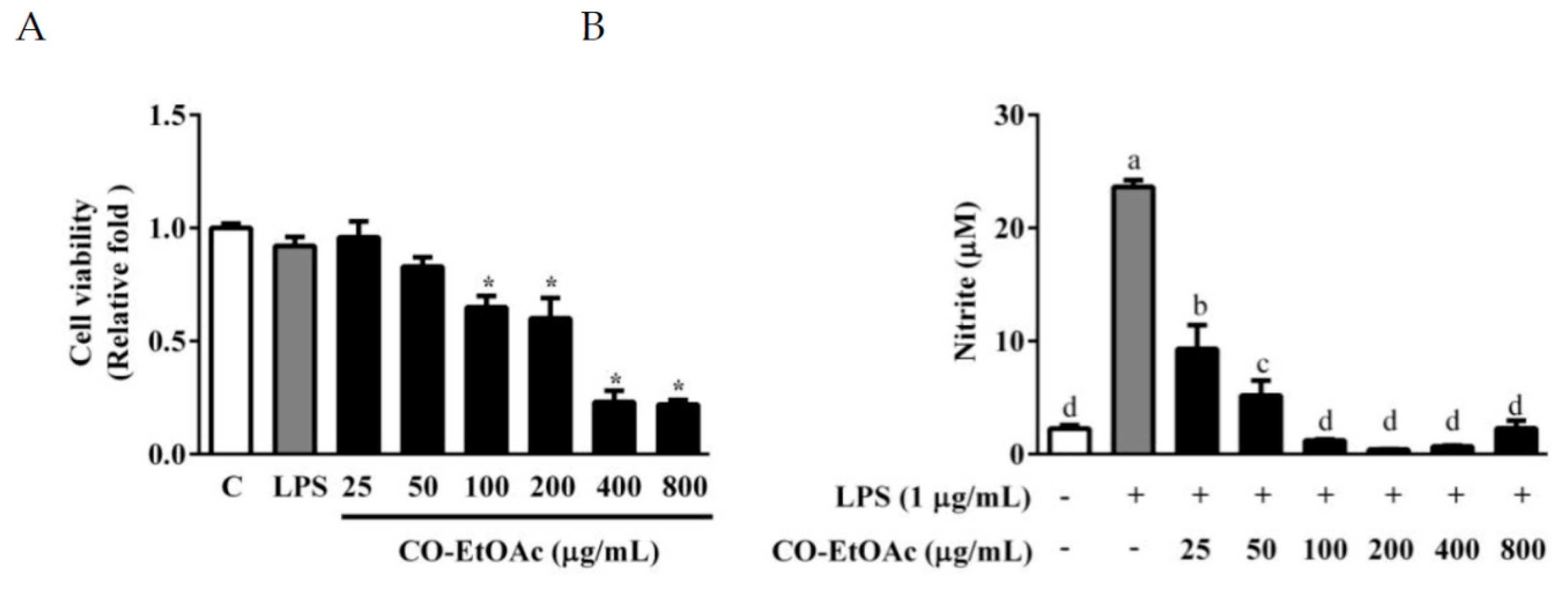
3.1. The Effects of CO-EtOAc on Cell Viability and Lipopolysaccharide (LPS)-Induced NO Production in RAW264.7 Cells
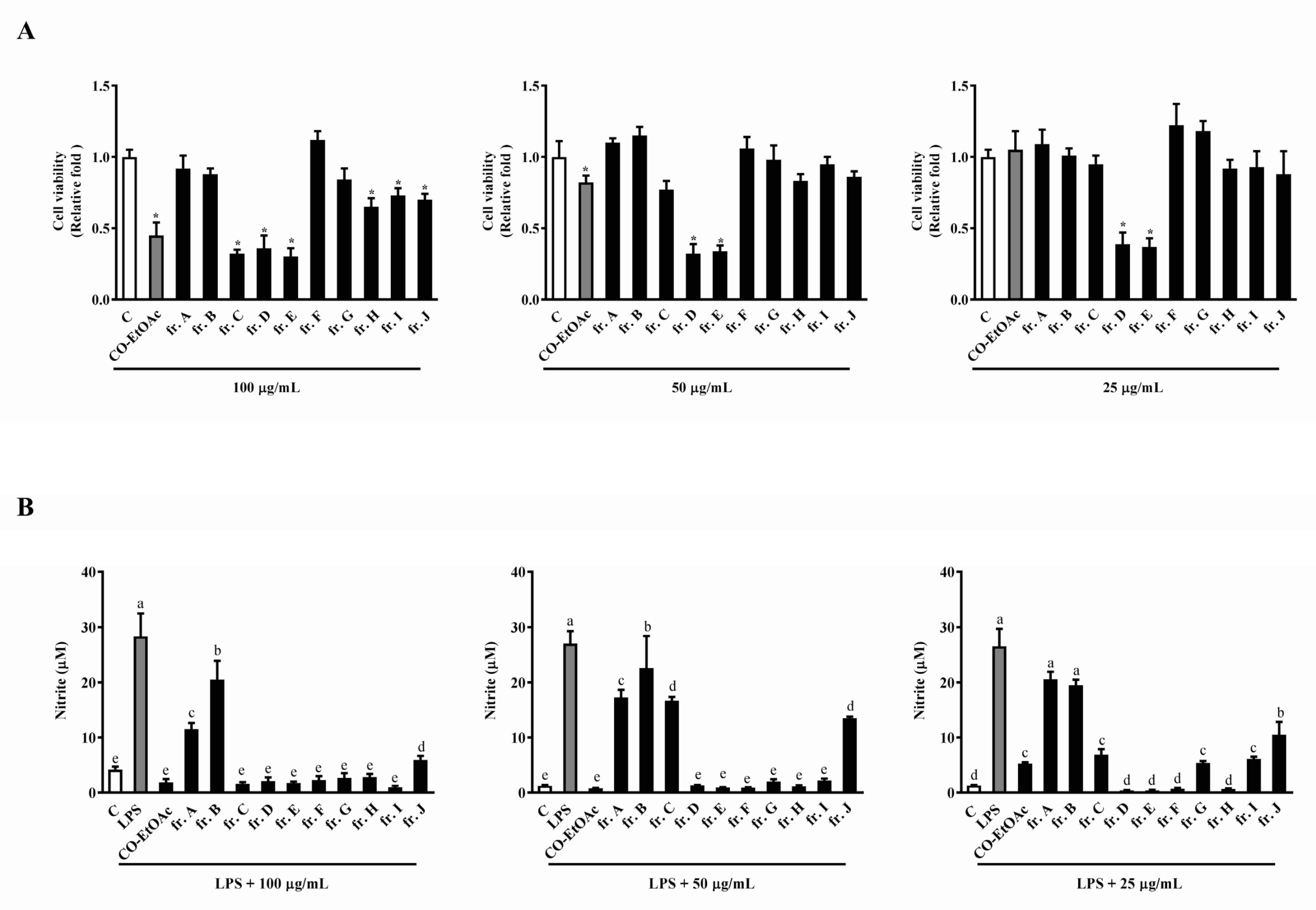
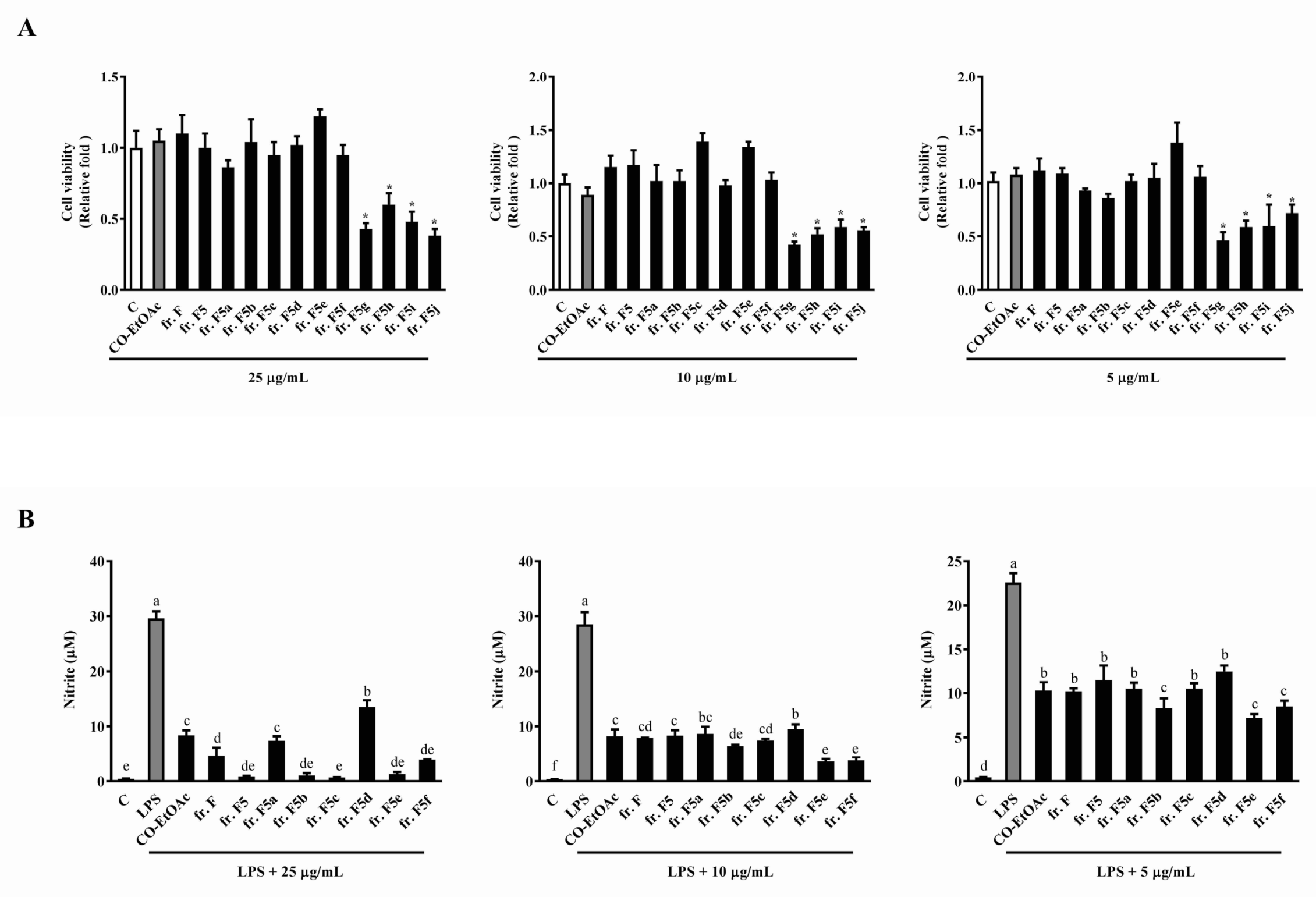
3.2. The Effects of the Subfractions from CO-EtOAc on Cell Viability and LPS-Induced NO Production in RAW264.7 Cells
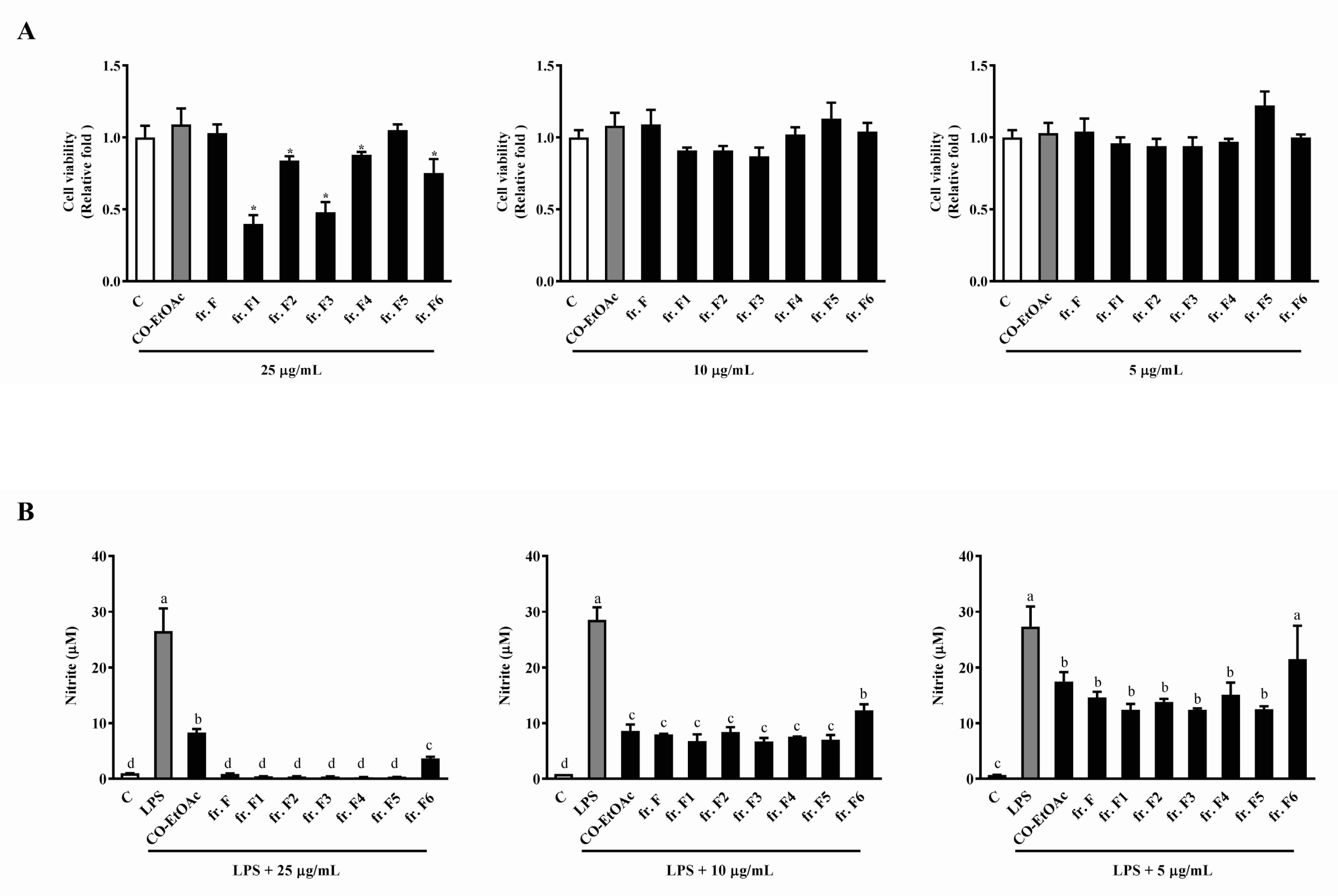
3.3. The Effects of the Fraction F Subfraction on Cell Viability and LPS-Induced NO Production in RAW264.7 Cells
3.4. The Effects of the Subfractions from Fraction F5 on Cell Viability and LPS-Induced NO Production in RAW264.7 Cells
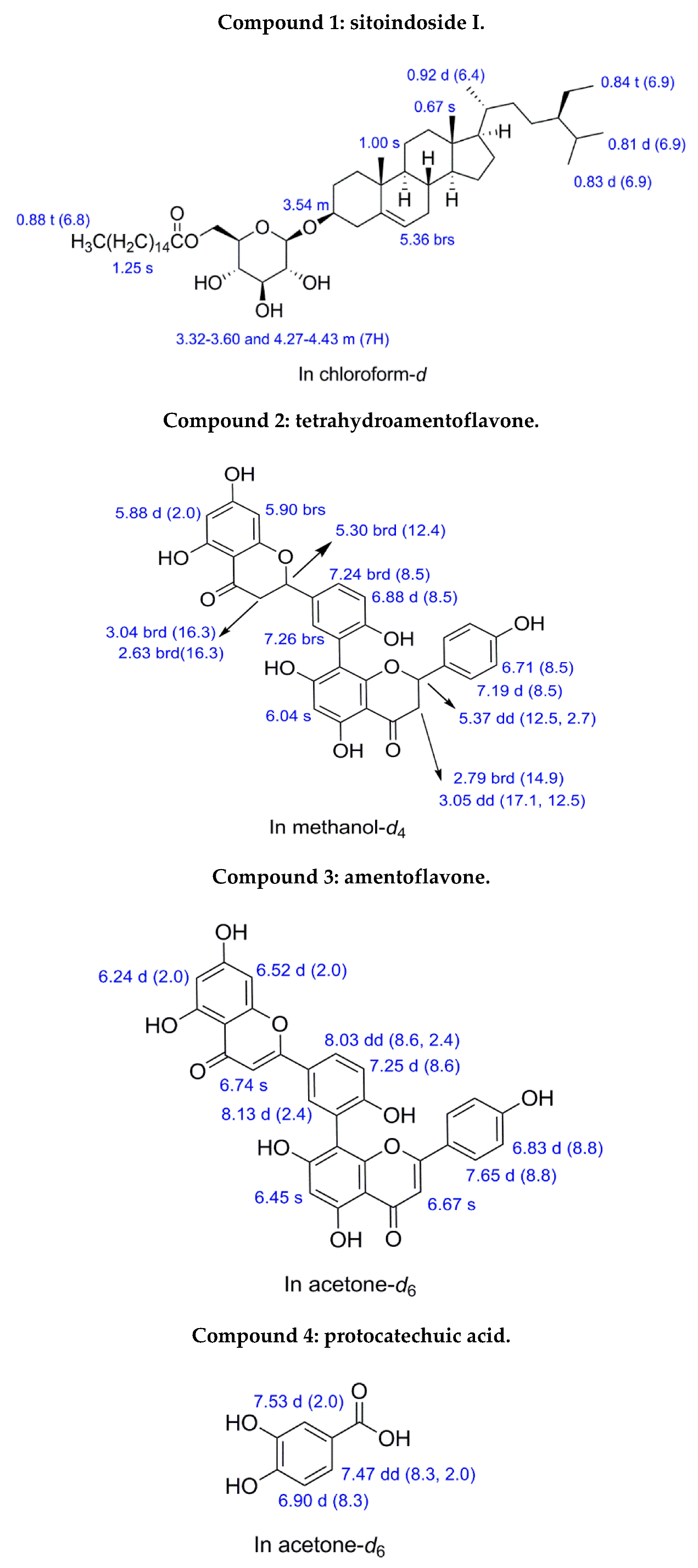
3.5. Identifying the Chromatographic Peaks in frs. F5b, F5e, and F5f
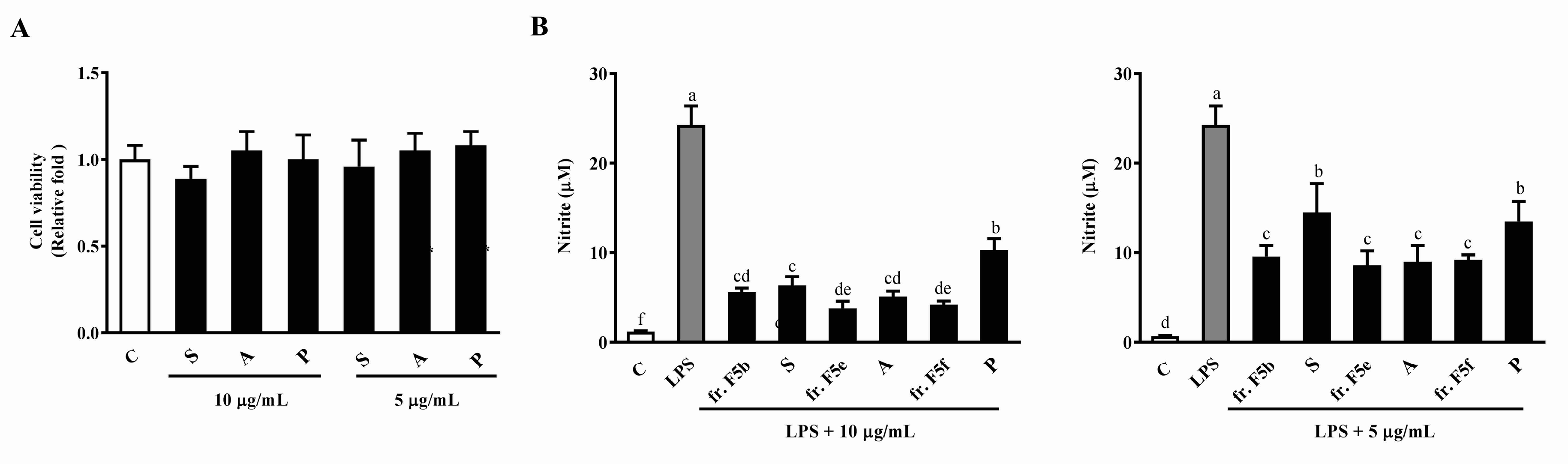
3.6. The Effects of Sitoindoside I, Amentoflavone, and Protocatechuic Acid on Cell Viability and LPS-Induced NO Production in RAW264.7 Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Kotas, M.E.; Medzhitov, R. Homeostasis, inflammation, and disease susceptibility. Cell 2015, 160, 816–827. [Google Scholar] [CrossRef] [PubMed]
- Nicholas, C.; Batra, S.; Vargo, M.A.; Voss, O.H.; Gavrilin, M.A.; Wewers, M.D.; Guttridge, D.C.; Grotewold, E.; Doseff, A.I. Apigenin blocks lipopolysaccharide-induced lethality in vivo and proinflammatory cytokines expression by inactivating nf-kappab through the suppression of p65 phosphorylation. J. Immunol. 2007, 179, 7121–7127. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.P.; Abraham, A. Inhibition of LPS induced pro-inflammatory responses in RAW 264.7 macrophage cells by PVP-coated naringenin nanoparticle via down regulation of nf-kappab/p38mapk mediated stress signaling. Pharmacol. Rep. 2017, 69, 908–915. [Google Scholar] [CrossRef]
- Wang, J.; Mazza, G. Effects of anthocyanins and other phenolic compounds on the production of tumor necrosis factor alpha in LPS/IFN-gamma-activated RAW 264.7 macrophages. J. Agric. Food Chem. 2002, 50, 4183–4189. [Google Scholar] [CrossRef] [PubMed]
- Gabay, C. Interleukin-6 and chronic inflammation. Arthritis Res. Ther. 2006, 8, S3. [Google Scholar] [CrossRef] [PubMed]
- Wang, A.; Al-Kuhlani, M.; Johnston, S.C.; Ojcius, D.M.; Chou, J.; Dean, D. Transcription factor complex ap-1 mediates inflammation initiated by chlamydia pneumoniae infection. Cell. Microbiol. 2013, 15, 779–794. [Google Scholar] [CrossRef]
- Nathan, C.; Shiloh, M.U. Reactive oxygen and nitrogen intermediates in the relationship between mammalian hosts and microbial pathogens. Proc. Natl. Acad. Sci. USA 2000, 97, 8841–8848. [Google Scholar] [CrossRef] [Green Version]
- Zirk, N.M.; Hashmi, S.F.; Ziegler, H.K. The polysaccharide portion of lipopolysaccharide regulates antigen-specific t-cell activation via effects on macrophage-mediated antigen processing. Infect. Immun. 1999, 67, 319–326. [Google Scholar]
- Fujiwara, N.; Kobayashi, K. Macrophages in inflammation. Curr. Drug Targets Inflamm. Allergy 2005, 4, 281–286. [Google Scholar] [CrossRef]
- Chung, H.T.; Pae, H.O.; Choi, B.M.; Billiar, T.R.; Kim, Y.M. Nitric oxide as a bioregulator of apoptosis. Biochem. Biophys. Res. Commun. 2001, 282, 1075–1079. [Google Scholar] [CrossRef]
- Taira, J.; Nanbu, H.; Ueda, K. Nitric oxide-scavenging compounds in agrimonia pilosa ledeb on lps-induced raw264. 7 macrophages. Food Chem. 2009, 115, 1221–1227. [Google Scholar] [CrossRef]
- Pacher, P.; Beckman, J.S.; Liaudet, L. Nitric oxide and peroxynitrite in health and disease. Physiol. Rev. 2007, 87, 315–424. [Google Scholar] [CrossRef] [PubMed]
- Kropf, P.; Kadolsky, U.D.; Rogers, M.; Cloke, T.E.; Müller, I. 13-the leishmaniasis model. Methods Microbiol. 2010, 37, 307–328. [Google Scholar]
- Maksimovic, Z.; Malencic, D.; Kovacevic, N. Polyphenol contents and antioxidant activity of maydis stigma extracts. Bioresour. Technol. 2005, 96, 873–877. [Google Scholar] [CrossRef] [PubMed]
- Mohsen, S.M.; Ammar, A.S. Total phenolic contents and antioxidant activity of corn tassel extracts. Food Chem. 2009, 112, 595–598. [Google Scholar] [CrossRef]
- Ding, B. Pharmacology of qingguo pills on relieving cough. China Tradit. Pat. Med. 1999, 21, 27–28. [Google Scholar]
- Kuo, C.T.; Liu, T.H.; Hsu, T.H.; Lin, F.Y.; Chen, H.Y. Antioxidant and antiglycation properties of different solvent extracts from chinese olive (Canarium album L.) fruit. Asian Pac. J. Trop. Med. 2015, 8, 1013–1021. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, S.C.; Hsieh, W.J.; Chiang, A.N.; Su, N.W.; Yeh, Y.T.; Liao, Y.C. The methanol-ethyl acetate partitioned fraction from Chinese olive fruits inhibits cancer cell proliferation and tumor growth by promoting apoptosis through the suppression of the nf-kappab signaling pathway. Food Funct. 2016, 7, 4797–4803. [Google Scholar] [CrossRef]
- Liu, Q.; Zhou, M.; Zheng, M.; Chen, N.; Zheng, X.; Zeng, S.; Zheng, B. Canarium album extract restrains lipid excessive accumulation in hepatocarcinoma cells. Int. J. Clin. Exp. Med. 2016, 9, 17509–17518. [Google Scholar]
- Yeh, Y.T.; Chiang, A.N.; Hsieh, S.C. Chinese olive (Canarium album L.) fruit extract attenuates metabolic dysfunction in diabetic rats. Nutrients 2017, 9, 1123. [Google Scholar] [CrossRef]
- Wang, C.H.; Lin, J.H.; Lu, T.J.; Chiang, A.N.; Chiou, S.T.; Chen, Y.A.; Pan, M.H.; Hsieh, S.C. Establishment of reporter platforms capable of detecting nf-κb mediated immuno-modulatory activity. J. Agric. Food Chem. 2013, 61, 12582–12587. [Google Scholar] [CrossRef] [PubMed]
- Pandey, K.B.; Rizvi, S.I. Plant polyphenols as dietary antioxidants in human health and disease. Oxid. Med. Cell. Longev. 2009, 2, 270–278. [Google Scholar] [CrossRef] [PubMed]
- Ogbe, R.J.; Ochalefu, D.O.; Mafulul, S.G.; Olaniru, O.B. A review on dietary phytosterols: Their occurrence, metabolism and health benefits. Asian J. Plant Sci. Res. 2015, 5, 10–21. [Google Scholar]
- Jessup, W.; Herman, A.; Chapman, M.J. Phytosterols in cardiovascular disease: Innocuous dietary components, or accelerators of atherosclerosis? Future Lipidol. 2008, 3, 301–310. [Google Scholar] [CrossRef]
- Weihrauch, J.L.; Gardner, J.M. Sterol content of foods of plant origin. J. Am. Diet. Assoc. 1978, 73, 39–47. [Google Scholar] [PubMed]
- Rivers, E.; Nguyen, B.; Havstad, S.; Ressler, J.; Muzzin, A.; Knoblich, B.; Peterson, E.; Tomlanovich, M.; Early Goal-Directed Therapy Collaborative Group. Early goal-directed therapy in the treatment of severe sepsis and septic shock. N. Engl. J. Med. 2001, 345, 1368–1377. [Google Scholar] [CrossRef] [PubMed]
- Saeidnia, S.; Manayi, A.; Gohari, A.R.; Abdollahi, M. The story of beta-sitosterol-a review. Eur. J. Med. Plants 2014, 4, 590. [Google Scholar] [CrossRef]
- Heitzman, M.E.; Neto, C.C.; Winiarz, E.; Vaisberg, A.J.; Hammond, G.B. Ethnobotany, phytochemistry and pharmacology of uncaria (rubiaceae). Phytochemistry 2005, 66, 5–29. [Google Scholar] [CrossRef]
- Bouic, P.; Etsebeth, S.; Liebenberg, R.; Albrecht, C.; Pegel, K.; Van Jaarsveld, P. Beta-sitosterol and beta-sitosterol glucoside stimulate human peripheral blood lymphocyte proliferation: Implications for their use as an immunomodulatory vitamin combination. Int. J. Immunopharmacol. 1996, 18, 693–700. [Google Scholar] [CrossRef]
- Hernandez-Valle, E.; Herrera-Ruiz, M.; Salgado, G.R.; Zamilpa, A.; Ocampo, M.L.; Aparicio, A.J.; Tortoriello, J.; Jimenez-Ferrer, E. Anti-inflammatory effect of 3-o-[(6’-o-palmitoyl)-beta-d-glucopyranosyl sitosterol] from agave angustifolia on ear edema in mice. Molecules 2014, 19, 15624–15637. [Google Scholar] [CrossRef]
- Lee, J.Y.; Jung, K.W.; Woo, E.R.; Kim, Y.M. Docking study of biflavonoids, allosteric inhibitors of protein tyrosine phosphatase 1b. Bull. Korean Chem. Soc. 2008, 29, 1479–1484. [Google Scholar]
- Chen, J.; Chang, H.W.; Kim, H.P.; Park, H. Synthesis of phospholipase a2 inhibitory biflavonoids. Bioorg. Med. Chem. Lett. 2006, 16, 2373–2375. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.; Yan, H.; Zhang, L.; Shan, M.; Chen, P.; Ding, A.; Li, S.F. A review on the phytochemistry, pharmacology, and pharmacokinetics of amentoflavone, a naturally-occurring biflavonoid. Molecules 2017, 22, 299. [Google Scholar] [CrossRef] [PubMed]
- He, Z.; Xia, W.; Chen, J. Isolation and structure elucidation of phenolic compounds in chinese olive (Canarium album L.) fruit. Eur. Food Res. Technol. 2008, 226, 1191–1196. [Google Scholar] [CrossRef]
- Funakoshi-Tago, M.; Okamoto, K.; Izumi, R.; Tago, K.; Yanagisawa, K.; Narukawa, Y.; Kiuchi, F.; Kasahara, T.; Tamura, H. Anti-inflammatory activity of flavonoids in nepalese propolis is attributed to inhibition of the il-33 signaling pathway. Int. Immunopharmacol. 2015, 25, 189–198. [Google Scholar] [CrossRef] [PubMed]
- Abdallah, H.M.; Almowallad, F.M.; Esmat, A.; Shehata, I.A.; Abdel-Sattar, E.A. Anti-inflammatory activity of flavonoids from chrozophora tinctoria. Phytochem. Lett. 2015, 13, 74–80. [Google Scholar] [CrossRef]
- Arwa, P.S.; Zeraik, M.L.; Ximenes, V.F.; da Fonseca, L.M.; da Silva Bolzani, V.; Silva, D.H.S. Redox-active biflavonoids from garcinia brasiliensis as inhibitors of neutrophil oxidative burst and human erythrocyte membrane damage. J. Ethnopharmacol. 2015, 174, 410–418. [Google Scholar] [CrossRef] [PubMed]
- Woo, E.R.; Lee, J.Y.; Cho, I.J.; Kim, S.G.; Kang, K.W. Amentoflavone inhibits the induction of nitric oxide synthase by inhibiting nf-kappab activation in macrophages. Pharmacol. Res. 2005, 51, 539–546. [Google Scholar] [CrossRef]
- Arimboor, R.; Rangan, M.; Aravind, S.G.; Arumughan, C. Tetrahydroamentoflavone (tha) from semecarpus anacardium as a potent inhibitor of xanthine oxidase. J. Ethnopharmacol. 2011, 133, 1117–1120. [Google Scholar] [CrossRef]
- Nomura, J.; Busso, N.; Ives, A.; Tsujimoto, S.; Tamura, M.; So, A.; Yamanaka, Y. Febuxostat, an inhibitor of xanthine oxidase, suppresses lipopolysaccharide-induced mcp-1 production via mapk phosphatase-1-mediated inactivation of jnk. PLoS ONE 2013, 8, e75527. [Google Scholar] [CrossRef]
- Selvam, C.; Jachak, S.M. A cyclooxygenase (cox) inhibitory biflavonoid from the seeds of semecarpus anacardium. J. Ethnopharmacol. 2004, 95, 209–212. [Google Scholar] [CrossRef] [PubMed]
- Robbins, R.J. Phenolic acids in foods: An overview of analytical methodology. J. Agric. Food Chem. 2003, 51, 2866–2887. [Google Scholar] [CrossRef] [PubMed]
- Kakkar, S.; Bais, S. A review on protocatechuic acid and its pharmacological potential. ISRN Pharmacol. 2014, 2014, 952943. [Google Scholar] [CrossRef] [PubMed]
- Min, S.W.; Ryu, S.N.; Kim, D.H. Anti-inflammatory effects of black rice, cyanidin-3-o-beta-d-glycoside, and its metabolites, cyanidin and protocatechuic acid. Int. Immunopharmacol. 2010, 10, 959–966. [Google Scholar] [CrossRef] [PubMed]
- Bendary, E.; Francis, R.; Ali, H.; Sarwat, M.; El Hady, S. Antioxidant and structure–activity relationships (sars) of some phenolic and anilines compounds. Ann. Agric. Sci. 2013, 58, 173–181. [Google Scholar] [CrossRef]







© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kuo, Y.-H.; Yeh, Y.-T.; Pan, S.-Y.; Hsieh, S.-C. Identification and Structural Elucidation of Anti-Inflammatory Compounds from Chinese Olive (Canarium Album L.) Fruit Extracts. Foods 2019, 8, 441. https://0-doi-org.brum.beds.ac.uk/10.3390/foods8100441
Kuo Y-H, Yeh Y-T, Pan S-Y, Hsieh S-C. Identification and Structural Elucidation of Anti-Inflammatory Compounds from Chinese Olive (Canarium Album L.) Fruit Extracts. Foods. 2019; 8(10):441. https://0-doi-org.brum.beds.ac.uk/10.3390/foods8100441
Chicago/Turabian StyleKuo, Yueh-Hsiung, Yu-Te Yeh, Sih-Ying Pan, and Shu-Chen Hsieh. 2019. "Identification and Structural Elucidation of Anti-Inflammatory Compounds from Chinese Olive (Canarium Album L.) Fruit Extracts" Foods 8, no. 10: 441. https://0-doi-org.brum.beds.ac.uk/10.3390/foods8100441