Enterotoxin Genes, Antibiotic Susceptibility, and Biofilm Formation of Low-Temperature-Tolerant Bacillus cereus Isolated from Green Leaf Lettuce in the Cold Chain

Abstract
:1. Introduction
2. Materials and Methods
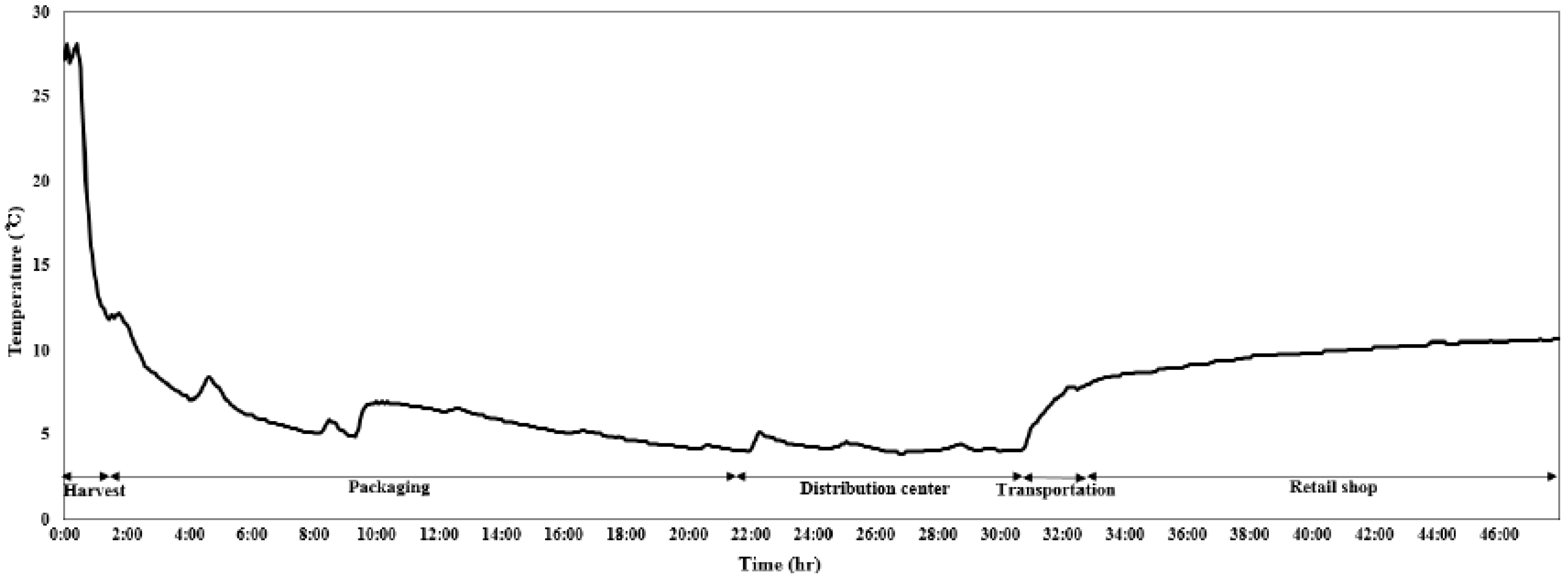
2.1. Collection of Samples
2.2. Microbiological Analysis for the B. cereus Group
2.3. Bacterial DNA Extraction
2.4. Identification of B. cereus and B. thuringiensis from the B. cereus Group
2.5. Growth Properties
2.6. Detection of Enterotoxin and Emetic Toxin Genes
2.7. Antibiotic Susceptibility Testing
2.8. Quantification of Biofilm Formation
2.9. Statistical Analysis
3. Results and Discussion
3.1. Prevalence and Identification of the B. cereus Group
3.2. Psychrotolerant Properties of B. cereus Isolated from Vegetables
3.3. Enterotoxigenic Potential of Psychrotolerant B. cereus Isolates
3.4. Antibiotic Resistance
3.5. Biofilm Formation
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Hertog, M.L.A.T.M.; Uysal, I.; McCarthy, U.; Verlinden, B.M.; Nicolai, B.M. Shelf life modelling for first-expired-first-out warehouse management. Philos. Trans. A Math. Phys. Eng. Sci. 2014, 372, 20130306. [Google Scholar] [CrossRef]
- Sorhaug, T.; Stepaniak, L. Psychrotrophs and their enzymes in milk and dairy products: Quality aspects. Trends Food Sci. Technol. 1997, 8, 35–41. [Google Scholar] [CrossRef]
- Wouters, J.A.; Rombouts, F.M.; Kuipers, O.P.; de Vos, W.M.; Abee, T. The role of cold-shock proteins in low-temperature adaptation of food-related bacteria. Syst. Appl. Microbiol. 2000, 23, 165–173. [Google Scholar] [CrossRef] [Green Version]
- Forghani, F.; Kim, J.B.; Oh, D.H. Enterotoxigenic profiling of emetic toxin and enterotoxin-producing Bacillus cereus, isolated from food, environmental, and clinical samples by Multiplex PCR. J. Food Sci. 2014, 79, 2288–2293. [Google Scholar] [CrossRef] [PubMed]
- Jo, M.J.; Jeong, A.R.; Kim, H.J.; Lee, N.R.; Oh, S.W.; Kim, Y.J.; Chun, H.S.; Koo, M.S. Microbiological quality of fresh-cut produce and organic vegetables. Korean J. Food Sci. Technol. 2011, 43, 91–97. [Google Scholar] [CrossRef] [Green Version]
- Agata, N.; Ohta, M.; Mori, M.; Isobe, M. A novel dodecadepsipeptide, cereulide, is an emetic toxin of Bacillus cereus. FEMS Microbiol. Lett. 1995, 129, 17–20. [Google Scholar]
- Kramer, J.M.; Gilbert, R.J. Bacillus cereus and other Bacillus species. In Foodborne Bacterial Pathogens; Marcel Dekker Inc.: New York, NY, USA, 1989. [Google Scholar]
- Granum, P.E. Bacillus cereus and its toxins. Soc. Appl. Bacteriol. Symp. Ser. 1994, 23, 61–66. [Google Scholar]
- Anderson, A.; Ronener, U.; Granum, P.E. What problems does the food industry have with the spore-forming pathogens Bacillus cereus and Clostridium perfringens? Int. J. Food Microbiol. 1995, 28, 145–155. [Google Scholar] [CrossRef]
- Lindsay, D.; Brözel, V.S.; Von Holy, A. Biofilm-spore response in Bacillus cereus and Bacillus subtilis during nutrient limitation. J. Food Prot. 2006, 69, 1168–1172. [Google Scholar] [CrossRef]
- Samie, N.; Noghabi, K.; Gharegozloo, Z.; Zahiri, H.; Ahmadian, G.; Sharafi, H. Psychrophilic α-amylase from Aeromonas veronii NS07 isolated from farm soils. Process. Biochem. 2012, 47, 1381–1387. [Google Scholar] [CrossRef]
- Larsen, H.D.; Jørgensen, K. The occurrence of Bacillus cereus in Danish pasteurized milk. Int. J. Food Microbiol. 1997, 34, 179–186. [Google Scholar] [CrossRef]
- Meer, R.R.; Baker, J.; Bodyfelt, F.W.; Griffiths, M.W. Psychrotrophic Bacillus spp. in fluid milk products: A review. J. Food Prot. 1991, 54, 969–979. [Google Scholar] [CrossRef] [PubMed]
- Huck, J.R.; Hammond, B.H.; Murphy, S.C.; Woodcock, N.H.; Boor, K.J. Tracking spore-forming bacterial contaminants in fluid milk-processing systems. J. Dairy Sci. 2007, 90, 4872–4883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baron, F.; Cochet, M.F.; Grosset, N.; Madec, M.N.; Briandet, R.; Dessaigne, S.; Chevalier, S.; Gautier, M.; Jan, S. Isolation and characterization of a psychrotolerant toxin producer, Bacillus weihenstephanensis in liquid egg products. J. Food Prot. 2007, 70, 2782–2791. [Google Scholar] [CrossRef] [PubMed]
- Stenfors Arnesen, L.; Granum, P.E.; Buisson, C.; Bohlin, J.; Nielsen-LeRoux, C. Using an insect model to assess correlation between temperature and virulence in Bacillus weihenstephanensis and Bacillus cereus. FEMS Microbiol. Lett. 2011, 317, 196–202. [Google Scholar] [CrossRef] [Green Version]
- Guerin, A.; Dargaignaratz, C.; Broussolle, V.; Clavel, T.; Nguyen-the, C. Combined effect of anaerobiosis, low pH and cold temperatures on the growth capacities of psychrotrophic Bacillus cereus. Food Microbiol. 2016, 59, 119–123. [Google Scholar] [CrossRef]
- Choma, C.; Guinebretiere, M.H.; Carlin, F.; Schmitt, P.; Velge, P.; Granum, P.E.; Nguyen-The, C. Prevalence, characterization and growth of Bacillus cereus in commercial cooked chilled foods containing vegetables. J. Appl. Microbiol. 2000, 88, 617–625. [Google Scholar] [CrossRef]
- Dancer, S.J. Mopping up hospital infection. J. Hosp. Infect. 1999, 43, 85–100. [Google Scholar] [CrossRef] [Green Version]
- Berthold-Pluta, A.; Garbowska, M.; Stefanska, I.; Pluta, A. Microbiological quality of selected ready-to-eat leaf vegetables, sprouts and non-pasteurized fresh fruit-vegetable juices including the presence of Cronobacter spp. Food Microbiol. 2017, 65, 221–230. [Google Scholar] [CrossRef]
- Park, K.M.; Jeong, M.; Park, K.J.; Koo, M. Prevalence, enterotoxin genes, and antibiotic resistance of Bacillus cereus isolated from raw vegetables in Korea. J. Food Prot. 2018, 81, 1590–1597. [Google Scholar] [CrossRef]
- Ozcelik, B.; Citak, S. Evaluation of antibiotic resistance of Bacillus cereus isolates in ice cream samples sold in Ancara. Turk. J. Pharm. Sci. 2009, 6, 231–238. [Google Scholar]
- Al-Khatib, M.S.; Khyami-Horani, H.; Badran, E.; Shehabi, A. Incidence and characterization of diarrheal enterotoxins of fecal Bacillus cereus isolates associated with diarrhea. Diagn. Microbiol. Infect. Dis. 2007, 59, 383–387. [Google Scholar] [CrossRef] [PubMed]
- van Netten, P.; van De Moosdijk, A.; van Hoensel, P.; Mossel, D.A.; Perales, I. Psychrotrophic strains of Bacillus cereus producing enterotoxin. J. Appl. Bacteriol. 1990, 69, 73–79. [Google Scholar] [CrossRef] [PubMed]
- Korea Food and Drug Administration (KFDA). Food Code. Available online: http://www.kfda.go.kr (accessed on 1 October 2019).
- Yamada, S.; Ohashi, E.; Agata, N.; Venkateswaran, K. Cloning and nucleotide sequence analysis of gyrB of Bacillus cereus, B. thuringiensis, B. mycoides, and B. anthracis and their application to the detection of B. cereus in rice. Appl. Environ. Microbiol. 1999, 65, 1483–1490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Święcicka, I.; Mahillon, J. Diversity of Commensal Bacillus cereus sensu lato isolated from the common sow bug (Porcellio scaber, Isopoda). FEMS Microbiol. Ecol. 2006, 56, 132–140. [Google Scholar] [CrossRef]
- Clinical and Laboratory Standards Institute (CLSI). Performance Standards for Antimicrobial Susceptibility Testing; Clinical and Laboratory Standards Institute: Tempe, AZ, USA, 2014. [Google Scholar]
- Singh, A.K.; Prakash, P.; Achra, A.; Singh, G.P.; Das, A.; Singh, R.K. Standardization and classification of in vitro biofilm formation by clinical isolates of Staphylococcus aureus. J. Glob. Infect. Dis. 2017, 9, 93–101. [Google Scholar]
- Chon, J.W.; Yim, J.H.; Kim, H.S.; Kim, D.H.; Kim, H.; Oh, D.H. Quantitative prevalence and toxin gene profile of Bacillus cereus from ready-to-eat vegetables in South Korea. Foodborne Pathog. Dis. 2015, 12, 795–799. [Google Scholar] [CrossRef]
- Kim, H.J.; Koo, M.; Hwang, D.; Choi, J.H.; Kim, S.M.; Oh, S.W. Contamination patterns and molecular typing of Bacillus cereus in fresh-cut vegetable salad processing. Appl. Biol. Chem. 2016, 59, 573–577. [Google Scholar] [CrossRef]
- Valero, M.; Hernandez-Herrero, L.A.; Fernandez, P.S.; Salmeron, M.C. Characterization of Bacillus cereus isolates from fresh vegetables and refrigerated minimally processed foods by biochemical and physiological tests. Food Microbiol. 2002, 19, 491–499. [Google Scholar] [CrossRef]
- European Food Safety Authority (EFSA). Risks for public health related to the presence of Bacillus cereus and other Bacillus spp. including Bacillus thuringiensis in foodstuffs. EFSA J. 2016, 14, 4524–4593. [Google Scholar]
- Walker, S.J.; Archer, P.; Banks, J.G. Growth of Listeria monocytogenes at refrigeration temperatures. J. Appl. Bacteriol. 1990, 68, 157–162. [Google Scholar] [CrossRef] [PubMed]
- Tasara, T.; Stephan, R. Cold stress tolerance of Listeria monocytogenes: A review of molecular adaptive mechanisms and food safety implications. J. Food Prot. 2006, 69, 1473–1484. [Google Scholar] [CrossRef] [PubMed]
- Samapundo, S.; Heyndrickx, M.; Xhaferi, R.; Devlieghere, F. Incidence, diversity and toxin gene characteristics of Bacillus cereus group strains isolated from food products marketed in Belgium. Int. J. Food Microbiol. 2011, 150, 34–41. [Google Scholar] [CrossRef] [PubMed]
- Bartoszewicz, M.; Bideshi, D.K.; Kraszewska, A.; Modzelewska, E.; Swiecicka, I. Natural isolates of Bacillus thuringiensis display genetic and psychrotrophic properties characteristic of Bacillus weihenstephanensis. J. Appl. Microbiol. 2009, 106, 1967–1975. [Google Scholar] [CrossRef]
- Guoping, Z.; Dasheng, Z.; Lina, D.; Quanxin, C.; Zhiming, Y. Occurrence of psychrotrophic Bacillus cereus group strains in ice creams. Int. J. Food Microbiol. 2010, 137, 143–146. [Google Scholar]
- Kovac, J.; Miller, R.A.; Carroll, L.M.; Kent, D.J.; Jian, J.; Beno, S.M. Production of hemolysin BL by Bacillus cereus group isolates of dairy origin is associated with whole-genome phylogenetic clade. BMC Genom. 2016, 17, 581–596. [Google Scholar] [CrossRef] [Green Version]
- Miller, R.A.; Beno, S.M.; Kent, D.J.; Carroll, L.M.; Martin, N.H.; Boor, K.J. Bacillus wiedmannii sp. nov. is a new psychrotrophic and cytotoxic Bacillus cereus group species isolated from dairy foods and environments in the USA. Int. J. Syst. Evol. Microbiol. 2016, 66, 4744–4753. [Google Scholar] [CrossRef]
- Miller, R.A.; Jian, J.; Beno, S.M.; Wiedmann, M.; Kovac, J. Genomic and phenotypic characterization of type strains and dairy-associated isolates in the Bacillus cereus group indicates considerable intra-clade variability in toxin production and cytotoxicity. Appl. Environ. Microbiol. 2018, 84, 2479–2517. [Google Scholar] [CrossRef] [Green Version]
- Ngamwongsatit, P.; Buasri, W.; Pianariyanon, P.; Pulsrikan, C.; Ohba, M. Broad distribution of enterotoxin genes (hblCDA, nhe ABC, cytK, and entFM) among Bacillus thuringiensis and Bacillus cereus as shown by novel primers. Int. J. Food Microbiol. 2008, 121, 352–356. [Google Scholar] [CrossRef]
- Ryan, P.A.; MacMillan, J.D.; Zilinskas, B.A. Molecular cloning and characterization of the genes encoding L1 and L2 components of hemolysin BL from Bacillus cereus. J. Bacteriol. 1997, 179, 2551–2556. [Google Scholar] [CrossRef] [Green Version]
- Merzougui, S.; Cohen, N.; Grosset, N.; Gautier, M.; Lkhider, M. Enterotoxigenic Profiles of psychrotrophic and mesophilic strains of the Bacillus cereus group isolated from food in Morocco. Int. J. Eng. Res. Appl. 2013, 3, 964–970. [Google Scholar]
- Oladipo, I.C.; Adejumobi, O.D. Incidence of antibiotic resistance in some bacterial pathogens from street vended food in Ogbomoso, Nigeria. Pak. J. Nutr. 2010, 9, 1061–1068. [Google Scholar] [CrossRef] [Green Version]
- Goldstein, F. The potential clinical impact of low level antibiotic resistance in Staphylococcus aureus. J. Antimicrob. Chemother. 2007, 59, 1–4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campbell, E.; Korzheva, N.; Mustaev, A.; Murakami, K.; Nair, S.; Goldfarb, A.; Darst, S.A. Structural mechanism for rifampicin inhibition of bacterial RNA polymerase. Cell 2001, 104, 901–912. [Google Scholar] [CrossRef]
- Park, Y.B.; Kim, J.B.; Shin, S.W.; Kim, J.C.; Cho, S.H.; Lee, B.K.; Ahn, J.; Kim, J.M.; Oh, D.H. Prevalence, genetic diversity, and antibiotic susceptibility of Bacillus cereus strains isolated from rice and cereals collected in Korea. J. Food Prot. 2009, 72, 612–617. [Google Scholar] [CrossRef] [PubMed]
- Agwa, O.K.; Uzoigwe, C.I.; Wokoma, E.C. Incidence and antibiotic sensitivity of Bacillus cereus isolated from ready to eat foods sold in some markets in portharcourt, rivers state, Nigeria. Asian J. Microbiol. Biotechnol. Environ. Sci. 2012, 14, 13–18. [Google Scholar]
- Owusu-Kwarteng, J.; Wuni, A.; Akabanda, F.; Tano-Debrah, K.; Jespersen, L. Prevalence, virulence factor genes and antibiotic resistance of Bacillus cereus sensu lato isolated from dairy farms and traditional dairy products. BMC Microbiol. 2017, 17, 65–72. [Google Scholar] [CrossRef] [Green Version]
- Chang, Q.; Wang, W.; Regev-Yochay, G.; Lipsitch, M.; Hanage, W.P. Antibiotics in agriculture and the risk to human health: How worried should we be? Evol. Appl. 2015, 8, 240–245. [Google Scholar] [CrossRef] [Green Version]
- Byington, C.L.; Enriquez, F.R.; Hoff, C.; Tuohy, R.; Taggart, E.W.; Hillyard, D.R.; Carroll, K.C.; Christenson, J.C. Serious bacterial infections in febrile infants 1 to 90 days old with and without viral infections. Pediatrics 2004, 113, 1662–1666. [Google Scholar] [CrossRef]
- Storz, G.; Hengge-Aronis, R. Bacterial Stress Responses; ASM Press: Washington, DC, USA, 2000. [Google Scholar]
- Poole, K. Bacterial stress responses as determinants of antimicrobial resistance. J. Antimicrob. Chemother. 2012, 67, 2069–2089. [Google Scholar] [CrossRef] [Green Version]
- Mc Mahon, M.A.S.; Blair, I.S.; Moore, J.E.; Mc Dowell, D.A. The rate of horizontal transmission of antibiotic resistance plasmids is increased in food preservation-stressed bacteria. J. Appl. Microbiol. 2007, 103, 1883–1888. [Google Scholar] [CrossRef]
- Peng, J.S.; Tsai, W.C.; Chou, C.C. Inactivation and removal of Bacillus cereus by sanitizer and detergent. Int. J. Food Microbiol. 2002, 77, 11–18. [Google Scholar] [CrossRef]
- Sarita, K.; Sarkar, P.K. In vitro model study for biofilm formation by Bacillus cereus in dairy chilling tanks and optimization of clean-in-place (CIP) regimes using response surface methodology. Food Control. 2014, 36, 153–158. [Google Scholar]
- Fernandes, M.D.S.; Fujimoto, G.; Schneid, I.; Kabuki, D.Y.; Kuaye, A.Y. Enterotoxigenic profile, antimicrobial susceptibility, and biofilm formation of Bacillus cereus isolated from ricotta processing. Int. Dairy J. 2014, 38, 16–23. [Google Scholar] [CrossRef]
- Karatan, E.; Watnick, P. Signals, regulatory networks, and materials that build and break bacterial biofilms. Microbiol. Mol. Biol. Rev. 2009, 73, 310–347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simoes, L.C.; Simoes, M.; Vieira, M.J. Adhesion and biofilm formation on polystyrene by drinking water-isolated bacteria. Antonie Leeuwenhoek Int. J. Gen. 2010, 98, 317–329. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Targent Gene | Primer | Sequence (5’–3’) | Melting Temp (°C) | Amplicon (bp) |
|---|---|---|---|---|
| hblA | hblA-F hblA-R | GTG CAG ATG TTG ATG CCG AT ATG CCA CTG CGT GGA CAT AT | 55 | 319 |
| hblC | hblC-F hblC-R | AAT GGT CAT CGG AAC TCT AT CTC GCT GTT CTG CTG TTA AT | 55 | 749 |
| hblD | hblD-F hblD-R | AAT CAA GAG CTG TCA CGA AT CAC CAA TTG ACC ATG CTA AT | 55 | 429 |
| nheA | nheA-F nheA-R | TAC GCT AAG GAG GGG CA GTT TTT ATT GCT TCA TCG GCT | 55 | 499 |
| nheB | nheB-F nheB-R | CTA TCA GCA CTT ATG GCA G ACT CCT AGC CGG TGT TCC | 55 | 769 |
| nheC | nheC-F nheC-R | CGG TAG TGA TTG CTG GG CAG CAT TCG TAC TTG CCA A | 55 | 581 |
| entFM | entFM-F entFM-R | ATG AAA AAA GTA ATT TGC AGG TTA GTA TGC TTT TGT GTA ACC | 55 | 1269 |
| cytK | cytK-F cytK-R | GTA ACT TTC ATT GAT GAT CC GAA TAC TAA ATA ATT GGT TTC C | 44 | 505 |
| ces | ces-F ces-R | GGT GAC ACA TTA TCA TAT AAG GTG GTA AGC GAA CCT GTC TGT AAC AAC A | 55 | 1271 |
| Sampling Point | No. of B. cereus Group Isolates (%) | Contamination Level (Log CFU/g) | ||
|---|---|---|---|---|
| B. cereus | B. thuringiensis | B. mycoides | ||
| Harvest | 29 (100) | 0 (0) | 0 (0) | 1.9 ± 0.22 a,(1),(2) |
| Packaging | 17 (100) | 0 (0) | 0 (0) | 2.9 ± 0.44 b |
| Distribution center | 15 (100) | 0 (0) | 0 (0) | 2.6 ± 0.68 b |
| Transportation | 10 (100) | 0 (0) | 0 (0) | 2.1 ± 0.26 ab |
| Retail shop | 30 (100) | 0 (0) | 0 (0) | 2.6 ± 0.41 b |
| Total | 101 (100) | 0 (0) | 0 (0) | 2.4 ± 0.39 |
| Sampling Point | No. of B. cereus Isolates | Temperature of Growth (°C) | |||
|---|---|---|---|---|---|
| 12 °C | 10 °C | 7 °C | 5 °C | ||
| Harvest | 29 | 10 (34.5) | 8 (27.6) | 0 (0) | 0 (0) |
| Packaging | 17 | 12 (70.6) | 12 (70.6) | 1 (5.9) | 0 (0) |
| Distribution center | 15 | 14 (93.3) | 14 (93.3) | 6 (33.3) | 0 (0) |
| Transportation | 10 | 7 (70) | 5 (50.0) | 4 (20.0) | 0 (0) |
| Retail shop | 30 | 16 (53.3) | 12 (40.0) | 7 (23.3) | 0 (0) |
| Total | 101 | 59 (58.4) | 51 (50.5) | 18 (17.8) | 0 (0) |
| Enterotoxin Genes | No. (%) of Entertoxigenic B. cereus | ||
|---|---|---|---|
| Non-psychrotolerant B. cereus (n = 83) | Psychrotolerant B. cereus (n = 18) | ||
| Frequency of Enterotoxin Genes | |||
| 1 | nheABC | 81 (97.6) | 17 (94.4) |
| 2 | hblACD | 25 (30.1) | 8 (44.4) |
| 3 | cytK | 80 (96.4) | 10 (55.6) |
| 4 | entFM | 73 (88.0) | 14 (77.8) |
| 5 | ces | 0 (0.0) | 0 (0.0) |
| Profile of Enterotoxin Genes | |||
| 1 | nheABC + hblACD + cytK + entFM | 19 (23.2) | 5 (27.8) |
| 2 | nheABC + hblACD + entFM | 0 (0.0) | 3 (16.7) |
| 3 | nheABC + cytK + entFM | 52 (63.4) | 3 (16.7) |
| 4 | nheABC + hblACD + entFM | 2 (2.4) | 0 (0.0) |
| 5 | nheABC + hblACD + cytK | 3 (3.7) | 0 (0.0) |
| 6 | nheABC + cytK | 4 (4.9) | 1 (5.6) |
| 7 | nheABC + entFM | 0 (0.0) | 1 (5.6) |
| 8 | cytK + entFM | 2 (2.4) | 1 (5.6) |
| 9 | entFM | 0 (0.0) | 1 (5.6) |
| 10 | nheABC | 1 (1.2) | 3 (16.7) |
| No. (%) of B. cereus Isolates with Indicated Response | ||||||
|---|---|---|---|---|---|---|
| Non-psychrotolerant B. cereus (n = 83) | Psychrotolerant B. cereus (n = 18) | |||||
| Resistance | Intermediate | Sensitive | Resistance | Intermediate | Sensitive | |
| Tetracycline | 13 (15.7) | 50 (60.2) | 20 (24.1) | 2 (11.1) | 3 (16.7) | 13 (72.2) |
| Gentamicin | 0 (0.0) | 0 (0.0) | 83 (100.0) | 0 (0.0) | 0 (0.0) | 18 (100.0) |
| Imipenem | 0 (0.0) | 0 (0.0) | 83 (100.0) | 0 (0.0) | 0 (0.0) | 18 (100.0) |
| Vancomycin | 1 (1.2) | 0 (0.0) | 82 (98.8) | 0 (0.0) | 0 (0.0) | 18 (100.0) |
| Ciprofloxacin | 0 (0.0) | 0 (0.0) | 83 (100.0) | 0 (0.0) | 0 (0.0) | 18 (100.0) |
| Erythromycin | 0 (0.0) | 0 (0.0) | 83 (100.0) | 0 (0.0) | 0 (0.0) | 18 (100.0) |
| Rifampin | 65 (78.3) | 13 (15.7) | 5 (6.0) | 4 (22.2) | 6 (33.3) | 8 (44.4) |
| Chloraphenicol | 0 (0.0) | 0 (0.0) | 83 (100.0) | 0 (0.0) | 0 (0.0) | 18 (100.0) |
| Clindamycin | 0 (0.0) | 0 (0.0) | 83 (100.0) | 0 (0.0) | 8 (44.4) | 10 (55.6) |
| Penicillin | 83 (100.0) | 0 (0.0) | 0 (0.0) | 18 (100.0) | 0 (0.0) | 0 (0.0) |
| Ceftriaxone | 83 (100.0) | 0 (0.0) | 0 (0.0) | 18 (100.0) | 0 (0.0) | 0 (0.0) |
| Cefotaxim | 83 (100.0) | 0 (0.0) | 0 (0.0) | 18 (100.0) | 0 (0.0) | 0 (0.0) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, K.M.; Kim, H.J.; Jeong, M.; Koo, M. Enterotoxin Genes, Antibiotic Susceptibility, and Biofilm Formation of Low-Temperature-Tolerant Bacillus cereus Isolated from Green Leaf Lettuce in the Cold Chain. Foods 2020, 9, 249. https://0-doi-org.brum.beds.ac.uk/10.3390/foods9030249
Park KM, Kim HJ, Jeong M, Koo M. Enterotoxin Genes, Antibiotic Susceptibility, and Biofilm Formation of Low-Temperature-Tolerant Bacillus cereus Isolated from Green Leaf Lettuce in the Cold Chain. Foods. 2020; 9(3):249. https://0-doi-org.brum.beds.ac.uk/10.3390/foods9030249
Chicago/Turabian StylePark, Kyung Min, Hyun Jung Kim, Mooncheol Jeong, and Minseon Koo. 2020. "Enterotoxin Genes, Antibiotic Susceptibility, and Biofilm Formation of Low-Temperature-Tolerant Bacillus cereus Isolated from Green Leaf Lettuce in the Cold Chain" Foods 9, no. 3: 249. https://0-doi-org.brum.beds.ac.uk/10.3390/foods9030249